Assessing Typhoon-Induced Canopy Damage Using Vegetation Indices in the Fushan Experimental Forest, Taiwan



Abstract
1. Introduction
2. Materials and Methods
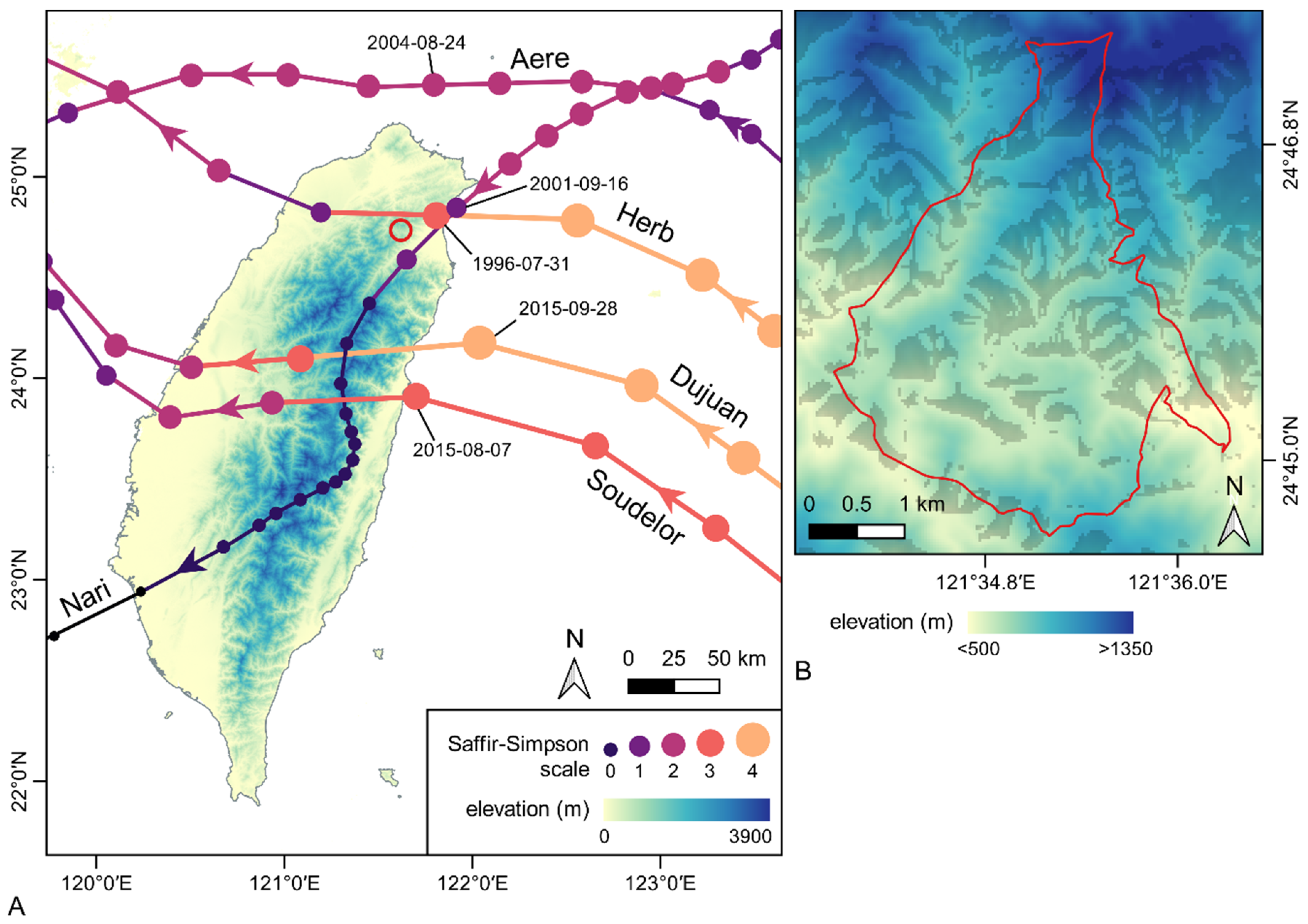
2.1. The Fushan Experimental Forest and Typhoons
2.2. Satellite Images
2.3. Pre-Processing
2.4. Processing
2.5. Analysis of Disturbances among Vegetation Indices
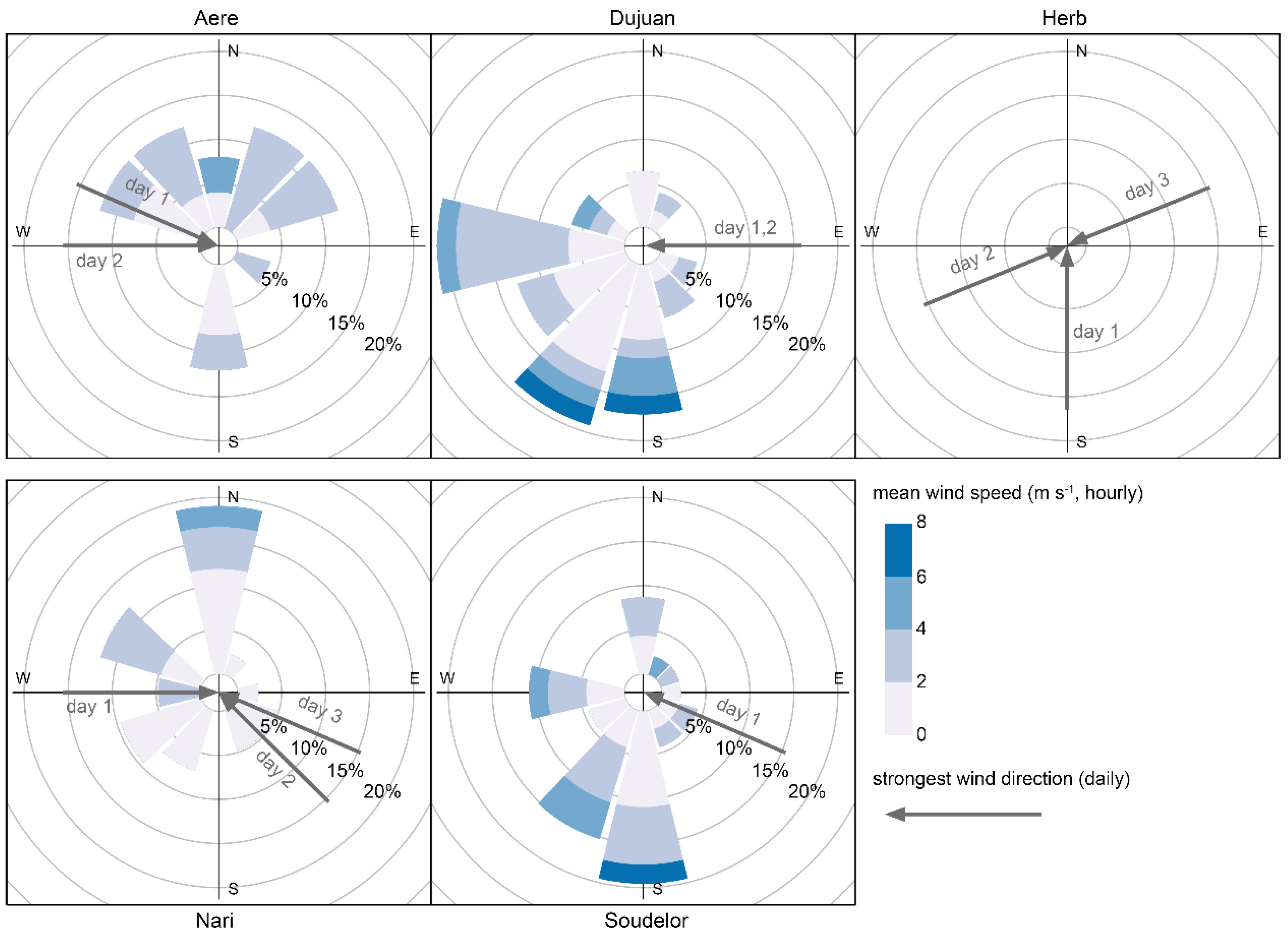
2.6. Typhoon Damages and Topography
2.7. Disturbance Frequencies and Intensity
2.8. Vegetation Recovery
3. Results
3.1. Vegetation Indices, Typhoons, and the Effect of Prior Vegetation Cover
3.2. Variation of ΔVIs among Typhoons
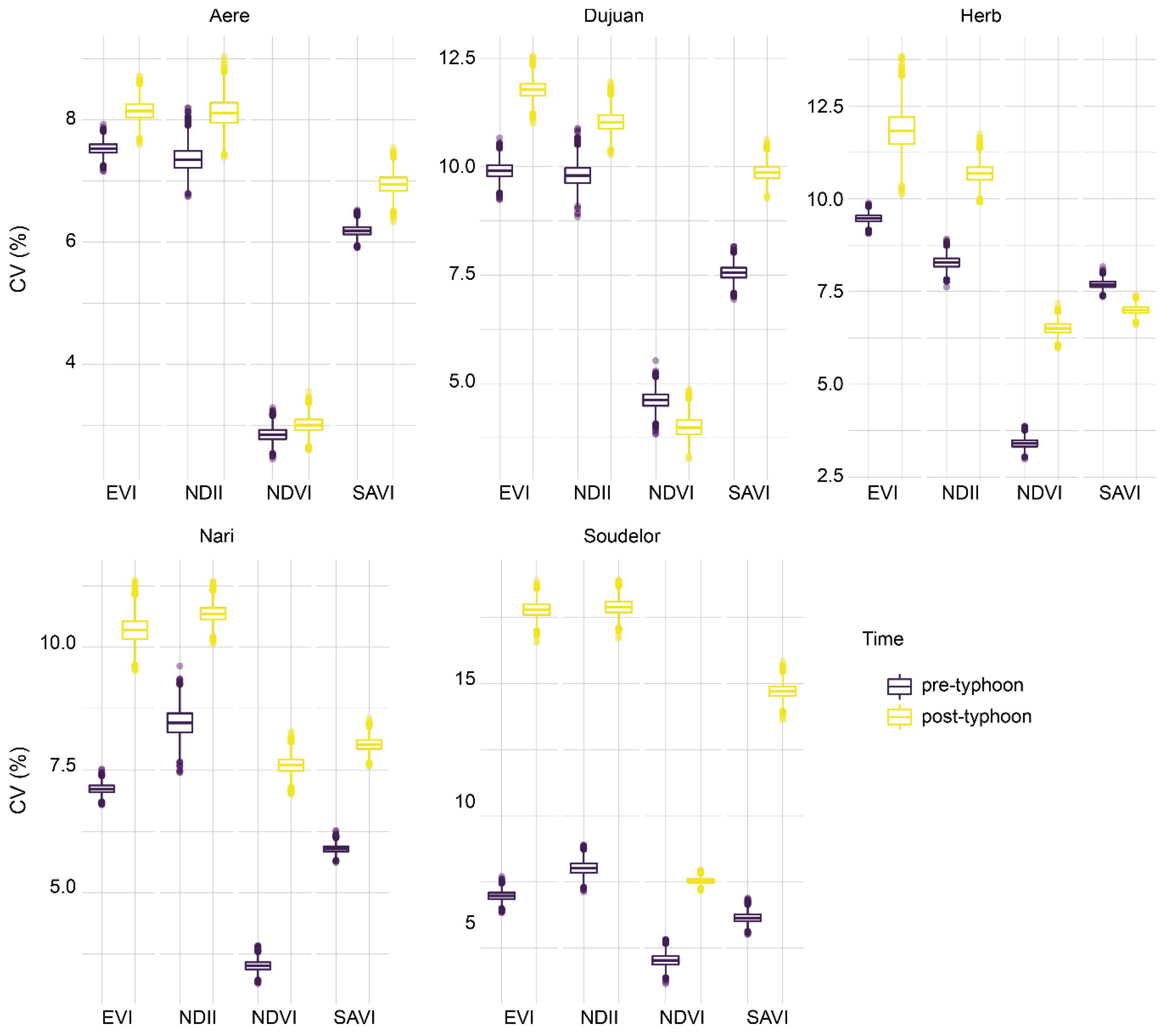
3.3. Effects on Vegetation Heterogeneity
3.4. Topography and Disturbance Severity
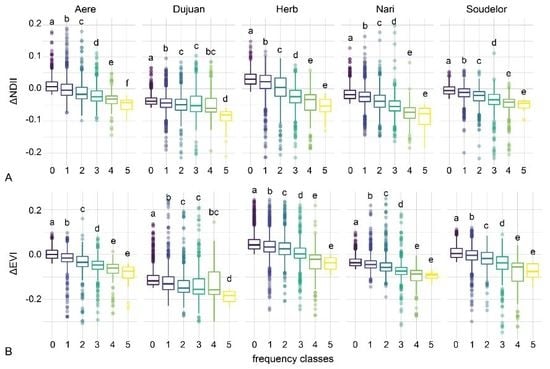
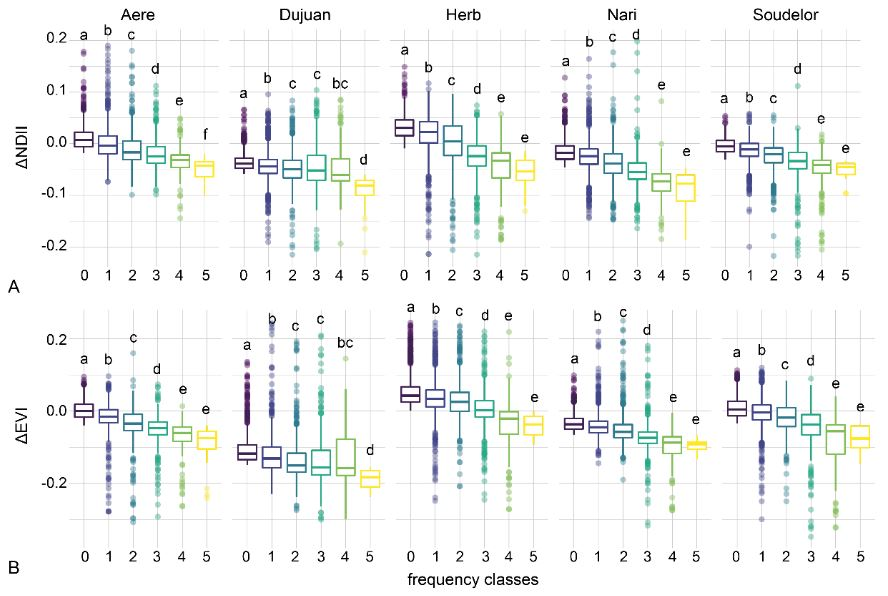
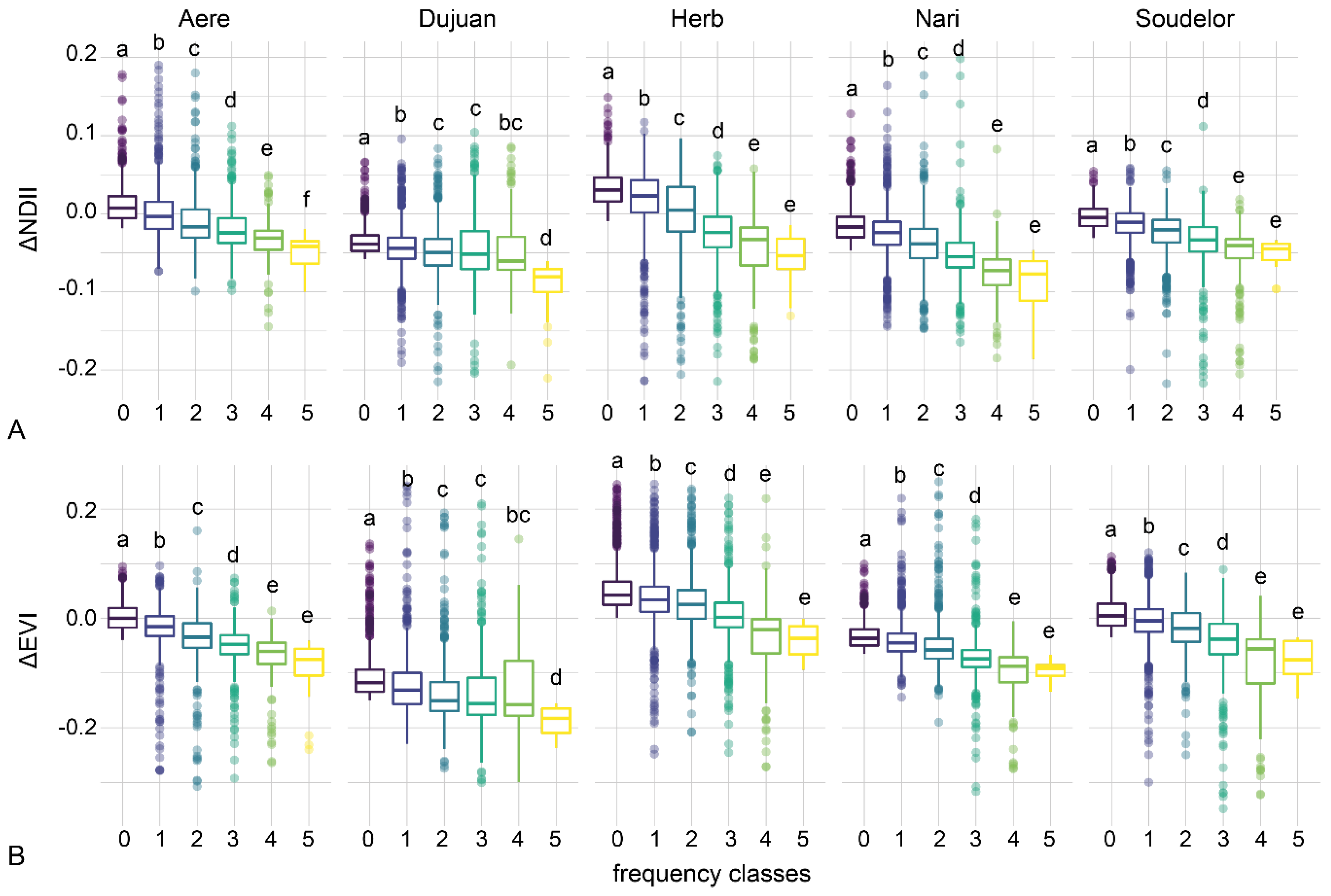
3.5. Disturbance Frequency and Severity
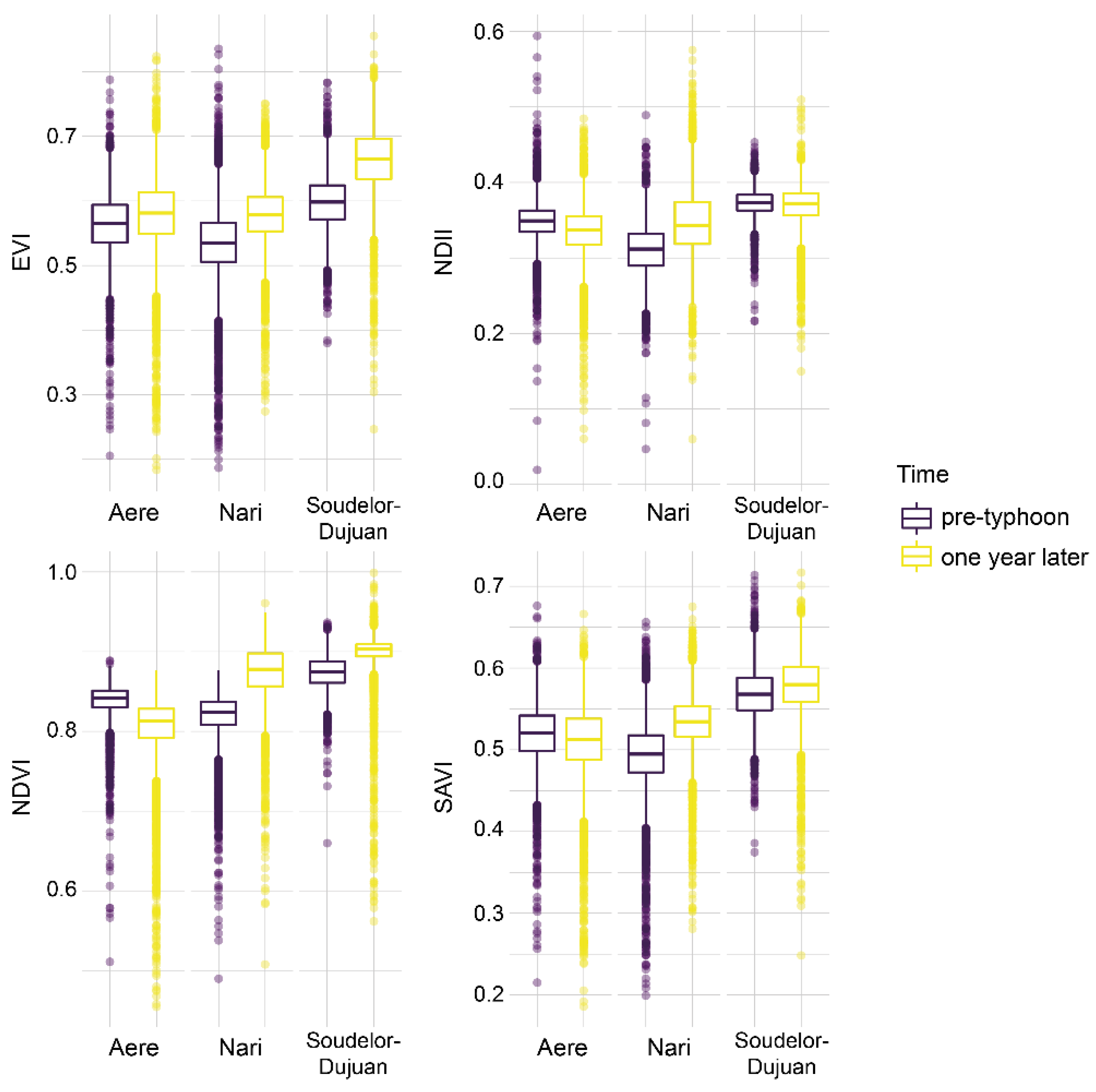
3.6. Recovery
4. Discussion
4.1. Consistency in the Damage Effects among Typhoons
4.2. ΔVIs in Relation to Topography
4.3. Typhoon Disturbance Frequency
4.4. Consistency among Vegetation Indices
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gentry, A.H. Tropical forest biodiversity: Distributional patterns and their conservational significance. Oikos 1992, 63, 19–28. [Google Scholar] [CrossRef]
- Malhi, Y.; Grace, J. Tropical forests and atmospheric carbon dioxide. Trends Ecol. Evol. 2000, 15, 332–337. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef]
- Saugier, B.; Roy, J.; Mooney, H.A. Estimations of global terrestrial productivity: Converging toward a single number? In Terrestrial Global Productivity, 1st ed.; Roy, J., Mooney, H., Saugier, B., Eds.; Academic Press: San Diego, CA, USA, 2001; pp. 543–557. [Google Scholar] [CrossRef]
- Putz, F.E.; Blate, G.M.; Redford, K.H.; Fimbel, R.; Robinson, J. Tropical forest management and conservation of biodiversity: An overview. Conserv. Biol. 2001, 15, 7–20. [Google Scholar] [CrossRef]
- Malhi, Y.; Gardner, T.A.; Goldsmith, G.R.; Silman, M.R.; Zelazowski, P. Tropical forests in the Anthropocene. Annu. Rev. Environ. Resour. 2014, 39, 125–159. [Google Scholar] [CrossRef]
- Baccini, A.; Walker, W.; Carvalho, L.; Farina, M.; Sulla-Menashe, D.; Houghton, R.A. Tropical forests are a net carbon source based on aboveground measurements of gain and loss. Science 2017, 358, 230–234. [Google Scholar] [CrossRef]
- Hubau, W.; Lewis, S.L.; Phillips, O.L.; Affum-Baffoe, K.; Beeckman, H.; Cuní-Sanchez, A.; Daniels, A.K.; Ewango, C.E.N.; Fauset, S.; Mukinzi, J.M.; et al. Asynchronous carbon sink saturation in African and Amazonian tropical forests. Nature 2020, 579, 80–87. [Google Scholar] [CrossRef]
- Whitmore, T.C.; Burslem, D.F.R.P. Major disturbances in tropical rainforests. In Dynamics of Tropical Communities; Newbery, D.M., Prins, H.H.T., Brown, N.D., Eds.; Blackwell Science Ltd: Oxford, UK, 1998; pp. 549–565. [Google Scholar]
- McDowell, N.; Allen, C.D.; Anderson-Teixeira, K.; Brando, P.; Brienen, R.; Chambers, J.; Christoffersen, B.; Davies, S.; Doughty, C.; Duque, A.; et al. Drivers and mechanisms of tree mortality in moist tropical forests. New Phytol. 2018, 219, 851–869. [Google Scholar] [CrossRef]
- Lin, T.-C.; Hogan, J.A.; Chang, C.-T. Tropical cyclone ecology: A scale-link perspective. Trends Ecol. Evol. 2020, in press. [Google Scholar] [CrossRef]
- Pruitt, J.N.; Little, A.G.; Majumdar, S.J.; Schoener, T.W.; Fisher, D.N. Call-to-action: A global consortium for tropical cyclone ecology. Trends Ecol. Evol. 2019, 34, 588–590. [Google Scholar] [CrossRef]
- Lugo, A.E. Visible and invisible effects of hurricanes on forest ecosystems: An international review. Austral Ecol. 2008, 33, 368–398. [Google Scholar] [CrossRef]
- Hogan, J.A.; Zimmerman, J.K.; Thompson, J.; Uriarte, M.; Swenson, N.G.; Condit, R.; Hubbell, S.; Johnson, D.J.; Sun, I.F.; Chang-Yang, C.-H.; et al. The frequency of cyclonic wind storms shapes tropical forest dynamism and functional trait dispersion. Forests 2018, 9, 404. [Google Scholar] [CrossRef]
- Harrington, R.A.; Fownes, J.H.; Scowcroft, P.G.; Vann, C.S. Impact of Hurricane Iniki on native Hawaiian Acacia koa forests: Damage and two-year recovery. J. Trop. Ecol. 1997, 13, 539–558. [Google Scholar] [CrossRef]
- Herbert, D.A.; Fownes, J.H.; Vitousek, P.M. Hurricane damage to a Hawaiian forest: Nutrient supply rate affects resistance and resilience. Ecology 1999, 80, 908–920. [Google Scholar] [CrossRef]
- Tanner, E.V.J.; Rodriguez-Sanchez, F.; Healey, J.R.; Holdaway, R.J.; Bellingham, P.J. Long-term hurricane damage effects on tropical forest tree growth and mortality. Ecology 2014, 95, 2974–2983. [Google Scholar] [CrossRef]
- Webb, E.L.; van de Bult, M.; Fa’aumu, S.; Webb, R.C.; Tualaulelei, A.; Carrasco, L.R. Factors affecting tropical tree damage and survival after catastrophic wind disturbance. Biotropica 2014, 46, 32–41. [Google Scholar] [CrossRef]
- Gannon, B.M.; Martin, P.H. Reconstructing hurricane disturbance in a tropical montane forest landscape in the Cordillera Central, Dominican Republic: Implications for vegetation patterns and dynamics. Arct. Antarct. Alp. Res. 2014, 46, 767–776. [Google Scholar] [CrossRef]
- Burslem, D.F.R.P.; Whitmore, T.C.; Brown, G.C. Short-term effects of cyclone impact and long-term recovery of tropical rain forest on Kolombangara, Solomon Islands. J. Ecol. 2000, 88, 1063–1078. [Google Scholar] [CrossRef]
- Elmqvist, T.; Rainey, W.E.; Pierson, E.D.; Cox, P.A. Effects of Tropical Cyclones Ofa and Val on the structure of a Samoan lowland rain forest. Biotropica 1994, 26, 384–391. [Google Scholar] [CrossRef]
- Hu, T.; Smith, R.B. The impact of Hurricane Maria on the vegetation of Dominica and Puerto Rico using multispectral remote sensing. Remote Sens. 2018, 10, 827. [Google Scholar] [CrossRef]
- Ayala-Silva, T.; Twumasi, Y.A. Hurricane Georges and vegetation change in Puerto Rico using AVHRR satellite data. Int. J. Remote Sens. 2004, 25, 1629–1640. [Google Scholar] [CrossRef]
- Turton, S.M. Landscape-scale impacts of Cyclone Larry on the forests of northeast Australia, including comparisons with previous cyclones impacting the region between 1858 and 2006. Austral Ecol. 2008, 33, 409–416. [Google Scholar] [CrossRef]
- Metcalfe, D.J.; Bradford, M.G.; Ford, A.J. Cyclone damage to tropical rain forests: Species- and community-level impacts. Austral Ecol. 2008, 33, 432–441. [Google Scholar] [CrossRef]
- Boose, E.R.; Foster, D.R.; Fluet, M. Hurricane impacts to tropical and temperate forest landscapes. Ecol. Monogr. 1994, 64, 369–400. [Google Scholar] [CrossRef]
- McLaren, K.; Denneko, L.; Tanner, E.; Bellingham, P.J.; Healey, J.R. Reconstructing the effects of hurricanes over 155 years on the structure and diversity of trees in two tropical montane rainforests in Jamaica. Agric. Forest Meteorol. 2019, 276-277, 107621. [Google Scholar] [CrossRef]
- Bellingham, P.J. Landforms influence patterns of hurricane damage: Evidence from Jamaican montane forests. Biotropica 1991, 23, 427–433. [Google Scholar] [CrossRef]
- Webb, E.L.; Seamon, J.O.; Fa’aumu, S. Frequent, low-amplitude disturbances drive high tree turnover rates on a remote, cyclone-prone Polynesian island. J. Biogeogr. 2011, 38, 1240–1252. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Y.; Jiang, H.; Wang, X. Remote-sensing assessment of forest damage by Typhoon Saomai and its related factors at landscape scale. Int. J. Remote Sens. 2013, 34, 7874–7886. [Google Scholar] [CrossRef]
- Negrón-Juárez, R.; Baker, D.B.; Chambers, J.Q.; Hurtt, G.C.; Goosem, S. Multi-scale sensitivty of Landsat and MODIS to forest disturbance associated with tropical cyclones. Remote Sens. Environ. 2014, 140, 679–689. [Google Scholar] [CrossRef]
- Luke, D.; McLaren, K.; Wilson, B. Modeling hurricane exposure in a Caribbean lower montane tropical wet forest: The effects of frequent, intermediate disturbances and topography on forest structural dynamics and composition. Ecosystems 2016, 19, 1178–1195. [Google Scholar] [CrossRef]
- Inagaki, Y.; Kuramoto, S.; Torii, A.; Shinomiya, Y.; Fukata, H. Effects of thinning on leaf-fall and leaf-litter nitrogen concentration in hinoki cypress (Chamaecyparis obtusa Endlicher) plantation stands in Japan. For. Ecol. Manag. 2008, 255, 1859–1867. [Google Scholar] [CrossRef]
- Brokaw, N.V.L.; Grear, J.S. Forest structure before and after Hurricane Hugo at three elevations in the Luquillo Mountains, Puerto Rico. Biotropica 1991, 23, 386–392. [Google Scholar] [CrossRef]
- Boose, E.R.; Serrano, M.I.; Foster, D.R. Landscape and regional impacts of hurricanes in Puerto Rico. Ecol. Monogr. 2004, 74, 335–352. [Google Scholar] [CrossRef]
- Heartsill Scalley, T.; Scatena, F.N.; Lugo, A.E.; Moya, S.; Estrada Ruiz, C.R. Changes in structure, composition, and nutrients during 15 years of hurricane-induced succession in a subtropical wet forest in Puerto Rico. Biotropica 2010, 42, 455–463. [Google Scholar] [CrossRef]
- Uriarte, M.; Thompson, J.; Zimmerman, J.K. Hurricane María tripled stem breaks and doubled tree mortality relative to other major storms. Nat. Commun. 2019, 10, 1362. [Google Scholar] [CrossRef] [PubMed]
- Hogan, J.A.; Zimmerman, J.K.; Thompson, J.; Nytch, C.J.; Uriarte, M. The interaction of land-use legacies and hurricane disturbance in subtropical wet forest: Twenty-one years of change. Ecosphere 2016, 7, e01405. [Google Scholar] [CrossRef]
- Hall, J.; Muscarella, R.; Quebbeman, A.; Arellano, G.; Thompson, J.; Zimmerman, J.K.; Uriarte, M. Hurricane-induced rainfall is a stronger predictor of tropical forest damage in Puerto Rico than maximum wind speeds. Sci. Rep. 2020, 10, 4318. [Google Scholar] [CrossRef]
- Comita, L.S.; Uriarte, M.; Forero-Montaña, J.; Kress, W.J.; Swenson, N.G.; Thompson, J.; Umaña, M.N.; Zimmerman, J.K. Changes in phylogenetic community structure of the seedling layer following hurricane disturbance in an human-impacted tropical forest. Forests 2018, 9, 556. [Google Scholar] [CrossRef]
- Mabry, C.M.; Hamburg, S.P.; Lin, T.-C.; Horng, F.-W.; King, H.-B.; Hsia, Y.-J. Typhoon disturbance and stand-level damage patterns at a subtropical forest in Taiwan. Biotropica 1998, 30, 238–250. [Google Scholar] [CrossRef]
- Lee, M.-F.; Lin, T.-C.; Vadeboncoeur, M.A.; Hwong, J.-L. Remote sensing assessment of forest damage in relation to the 1996 strong Typhoon Herb at Lienhuachi Experimental Forest, Taiwan. For. Ecol. Manag. 2008, 255, 3297–3306. [Google Scholar] [CrossRef]
- De Beurs, K.M.; McThompson, N.S.; Owsley, B.C.; Henebry, G.M. Hurricane damage detection on four major Caribbean islands. Remote Sens. Environ. 2019, 229, 1–13. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Knapp, D.E.; Tupayachi, R.; Anderson, C.B.; Sinca, F.; Vaughn, N.R.; Llactayo, W. Airborne laser-guided imaging spectroscopy to map forest trait diversity and guide conservation. Science 2017, 355, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Asner, G.P.; Brodrick, P.G.; Philipson, C.; Vaughn, N.R.; Martin, R.E.; Knapp, D.E.; Heckler, J.; Evans, L.J.; Jucker, T.; Goossens, B.; et al. Mapped aboveground carbon stocks to advance forest conservation and recovery in Malaysian Borneo. Biol. Conserv. 2018, 217, 289–310. [Google Scholar] [CrossRef]
- Abbas, S.; Nichol, J.E.; Fischer, G.A.; Wong, M.S.; Irteza, S.M. Impact assessment of a super-typhoon on Hong Kong’s secondary vegetation and recommendations for restoration of resilience in the forest succession. Agric. For. Meteorol. 2020, 280. [Google Scholar] [CrossRef]
- Huete, A.R. Vegetation indices, remote sensing and forest monitoring. Geogr. Compass 2012, 6, 513–532. [Google Scholar] [CrossRef]
- Glenn, E.P.; Huete, A.R.; Nagler, P.L.; Nelson, S.G. Relationship between remotely-sensed vegetation indices, canopy attributes and plant physiological processes: What vegetation indices can and cannot tell us about the landscape. Sensors (Basel) 2008, 8, 2136–2160. [Google Scholar] [CrossRef]
- Kerr, J.T.; Ostrovsky, M. From space to species: Ecological applications for remote sensing. Trends Ecol. Evol. 2003, 18, 299–305. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef]
- Baret, F.; Guyot, G. Potentials and limits of vegetation indices for LAI and APAR assessment. Remote Sens. Environ. 1991, 35, 161–173. [Google Scholar] [CrossRef]
- Lobell, D.B.; Asner, G.P.; Law, B.E.; Treuhaft, R.N. Subpixel canopy cover estimation of coniferous forests in Oregon using SWIR imaging spectrometry. J. Geophys. Res. Atmos. 2001, 106, 5151–5160. [Google Scholar] [CrossRef]
- Tian, Y.; Zhu, Y.; Cao, W. Monitoring leaf photosynthesis with canopy spectral reflectance in rice. Photosynthetica 2005, 43, 481–489. [Google Scholar] [CrossRef]
- Huete, A.R.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Huete, A.R.; Didan, K.; Shimabukuro, Y.E.; Ratana, P.; Saleska, S.R.; Hutyra, L.R.; Yang, W.; Nemani, R.R.; Myneni, R. Amazon rainforests green-up with sunlight in dry season. Geophys. Res. Lett. 2006, 33, L06405. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.; van Leeuwen, W.J. The use of vegetation indices in forested regions: Issues of linearity and saturation. In Proceedings of the IGARSS’97. 1997 IEEE International Geoscience and Remote Sensing Symposium Proceedings. Remote Sensing—A Scientific Vision for Sustainable Development, Singapore, 3–8 August 1997; pp. 1966–1968. [Google Scholar]
- White, J.D.; Running, S.W.; Nemani, R.; Keane, R.E.; Ryan, K.C. Measurement and remote sensing of LAI in Rocky Mountain montane ecosystems. Can. J. For. Res. 1997, 27, 1714–1727. [Google Scholar] [CrossRef]
- Gao, X.; Huete, A.R.; Ni, W.; Miura, T. Optical-biophysical relationships of vegetation spectra without background contamination. Remote Sens. Environ. 2000, 74, 609–620. [Google Scholar] [CrossRef]
- Hardisky, M.A.; Klemas, V.; Smart, R.M. The influences of soil salinity, growth form, and leaf moisture on the spectral reflectance of Spartina alterniflora canopies. Photogramm. Eng. Remote Sens. 1983, 49, 77–83. [Google Scholar]
- Cheng, Y.-B.; Zarco-Tejada, P.J.; Riaño, D.; Rueda, C.A.; Ustin, S.L. Estimating vegetation water content with hyperspectral data for different canopy scenarios: Relationships between AVIRIS and MODIS indexes. Remote Sens. Environ. 2006, 105, 354–366. [Google Scholar] [CrossRef]
- Anderson, M.C.; Neale, C.M.U.; Li, F.; Norman, J.M.; Kustas, W.P.; Jayanthi, H.; Chavez, J. Upscaling ground observations of vegetation water content, canopy height, and leaf area index during SMEX02 using aircraft and Landsat imagery. Remote Sens. Environ. 2004, 92, 447–464. [Google Scholar] [CrossRef]
- Lawrence, R.L.; Ripple, W.J. Comparisons among vegetation indices and bandwise regression in a highly disturbed, heterogeneous landscape: Mount St. Helens, Washington. Remote Sens. Environ. 1998, 64, 91–102. [Google Scholar] [CrossRef]
- Lin, T.-C.; Hamburg, S.P.; Lin, K.-C.; Wang, L.-J.; Chang, C.-T.; Hsia, Y.-J.; Vadeboncoeur, M.A.; Mabry McMullen, C.M.; Liu, C.-P. Typhoon disturbance and forest dynamics: Lessons from a northwest Pacific subtropical forest. Ecosystems 2011, 14, 127–143. [Google Scholar] [CrossRef]
- Chi, C.-H.; McEwan, R.W.; Chang, C.-T.; Zheng, C.; Yang, Z.; Chiang, J.-M.; Lin, T.-C. Typhoon disturbance mediates elevational patterns of forest structure, but not species diversity, in humid monsoon Asia. Ecosystems 2015, 18, 1410–1423. [Google Scholar] [CrossRef]
- Peereman, J.; Hogan, J.A.; Lin, T.-C. Landscape representation by a permanent forest plot and alternative plot designs in a typhoon hotspot, Fushan, Taiwan. Remote Sens. 2020, 12, 660. [Google Scholar] [CrossRef]
- Lin, S.-Y.; Shaner, P.-J.L.; Lin, T.-C. Characteristics of old-growth and secondary forests in relation to age and typhoon disturbance. Ecosystems 2018, 21, 1521–1532. [Google Scholar] [CrossRef]
- Su, S.-H.; Chang-Yang, C.H.; Lu, C.-L.; Tsui, C.-C.; Lin, T.-T.; Lin, C.-L.; Chiou, W.-L.; Kuan, L.-H.; Chen, Z.-S.; Hsieh, C.-F. Fushan Subtropical Forest Dynamics Plot: Tree Species Characteristics and Distribution Patterns; Taiwan Forestry Research Institute: Taipei, Taiwan, 2007. [Google Scholar]
- Wang, H.-H.; Pan, F.-J.; Liu, C.-K.; Yu, Y.-H.; Hung, S.-F. Vegetation classification and ordination of a permanent plot in the Fushan Experimental Forest, northern Taiwan. Taiwan J. For. Sci. 2000, 15, 411–428. [Google Scholar] [CrossRef]
- Knapp, K.R.; Kruk, M.C.; Levinson, D.H.; Diamond, H.J.; Neumann, C.J. The International Best Track Archive for Climate Stewardship (IBTrACS): Unifying tropical cyclone best track data. Bull. Am. Meteorol. Soc. 2010, 91, 363–376. [Google Scholar] [CrossRef]
- Knapp, K.R.; Diamond, H.J.; Kossin, J.P.; Kruk, M.C.; Schreck, C.J. International Best Track Archive for Climate Stewardship (IBTrACS) Project, Version 4. [IBTrACS.EP.list.v04r00]; NOAA National Centers for Environmental Information: Asheville, NC, USA, 2018. [CrossRef]
- Walker, L.R.; Voltzow, J.; Ackerman, J.D.; Fernández, D.S.; Fetcher, N. Immediate impact of Hurricane Hugo on a Puerto Rican rain forest. Ecology 1992, 73, 691–694. [Google Scholar] [CrossRef]
- Lin, K.-C.; Hamburg, S.P.; Wang, L.; Duh, C.-T.; Huang, C.-M.; Chang, C.-T.; Lin, T.-C. Impacts of increasing typhoons on the structure and function of a subtropical forest: Reflections of a changing climate. Sci. Rep. 2017, 7, 4911. [Google Scholar] [CrossRef]
- Chang, C.-T.; Wang, L.-J.; Huang, J.-C.; Liu, C.-P.; Wang, C.-P.; Lin, N.-H.; Wang, L.; Lin, T.-C. Precipitation controls on nutrient budgets in subtropical and tropical forests and the implications under changing climate. Adv. Water Resour. 2017, 103, 44–50. [Google Scholar] [CrossRef]
- Simpson, R.H.; Riehl, H. The Hurricane and Its Impact; Louisiana State University Press: Baton Rouge, LA, USA, 1981; p. 398. [Google Scholar]
- Carslaw, D.C.; Ropkins, K. openair - An R package for air quality data analysis. Environ. Model. Softw. 2012, 27–28, 52–61. [Google Scholar] [CrossRef]
- Chang-Yang, C.H.; Lu, C.-L.; Sun, I.-F.; Hsieh, C.-F. Flowering and fruiting patterns in a subtropical rain forest, Taiwan. Biotropica 2013, 45, 165–174. [Google Scholar] [CrossRef]
- Lin, K.-C.; Hwanwu, C.-B.; Liu, C.-C. Phenology of broadleaf tree species in the Fushan Experimental Forest of northeastern Taiwan. Taiwan J. For. Sci. 1997, 12, 347–355. [Google Scholar] [CrossRef]
- USGS. Earthexplorer. Available online: https://earthexplorer.usgs.gov/ (accessed on 4 July 2019).
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-resolution global maps of 21st-century forest cover change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Young, N.E.; Anderson, R.S.; Chignell, S.M.; Vorster, A.G.; Lawrence, R.; Evangelista, P.H. A survival guide to Landsat preprocessing. Ecology 2017, 98, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Leutner, B.; Horning, N.; Schwalb-Willmann, J. RStoolbox: Tools for Remote Sensing Data Analysis; 2019; Available online: https://cran.r-project.org/web/packages/RStoolbox/index.html (accessed on 2 May 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Foga, S.; Scaramuzza, P.L.; Guo, S.; Zhu, Z.; Dilley, R.D., Jr.; Beckmann, T.; Schmidt, G.L.; Dwyer, J.L.; Hughes, M.J.; Laue, B. Cloud detection algorithm comparison and validation for operational Landsat data products. Remote Sens. Environ. 2017, 194, 379–390. [Google Scholar] [CrossRef]
- Achard, F.; Beuchle, R.; Mayaux, P.; Stibig, H.J.; Bodart, C.; Brink, A.; Carboni, S.; Desclée, B.; Donnay, F.; Eva, H.D.; et al. Determination of tropical deforestation rates and related carbon losses from 1990 to 2010. Glob. Chang. Biol. 2014, 20, 2540–2554. [Google Scholar] [CrossRef]
- Vieilledent, G.; Grinand, C.; Rakotomalala, F.A.; Ranaivosoa, R.; Rakotoarijaona, J.-R.; Allnutt, T.F.; Achard, F. Combining global tree cover loss data with historical national forest cover maps to look at six decades of deforestation and forest fragmentation in Madagascar. Biol. Conserv. 2018, 222, 189–197. [Google Scholar] [CrossRef]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling; 2.9-23; 2019; Available online: https://rdrr.io/cran/raster/ (accessed on 2 May 2020).
- Weiss, A. Topographic Position and Landform Analysis. In Poster Presentation; ESRI User Conference: San Diego, CA, USA, 2001. [Google Scholar]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W.; Harlan, J.C. Monitoring the Vernal Advancement of Retrogradation (Green Wave Effect) of Natural Vegetation; Type III Final Rep; NASA/GSFC: Greenbelt, MD, USA, 1974; p. 371.
- Huete, A.R. A Soil-Adjusted Vegetation Index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Chang, C.-T.; Wang, S.-F.; Vadeboncoeur, M.A.; Lin, T.-C. Relating vegetation dynamics to temperature and precipitation at monthly and annual timescales in Taiwan using MODIS vegetation indices. Int. J. Remote Sens. 2014, 35, 598–620. [Google Scholar] [CrossRef]
- Townsend, P.A.; Singh, A.; Foster, J.R.; Rehberg, N.J.; Kingdon, C.C.; Eshleman, K.N.; Seagle, S.W. A general Landsat model to predict canopy defoliation in broadleaf deciduous forests. Remote Sens. Environ. 2012, 119, 255–265. [Google Scholar] [CrossRef]
- Wang, W.; Qu, J.J.; Hao, X.; Liu, Y.; Stanturf, J.A. Post-hurricane forest damage assessment using satellite remote sensing. Agric. For. Meteorol. 2010, 150, 122–132. [Google Scholar] [CrossRef]
- Zhang, K.; Thapa, B.; Ross, M.; Gann, D. Remote sensing of seasonal changes and disturbances in mangrove forest: A case study from South Florida. Ecosphere 2016, 7, e01366. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q.; Batchily, K.; van Leeuwen, W. A comparison of vegetation indices over a global set of TM images for EOS-MODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; 1.9.12; 2019; Available online: https://www.scholars.northwestern.edu/en/publications/psych-procedures-for-personality-and-psychological-research (accessed on 2 May 2020).
- Fisher, J.I.; Hurtt, G.C.; Thomas, R.Q.; Chambers, J.Q. Clustered disturbances lead to bias in large-scale estimates based on forest sample plots. Ecol. Lett. 2008, 11, 554–563. [Google Scholar] [CrossRef]
- Saatchi, S.; Mascaro, J.; Xu, L.; Keller, M.; Yang, Y.; Duffy, P.; Espírito-Santo, F.; Baccini, A.; Chambers, J.; Schimel, D. Seeing the forest beyond the trees. Glob. Ecol. Biogeogr. 2015, 24, 606–610. [Google Scholar] [CrossRef]
- Turner, D.P.; Cohen, W.B.; Kennedy, R.E.; Fassnacht, K.S.; Briggs, J.M. Relationships between Leaf Area Index and Landsat TM spectral vegetation indices across three temperate zone sites. Remote Sens. Environ. 1999, 70, 52–68. [Google Scholar] [CrossRef]
- Herberich, E.; Sikorski, J.; Hothorn, T. A robust procedure for comparing multiple means under heteroscedasticity in unbalanced designs. PLoS ONE 2010, 5, e9788. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Zeileis, A. Econometric computing with HC and HAC covariance matrix estimators. J. Stat. Softw. 2004, 11, 1–17. [Google Scholar] [CrossRef]
- Zeileis, A. Object-oriented computation of sandwich estimators. J. Stat. Softw. 2006, 16, 1–16. [Google Scholar] [CrossRef]
- Rossi, E.; Rogan, J.; Schneider, L. Mapping forest damage in northern Nicaragua after Hurricane Felix (2007) using MODIS enhanced vegetation index data. GIsci. Remote Sens. 2013, 50, 385–399. [Google Scholar] [CrossRef]
- Tsai, H.P.; Yang, M.-D. Relating vegetation dynamics to climate variables in Taiwan using 1982-2012 NDVI3g data. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2016, 9, 1624–1639. [Google Scholar] [CrossRef]
- Kang, R.-L.; Lin, T.-C.; Jan, J.-F.; Hwong, J.-L. Changes in the Normalized Difference Vegetation Index (NDVI) at the Fushan Experimental Forest in relation to Typhoon Bilis of 2000. Taiwan J. For. Sci. 2005, 20, 73–87. [Google Scholar] [CrossRef]
- Liu, X.; Zeng, X.; Zou, X.; González, G.; Wang, C.; Yang, S. Litterfall production prior to and during Hurricanes Irma and Maria in four Puerto Rican forests. Forests 2018, 9, 367. [Google Scholar] [CrossRef]
- Walker, L.R. Tree damage and recovery from Hurricane Hugo in Luquillo experimental forest, Puerto Rico. Biotropica 1991, 23, 379–385. [Google Scholar] [CrossRef]
- Lodge, D.J.; Scatena, F.N.; Asbury, C.E.; Sanchez, M.J. Fine litterfall and related nutrient inputs resulting from Hurricane Hugo in subtropical wet and lower montane rain forests of Puerto Rico. Biotropica 1991, 23, 336–342. [Google Scholar] [CrossRef]
- Lin, T.-C.; Hamburg, S.P.; Hsia, Y.-J.; Lin, T.-T.; King, H.-B.; Wang, L.-J.; Lin, K.-C. Influence of typhoon disturbances on the understory light regime and stand dynamics of a subtropical rain forest in northeastern Taiwan. J. For. Res. 2003, 8, 139–145. [Google Scholar] [CrossRef]
- Grimbacher, P.S.; Catterall, C.P.; Stork, N.E. Do edge effects increase the susceptibility of rainforest fragments to structural damage resulting from a severe tropical cyclone? Austral Ecol. 2008, 33, 525–531. [Google Scholar] [CrossRef]
- Everham, E.M.; Brokaw, N.V.L. Forest damage and recovery from catastrophic wind. Bot. Rev. 1996, 62, 113–185. [Google Scholar] [CrossRef]
- Lin, M.L.; Jeng, F.S. Characteristics of hazards induced by extremely heavy rainfall in Central Taiwan—Typhoon Herb. Eng. Geol. 2000, 58, 191–207. [Google Scholar] [CrossRef]
- Knutson, T.R.; McBride, J.L.; Chan, J.; Emanuel, K.; Holland, G.; Landsea, C.; Held, I.; Kossin, J.P.; Srivastava, A.K.; Sugi, M. Tropical cyclones and climate change. Nat. Geosci. 2010, 3, 157–163. [Google Scholar] [CrossRef]
- Walsh, K.J.E.; McBride, J.L.; Klotzbach, P.J.; Balachandran, S.; Camargo, S.J.; Holland, G.; Knutson, T.R.; Kossin, J.; Lee, T.-c.; Sobel, A.; et al. Tropical cyclones and climate change. Wiley Interdiscip. Rev. Clim. Chang. 2016, 7, 65–89. [Google Scholar] [CrossRef]
- Knutson, T.R.; Sirutis, J.J.; Zhao, M.; Tuleya, R.E.; Bender, M.; Vecchi, G.A.; Villarini, G.; Chavas, D. Global projections of intense tropical cyclone activity for the late twenty-first century from dynamical downscaling of CMIP5/RCP4.5 scenarios. J. Clim. 2015, 28, 7203–7224. [Google Scholar] [CrossRef]
- McEwan, R.W.; Lin, Y.-C.; Sun, I.-F.; Hsieh, C.-F.; Su, S.-H.; Chang, L.-W.; Song, G.-Z.M.; Wang, H.-H.; Hwong, J.-L.; Lin, K.-C.; et al. Topographic and biotic regulation of aboveground carbon storage in subtropical broad-leaved forests of Taiwan. For. Ecol. Manag. 2011, 262, 1817–1825. [Google Scholar] [CrossRef]
- Ostertag, R.; Silver, W.L.; Lugo, A.E. Factors affecting mortality and resistance to damage following hurricanes in a rehabilitated subtropical moist forest. Biotropica 2005, 37, 16–24. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; van Leeuwen, W.; Miura, T.; Glenn, E. MODIS Vegetation Indices. In Land Remote Sensing and Global Environmental Change: NASA’s Earth Observing System and the Science of ASTER and MODIS; Ramachandran, B., Justice, C.O., Abrams, M.J., Eds.; Springer: New York, NY, USA, 2011; pp. 579–602. [Google Scholar] [CrossRef]
- Irons, J.R.; Dwyer, J.L.; Barsi, J.A. The next Landsat satellite: The Landsat data continuity mission. Remote Sens. Environ. 2012, 122, 11–21. [Google Scholar] [CrossRef]
- Markham, B.L.; Helder, D.L. Forty-year calibrated record of earth-reflected radiance from Landsat: A review. Remote Sens. Environ. 2012, 122, 30–40. [Google Scholar] [CrossRef]
- Markham, B.; Barsi, J.; Kvaran, G.; Ong, L.; Kaita, E.; Biggar, S.; Czapla-Myers, J.; Mishra, N.; Helder, D. Landsat-8 operational land imager radiometric calibration and stability. Remote Sens. 2014, 6, 12275–12308. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Typhoon | Dates | Total Rainfall (mm) | Max Instantaneous Wind Speed (m s−1) |
|---|---|---|---|
| Herb | 1996-07-30 to 1996-08-01 | 720 | 36.8 |
| Nari | 2001-09-16 to 2001-09-18 | 1300 | 25.1 |
| Aere | 2004-08-24 to 2004-08-25 | 710 | 25.0 |
| Soudelor | 2015-08-07 to 2015-08-08 | 790 | 21.8 |
| Dujuan | 2015-09-28 to 2018-09-29 | 690 | 21.9 |
| Typhoon | Image Acquisition Dates | Sensors | Resolution (m) | Null Cells (%) | |
|---|---|---|---|---|---|
| disturbance | recovery | ||||
| Herb | 1996-07-06 & 08-23 | - | TM5 | 30 | 19.4 |
| Nari | 2001-09-14 & 10-08 | 2001-06-18 & 2002-06-29 | TM5, ETM+ | 30 | 17.5 |
| Aere | 2004-07-12 & 09-30 | 2004-07-12 & 2005-07-15 | TM5 | 30 | 30.8 |
| Soudelor | 2015-06-09 & 08-12 | 2015-06-09 & 2016-07-13 | OLI | 30 | 48.8 |
| Dujuan | 2015-09-13 & 11-16 | OLI | 30 | 33.9 | |
| Slope Position | TPI | Slope (°) |
|---|---|---|
| Ridge | SD < x | – |
| Upper slope | 0.5 SD < x < SD | – |
| Middle slope | −0.5 SD < x < 0.5 SD | >5 |
| Flat slope | −0.5 SD < x < 0.5 SD | <5 |
| Lower slope | −SD < x < −0.5 SD | – |
| Valley | x < −SD | – |
| Vegetation change | ||||
|---|---|---|---|---|
| Typhoon | ΔEVI | ΔNDII | ΔNDVI | ΔSAVI |
| Aere | −0.021 (0.047) | −0.006 (0.034) | −0.012 (0.027) | −0.029 (0.038) |
| variation (%) | −3.39 | −1.18 | −1.39 | −5.34 |
| Dujuan | −0.121 (0.057) | −0.044 (0.031) | −0.012 (0.034) | −0.089 (0.041) |
| variation (%) | −19.87 | −12.09 | −1.19 | −16.23 |
| Herb | 0.048 (0.099) | 0.008 (0.045) | −0.035 (0.067) | 0.014 (0.423) |
| variation (%) | 11.06 | 3.31 | −4.01 | 4.01 |
| Nari | −0.046 (0.050) | −0.034 (0.037) | −0.044 (0.071) | −0.052 (0.040) |
| variation (%) | −8.62 | −9.87 | −5.51 | −12.50 |
| Soudelor | −0.010 (0.045) | −0.017 (0.027) | −0.022 (0.033) | −0.010 (0.037) |
| variation (%) | −1.34 | −4.40 | −2.52 | −1.55 |
| Correlation (ρ) | ||||||
|---|---|---|---|---|---|---|
| Typhoon | ΔEVI-ΔNDVI | ΔEVI-ΔNDII | ΔEVI-ΔSAVI | ΔNDVI-ΔNDII | ΔNDVI-ΔSAVI | ΔNDII-ΔSAVI |
| Aere | 0.59 | 0.23 | 0.90 | 0.28 | 0.64 | 0.31 |
| Dujuan | 0.27 | 0.21 | 0.90 | 0.48 | 0.19 | 0.19 |
| Herb | −0.23 | −0.27 | 0.60 | 0.64 | 0.42 | 0.26 |
| Nari | 0.10 | 0.05 | 0.48 | 0.56 | 0.80 | 0.55 |
| Soudelor | 0.39 | 0.46 | 0.97 | 0.64 | 0.48 | 0.49 |
| Topography | Nari | Herb | ||||||
|---|---|---|---|---|---|---|---|---|
| ΔEVI | ΔNDII | ΔNDVI | ΔSAVI | ΔEVI | ΔNDII | ΔNDVI | ΔSAVI | |
| Elevation (m) | 0.00005 * | 0.00002 ‡ | 0.00008 * | 0.00006 * | 0.0002 * | 0.0001 * | 0.0002 * | 0.0002 * |
| Slope (°) | 0.0006 * | −0.001 * | −0.002 * | −0.0008 * | 0.002 * | −0.0007 * | −0.001 * | 0.0002 * |
| TPI category | ||||||||
| Lower slope | −0.008 | 0.003 | 0.007 | 0.003 | 0.02 † | 0.02 * | 0.02 * | 0.02 * |
| Middle slope | −0.005 | 0.003 | 0.008 | 0.005 | 0.03 * | 0.03 * | 0.02 * | 0.02 * |
| Ridge | −0.0005 | 0.0004 | 0.004 | 0.008 ‡ | 0.03 * | 0.03 * | 0.02 * | 0.02 * |
| Upper slope | −0.002 | 0.002 | 0.005 | 0.006 † | 0.03 * | 0.03 * | 0.02 * | 0.02 * |
| Valley | −0.008 | 0.0007 | 0.003 | 0.0001 | 0.01 | 0.02 * | 0.01 * | 0.01 * |
| Flat slope | - | - | - | - | - | - | - | - |
| Aspects | ||||||||
| North | 0.007 ‡ | −0.03 * | −0.07 * | −0.03 * | 0.03 * | −0.03 * | −0.05 * | −0.005 ‡ |
| Northeast | −0.002 | −0.02 * | −0.03 * | −0.01 * | 0.005 | −0.006 ‡ | −0.01 * | −0.003 |
| Northwest | 0.02 * | −0.03 * | −0.09 * | −0.04 * | 0.05 * | −0.06 * | −0.09 * | −0.02 * |
| South | −0.006 ‡ | −0.0005 | −0.001 | −0.006 * | 0.02 * | −0.03 * | −0.04 * | 0.0004 |
| Southeast | −0.003 | −0.0002 | 0.006 † | −0.003 | 0.006 | −0.007 * | −0.01 * | −0.001 |
| Southwest | −0.006 † | −0.01 * | −0.03 * | −0.02 * | 0.05 * | −0.05 * | −0.07 * | −0.0005 |
| West | 0.009 * | −0.02 * | −0.06 * | −0.03 * | 0.07 * | −0.06 * | −0.1 * | −0.007 * |
| East | - | - | - | - | - | - | - | - |
| R2 | 0.07899 | 0.2163 | 0.4684 | 0.2786 | 0.2222 | 0.3417 | 0.5179 | 0.2046 |
| Adjusted R2 | 0.07618 | 0.2139 | 0.4668 | 0.2764 | 0.2198 | 0.3397 | 0.5164 | 0.2021 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peereman, J.; Hogan, J.A.; Lin, T.-C. Assessing Typhoon-Induced Canopy Damage Using Vegetation Indices in the Fushan Experimental Forest, Taiwan. Remote Sens. 2020, 12, 1654. https://doi.org/10.3390/rs12101654
Peereman J, Hogan JA, Lin T-C. Assessing Typhoon-Induced Canopy Damage Using Vegetation Indices in the Fushan Experimental Forest, Taiwan. Remote Sensing. 2020; 12(10):1654. https://doi.org/10.3390/rs12101654
Chicago/Turabian StylePeereman, Jonathan, James Aaron Hogan, and Teng-Chiu Lin. 2020. "Assessing Typhoon-Induced Canopy Damage Using Vegetation Indices in the Fushan Experimental Forest, Taiwan" Remote Sensing 12, no. 10: 1654. https://doi.org/10.3390/rs12101654
APA StylePeereman, J., Hogan, J. A., & Lin, T.-C. (2020). Assessing Typhoon-Induced Canopy Damage Using Vegetation Indices in the Fushan Experimental Forest, Taiwan. Remote Sensing, 12(10), 1654. https://doi.org/10.3390/rs12101654