Multi-Temporal Predictive Modelling of Sorghum Biomass Using UAV-Based Hyperspectral and LiDAR Data


Abstract
1. Introduction
2. Related Work
3. Materials and Methods
3.1. Experimental Site
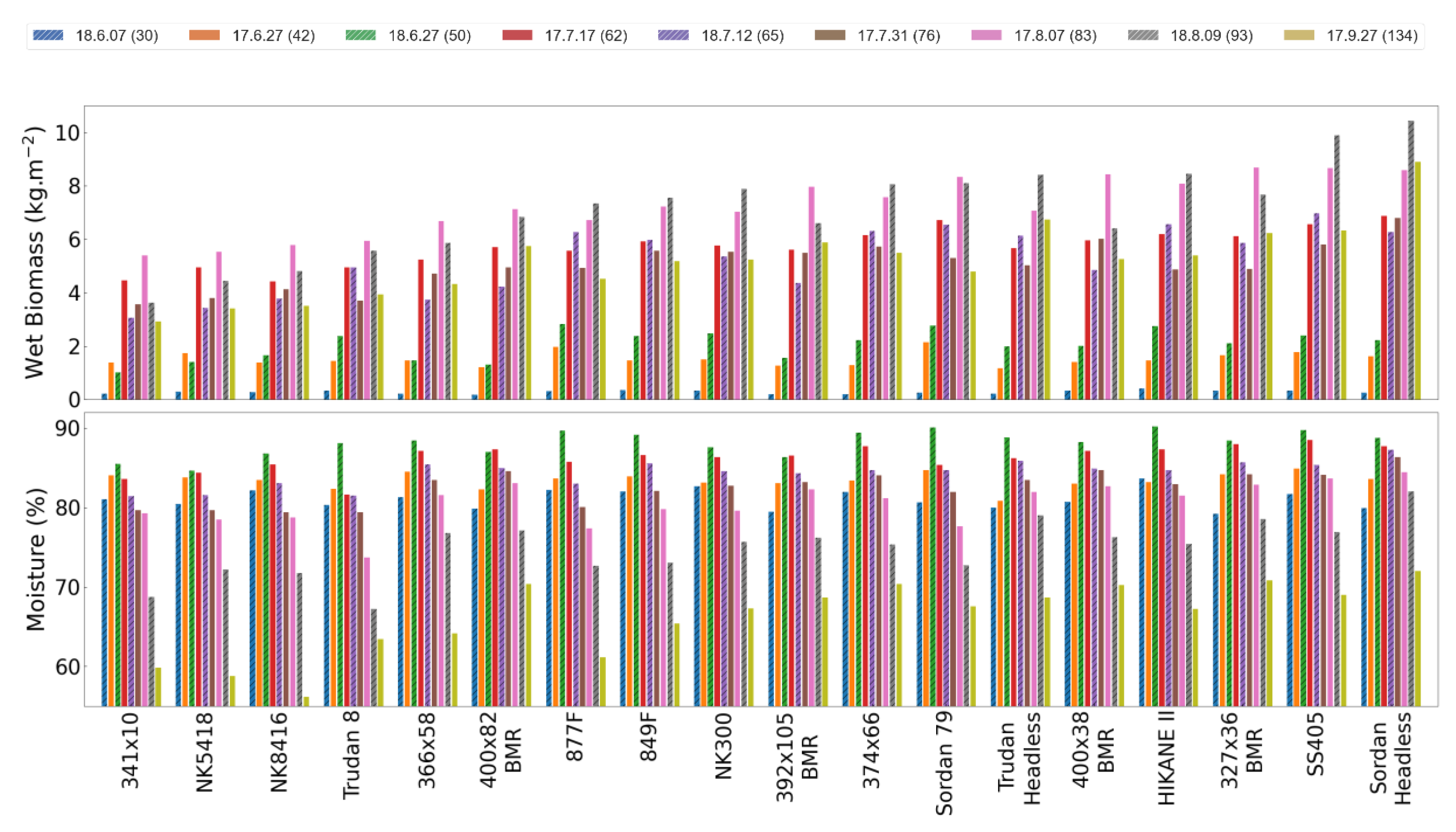
3.2. Ground Reference Data
3.3. Remote Sensing Data
3.4. Feature Extraction
3.4.1. Hyperspectral-Based Features
- Spectral Reflectance
- Vegetation Indices
- Integration Features
- Derivative Features
3.4.2. LiDAR-Based Features
- Height Percentile
- Canopy Volume
- Canopy Cover
- Height Statistics
3.5. Regression-Based Modeling Approaches
3.6. Statistical Analysis
4. Results
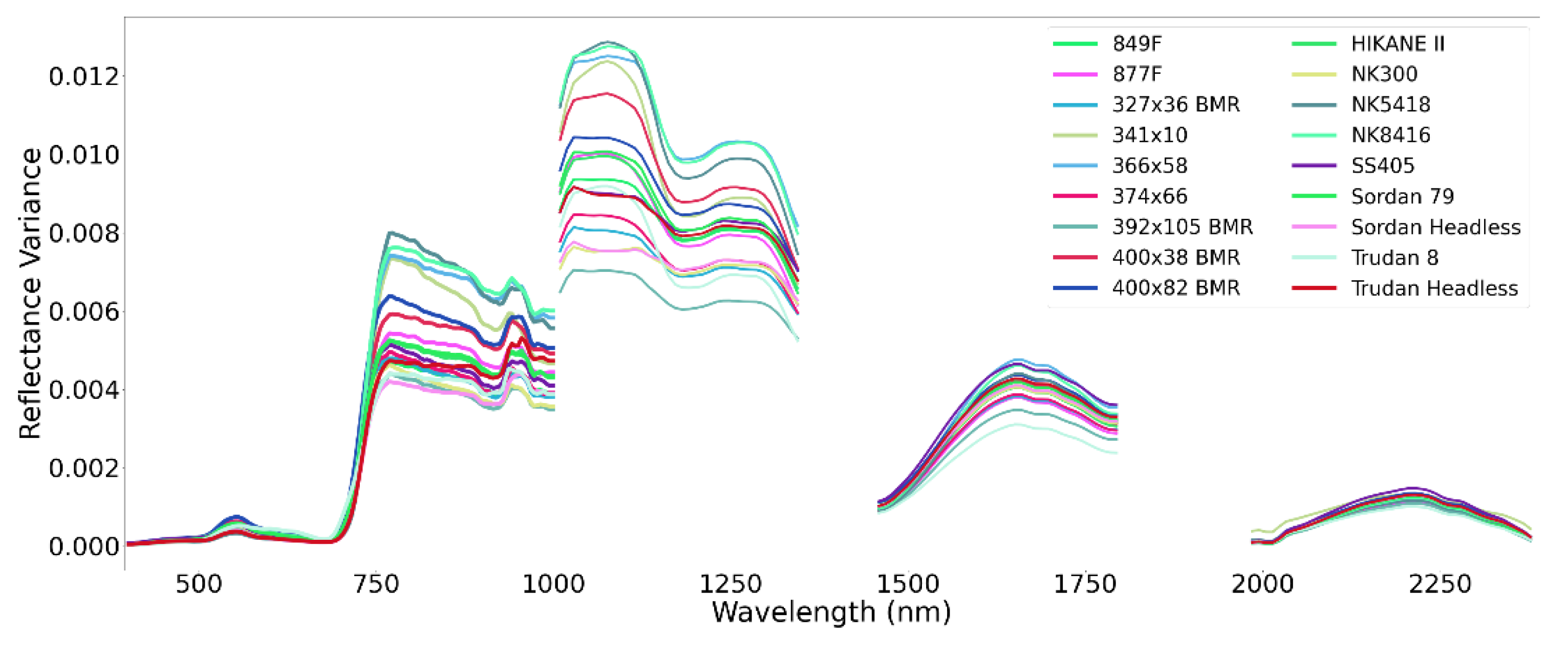
4.1. Data Screening
4.1.1. Time Series of Biomass Data
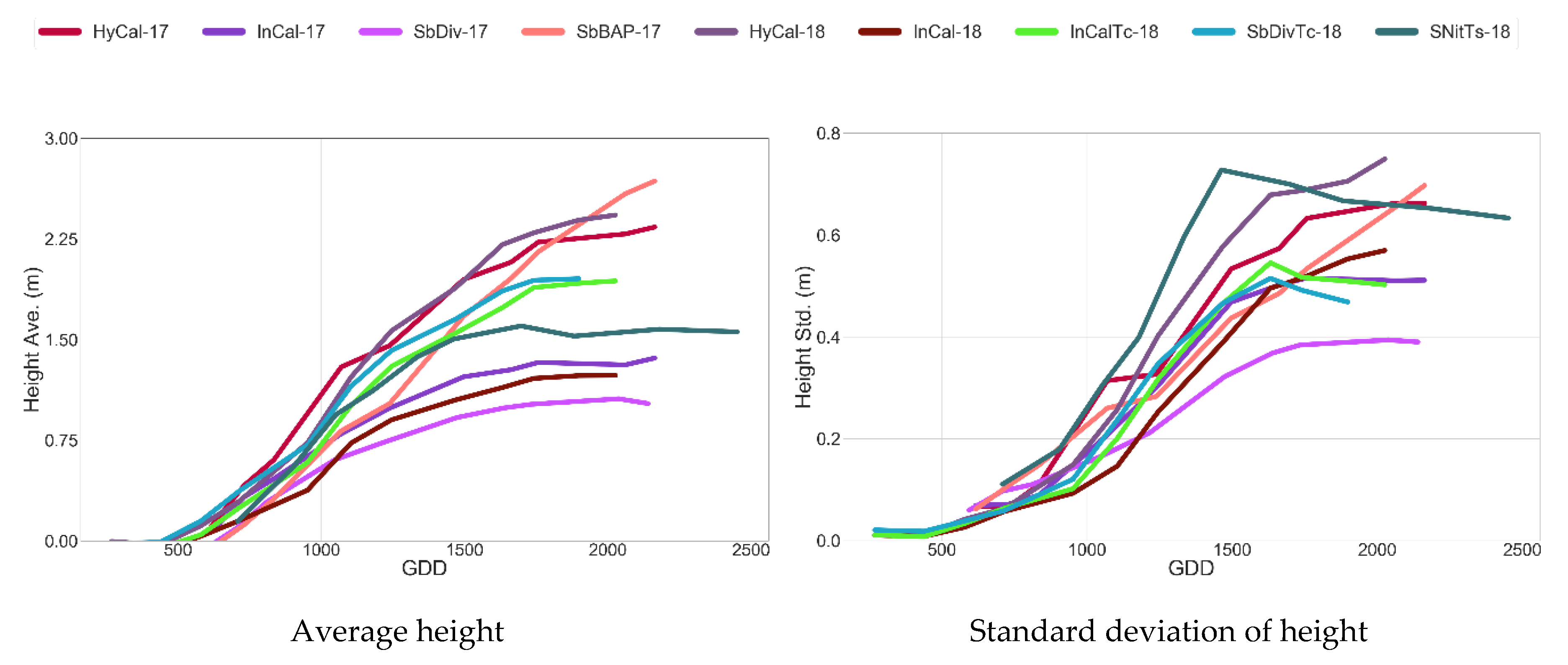
4.1.2. Time Series of Remote Sensing Data
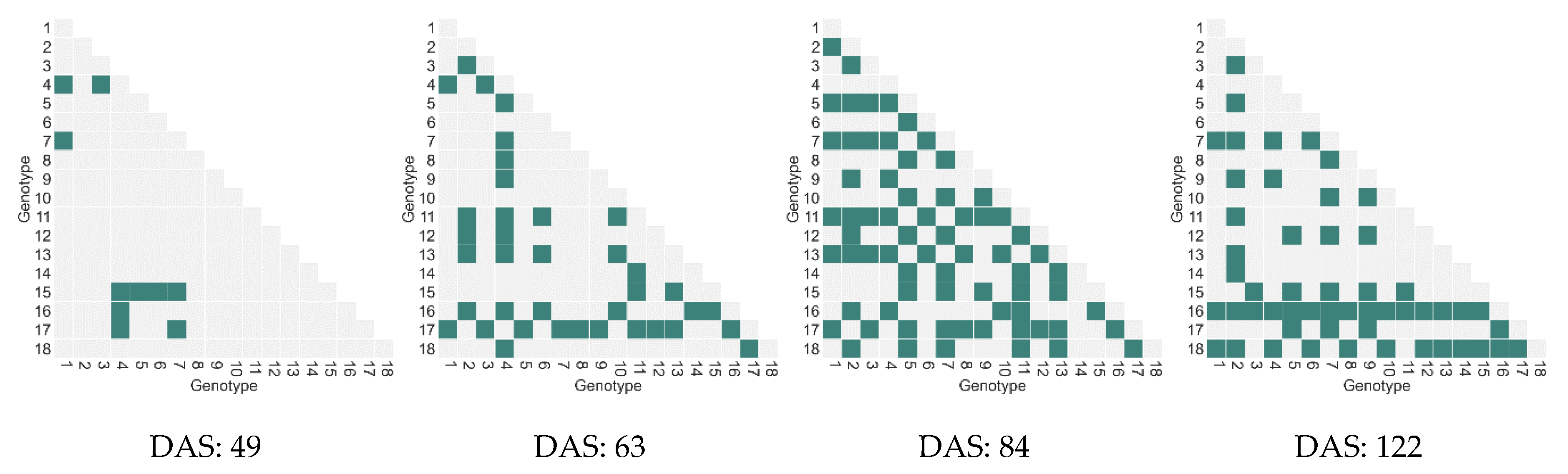
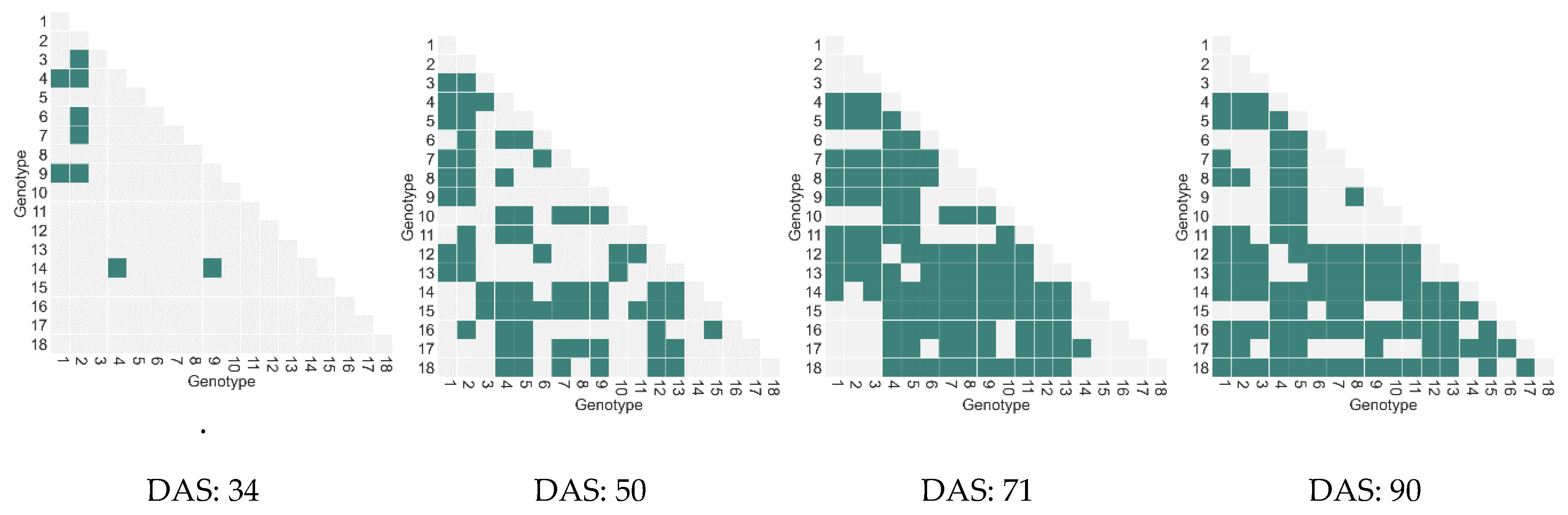
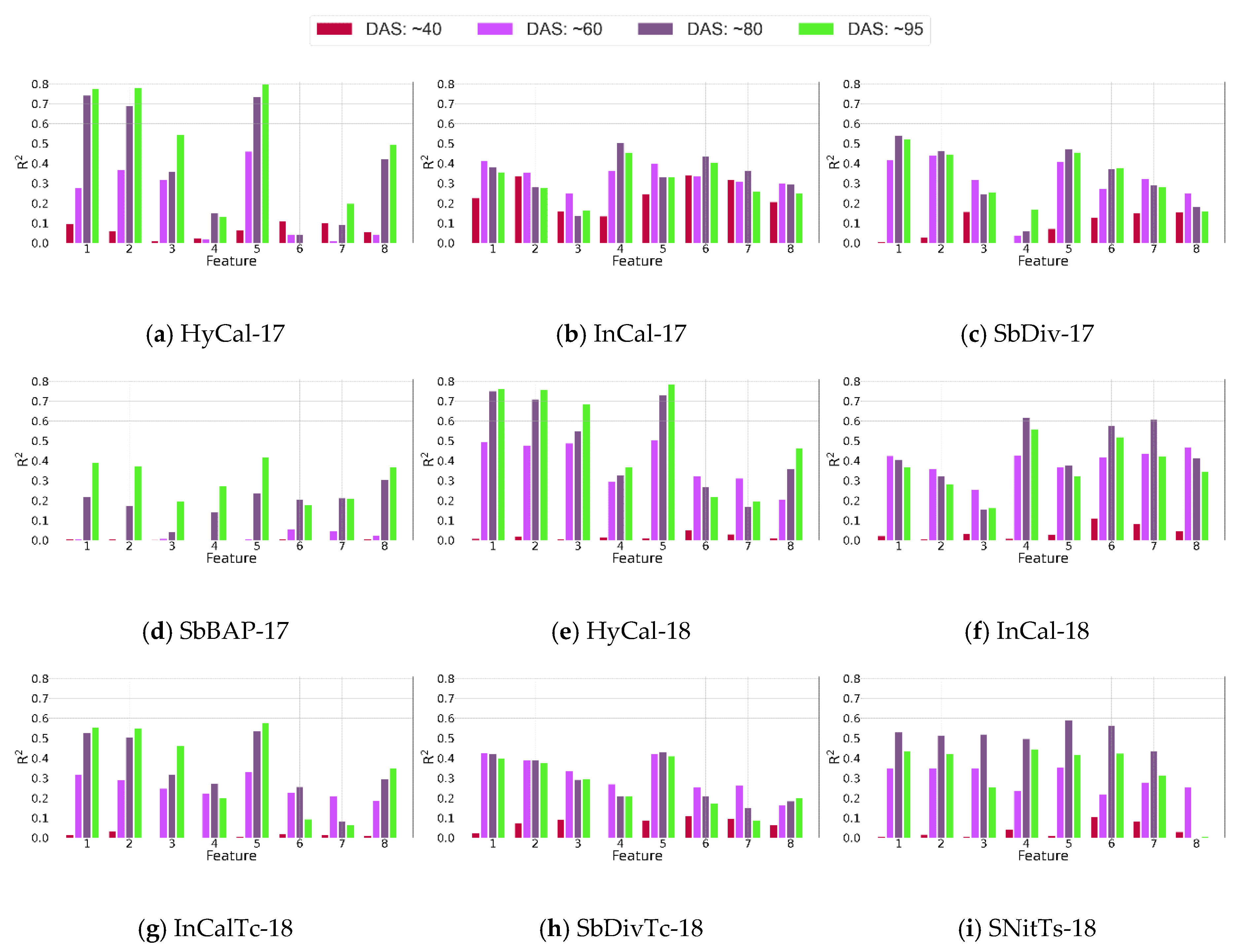
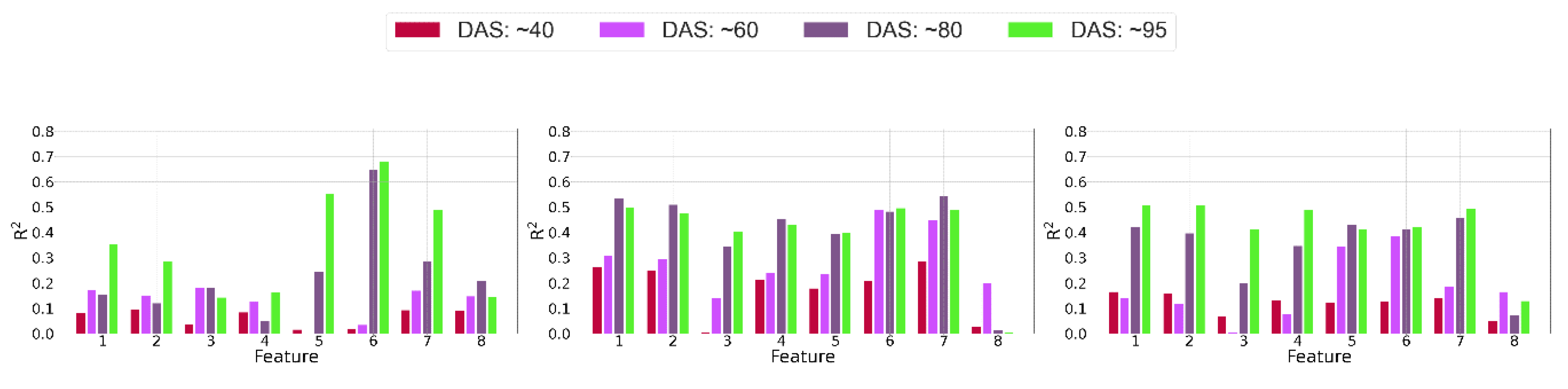
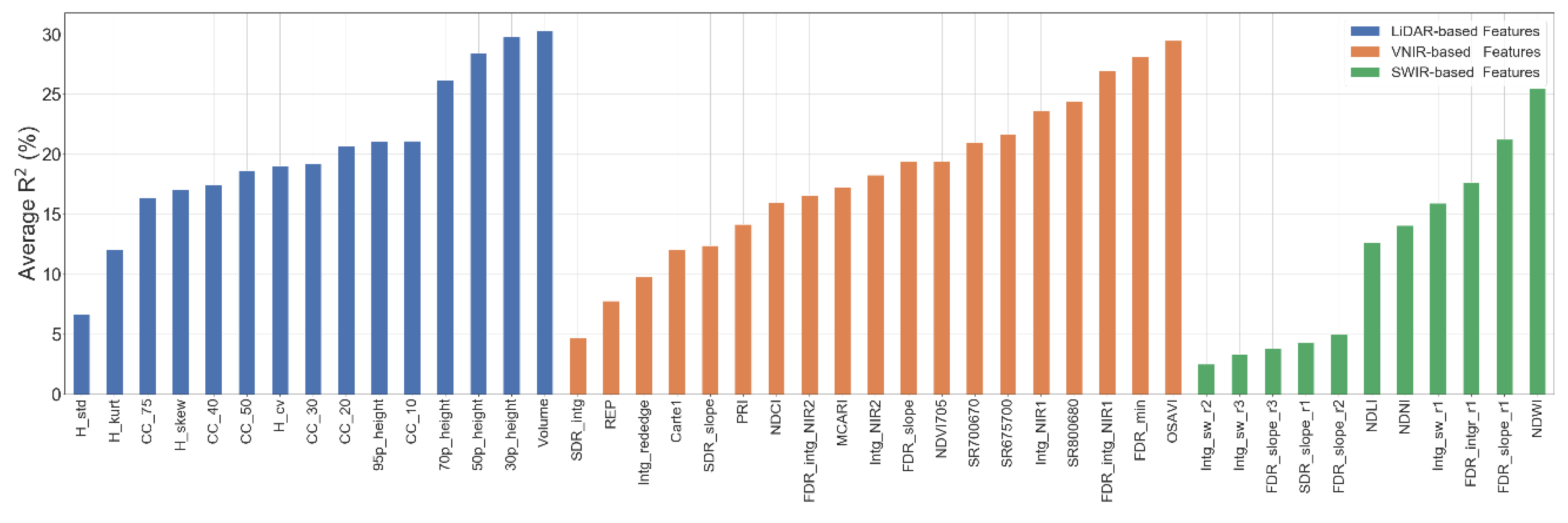
4.1.3. Relationship between Features and Biomass
4.2. Biomass Predictive Models
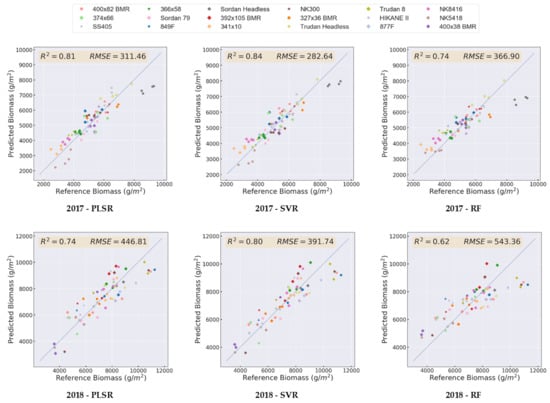
4.2.1. Impact of the Data Source and Regression Method on the Prediction Results
- (i)
- Diversity in the samples: the regression models are better able to learn the pattern in the data when the samples are more diverse.
- (ii)
- Number of data samples: the larger the number of data points in an experiment, the higher accuracies are typically achieved for the prediction.
- (iii)
- Similarity between the training and test data sets; if the training and test data sets are very different, then overfitting can occur for the training data set, resulting in decreased accuracy of the predictions. Note that this can happen when the number of data samples is limited, which causes unlike training and test sets, even when the samples are selected randomly. Additionally, if there is a significant range of biomass values in one experiment, there is more chance to have dissimilar training and test sets.
4.2.2. Predictions in Time
4.2.3. Multi-Temporal Predictions of End-of-Season Biomass
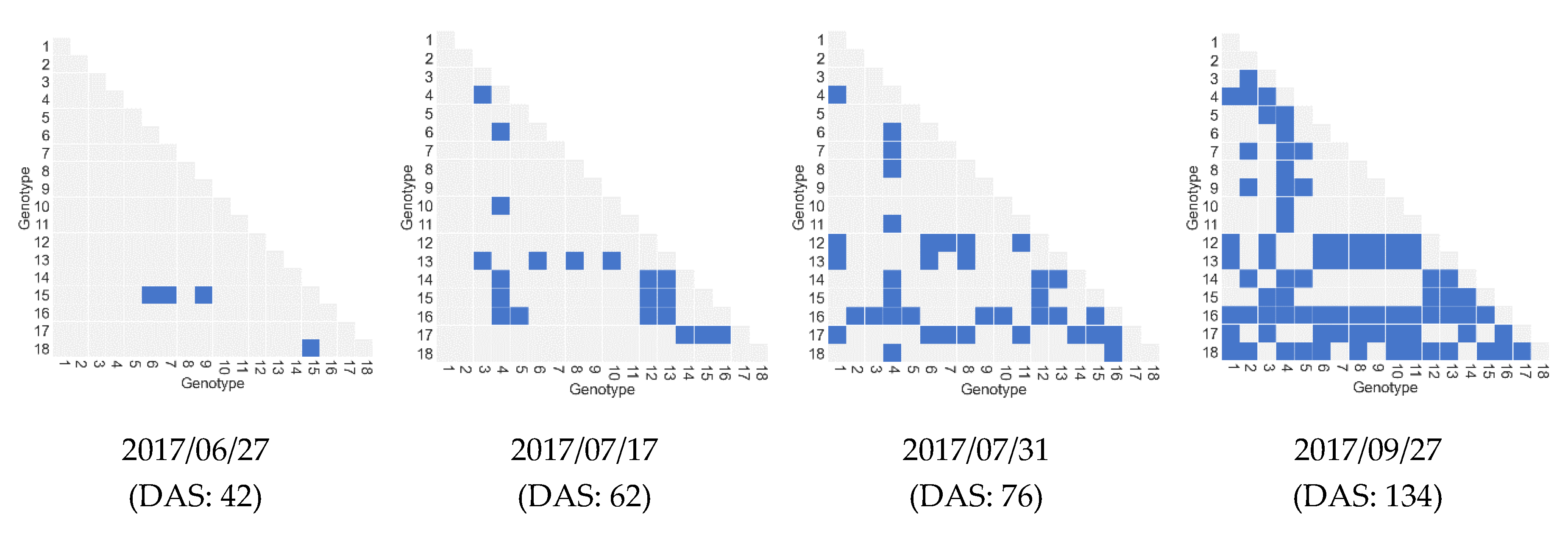
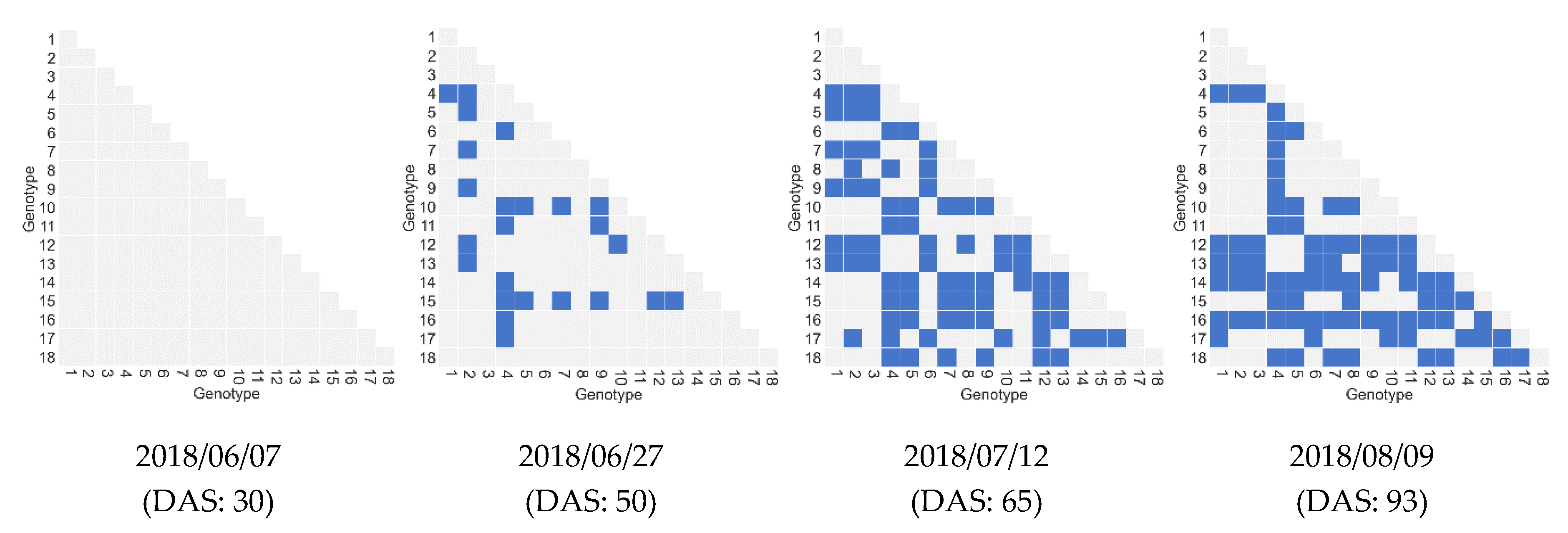
4.2.4. Impact of the Number of Features and Samples on Biomass Prediction
5. Discussion
Recommendations for Biomass Prediction
- Regression Model
- Data Source
- Flight Time and Frequency
- Required Reference Data
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Variety Name | Sorghum Type | Company |
|---|---|---|---|
| 1 | 849F | Forage | Pioneer |
| 2 | 877F | Forage | Pioneer |
| 3 | 327 × 36 BMR | Forage | Richardson |
| 4 | 341 × 10 | Forage | Richardson |
| 5 | 366 × 58 | Food grain | Richardson |
| 6 | 374 × 66 | Food grain | Richardson |
| 7 | 392 × 105 BMR | Forage | Richardson |
| 8 | 400 × 38 BMR | Sudangrass | Richardson |
| 9 | 400 × 82 BMR | Sudangrass | Richardson |
| 10 | HIKANE II | Forage | Sorghum Partners |
| 11 | NK300 | Forage | Sorghum Partners |
| 12 | NK5418 | Grain | Sorghum Partners |
| 13 | NK8416 | Grain | Sorghum Partners |
| 14 | SS405 | Forage | Sorghum Partners |
| 15 | Sordan 79 | Forage | Sorghum Partners |
| 16 | Sordan Headless | Forage (photoperiod sensitive) | Sorghum Partners |
| 17 | Trudan 8 | Forage | Sorghum Partners |
| 18 | Trudan Headless | Forage (photoperiod sensitive) | Sorghum Partners |
| Year | Data Type | Field | Dates |
|---|---|---|---|
| 2017 | RGB and LiDAR | InCal, HyCal, SbDiv, and SbBAP | 16/06, 21/06, 27/06, 05/07, 11/07, 14/07, 17/07, 25/07, 02/08, 08/08, 23/08, 3008 |
| VNIR | HyCal and InCal | 21/06, 28/06, 04/07, 12/07, 18/07, 25/07, 08/08, 23/08, 30/08, 10/09, 15/09 | |
| SbDiv | 21/06, 27/06, 04/07, 18/07, 25/07, 30/07, 08/08, 14/08, 23/08, 10/09, 24/09, 30/09 | ||
| SbBAP | 21/06, 27/06, 04/07, 18/07, 25/07, 30/07, 08/08, 14/08, 23/08, 10/09, 24/09 | ||
| SWIR | InCal and HyCal | 23/08, 30/08, 10/09, 15/09 | |
| SbDiv | 02/08, 08/08, 14/08, 23/08, 30/08, 10/09, 30/09 | ||
| SbBAP | 02/08, 08/08, 14/08, 23/08, 30/08, 10/09 | ||
| 2018 | RGB and LiDAR | HyCal, InCal, InCalTc, and SbDivTc | 22/05, 29/05, 04/05, 11/06, 20/06, 27/06, 02/07, 11/07, 18/07, 23/07, 01/08, 06/08 |
| SNitTs | 28/06, 03/07, 11/07, 17/17, 23/07, 01/08, 06/08, 16/08, 25/08, 05/09, 19/09 | ||
| VNIR and SWIR | HyCal, InCal, InCalTc | 04/06, 08/06, 14/06, 29/06, 03/07, 06/07, 11/07, 18/07, 25/07, 02/08, 09/08 | |
| SbDivTc | 04/06, 08/06, 14/06, 29/06, 03/07, 06/07, 10/07, 11/07, 25/07, 02/08 | ||
| SNitTs | 28/06, 03/07, 11/07, 18/07, 25/07, 02/08, 13/08, 28/08, 04/09, 12/09, 18/09 |
| Data Type | Index Name | Formulation | References |
|---|---|---|---|
| VNIR | NDVI | (R750 − R705)/(R750 + R705) | [70] |
| NDCI | (R762 − R527)/(R762 + R527) | [71] | |
| Carte1 | R695/R420 | [72] | |
| SR800,680 | R800/R680 | [73] | |
| SR675,700 | R675/R700 | [74] | |
| SR700,670 | R700/R670 | [75] | |
| OSAVI | (1 + 0.16) (R800 − R670)/(R800 + R670 + 0.16) | [76] | |
| MCARI | [(R700 − R670) − 0.2(R700-R550)](R700/R670) | [77] | |
| REP | 700 + 40[(R670 + R780)/2 − R700)]/(R740 − R700)) | [78] | |
| PRI | (R531 − R570)/(R531 + R570) | [79] | |
| SWIR | NDWI | (R860 − R1240)/(R860 + R1240) | [80] |
| NDLI | [log(1/R1754) − log(1/R1680)]/[log(1/R1754) + log(1/R1680)] | [81] | |
| NDNI | [log(1/R1510) − log(1/R1680)]/[log(1/R1510) + log(1/R1680)] | [81] |
| Data Type | Feature Name | λa | λb |
|---|---|---|---|
| VNIR | Intg_rededge | 685 | 745 |
| Intg_NIR1 | 770 | 910 | |
| Intg_NIR2 | 910 | 1000 | |
| SWIR | Intg_SWIR_r1 | 920 | 1353 |
| Intg_SWIR_r2 | 1430 | 1800 | |
| Intg_SWIR_r3 | 1952 | 2385 |
| Data Type | Feature Name | Description |
|---|---|---|
| VNIR | FDR_slope | slope of the line that passes through the minimum of FDR and the maximum of FDR in 660–690 nm range |
| FDR_min | minimum of FDR | |
| FDR_intg_ NIR1 | integration of FDR in bands between 670 and 780 nm | |
| FDR_intg_ NIR2 | integration of FDR in bands between 910 and 1000 nm | |
| SDR_slope_rededge | slope of the line passing through the maximum and the minimum of SDR | |
| SDR_intg | integration of SDR in all bands | |
| SWIR | FDR_slope_r1 | slope of the line that passes through the maximum of FDR in 1000–1050 nm range and the minimum of FDR in 1100–1200 nm range |
| FDR_slope_r2 | slope of the line that passes through the maximum of FDR in 1475–1525 nm range and the minimum of FDR in 1675–1725 nm range | |
| FDR_slope_r3 | slope of the line that passes through the maximum of FDR in 2000–2050 nm range and the minimum of FDR in 2200–2300 nm range | |
| FDR_intg-r1 | integration of FDR in bands between 920 and 1353 nm | |
| SDR_ slope_r1 | slope of the line that passes through the maximum of SDR in 1100–1200 nm range and the minimum of SDR in 1000–1100 nm range |
| Algorithm | Hyperparameter | Values Tested |
|---|---|---|
| PLSR | Number of components | 2, 3, 5, 10, 15, 20 |
| SVR (RBF) | C | 10, 100, 1000, number of features |
| gamma | 1/n, 0.0001, 0.001, 0.01, 0.1 | |
| Random Forest (RF) | Max tree depth | 5, 10, 100 |
| Min sample split | 2, 10 | |
| Number of trees | 50, 100, 500 |
Appendix B






References
- Davey, J.W.; Hohenlohe, P.A.; Etter, P.D.; Boone, J.Q.; Catchen, J.M.; Blaxter, M.L. Genome-wide genetic marker discovery and genotyping using next-generation sequencing. Nat. Rev. Genet. 2011, 12, 499–510. [Google Scholar] [CrossRef]
- Potgieter, A.B.; George-Jaeggli, B.; Chapman, S.C.; Laws, K.; Suárez Cadavid, L.A.; Wixted, J.; Watson, J.; Eldridge, M.; Jordan, D.R.; Hammer, G.L. Multi-Spectral Imaging from an Unmanned Aerial Vehicle Enables the Assessment of Seasonal Leaf Area Dynamics of Sorghum Breeding Lines. Front. Plant Sci. 2017, 8, 1532–1546. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Di, L.; Zhang, L.; Deng, M.; Qin, Z.; Zhao, S.; Lin, H. Estimation of crop LAI using hyperspectral vegetation indices and a hybrid inversion method. Remote Sens. Environ. 2015, 165, 123–134. [Google Scholar] [CrossRef]
- Chu, T.; Starek, M.J.; Brewer, M.J.; Murray, S.C.; Pruter, L.S. Characterizing canopy height with UAS structure-from-motion photogrammetry—Results analysis of a maize field trial with respect to multiple factors. Remote Sens. Lett. 2018, 9, 753–762. [Google Scholar] [CrossRef]
- Pugh, N.A.; Horne, D.W.; Murray, S.C.; Carvalho, G.; Malambo, L.; Jung, J.; Chang, A.; Maeda, M.; Popescu, S.; Chu, T.; et al. Temporal Estimates of Crop Growth in Sorghum and Maize Breeding Enabled by Unmanned Aerial Systems. Plant Phenome J. 2018, 1, 1–10. [Google Scholar] [CrossRef]
- Maimaitijiang, M.; Ghulam, A.; Sidike, P.; Hartling, S.; Maimaitiyiming, M.; Peterson, K.; Shavers, E.; Fishman, J.; Peterson, J.; Kadam, S.; et al. Unmanned Aerial System (UAS)-based phenotyping of soybean using multi-sensor data fusion and extreme learning machine. ISPRS J. Photogramm. Remote Sens. 2017, 134, 43–58. [Google Scholar] [CrossRef]
- Tattaris, M.; Reynolds, M.P.; Chapman, S.C. A Direct Comparison of Remote Sensing Approaches for High-Throughput Phenotyping in Plant Breeding. Front. Plant Sci. 2016, 7, 1–9. [Google Scholar] [CrossRef]
- Eitel, J.U.H.; Magney, T.S.; Vierling, L.A.; Greaves, H.E.; Zheng, G. An automated method to quantify crop height and calibrate satellite-derived biomass using hypertemporal lidar. Remote Sens. Environ. 2016, 187, 414–422. [Google Scholar] [CrossRef]
- Li, J.; Shi, Y.; Veeranampalayam-Sivakumar, A.N.; Schachtman, D.P. Elucidating sorghum biomass, nitrogen and chlorophyll contents with spectral and morphological traits derived from unmanned aircraft system. Front. Plant Sci. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Sun, C.; Feng, L.; Zhang, Z.; Ma, Y.; Crosby, T.; Naber, M.; Wang, Y. Prediction of end-of-season tuber yield and tuber set in potatoes using in-season uav-based hyperspectral imagery and machine learning. Sensors 2020, 20, 5293. [Google Scholar] [CrossRef]
- Li, B.; Xu, X.; Zhang, L.; Han, J.; Bian, C.; Li, G.; Liu, J.; Jin, L. Above-ground biomass estimation and yield prediction in potato by using UAV-based RGB and hyperspectral imaging. ISPRS J. Photogramm. Remote Sens. 2020, 162, 161–172. [Google Scholar] [CrossRef]
- Duan, T.; Chapman, S.C.; Guo, Y.; Zheng, B. Dynamic monitoring of NDVI in wheat agronomy and breeding trials using an unmanned aerial vehicle. Field Crop. Res. 2017, 210, 71–80. [Google Scholar] [CrossRef]
- Stanton, C.; Starek, M.J.; Elliott, N.; Brewer, M.; Maeda, M.M.; Chu, T. Unmanned aircraft system-derived crop height and normalized difference vegetation index metrics for sorghum yield and aphid stress assessment. J. Appl. Remote Sens. 2017, 11, 026035. [Google Scholar] [CrossRef]
- Gracia-Romero, A.; Kefauver, S.C.; Fernandez-Gallego, J.A.; Vergara-Díaz, O.; Nieto-Taladriz, M.T.; Araus, J.L. UAV and ground image-based phenotyping: A proof of concept with durum wheat. Remote Sens. 2019, 11, 1244. [Google Scholar] [CrossRef]
- Perich, G.; Hund, A.; Anderegg, J.; Roth, L.; Boer, M.P.; Walter, A.; Liebisch, F.; Aasen, H. Assessment of Multi-Image Unmanned Aerial Vehicle Based High-Throughput Field Phenotyping of Canopy Temperature. Front. Plant Sci. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Borra-Serrano, I.; Swaef, T.D.; Quataert, P.; Aper, J.; Saleem, A.; Saeys, W.; Somers, B.; Roldán-Ruiz, I.; Lootens, P. Closing the phenotyping gap: High resolution UAV time series for soybean growth analysis provides objective data from field trials. Remote Sens. 2020, 12, 1644. [Google Scholar] [CrossRef]
- Zhang, Z.; Masjedi, A.; Zhao, J.; Crawford, M.M. Prediction of sorghum biomass based on image based features derived from time series of UAV images. In Proceedings of the International Geoscience and Remote Sensing Symposium (IGARSS), Fort Worth, TX, USA, 23–28 July 2017; pp. 6154–6157. [Google Scholar]
- Lewis, B.; Smith, I.; Fowler, M.; Licato, J. The robot mafia: A test environment for deceptive robots. In Proceedings of the 28th Modern Artificial Intelligence and Cognitive Science Conference, MAICS 2017, Fort Wayne, IN, USA, 28–29 April 2017; pp. 189–190. [Google Scholar]
- Masjedi, A.; Zhao, J.; Thompson, A.M.; Yang, K.W.; Flatt, J.E.; Crawford, M.M.; Ebert, D.S.; Tuinstra, M.R.; Hammer, G.; Chapman, S. Sorghum biomass prediction using uav-based remote sensing data and crop model simulation. In Proceedings of the International Geoscience and Remote Sensing Symposium (IGARSS), Valencia, Spain, 23–27 July 2018; pp. 7719–7722. [Google Scholar]
- Ostos-Garrido, F.J.; de Castro, A.I.; Torres-Sánchez, J.; Pistón, F.; Peña, J.M. High-Throughput Phenotyping of Bioethanol Potential in Cereals Using UAV-Based Multi-Spectral Imagery. Front. Plant Sci. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Sagan, V.; Maimaitijiang, M.; Sidike, P.; Eblimit, K.; Peterson, K.T.; Hartling, S.; Esposito, F.; Khanal, K.; Newcomb, M.; Pauli, D.; et al. UAV-based high resolution thermal imaging for vegetation monitoring, and plant phenotyping using ICI 8640 P, FLIR Vue Pro R 640, and thermomap cameras. Remote Sens. 2019, 11, 330. [Google Scholar] [CrossRef]
- Holman, F.H.; Riche, A.B.; Castle, M.; Wooster, M.J.; Hawkesford, M.J. Radiometric calibration of “commercial offthe shelf” cameras for UAV-based high-resolution temporal crop phenotyping of reflectance and NDVI. Remote Sens. 2019, 11, 1657. [Google Scholar] [CrossRef]
- Enciso, J.; Avila, C.A.; Jung, J.; Elsayed-Farag, S.; Chang, A.; Yeom, J.; Landivar, J.; Maeda, M.; Chavez, J.C. Validation of agronomic UAV and field measurements for tomato varieties. Comput. Electron. Agric. 2019, 158, 278–283. [Google Scholar] [CrossRef]
- Ampatzidis, Y.; Partel, V. UAV-based high throughput phenotyping in citrus utilizing multispectral imaging and artificial intelligence. Remote Sens. 2019, 11, 410. [Google Scholar] [CrossRef]
- Fernandes, S.B.; Dias, K.O.G.; Ferreira, D.F.; Brown, P.J. Efficiency of multi-trait, indirect, and trait-assisted genomic selection for improvement of biomass sorghum. Theor. Appl. Genet. 2018, 131, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Ogbaga, C.C.; Bajhaiya, A.K.; Gupta, S.K. Improvements in biomass production: Learning lessons from the bioenergy plants maize and sorghum. J. Environ. Biol. 2019, 40, 400–406. [Google Scholar] [CrossRef]
- Prabhakara, K.; Dean Hively, W.; McCarty, G.W. Evaluating the relationship between biomass, percent groundcover and remote sensing indices across six winter cover crop fields in Maryland, United States. Int. J. Appl. Earth Obs. Geoinf. 2015, 39, 88–102. [Google Scholar] [CrossRef]
- Moghimi, A.; Yang, C.; Anderson, J.A. Aerial hyperspectral imagery and deep neural networks for high-throughput yield phenotyping in wheat. Comput. Electron. Agric. 2020, 172, 105299. [Google Scholar] [CrossRef]
- Zhao, J.; Karimzadeh, M.; Masjedi, A.; Wang, T.; Zhang, X.; Crawford, M.M.; Ebert, D.S. FeatureExplorer: Interactive Feature Selection and Exploration of Regression Models for Hyperspectral Images. In Proceedings of the 2019 IEEE Visualization Conference VIS, Vancouver, BC, Canada, 20–25 October 2019; pp. 161–165. [Google Scholar] [CrossRef]
- Feng, W.; Guo, B.B.; Zhang, H.Y.; He, L.; Zhang, Y.S.; Wang, Y.H.; Zhu, Y.J.; Guo, T.C. Remote estimation of above ground nitrogen uptake during vegetative growth in winter wheat using hyperspectral red-edge ratio data. Field Crop. Res. 2015, 180, 197–206. [Google Scholar] [CrossRef]
- Foster, A.J.; Kakani, V.G.; Mosali, J. Estimation of bioenergy crop yield and N status by hyperspectral canopy reflectance and partial least square regression. Precis. Agric. 2017, 18, 192–209. [Google Scholar] [CrossRef]
- Yue, J.; Feng, H.; Yang, G.; Li, Z. A comparison of regression techniques for estimation of above-ground winter wheat biomass using near-surface spectroscopy. Remote Sens. 2018, 10, 66. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Hartig, F.; Latifi, H.; Berger, C.; Hernández, J.; Corvalán, P.; Koch, B. Importance of sample size, data type and prediction method for remote sensing-based estimations of aboveground forest biomass. Remote Sens. Environ. 2014, 154, 102–114. [Google Scholar] [CrossRef]
- Vaglio Laurin, G.; Puletti, N.; Chen, Q.; Corona, P.; Papale, D.; Valentini, R. Above ground biomass and tree species richness estimation with airborne lidar in tropical Ghana forests. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 371–379. [Google Scholar] [CrossRef]
- Harkel, J.T.; Bartholomeus, H.; Kooistra, L. Biomass and crop height estimation of different crops using UAV-based LiDAR. Remote Sens. 2020, 12, 17. [Google Scholar] [CrossRef]
- McGlinchy, J.; Van Aardt, J.A.N.; Erasmus, B.; Asner, G.P.; Mathieu, R.; Wessels, K.; Knapp, D.; Kennedy-Bowdoin, T.; Rhody, H.; Kerekes, J.P.; et al. Extracting structural vegetation components from small-footprint waveform lidar for biomass estimation in savanna ecosystems. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 480–490. [Google Scholar] [CrossRef]
- Shao, G.; Shao, G.; Gallion, J.; Saunders, M.R.; Frankenberger, J.R.; Fei, S. Improving Lidar-based aboveground biomass estimation of temperate hardwood forests with varying site productivity. Remote Sens. Environ. 2018, 204, 872–882. [Google Scholar] [CrossRef]
- Phua, M.H.; Johari, S.A.; Wong, O.C.; Ioki, K.; Mahali, M.; Nilus, R.; Coomes, D.A.; Maycock, C.R.; Hashim, M. Synergistic use of Landsat 8 OLI image and airborne LiDAR data for above-ground biomass estimation in tropical lowland rainforests. For. Ecol. Manag. 2017, 406, 163–171. [Google Scholar] [CrossRef]
- Vastaranta, M.; Holopainen, M.; Karjalainen, M.; Kankare, V.; Hyyppa, J.; Kaasalainen, S. TerraSAR-X stereo radargrammetry and airborne scanning LiDAR height metrics in imputation of forest aboveground biomass and stem volume. IEEE Trans. Geosci. Remote Sens. 2014, 52, 1197–1204. [Google Scholar] [CrossRef]
- Zhao, K.; Suarez, J.C.; Garcia, M.; Hu, T.; Wang, C.; Londo, A. Utility of multitemporal lidar for forest and carbon monitoring: Tree growth, biomass dynamics, and carbon flux. Remote Sens. Environ. 2018, 204, 883–897. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, C.; Yang, H.; Yang, G.; Han, L.; Li, Z.; Feng, H.; Xu, B.; Wu, J.; Lei, L. Estimation of maize above-ground biomass based on stem-leaf separation strategy integrated with LiDAR and optical remote sensing data. PeerJ 2019, 7, 1–30. [Google Scholar] [CrossRef]
- Luo, S.; Wang, C.; Xi, X.; Nie, S.; Fan, X.; Chen, H.; Yang, X.; Peng, D.; Lin, Y.; Zhou, G. Combining hyperspectral imagery and LiDAR pseudo-waveform for predicting crop LAI, canopy height and above-ground biomass. Ecol. Indic. 2019, 102, 801–812. [Google Scholar] [CrossRef]
- Chao, Z.; Liu, N.; Zhang, P.; Ying, T.; Song, K. Estimation methods developing with remote sensing information for energy crop biomass: A comparative review. Biomass Bioenergy 2019, 122, 414–425. [Google Scholar] [CrossRef]
- Vaglio Laurin, G.; Chen, Q.; Lindsell, J.A.; Coomes, D.A.; Frate, F.D.; Guerriero, L.; Pirotti, F.; Valentini, R. Above ground biomass estimation in an African tropical forest with lidar and hyperspectral data. ISPRS J. Photogramm. Remote Sens. 2014, 89, 49–58. [Google Scholar] [CrossRef]
- Ravi, R.; Lin, Y.J.; Elbahnasawy, M.; Shamseldin, T.; Habib, A. Simultaneous System Calibration of a Multi-LiDAR Multicamera Mobile Mapping Platform. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2018, 11, 1694–1714. [Google Scholar] [CrossRef]
- LaForest, L.; Hasheminasab, S.M.; Zhou, T.; Flatt, J.E.; Habib, A. New strategies for time delay estimation during system calibration for UAV-Based GNSS/INS-Assisted imaging systems. Remote Sens. 2019, 11, 1811. [Google Scholar] [CrossRef]
- He, F.; Zhou, T.; Xiong, W.; Hasheminnasab, S.M.; Habib, A. Automated aerial triangulation for UAV-based mapping. Remote Sens. 2018, 10, 1952. [Google Scholar] [CrossRef]
- Hasheminasab, S.M.; Zhou, T.; Habib, A. GNSS/INS-Assisted structure from motion strategies for UAV-Based imagery over mechanized agricultural fields. Remote Sens. 2020, 12, 351. [Google Scholar] [CrossRef]
- Habib, A.; Zhou, T.; Masjedi, A.; Zhang, Z.; Evan Flatt, J.; Crawford, M. Boresight Calibration of GNSS/INS-Assisted Push-Broom Hyperspectral Scanners on UAV Platforms. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2018, 11, 1734–1749. [Google Scholar] [CrossRef]
- Liu, Y.-K.; Li, C.-R.; Ma, L.-L.; Qian, Y.-G.; Wang, N.; Gao, C.-X.; Tang, L.-L. Land surface reflectance retrieval from optical hyperspectral data collected with an unmanned aerial vehicle platform. Opt. Express 2019, 27, 7174. [Google Scholar] [CrossRef]
- Thorp, K.R.; Wang, G.; Bronson, K.F.; Badaruddin, M.; Mon, J. Hyperspectral data mining to identify relevant canopy spectral features for estimating durum wheat growth, nitrogen status, and grain yield. Comput. Electron. Agric. 2017, 136, 1–12. [Google Scholar] [CrossRef]
- Demetriades-Shah, T.H.; Steven, M.D.; Clark, J.A. High resolution derivative spectra in remote sensing. Remote Sens. Environ. 1990, 33, 55–64. [Google Scholar] [CrossRef]
- Feng, W.; Guo, B.B.; Wang, Z.J.; He, L.; Song, X.; Wang, Y.H.; Guo, T.C. Measuring leaf nitrogen concentration in winter wheat using double-peak spectral reflection remote sensing data. Field Crop. Res. 2014, 159, 43–52. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M.J.E. Smoothing and Differentiation of Data by Simplified Least Squares Procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E. Spectral and chemical analysis of tropical forests: Scaling from leaf to canopy levels. Remote Sens. Environ. 2008, 112, 3958–3970. [Google Scholar] [CrossRef]
- Zhao, Y.R.; Li, X.; Yu, K.Q.; Cheng, F.; He, Y. Hyperspectral Imaging for Determining Pigment Contents in Cucumber Leaves in Response to Angular Leaf Spot Disease. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Féret, J.B.; François, C.; Gitelson, A.; Asner, G.P.; Barry, K.M.; Panigada, C.; Richardson, A.D.; Jacquemoud, S. Optimizing spectral indices and chemometric analysis of leaf chemical properties using radiative transfer modeling. Remote Sens. Environ. 2011, 115, 2742–2750. [Google Scholar] [CrossRef]
- Ullah, S.; Skidmore, A.K.; Ramoelo, A.; Groen, T.A.; Naeem, M.; Ali, A. Retrieval of leaf water content spanning the visible to thermal infrared spectra. ISPRS J. Photogramm. Remote Sens. 2014, 93, 56–64. [Google Scholar] [CrossRef]
- Thulin, S.; Hill, M.J.; Held, A.; Jones, S.; Woodgate, P. Predicting Levels of Crude Protein, Digestibility, Lignin and Cellulose in Temperate Pastures Using Hyperspectral Image Data. Am. J. Plant Sci. 2014, 05, 997–1019. [Google Scholar] [CrossRef]
- Ecarnot, M.; Compan, F.; Roumet, P. Assessing leaf nitrogen content and leaf mass per unit area of wheat in the field throughout plant cycle with a portable spectrometer. Field Crop. Res. 2013, 140, 44–50. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Bao, Y.; Luo, J.; Jin, X.; Xu, X.; Song, X.; Yang, G. Exploring the best hyperspectral features for LAI estimation using partial least squares regression. Remote Sens. 2014, 6, 6221–6241. [Google Scholar] [CrossRef]
- Zhang, T. An Introduction to Support Vector Machines and Other Kernel-Based Learning Methods A Review; Cambridge University Press: Cambridge, UK, 2001; Volume 22, ISBN 0521780195. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Belgiu, M.; Drăgu, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Blondel, M.; Brucher, M.; Buitinck, L.; Cournapeau, D.; Dawe, N.; Du, S.; Dubourg, V.; Duchesnay, E.; Fabisch, A.; Fritsch, V.; et al. Scikit-learn. J. Mach. Learn. Res. 2015, 12, 2825–2830. [Google Scholar] [CrossRef]
- Sokal, R.R.; James Rohlf, F. Biometry: The Principles and Practice of Statistics in Biological Research; W. H. Freeman: New York, NY, USA, 1995. [Google Scholar]
- Seabold, S.; Perktold, J. Statsmodels: Econometric and Statistical Modeling with Python. In Proceedings of the 9th Python in Science Conference, Austin, TX, USA, 28 June–3 July 2010; pp. 92–96. [Google Scholar]
- Gerik, T.; Bean, B.; Vanderlip, R. Sorghum Growth and Development; Texas FARMER Collection, Texas Agrilife Extension, Texas A&M University: College Station, TX, USA, 2003. [Google Scholar]
- De Almeida, C.T.; Galvão, L.S.; de Aragão, L.E.O.C.e.; Ometto, J.P.H.B.; Jacon, A.D.; de Pereira, F.R.S.; Sato, L.Y.; Lopes, A.P.; de Graça, P.M.L.A.; de Silva, C.V.J.; et al. Combining LiDAR and hyperspectral data for aboveground biomass modeling in the Brazilian Amazon using different regression algorithms. Remote Sens. Environ. 2019, 232, 111323. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M.N. Quantitative estimation of chlorophyll-a using reflectance spectra: Experiments with autumn chestnut and maple leaves. J. Photochem. Photobiol. B Biol. 1994, 22, 247–252. [Google Scholar] [CrossRef]
- Marshak, A.; Knyazikhin, Y.; Davis, A.B.; Wiscombe, W.J.; Pilewskie, P. Cloud-vegetation interaction: Use of normalized difference cloud index for estimation of cloud optical thickness. Geophys. Res. Lett. 2000, 27, 1695–1698. [Google Scholar] [CrossRef]
- Carter, G.A. Ratios of leaf reflectances in narrow wavebands as indicators of plant stress. Int. J. Remote Sens. 1994, 15, 517–520. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance acrossa wide range of species, leaf structures and developmental stages. Int. J. Remote Sens. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a green channel in remote sensing of global vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- McMurtrey, J.E.; Chappelle, E.W.; Kim, M.S.; Meisinger, J.J.; Corp, L.A. Distinguishing nitrogen fertilization levels in field corn (Zea mays L.) with actively induced fluorescence and passive reflectance measurements. Remote Sens. Environ. 1994, 47, 36–44. [Google Scholar] [CrossRef]
- Rondeaux, G.; Steven, M.; Baret, F. Optimization of soil-adjusted vegetation indices. Remote Sens. Environ. 1996, 55, 95–107. [Google Scholar] [CrossRef]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; de Colstoun, E.B.; McMurtrey, J.E. Estimating Corn Leaf Chlorophyll Concentration from Leaf and Canopy Reflectance. Remote Sens. Environ. 2000, 35, 229–239. [Google Scholar] [CrossRef]
- Clevers, J.G.P.W. Imaging Spectrometry in Agriculture—Plant Vitality And Yield Indicators BT—Imaging Spectrometry—A Tool for Environmental Observations; Hill, J., Mégier, J., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 193–219. ISBN 978-0-585-33173-7. [Google Scholar]
- Gamon, J.A.; Peñuelas, J.; Field, C.B. A Narrow-Waveband Spectral Index That Tracks Diurnal Changes in Photosynthetic Efficiency. Remote Sens. Environ. 1992, 44, 35–44. [Google Scholar] [CrossRef]
- Gao, B.-C. NDWI A Normalized Difference Water Index for Remote Sensing of Vegetation Liquid Water From Space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Serrano, L.; Peñuelas, J.; Ustin, S.L. Remote sensing of nitrogen and lignin in Mediterranean vegetation from AVIRIS data: Decomposing biochemical from structural signals. Remote Sens. Environ. 2002, 81, 355–364. [Google Scholar] [CrossRef]



























| Trial | Year | Genotype | # of Plots | # of Genotypes | Sowing Date | Harvest Date | Available Biomass Data |
|---|---|---|---|---|---|---|---|
| HyCal-17 | 2017 | Hybrid | 72 | 18 | 16/05 | 27/09 | 27/06, 17/07, 31/07, 08/08, 27/09 |
| InCal-17 | 2017 | Inbred | 120 | 60 | 16/05 | 27/09 | 27/09 |
| SbBAP-17 | 2017 | Inbred | 760 | 350 | 16/05 | 28/09 | 28/09 |
| SbDiv-17 | 2017 | Inbred | 1800 | 840 | 17/05 | 09/11 | 09/11 |
| HyCal-18 | 2018 | Hybrid | 72 | 18 | 08/05 | 09/08 | 27/06, 12/07, 09/08 |
| InCal-18 | 2018 | Inbred | 108 | 54 | 08/05 | 09/08 | 09/08 |
| InCalTc-18 | 2018 | Hybrid | 108 | 54 | 08/05 | 06/08 | 06/08 |
| SbDivTc-18 | 2018 | Hybrid | 1260 | 630 | 08/05 | 02/08 | 02/08 and 14/08 |
| SNitTs-18 | 2018 | Inbred | 112 | 4 | 04/06 | 02/10 | 02/10 |
| Sensor | Description |
|---|---|
| RGB | Sony Alpha ILCE-7R Sony 35mm Lens Full-frame 36.4MP |
| LiDAR | Velodyne VLP-16 600 rotations per minute (RPM), 360-degree horizontal FOV Maximum range of 100 m |
| VNIR | Headwall Photonics Nano-Hyperspec imaging sensor 272 spectral bands at 2.2 nm/band from 400 nm to 1000 nm 640 spatial channels at 7.4 µm/pixel, 12 mm lens (in 2017) and 8 mm lens (in 2018) |
| SWIR | Headwall Photonics Micro-Hyperspec pushbroom 166 spectral bands at 10 nm/band from 900 nm to 2500 nm 384 spatial channels at 24 µm/pixel, 25 mm lens |
| HyCal-17 and 18 | InCal-17 and 18 | SbBAP-17 | SbDiv-17 | InCalTc-18 | SbDivTc-18 | SNitTs | |
|---|---|---|---|---|---|---|---|
| Diversity | Very High | Low | Very High | Low | High | High | High |
| Number of samples | Low | Low | High | High | Low | High | Low |
| Test-training set dissimilarity (range of biomass) | High | Low | Very High | Low | High | High | Low |
| Prediction accuracy (maximum R2) | High (0.80) | Medium (0.71) | Medium (0.67) | Medium (0.74) | Medium (0.69) | High (0.77) | Very high (0.88) |
| Factor | Sum of Squares | Degree of Freedom | F Value | p-Value | ɳ2 (%) |
|---|---|---|---|---|---|
| Data source | 103.70 | 5 | 564.09 | <2 × 10−14 | 68.59 |
| Method | 17.15 | 2 | 233.23 | <2 × 10−14 | 28.36 |
| Data source:method | 9.24 | 10 | 25.14 | <2 × 10−14 | 3.06 |
| Residuals | 594.95 | 16,182 |
| Factor | Sum of Squares | Degree of Freedom | F Value | p-Value | ɳ2 (%) |
|---|---|---|---|---|---|
| Data source | 80.95 | 5 | 412.58 | <2 × 10−6 | 44.18 |
| Method | 19.29 | 2 | 245.79 | <2 × 10−6 | 26.32 |
| Cultivar type | 0.35 | 1 | 8.86 | 0.003 | 0.95 |
| Data source:method | 6.84 | 10 | 17.44 | <2 × 10−6 | 1.87 |
| Data source:cultivar type | 4.03 | 5 | 20.53 | <2 × 10−6 | 2.20 |
| Method:cultivar type | 17.59 | 2 | 224.12 | <2 × 10−6 | 24.00 |
| Data source:method:cultivar type | 1.82 | 10 | 4.63 | 0.093 | 0.50 |
| Residuals | 422.37 | 10,764 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masjedi, A.; Crawford, M.M.; Carpenter, N.R.; Tuinstra, M.R. Multi-Temporal Predictive Modelling of Sorghum Biomass Using UAV-Based Hyperspectral and LiDAR Data. Remote Sens. 2020, 12, 3587. https://doi.org/10.3390/rs12213587
Masjedi A, Crawford MM, Carpenter NR, Tuinstra MR. Multi-Temporal Predictive Modelling of Sorghum Biomass Using UAV-Based Hyperspectral and LiDAR Data. Remote Sensing. 2020; 12(21):3587. https://doi.org/10.3390/rs12213587
Chicago/Turabian StyleMasjedi, Ali, Melba M. Crawford, Neal R. Carpenter, and Mitchell R. Tuinstra. 2020. "Multi-Temporal Predictive Modelling of Sorghum Biomass Using UAV-Based Hyperspectral and LiDAR Data" Remote Sensing 12, no. 21: 3587. https://doi.org/10.3390/rs12213587
APA StyleMasjedi, A., Crawford, M. M., Carpenter, N. R., & Tuinstra, M. R. (2020). Multi-Temporal Predictive Modelling of Sorghum Biomass Using UAV-Based Hyperspectral and LiDAR Data. Remote Sensing, 12(21), 3587. https://doi.org/10.3390/rs12213587