Mapping Succession in Non-Forest Habitats by Means of Remote Sensing: Is the Data Acquisition Time Critical for Species Discrimination?

 , ,
, ,  ,
,  , ,
, , 
Abstract

1. Introduction
2. Study Areas
3. Source Data
3.1. Remote Sensing Data
3.2. Field Data
4. Methodology
4.1. Overview
4.2. Hyperspectral Data Processing
4.3. LiDAR Data Processing
4.4. Reference Data Preparation
4.5. Reference Data Analysis
4.6. Feature Selection and Classification
4.7. Masking
4.8. Accuracy Assessment and Analysis of the Results
5. Results
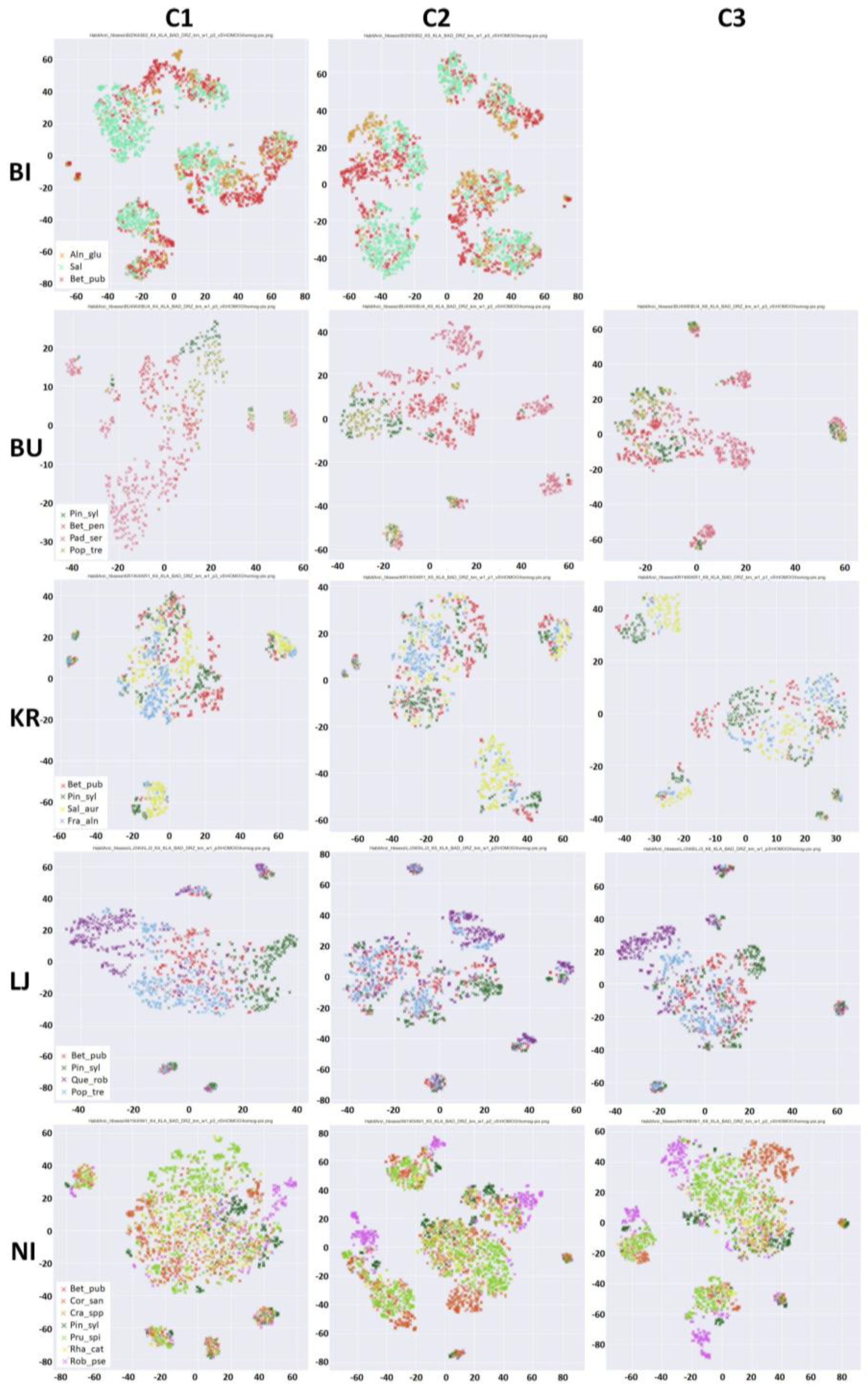
5.1. Results of Implementation of the t-SNE Algorithm
5.2. Results of Accuracy Assessment
5.3. Comparison of the Extent of Species in Different Seasons
6. Discussion
6.1. Influence of Acquisition Date on the Classification Results
6.1.1. BI Research Area
6.1.2. BU Research Area
6.1.3. KR Research Area
6.1.4. LJ Research Area
6.1.5. NI Research Area
6.2. Comparative Analysis of Species’ Extent in Different Seasons
6.3. Comparison of the Obtained Results with Previous Research Studies
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors of the Study | Acquisition Data | Type of Data | Methods | No. of Classes/OA/K | Species of Interest | Accuracy |
|---|---|---|---|---|---|---|
| Dalponte et al., 2008 [56] | 28 June 2006 | HS, 1 m; LiDAR, 5.6 pt/m2 | Support vector machines | 23/-/0.88 |
|
|
| Dalponte et al., 2012 [36] | 16 July 2008 | HS, 1 m; LiDAR, 8.6 pt/m2 | Support vector machines | 8/-/0.772 | Pinus sylvestris | PA: 0.843, UA: 0.820 |
| Immitzer et al., 2012 [37] | 10 July 2010 | WorldView-2, 2 m | Random forest | 10/0.728/0.678 |
|
|
| Priedītis et al., 2015 [91] | 27 October 2014 | HS, 0.5 m | Linear discriminant analysis | 5/-/- |
|
|
| Raczko & Zagajewski 2017 [88] | 10 September 2012 | HS, 2.5 m | Random forest | 5/0.62/0.52 | Betula pendula | PA: 0.50, UA: 0.55 |
| Raczko & Zagajewski 2017 [88] | 10 September 2012 | HS, 2.5 m | Support vector machines | 5/0.68/0.61 | Betula pendula | PA: 0.59, UA: 0.57 |
| Raczko & Zagajewski 2017 [88] | 10 September 2012 | HS, 2.5 m | Artificial neural networks | 5/0.77/0.72 | Betula pendula | PA: 0.68, UA: 0.69 |
| Richter et al., 2016 [64] | 2 August 2013 | HS, 2 m | Partial least square discriminant analysis | 10/0.705/- | Alnus glutinosa Quercus robur | PA: 0.78, UA: 0.69 PA: 0.72, UA: 0.58 |
| Richter et al., 2016 [64] | 25 September 2011 | HS, 2 m | Partial least square discriminant analysis | 10/0.746/- | Alnus glutinosa Quercus robur | PA: 0.56, UA: 0.55 PA: 0.44, UA: 0.52 |
| Richter et al., 2016 [64] | 2 August 2013 and 25 September 2011 | HS, 2 m | Partial least square discriminant analysis | 10/0.784/- | Alnus glutinosa Quercus robur | PA: 0.89, UA: 0.86 PA: 0.59, UA: 0.64 |
| Trier et al., 2018 [46] | 2 May - 20 June 2015 | HS, 1 m; LiDAR, 5 pt/m2 | Deep neural network | 3/0.78/- | Pine Birch | PA: 1.00, UA: 0.88 PA: 0.70, UA: 0.61 |
| Tuominen et al., 2018 [91] | 21 August 2015 | HS, 20 cm; DSM | K-nearest neighbour method + genetic algorithm | 14/0.823/0.808 |
|
|
| Wietecha et al., 2019 [89] | 19 August 2015 | HS, 2.5 m, 5 m; LiDAR, 10 pt/m2 | Support vector machines | 5/0.922/0.810 |
|
|
Appendix B




References
- EEA. EU Biodiversity Strategy to 2020. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:52011DC0244&from=EN (accessed on 10 June 2019).
- Waldron, A.; Miller, D.C.; Redding, D.; Mooers, A.; Kuhn, T.S.; Nibbelink, N.; Roberts, J.T.; Tobias, J.A.; Gittleman, J.L. Reductions in global biodiversity loss predicted from conservation spending. Nature 2017, 551, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- EEA. EU Biodiversity Action Plan. Available online: http://ec.europa.eu/environment/nature/ biodiversity/comm2006/index_en.htm (accessed on 10 June 2019).
- Chan, J.C.W.; Spanhove, T.; Ma, J.; Borre, J.V.; Paelinckx, D.; Canters, F. Natura 2000 habitat identification and conservation status assessment with superresolution enhanced hyperspectral (CHRIS/Proba) Imagery. In Proceedings of the GEOBIA 2010 Geographic Object-Based Image Analysis, Ghent, Belgium, 29 June–2 July 2010. [Google Scholar]
- Lucas, R.; Medcalf, K.; Brown, A.; Bunting, P.; Breyer, J.; Clewley, D.; Keyworth, S.; Blackmore, P. Updating the phase 1 habitat map of Wales UK, using satellite sensor data. ISPRS J. Photogrammetry Remote Sens. 2011, 66, 81–102. [Google Scholar] [CrossRef]
- Schuster, C.; Schmidt, T.; Conrad, C.; Kleinschmit, B.; Förster, M. Grassland habitat mapping by intra-annual time series analysis–comparison of RapidEye and TerraSAR-X satellite data. Int. J. Appl. Earth Obs. Geoinf. 2015, 34, 25–34. [Google Scholar] [CrossRef]
- Haest, B.; Vanden Borre, J.; Spanhove, T.; Thoonen, G.; Delalieux, S.; Kooistra, L.; Mücher, C.A.; Paelinckx, D.; Scheunders, P.; Kempeneers, P. Habitat mapping and quality assessment of NATURA 2000 heathland using airborne imaging spectroscopy. Remote Sens. 2017, 9, 266. [Google Scholar] [CrossRef]
- Mahdianpari, M.; Salehi, B.; Mohammadimanesh, F.; Homayouni, S.; Gill, E. The first wetland inventory map of Newfoundland at a spatial resolution of 10 m using Sentinel-1 and Sentinel-2 data on the Google Earth engine cloud computing platform. Remote Sens. 2019, 11, 43. [Google Scholar] [CrossRef]
- Zlinszky, A.; Deák, B.; Kania, A.; Schroiff, A.; Pfeifer, N. Mapping Natura 2000 habitat conservation status in a Pannonic salt steppe with airborne laser scanning. Remote Sens. 2015, 7, 2991–3019. [Google Scholar] [CrossRef]
- Schmidt, J.; Fassnacht, F.E.; Forster, F.; Schmidtlein, S. Synergetic use of Sentinel-1 and Sentinel-2 for assessments of heathland conservation status. Remote Sens. Ecol. Conserv. 2017, 4, 225–239. [Google Scholar] [CrossRef]
- Huang, C.Y.; Asner, G.P. Applications of remote sensing to alien invasive plant studies. Sensors 2009, 9, 4869–4889. [Google Scholar] [CrossRef] [PubMed]
- Dorigo, W.; Lucieer, A.; Podobnikar, T.; Carni, A. Mapping invasive Fallopia japonica by combined spectral, spatial, and temporal analysis of digital orthophotos. Int. J. Appl. Earth Obs. Geoinf. 2012, 19, 185–195. [Google Scholar] [CrossRef]
- Bradley, B.A. Remote detection of invasive plants: A review of spectral, textural and phenological approaches. Biol. Invasions 2014, 16, 1411–1425. [Google Scholar] [CrossRef]
- Marcinkowska-Ochtyra, A.; Jarocińska, A.; Bzdęga, K.; Tokarska-Guzik, B. Classification of expansive grassland species in different growth stages based on hyperspectral and LiDAR data. Remote Sens. 2018, 10, 2019. [Google Scholar] [CrossRef]
- Batistella, M.; Lu, D. Integrating field data and remote sensing to identify secondary succession stages in the Amazon. In Proceedings of the 29th International Symposium on Remote Sensing of Environment, Buenos Aires, Argentina, 8–12 April 2002. [Google Scholar]
- Szostak, M.; Wężyk, P.; Hawryło, P.; Puchała, M. Monitoring the secondary forest succession and land cover/use changes of the Błędów Desert (Poland) using geospatial analyses. Quaest. Geogr. 2016, 35, 5–13. [Google Scholar] [CrossRef]
- Osińska-Skotak, K.; Jełowicki, Ł.; Bakuła, K.; Michalska-Hejduk, D.; Wylazłowska, J.; Kopeć, D. Analysis of using dense image matching techniques to study the process of secondary succession in non-forest Natura 2000 habitats. Remote Sens. 2019, 11, 893. [Google Scholar] [CrossRef]
- Dabrowska-Zielinska, K.; Musial, J.; Malinska, A.; Budzynska, M.; Gurdak, R.; Kiryla, W.; Bartold, M.; Grzybowski, P. Soil moisture in the Biebrza Wetlands retrieved from Sentinel-1 imagery. Remote Sens. 2018, 10, 1979. [Google Scholar] [CrossRef]
- Perennou, C.; Guelmami, A.; Paganini, M.; Philipson, P.; Poulin, B.; Strauch, A.; Tottrup, C.; Truckenbrodt, J.; Geijzendorffer, I.R. Mapping Mediterranean wetlands with remote sensing: A good-looking map is not always a good map. Adv. Ecol. Res. 2018, 58, 243–277. [Google Scholar]
- Ciężkowski, W.; Jóźwiak, J.; Szporak-Wasilewska, S.; Dabrowski, P.; Góraj, M.; Kleniewska, M.; Chormański, J. Water stress index for bogs and mires based on UAV land surface measurements and its dependency on airborne hyperspectral data. In Proceedings of the 2018 IEEE International Geoscience and Remote Sensing Symposium, Valencia, Spain, 22–27 July 2018. [Google Scholar]
- Ostermann, O.P. The need for management of nature conservation sites designated under Natura 2000. J. Appl. Ecol. 1998, 35, 968–973. [Google Scholar] [CrossRef]
- Halada, L.; Evans, D.; Romão, C.; Petersen, J.E. Which habitats of European importance depend on agricultural practices? Biodivers. Conserv. 2011, 20, 2365–2378. [Google Scholar] [CrossRef]
- Prévosto, B.; Kuiters, L.; Bernhardt-Römermann, M.; Dölle, M.; Schmidt, W.; Hoffmann, M.; Van Uytvanck, J.; Bohner, A.; Kreiner, D.; Stadler, J.; et al. Impacts of Land Abandonment on Vegetation: Successional Pathways in European Habitats. Folia Geobot. 2011, 46, 303–325. [Google Scholar] [CrossRef]
- Barabasz, B. Wpływ modyfikacji tradycyjnych metod gospodarowania na przemiany roślinności łąk z klasy Molinio-Arrhenatheretea. (The effect of traditional management methods modifications on changes in meadows of Molinio-Arrhenatheretea class). Wiadomości Bot. 1994, 38, 85–94. [Google Scholar]
- Falińska, K. Plant population processes in the course of forest succession in abandoned meadows. I. Variability and diversity of floristic composition, and biological mechanisms of species turnover. Acta Soc. Bot. Pol. 1989, 58, 439–465. [Google Scholar] [CrossRef][Green Version]
- Falińska, K. Plant population processes in the course of forest succession in abandoned meadows. II. Demography and succession promotors. Acta Soc. Bot. Pol. 1989, 58, 467–491. [Google Scholar] [CrossRef][Green Version]
- Mróz, W. Monitoring Siedlisk Przyrodniczych. Przewodnik Metodyczny [Natura 2000 Habitat Monitoring]; Part I; Biblioteka Monitoringu Środowiska: Warszawa, Poland, 2010. [Google Scholar]
- Mróz, W. Monitoring Siedlisk Przyrodniczych. Przewodnik Metodyczny [Natura 2000 Habitat Monitoring]; Part II; Biblioteka Monitoringu Środowiska: Warszawa, Poland, 2012. [Google Scholar]
- Mróz, W. Monitoring Siedlisk Przyrodniczych. Przewodnik Metodyczny [Natura 2000 Habitat Monitoring]; Part III; Biblioteka Monitoringu Środowiska: Warszawa, Poland, 2013. [Google Scholar]
- Rohde, W.G.; Olson, C.E., Jr. Multispectral sensing of forest tree species. Photogramm. Eng. Remote. Sens. 1972, 38, 1209–1215. [Google Scholar]
- Rignot, E.J.M.; Williams, C.L.; Way, J.; Viereck, L.A. Mapping of forest types in Alaskan boreal forests using SAR imagery. IEEE Trans. Geosci. Remote Sens. 1994, 32, 1051–1059. [Google Scholar] [CrossRef]
- Saatchi, S.; Rignot, E. Classification of boreal forest cover types using SAR images. Remote Sens. Environ. 1997, 60, 270–281. [Google Scholar] [CrossRef]
- Boschetti, M.; Boschetti, L.; Oliveri, S.; Casati, L.; Canova, I. Tree species mapping with airborne hyper-spectral MIVIS data: The Ticino Park study case. Int. J. Remote Sens. 2007, 28, 1251–1261. [Google Scholar] [CrossRef]
- Suratno, A.; Seielstad, C.; Queen, L. Tree species identification in mixed coniferous forest using airborne laser scanning. ISPRS J. Photogramm. Remote Sens. 2009, 64, 683–693. [Google Scholar] [CrossRef]
- Dalponte, M.; Bruzzone, L.; Gianelle, D. Tree species classification in the Southern Alps based on the fusion of very high geometrical resolution multispectral/hyperspectral images and LiDAR data. Remote Sens. Environ. 2012, 123, 258–270. [Google Scholar] [CrossRef]
- Immitzer, M.; Atzberger, C.; Koukal, T. Tree species classification with random forest using very high spatial resolution 8-band WorldView-2 satellite data. Remote Sens. 2012, 4, 2661–2693. [Google Scholar] [CrossRef]
- Ørka, H.O.; Dalponte, M.; Gobakken, T.; Næsset, E.; Ene, L.T. Characterizing forest species composition using multiple remote sensing data sources and inventory approaches. Scand. J. For. Res. 2013, 28, 677–688. [Google Scholar] [CrossRef]
- Korpela, I.; Mehtätalo, L.; Seppänen, A.; Kangas, A. Tree species identification in aerial image data using directional reflectance signatures. Silva Fenn. 2014, 48, 1087. [Google Scholar] [CrossRef]
- Somers, B.; Asner, G.P. Tree species mapping in tropical forests using multi-temporal imaging spectroscopy: Wavelength adaptive spectral mixture analysis. Int. J. Appl. Earth Obs. Geoinf. 2014, 31, 57–66. [Google Scholar] [CrossRef]
- Ballanti, L.; Blesius, L.; Hines, E.; Kruse, B. Tree species classification using hyperspectral imagery: A comparison of two classifiers. Remote Sens. 2016, 8, 445. [Google Scholar] [CrossRef]
- Hauglin, M.; Ørka, H.O. Discriminating between native Norway spruce and invasive Sitka spruce—A comparison of multitemporal Landsat 8 imagery, aerial images and airborne laser scanner data. Remote Sens. 2016, 8, 363. [Google Scholar] [CrossRef]
- Ørka, H.O.; Hauglin, M. Use of remote sensing for mapping of non-native conifer species. INA Fagrapp. 2016, 33, 76. [Google Scholar]
- Piiroinen, R.; Heiskanen, J.; Maeda, E.; Viinikka, A.; Pellikka, P. Classification of tree species in a diverse African agroforestry landscape using imaging spectroscopy and laser scanning. Remote Sens. 2017, 9, 875. [Google Scholar] [CrossRef]
- Shen, X.; Cao, L. Tree-species classification in subtropical forests using airborne hyperspectral and LiDAR data. Remote Sens. 2017, 9, 1180. [Google Scholar] [CrossRef]
- Trier, Ø.D.; Salberg, A.B.; Kermit, M.; Rudjord, Ø.; Gobakken, T.; Næsset, E.; Aarsten, D. Tree species classification in Norway from airborne hyperspectral and airborne laser scanning data. Eur. J. Remote Sens. 2018, 51, 336–351. [Google Scholar] [CrossRef]
- Grabska, E.; Hostert, P.; Pflugmacher, D.; Ostapowicz, K. Forest stand species mapping using the Sentinel-2 time series. Remote Sens. 2019, 11, 1197. [Google Scholar] [CrossRef]
- Voss, M.; Sugumaran, R. Seasonal effect on tree species classification in an urban environment using hyperspectral data, LiDAR, and an object-oriented approach. Sensors 2008, 8, 3020–3036. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.R.; Hardin, P.J.; Hardin, A.J. Classification of urban tree species using hyperspectral imagery. Geocarto Int. 2012, 27, 443–458. [Google Scholar] [CrossRef]
- Zhang, K.; Hu, B. Individual urban tree species classification using very high spatial resolution airborne multi-spectral imagery using longitudinal profiles. Remote Sens. 2012, 4, 1741–1757. [Google Scholar] [CrossRef]
- Zhang, C.; Qiu, F. Mapping individual tree species in an urban forest using airborne LiDAR data and hyperspectral imagery. Photogramm. Eng. Remote Sens. 2012, 78, 1079–1087. [Google Scholar] [CrossRef]
- Alonzo, M.; Bookhagen, B.; Roberts, D.A. Urban tree species mapping using hyperspectral and LiDAR data fusion. Remote Sens. Environ. 2014, 148, 70–83. [Google Scholar] [CrossRef]
- Chance, C.M.; Coops, N.C.; Crosby, K.; Aven, N. Spectral wavelength selection and detection of two invasive plant species in an urban area. Can. J. Remote. Sens. 2016, 41, 1–14. [Google Scholar] [CrossRef]
- Brabant, C.; Alvarez-Vanhard, E.; Laribi, A.; Morin, G.; Thanh Nguyen, K.; Thomas, A.; Houet, T. Comparison of hyperspectral techniques for urban tree diversity classification. Remote Sens. 2019, 11, 1269. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Latifi, H.; Stereńczak, K.; Modzelewska, A.; Lefsky, M.; Waser, L.T.; Straub, C.; Ghosh, A. Review of studies on tree species classification from remotely sensed data. Rem. Sens. Environ. 2016, 186, 64–87. [Google Scholar] [CrossRef]
- Dalponte, M.; Bruzzone, L.; Gianelle, D. Fusion of hyperspectral and LIDAR remote sensing data for classification of complex forest areas. IEEE Trans. Geosci. Remote Sens. 2008, 46, 1416–1427. [Google Scholar] [CrossRef]
- Coops, N.C.; Hilker, T.; Wulder, M.A.; St-Onge, B.; Newnham, G.; Siggins, A.; Trofymow, J.A. Estimating canopy structure of Douglas-fir forest stands from discrete-return LiDAR. Trees-Struct. Funct. 2007, 21, 295–310. [Google Scholar] [CrossRef]
- Ørka, H.O.; Næsset, E.; Bollandsås, M. Classifying species of individual trees by intensity and structure features derived from airborne laser scanner data. Remote Sens. Environ. 2009, 113, 1163–1174. [Google Scholar] [CrossRef]
- Key, T.; Warner, T.A.; McGraw, J.B.; Fajvan, M.A. A comparison of multispectral and multitemporal information in high spatial resolution imagery for classification of individual tree species in a temperate hardwood forest. Remote Sens. Environ. 2001, 75, 100–112. [Google Scholar] [CrossRef]
- Matthew, E.F.; DeFries, R.S.; Sesnie, S.E.; Arroyo-Mora, J.P.; Soto, C.; Singh, A.; Townsend, P.A.; Chazdon, R.L. Mapping species composition of forests and tree plantations in Northeastern Costa Rica with an integration of hyperspectral and multitemporal Landsat imagery. Remote Sens. 2015, 7, 5660–5696. [Google Scholar]
- Hościło, A.; Lewandowska, A. Mapping forest type and tree species on a regional scale using multi-temporal Sentinel-2 data. Remote Sens. 2019, 11, 929. [Google Scholar] [CrossRef]
- Mickelson, J.G.; Civco, D.L.; Silander, J.A. Delineating forest canopy species in the northeastern United States using multi-temporal TM imagery. Photogramm. Eng. Remote. Sens. 1998, 64, 891–904. [Google Scholar]
- Dymond, C.C.; Mladenoff, D.J.; Radeloff, V.C. Phenological differences in Tasseled Cap indices improve deciduous forest classification. Remote Sens. Environ. 2002, 80, 460–472. [Google Scholar] [CrossRef]
- Richter, R.; Reu, B.; Wirtha, C.; Doktor, D.; Vohland, M. The use of airborne hyperspectral data for tree species classification in a species-rich Central European forest area. Int. J. Appl. Earth Obs. 2016, 52, 464–474. [Google Scholar] [CrossRef]
- Strzyż, M.; Wójtowicz, B.; Winiarczyk-Raźniak, A. The influence of areas connected with Natura 2000 on the development and functioning of the environmental potential based on the example of Ponidzie Administrative District. In Proceedings of the 3rd International Geography Symposium–GEOMED, Kemer-Antalya, Turkey, 10–13 June 2013; pp. 645–660. [Google Scholar]
- Strzyż, M. The active protection and monitoring of xerothermic reserves based on the example of the Skowronno reserve in Ponidzie. Rocz. Świętokrzyski. Ser. B–Nauk. Przyr. 2014, 35, 133–162. [Google Scholar]
- Sławik, Ł.; Niedzielko, J.; Kania, A.; Piórkowski, H.; Kopeć, D. Multiple flights or single flight instrument fusion of hyperspectral and ALS data? A comparison of their performance for vegetation mapping. Remote Sens. 2019, 11, 970. [Google Scholar] [CrossRef]
- IMGiW. Climate Maps. Available online: http://klimat.pogodynka.pl/pl/climate-maps/ (accessed on 6 May 2019).
- Van der Maaten, L.J.P.; Hinton, G.E. Visualizing high-dimensional data using t-SNE. J. Mach. Learn. Res. 2008, 9, 2579–2605. [Google Scholar]
- HySpex Products. Available online: https://www.hyspex.no/products/ (accessed on 22 February 2018).
- Schläpfer, D. Parametric Geocoding, PARGE User Guide; ver. 3.3; ReSe Applications Schläpfer: Wil, Switzerland, 2016; p. 270. [Google Scholar]
- Richter, R.; Schlapfer, D. ATCOR4 Manual. ReSe Applications. Available online: https://www.rese-apps.com/pdf/atcor4_manual.pdf (accessed on 1 March 2018).
- Green, A.A.; Berman, M.; Switzer, P.; Craig, M.D. A transformation for ordering multispectral data in terms of image quality with implications for noise removal. IEEE Trans. Geosci. Remote Sens. 1988, 26, 65–74. [Google Scholar] [CrossRef]
- Clark, M.L.; Roberts, D.A.; Clark, D.B. Hyperspectral discrimination of tropical rain forest tree species at leaf to crown scales. Remote Sens. Environ. 2005, 96, 375–398. [Google Scholar] [CrossRef]
- Alonzo, M.; Roth, K.; Roberts, D. Identifying Santa Barbara’s urban tree species from AVIRIS imagery using canonical discriminant analysis. Remote Sens. Lett. 2013, 4, 513–521. [Google Scholar] [CrossRef]
- Ghosh, A.; Fassnacht, F.E.; Joshia, P.K.; Koch, B. A framework for mapping tree species combining hyperspectral and LiDAR data: Role of selected classifiers and sensor across three spatial scales. Int. J. Appl. Earth Obs. 2014, 26, 49–63. [Google Scholar] [CrossRef]
- Ghiyamat, A.; Shafri, H.Z.M.; Shariff, A.R.M. Influence of tree species complexity on discrimination performance of vegetation indices. Eur. J. Remote Sens. 2016, 49, 15–37. [Google Scholar] [CrossRef]
- Axelsson, P. Processing of laser scanner data-algorithms and applications. ISPRS J. Photogramm. 1999, 54, 138–147. [Google Scholar] [CrossRef]
- Mandlburger, G.; Otepka, J.; Karel, W.; Hollaus, M.; Ressl, C.; Haring, A.; Briese, C.; Molnar, G.; Höfle, B. OPALS–A comprehensive laser scanning software for geomorphological analysis. Geophys. Res. Abs. 2011, 13, 2011–7933. [Google Scholar]
- Höfle, B.; Mücke, W.; Dutter, M.; Rutzinger, M.; Dorninger, P. Detection of building regions using airborne LiDAR–a new combination of raster and point cloud based GIS methods. In Proceedings of the GI-Forum 2009-International Conference on Applied Geoinformatics, Salzburg, Austria, 7–10 July 2009; pp. 66–75. [Google Scholar]
- BCAL, Boise, ID, USA. BCAL LiDAR Tools. 2016. Available online: https://bcal.boisestate.edu/tools/lidar (accessed on 18 June 2019).
- Evans, J.S.; Hudak, A.T.; Faux, R.; Smith, A. Discrete return LiDAR in natural resources: Recommendations for project planning, data processing, and deliverables. Remote Sens. 2009, 1, 776–794. [Google Scholar] [CrossRef]
- Kohavi, R.; John, G.H. Wrappers for feature subset selection. Artif. Intell. 1997, 97, 273–324. [Google Scholar] [CrossRef]
- Guyon, I.; Elisseeff, A. An Introduction to Variable and Feature Selection. J. Mach. Learn. Res. 2003, 3, 1157–1182. [Google Scholar]
- Breiman, L. Random forest. Mach. Lear. 2001, 45, 1–33. [Google Scholar]
- Congalton, R.G.; Green, K. Assessing the Accuracy of Remotely Sensed Data: Principles and Practices; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Powers, D.M.W. Evaluation: From Precision, Recall and F-Measure to ROC, Informedness, Markedness & Correlation. J. Mach. Learn. Technol. 2011, 2, 37–63. [Google Scholar]
- Raczko, E.; Zagajewski, B. Comparison of support vector machine, random forest and neural network classifiers for tree species classification on airborne hyperspectral APEX images. Eur. J. Remote Sens. 2017, 50, 144–154. [Google Scholar] [CrossRef]
- Wietecha, M.; Jełowicki, M.; Mitelsztedt, K.; Miścicki, S.; Stereńczak, K. The capability of species-related forest stand characteristics determination with the use of hyperspectral data. Remote Sens. Environ. 2019, 231, 111232. [Google Scholar] [CrossRef]
- Priedītis, G.; Šmits, I.; Daģis, S.; Paura, L.; Krūmiņš, J.; Dubrovskis, D. Assessment of hyperspectral data analysis methods to classify tree species. Res. Rural Dev. 2015, 2, 7–13. [Google Scholar]
- Tuominen, S.; Näsi, R.; Honkavaara, E.; Balazs, A.; Hakala, T.; Viljanen, N.; Pölönen, I.; Saari, H.; Ojanen, H. Assessment of classifiers and remote sensing features of hyperspectral imagery and stereo-photogrammetric point clouds for recognition of tree species in a forest area of high species diversity. Remote Sens. 2018, 10, 714. [Google Scholar] [CrossRef]










| Natura 2000 Site | Natura 2000 Code | Geographical Coordinates | Acronym | Study Area (km2) | Type of Natura 2000 Habitat Studied (code) | Promoters of Succession |
|---|---|---|---|---|---|---|
| Biebrza River Valley | PLH200008 | 53°17′10″N; 22°37′45″E | BI | 37 | Alkaline fens (7230) |
|
| Lucynów-Mostówka Inland Dunes | PLH140013 | 52°35′34″N; 21°27′30″E | BU | 7 | European dry heaths (4030) |
|
| Nidziańska Refuge | PLH26003 | 50°32′14″N; 20°30′42″E | NI | 23 | Semi-natural dry grasslands and scrubland facies on calcareous substrates (Festuco-Brometalia) (6210) |
|
| Krasna Valley | PLH260001 | 50°45″N; 19°17″E | KR | 41 | European dry heaths (4030); species-rich Nardus grasslands on siliceous substrates in mountain areas (6230); Molinia meadows on calcareous, peaty or clayey, silt-laden soils (Molinion caeruleae) (6410) |
|
| Janowskie Forests Ranges | PLH060031 | 50°43″N; 22°0″E | LJ | 43 | European dry heaths (4030); transition mires and quaking bogs (7140) |
|
| Research Area | Remote Sensing Data Acquisition Time | ||
|---|---|---|---|
| C1 Flight Campaign (spring) | C2 Flight Campaign (summer) | C3 Flight Campaign (autumn) | |
| BI | 27 June 2017 | 9 August 2017 | - |
| BU | 28 May 2017 | 10 July 2017 | 9 September 2017 |
| KR | 1 June 2017 | 7 July 2017 | 27 September 2017 |
| LJ | 2 June 2017 | 19 July 2017 | 9 September 2017 |
| NI | 18 May 2017 | 30 July 2017 | 27 September 2017 |
| Technical Parameters | Sensor Type | ||
|---|---|---|---|
| Airborne Laser Scanner (FWF) Riegl LMS-Q680i | HySpex Hyperspectral Camera | ||
| VNIR-1800 | SWIR-384 | ||
| Flight Altitude AGL (m) | 500 | ||
| Point density (points/m2) | 7 | - | - |
| Spatial resolution (m) | - | 0.5 | 1 |
| Spectral resolution | 1.55 µm | 430 bands encompassing 0.4–2.4 µm spectral range | |
| Spectral sampling (nm) | - | 3.26 | 5.45 |
| FOV max (degrees) | 60 | 34 | 32 |
| Overlap (%) | 62.7 | 30 | 30 |
| Overlap width (m) | 855 | 450 | 450 |
| Feature Type | Statistics Generated/Description | ALS Classes |
|---|---|---|
| DTM | SigmaZ, slope, exposition, TPI | GRD |
| DSM | SigmaZ, slope, exposition | ALL |
| nDSM | Min, max, mean, variance, median, range, RMS, Percentiles: 5, 10, 25, 75, 90, 95 | ALL, VEG |
| Amplitude | Min, max, mean, variance, median, range, RMS | ALL, GRD, VEG |
| Echo ratio | Min, max, mean, variance, median, range, RMS | ALL, GRD, VEG |
| Point density | Number of pts/m2 | ALL, GRD, VEG |
| Total vegetation density | Ratio of vegetation to ground points | - |
| Vegetation cover | Ratio of vegetation to all points | - |
| IQR (interquartile range of height) | Range between quantiles 0.75 and 0.25 | - |
| Research Area | Number of Reference Polygons Per Class | Number of Classes | Total Number of Reference Polygons |
|---|---|---|---|
| BI | 70 | 3 | 210 |
| BU | 20 | 4 | 80 |
| KR | 29 | 4 | 116 |
| LJ | 25 | 4 | 100 |
| NI | 29 | 6 | 174 |
| Study Area | |||||
|---|---|---|---|---|---|
| Flight Campaigns | BI | BU | KR | LJ | NI |
| spring (C1)–summer (C2) | 66.8% | 59.4% | 48.7% | 51.9% | 46.1% |
| spring (C1)–autumn (C3) | - | 50.7% | 49.1% | 57.2% | 43.8% |
| summer (C2)–autumn (C3) | - | 49.1% | 51.9% | 53.0% | 52.6% |
| spring (C1)–summer (C2)–autumn (C3) | - | 33.7% | 31.8% | 35.7% | 32.2% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osińska-Skotak, K.; Radecka, A.; Piórkowski, H.; Michalska-Hejduk, D.; Kopeć, D.; Tokarska-Guzik, B.; Ostrowski, W.; Kania, A.; Niedzielko, J. Mapping Succession in Non-Forest Habitats by Means of Remote Sensing: Is the Data Acquisition Time Critical for Species Discrimination? Remote Sens. 2019, 11, 2629. https://doi.org/10.3390/rs11222629
Osińska-Skotak K, Radecka A, Piórkowski H, Michalska-Hejduk D, Kopeć D, Tokarska-Guzik B, Ostrowski W, Kania A, Niedzielko J. Mapping Succession in Non-Forest Habitats by Means of Remote Sensing: Is the Data Acquisition Time Critical for Species Discrimination? Remote Sensing. 2019; 11(22):2629. https://doi.org/10.3390/rs11222629
Chicago/Turabian StyleOsińska-Skotak, Katarzyna, Aleksandra Radecka, Hubert Piórkowski, Dorota Michalska-Hejduk, Dominik Kopeć, Barbara Tokarska-Guzik, Wojciech Ostrowski, Adam Kania, and Jan Niedzielko. 2019. "Mapping Succession in Non-Forest Habitats by Means of Remote Sensing: Is the Data Acquisition Time Critical for Species Discrimination?" Remote Sensing 11, no. 22: 2629. https://doi.org/10.3390/rs11222629
APA StyleOsińska-Skotak, K., Radecka, A., Piórkowski, H., Michalska-Hejduk, D., Kopeć, D., Tokarska-Guzik, B., Ostrowski, W., Kania, A., & Niedzielko, J. (2019). Mapping Succession in Non-Forest Habitats by Means of Remote Sensing: Is the Data Acquisition Time Critical for Species Discrimination? Remote Sensing, 11(22), 2629. https://doi.org/10.3390/rs11222629