Nitrogen and Phosphorus Effect on Sun-Induced Fluorescence and Gross Primary Productivity in Mediterranean Grassland

 , , ,
, , ,  , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Nutrient Manipulation Experiment, Gross Primary Production and Ancillary Data
2.3. Transpiration Estimates
2.4. Field Spectroscopy, Retrieval of Sun-Induced Fluorescence and Biophysical Properties
2.5. SCOPE Model Simulations
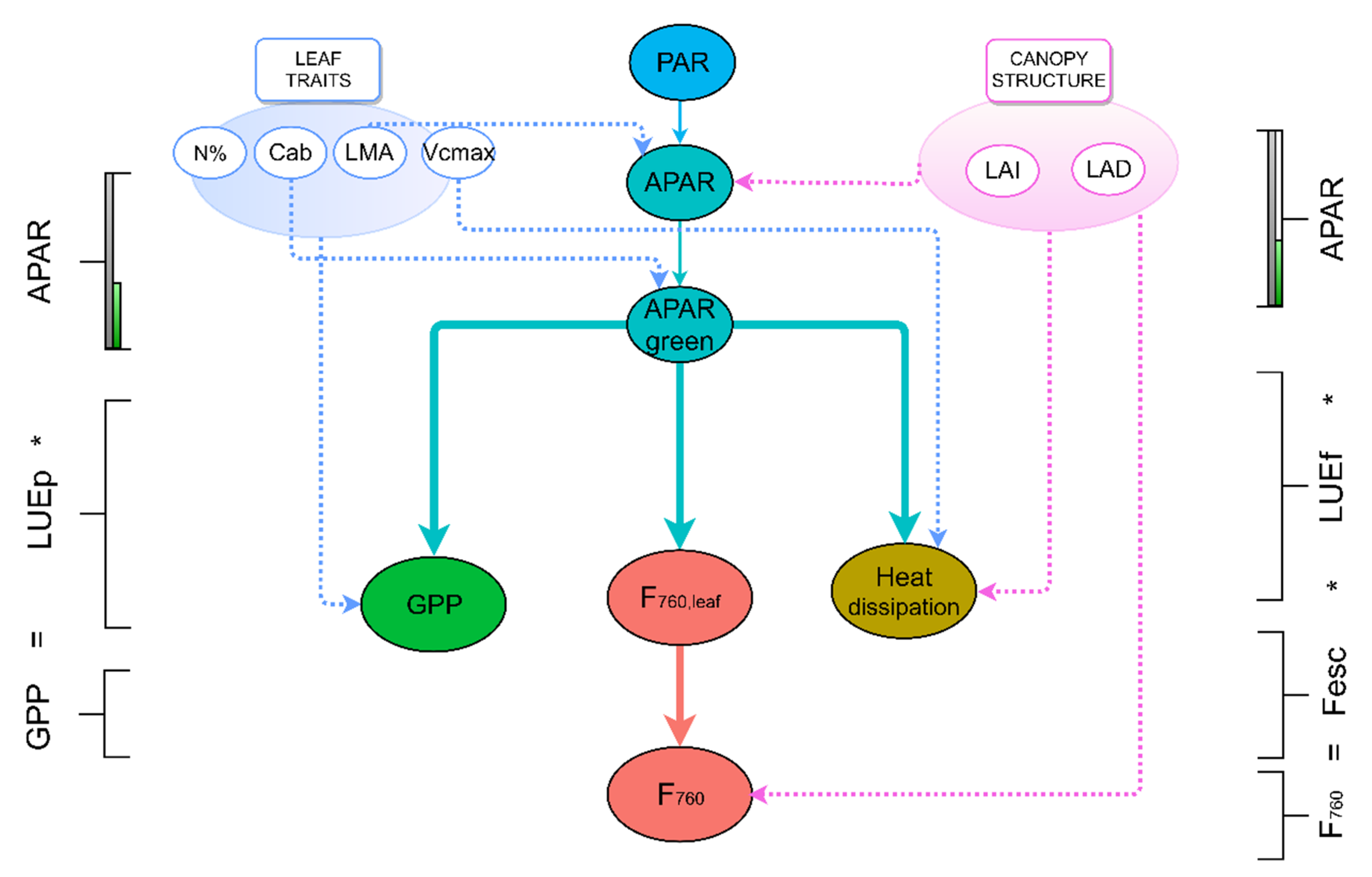
2.6. Calculation of the Light Use Efficiency of Photosynthesis (LUEp), Light use Efficiency of Fluorescence Emission (LUEf) and Escape Probability of F760 (Fesc)
2.7. Statistical Analysis
3. Results
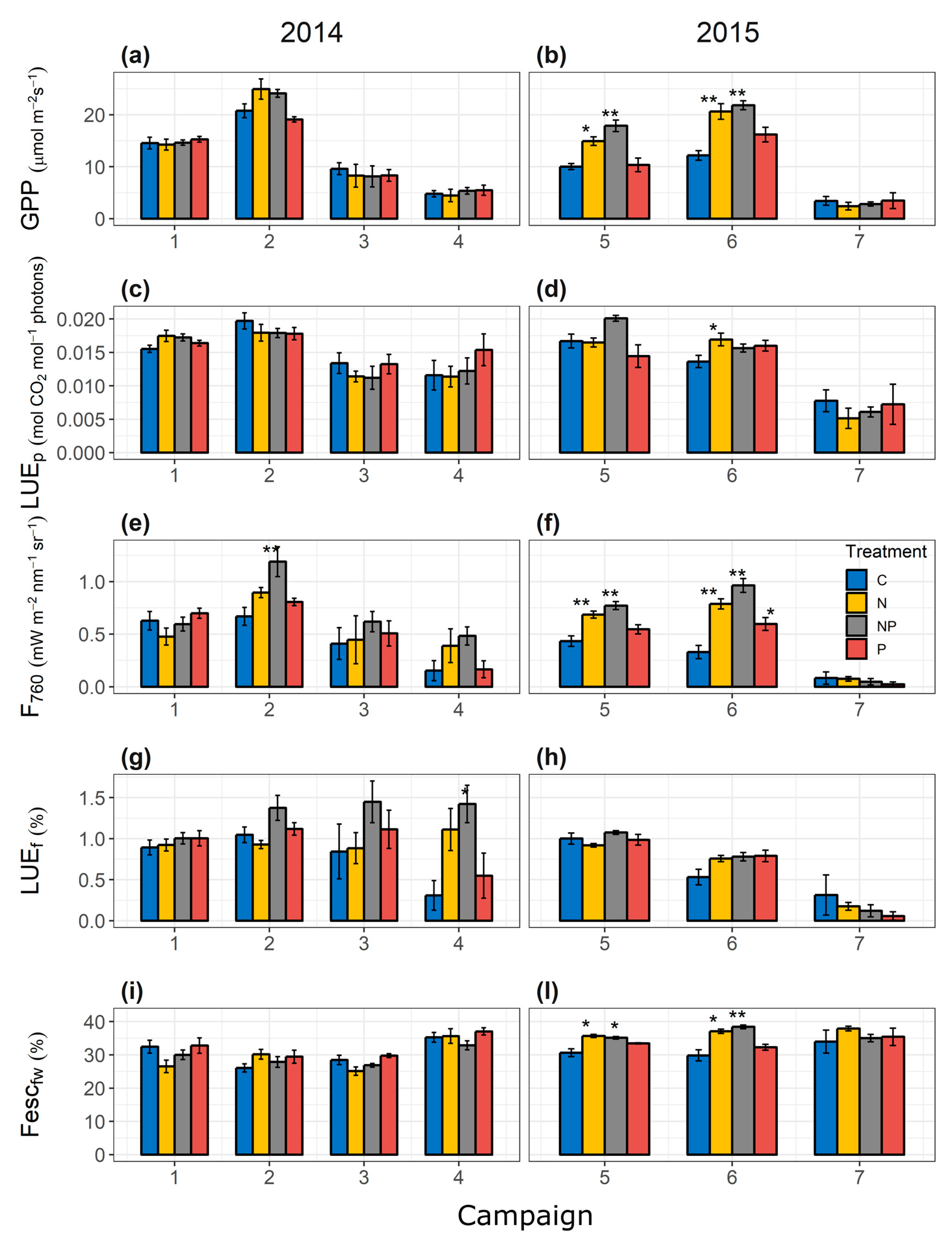
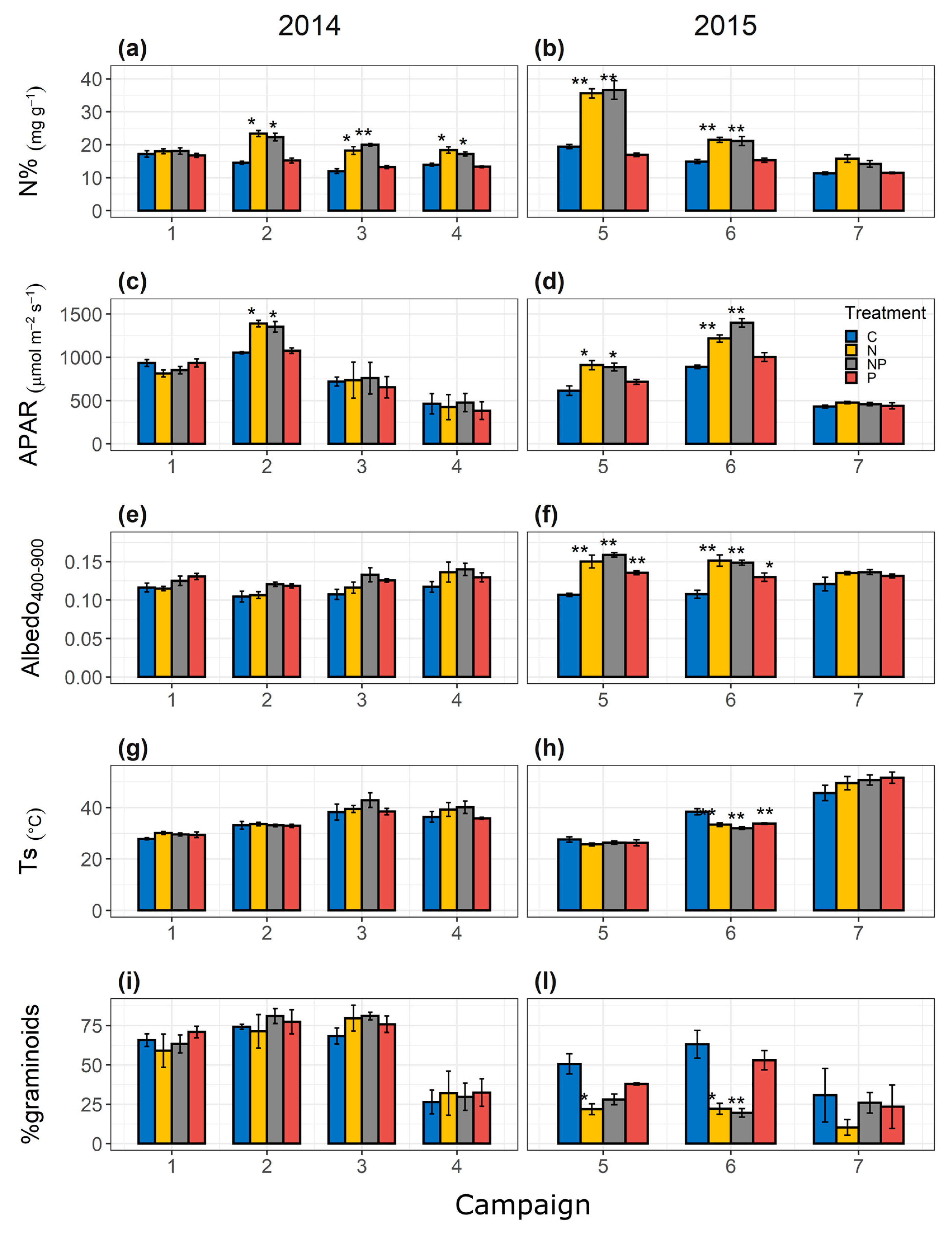
3.1. Description of Fertilization Effects on Fluxes, Optical Data, and Vegetation Characteristics
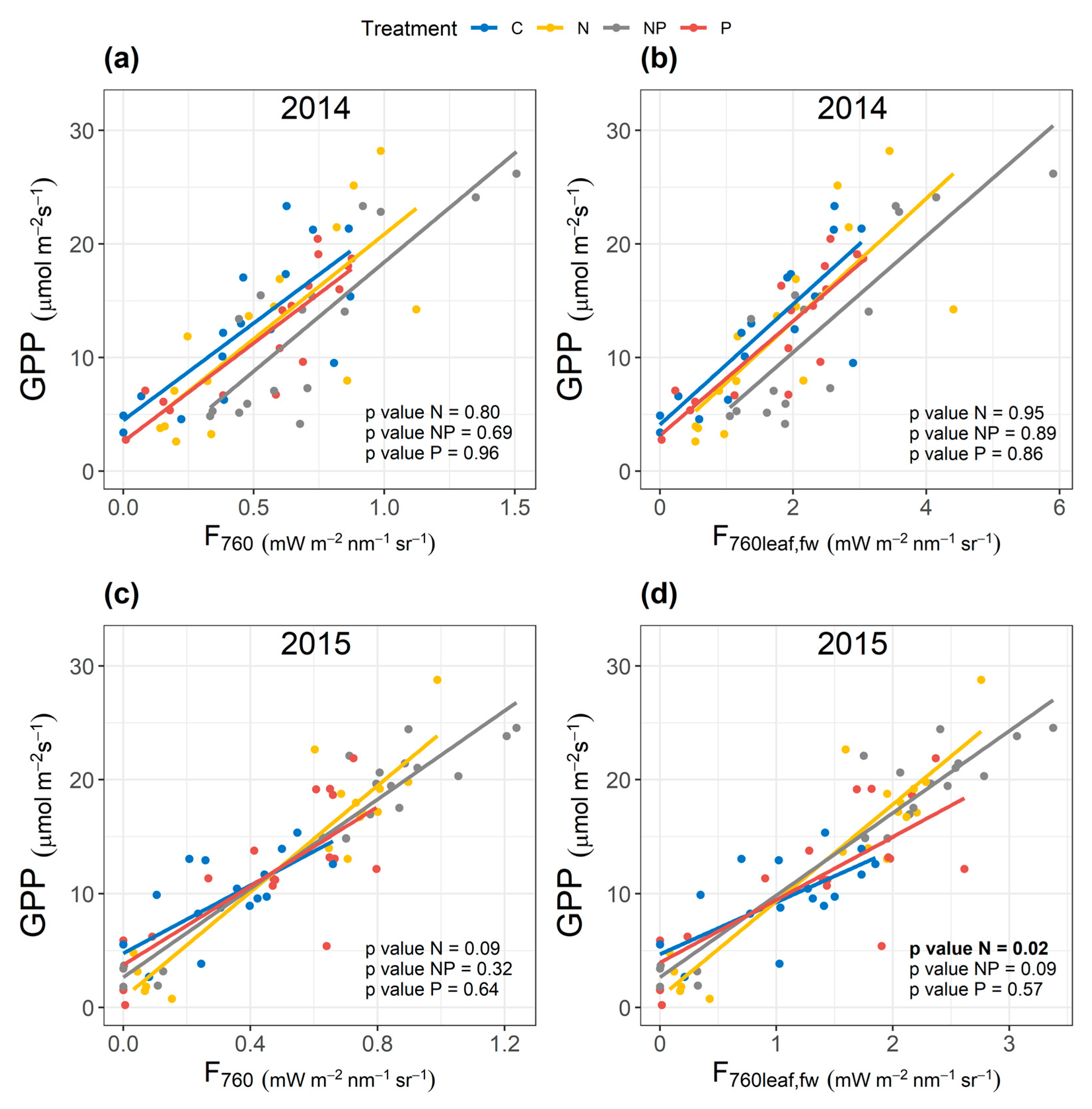
3.2. Temporal Variability of GPP–F760 and GPP–F760leaf.fw Relationship among Treatments
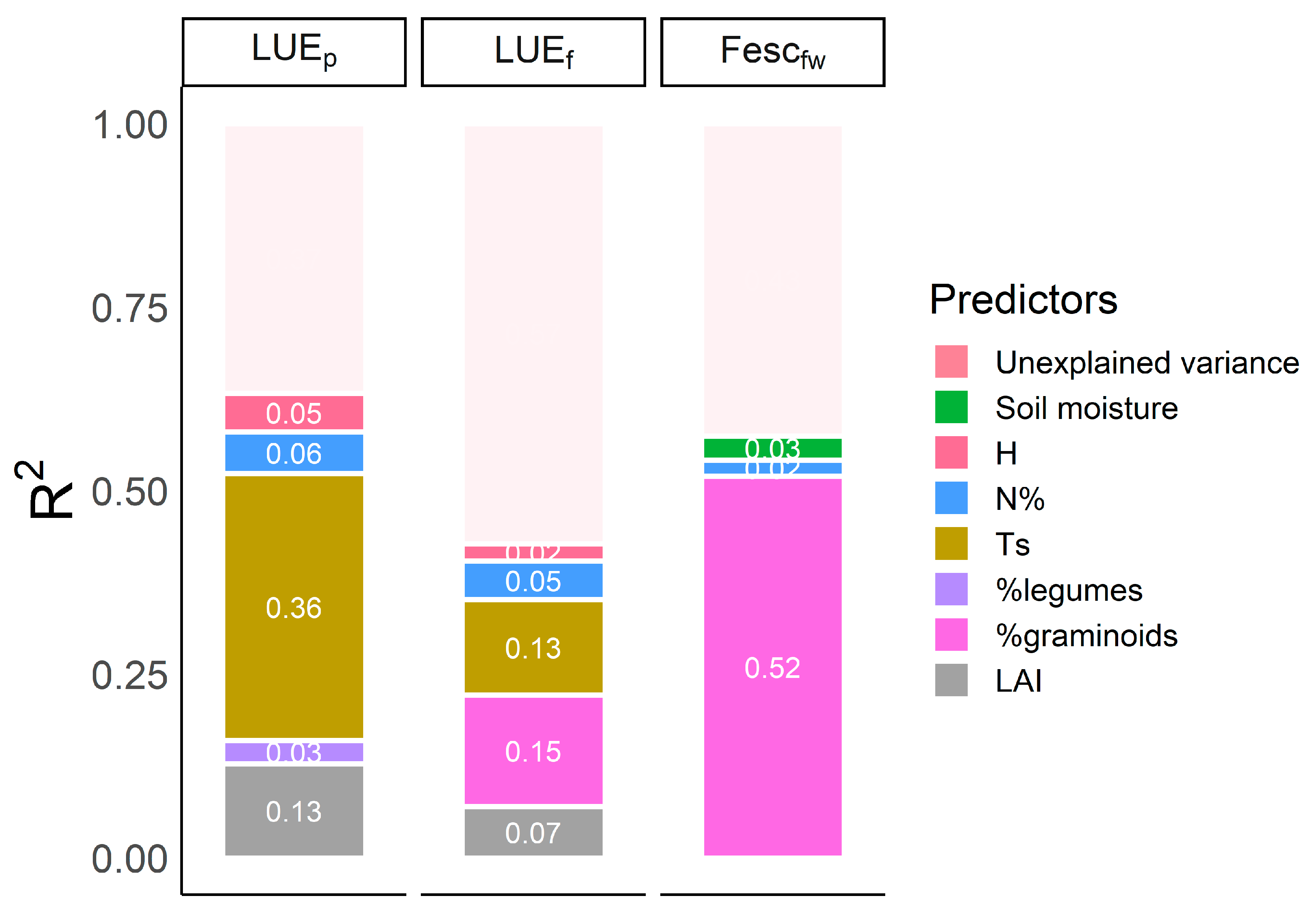
3.3. Factors Controlling the Parameters of Light Use Efficiency Equation (LUEp, LUEf and Fesc)
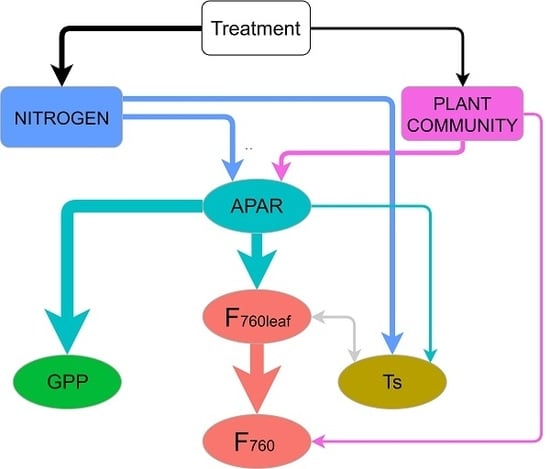
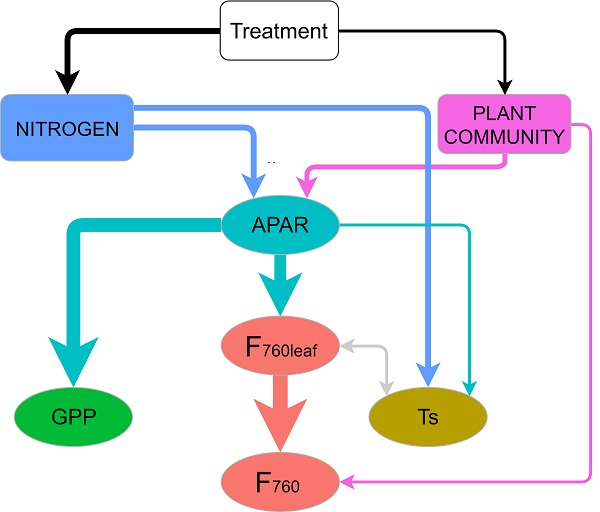
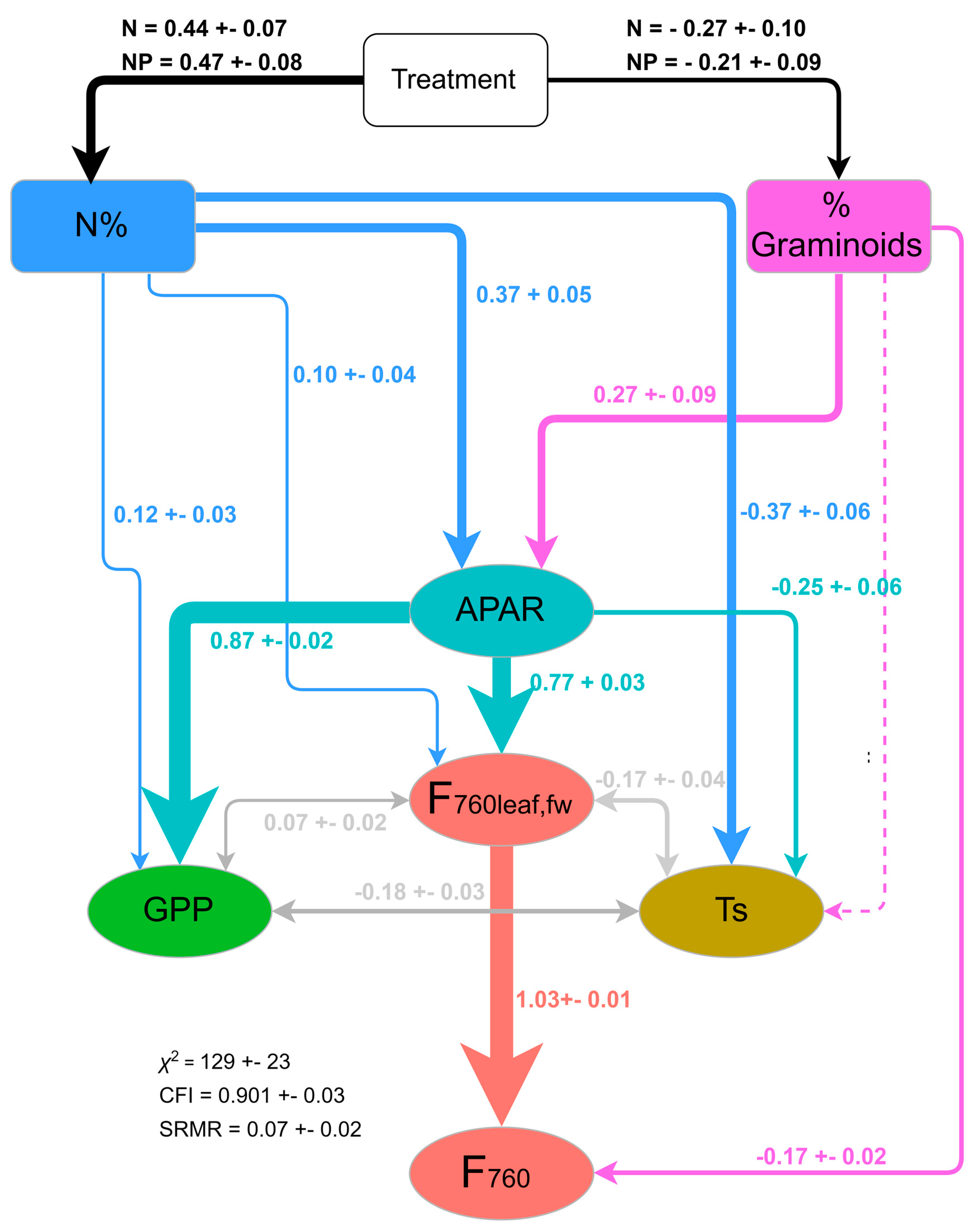
3.4. Mechanisms behind the Treatment Effect on GPP and F760 at Leaf and Canopy Scale
4. Discussion
4.1. Treatment Effect on LUEp, LUEf, Fescfw
4.2. Predictors of the Terms of the Light Use Efficiency Equation
4.3. Mechanisms behind the Treatment Effect on GPP and F760 at Leaf and Canopy Scale
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rodenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Monteith, J. Solar radiation and productivity in tropical ecosystems. J. Appl. Ecol. 1972, 9, 747–766. [Google Scholar] [CrossRef]
- Guanter, L.; Zhang, Y.G.; Jung, M.; Joiner, J.; Voigt, M.; Berry, J.A.; Frankenberg, C.; Huete, A.R.; Zarco-Tejada, P.; Lee, J.E.; et al. Global and time-resolved monitoring of crop photosynthesis with chlorophyll fluorescence. Proc. Natl. Acad. Sci. USA 2014, 111, E1327–E1333. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Tang, J.; Mustard, J.F.; Lee, J.E.; Rossini, M.; Joiner, J.; Munger, J.W.; Kornfeld, A.; Richardson, A.D. Solar--induced chlorophyll fluorescence that correlates with canopy photosynthesis on diurnal and seasonal scales in a temperate deciduous forest. Geophys. Res. Lett. 2015, 42, 2977–2987. [Google Scholar] [CrossRef]
- Zhang, Y.; Guanter, L.; Berry, J.A.; Joiner, J.; van der Tol, C.; Huete, A.; Gitelson, A.; Voigt, M.; Köhler, P. Estimation of vegetation photosynthetic capacity from space--based measurements of chlorophyll fluorescence for terrestrial biosphere models. Glob. Chang. Biol. 2014, 20, 3727–3742. [Google Scholar] [CrossRef]
- Meroni, M.; Rossini, M.; Guanter, L.; Alonso, L.; Rascher, U.; Colombo, R.; Moreno, J. Remote sensing of solar-induced chlorophyll fluorescence: Review of methods and applications. Remote Sens. Environ. 2009, 113, 2037–2051. [Google Scholar] [CrossRef]
- Damm, A.; Elbers, J.; Erler, A.; Gioli, B.; Hamdi, K.; Hutjes, R.; Kosvancova, M.; Meroni, M.; Miglietta, F.; Moersch, A. Remote sensing of sun--induced fluorescence to improve modeling of diurnal courses of gross primary production (gpp). Glob. Chang. Biol. 2010, 16, 171–186. [Google Scholar] [CrossRef]
- Migliavacca, M.; Perez-Priego, O.; Rossini, M.; El-Madany, T.S.; Moreno, G.; van der Tol, C.; Rascher, U.; Berninger, A.; Bessenbacher, V.; Burkart, A.; et al. Plant functional traits and canopy structure control the relationship between photosynthetic co2 uptake and far-red sun-induced fluorescence in a mediterranean grassland under different nutrient availability. New Phytol. 2017, 214, 1078–1091. [Google Scholar] [CrossRef]
- Rossini, M.; Meroni, M.; Migliavacca, M.; Manca, G.; Cogliati, S.; Busetto, L.; Picchi, V.; Cescatti, A.; Seufert, G.; Colombo, R. High resolution field spectroscopy measurements for estimating gross ecosystem production in a rice field. Agr. For. Meteorol. 2010, 150, 1283–1296. [Google Scholar] [CrossRef]
- Wieneke, S.; Ahrends, H.; Damm, A.; Pinto, F.; Stadler, A.; Rossini, M.; Rascher, U. Airborne based spectroscopy of red and far-red sun-induced chlorophyll fluorescence: Implications for improved estimates of gross primary productivity. Remote Sens. Environ. 2016, 184, 654–667. [Google Scholar] [CrossRef]
- Guan, K.; Berry, J.A.; Zhang, Y.; Joiner, J.; Guanter, L.; Badgley, G.; Lobell, D.B. Improving the monitoring of crop productivity using spaceborne solar--induced fluorescence. Glob. Chang. Biol. 2016, 22, 716–726. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-E.; Frankenberg, C.; van der Tol, C.; Berry, J.A.; Guanter, L.; Boyce, C.K.; Fisher, J.B.; Morrow, E.; Worden, J.R.; Asefi, S. Forest productivity and water stress in amazonia: Observations from gosat chlorophyll fluorescence. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130171. [Google Scholar] [CrossRef] [PubMed]
- Parazoo, N.C.; Bowman, K.; Fisher, J.B.; Frankenberg, C.; Jones, D.B.; Cescatti, A.; Pérez--Priego, Ó.; Wohlfahrt, G.; Montagnani, L. Terrestrial gross primary production inferred from satellite fluorescence and vegetation models. Glob. Chang. Biol. 2014, 20, 3103–3121. [Google Scholar] [CrossRef]
- Ač, A.; Malenovský, Z.; Olejníčková, J.; Gallé, A.; Rascher, U.; Mohammed, G. Meta-analysis assessing potential of steady-state chlorophyll fluorescence for remote sensing detection of plant water, temperature and nitrogen stress. Remote Sens. Environ. 2015, 168, 420–436. [Google Scholar] [CrossRef]
- Cendrero-Mateo, M.P.; Moran, M.S.; Papuga, S.A.; Thorp, K.; Alonso, L.; Moreno, J.; Ponce-Campos, G.; Rascher, U.; Wang, G. Plant chlorophyll fluorescence: Active and passive measurements at canopy and leaf scales with different nitrogen treatments. J. Exp. Bot. 2015, 67, 275–286. [Google Scholar] [CrossRef]
- Bilger, W.; Bjorkman, O. Temperature-dependence of violaxanthin deepoxidation and nonphotochemical fluorescence quenching in intact leaves of gossypium-hirsutum l and malva-parviflora l. Planta 1991, 184, 226–234. [Google Scholar] [CrossRef]
- Govindjee. 63 years since kautsky - chlorophyll-a fluorescence. Aust. J. Plant Physiol. 1995, 22, 131–160. [Google Scholar]
- Porcar-Castell, A.; Tyystjarvi, E.; Atherton, J.; van der Tol, C.; Flexas, J.; Pfundel, E.E.; Moreno, J.; Frankenberg, C.; Berry, J.A. Linking chlorophyll a fluorescence to photosynthesis for remote sensing applications: Mechanisms and challenges. J. Exp. Bot. 2014, 65, 4065–4095. [Google Scholar] [CrossRef]
- van der Tol, C.; Berry, J.A.; Campbell, P.K.E.; Rascher, U. Models of fluorescence and photosynthesis for interpreting measurements of solar-induced chlorophyll fluorescence. J. Geophys. Res. Biogeosci. 2014, 119, 2312–2327. [Google Scholar]
- Damm, A.; Guanter, L.; Paul-Limoges, E.; van der Tol, C.; Hueni, A.; Buchmann, N.; Eugster, W.; Ammann, C.; Schaepman, M.E. Far-red sun-induced chlorophyll fluorescence shows ecosystem-specific relationships to gross primary production: An assessment based on observational and modeling approaches. Remote Sens. Environ. 2015, 166, 91–105. [Google Scholar] [CrossRef]
- Grime, J.P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences. J. Veg. Sci. 2006, 17, 255–260. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Kull, O.; Tenhunen, J.D. Variability in leaf morphology and chemical composition as a function of canopy light environment in coexisting deciduous trees. Int. J. Plant Sci. 1999, 160, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of c 3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Houborg, R.; Cescatti, A.; Migliavacca, M.; Kustas, W. Satellite retrievals of leaf chlorophyll and photosynthetic capacity for improved modeling of gpp. Agr. For. Meteorol. 2013, 177, 10–23. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.v.; Berry, J. A biochemical model of photosynthetic co 2 assimilation in leaves of c 3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, A.S.; Demmig-Adams, B.; Adams III, W.W. Enhanced employment of the xanthophyll cycle and thermal energy dissipation in spinach exposed to high light and n stress. Plant Physiol. 1997, 113, 817–824. [Google Scholar] [CrossRef]
- Jiang, M.; Caldararu, S.; Zaehle, S.; Ellsworth, D.S.; Medlyn, B.E. Towards a more physiological representation of vegetation phosphorus processes in land surface models. New Phytol. 2019, 222, 1223–1229. [Google Scholar] [CrossRef]
- Singh, S.; Reddy, V.; Fleisher, D.; Timlin, D. Relationship between photosynthetic pigments and chlorophyll fluorescence in soybean under varying phosphorus nutrition at ambient and elevated co 2. Photosynthetica 2017, 55, 421–433. [Google Scholar] [CrossRef]
- Wohlfahrt, G.; Bahn, M.; Tappeiner, U.; Cernusca, A. A multi-component, multi-species model of vegetation-atmosphere co2 and energy exchange for mountain grasslands. Agr. For. Meteorol. 2001, 106, 261–287. [Google Scholar] [CrossRef]
- Porcar-Castell, A.; Juurola, E.; Ensminger, I.; Berninger, F.; Hari, P.; Nikinmaa, E. Seasonal acclimation of photosystem ii in pinus sylvestris. Ii. Using the rate constants of sustained thermal energy dissipation and photochemistry to study the effect of the light environment. Tree Physiol. 2008, 28, 1483–1491. [Google Scholar] [CrossRef]
- Yang, P.Q.; van der Tol, C. Linking canopy scattering of far-red sun-induced chlorophyll fluorescence with reflectance. Remote Sens. Environ. 2018, 209, 456–467. [Google Scholar] [CrossRef]
- Liu, X.; Guanter, L.; Liu, L.; Damm, A.; Malenovský, Z.; Rascher, U.; Peng, D.; Du, S.; Gastellu-Etchegorry, J.-P. Downscaling of solar-induced chlorophyll fluorescence from canopy level to photosystem level using a random forest model. Remote Sens. Environ. 2018, 231, 110772. [Google Scholar] [CrossRef]
- Zeng, Y.; Badgley, G.; Dechant, B.; Ryu, Y.; Chen, M.; Berry, J.A. A practical approach for estimating the escape ratio of near-infrared solar-induced chlorophyll fluorescence. Remote Sens. Environ. 2019, 232, 111209. [Google Scholar] [CrossRef]
- Galmés, J.; Ribas-Carbó, M.; Medrano, H.; Flexas, J. Response of leaf respiration to water stress in mediterranean species with different growth forms. J. Arid Environ. 2007, 68, 206–222. [Google Scholar] [CrossRef]
- Alonso, L.; Van Wittenberghe, S.; Amorós-López, J.; Vila-Francés, J.; Gómez-Chova, L.; Moreno, J. Diurnal cycle relationships between passive fluorescence, pri and npq of vegetation in a controlled stress experiment. Remote Sens. 2017, 9, 770. [Google Scholar] [CrossRef]
- Hilker, T.; Coops, N.C.; Hall, F.G.; Black, T.A.; Wulder, M.A.; Nesic, Z.; Krishnan, P. Separating physiologically and directionally induced changes in pri using brdf models. Remote Sens. Environ. 2008, 112, 2777–2788. [Google Scholar] [CrossRef]
- Perez-Priego, O.; Guan, J.; Rossini, M.; Fava, F.; Wutzler, T.; Moreno, G.; Carvalhais, N.; Carrara, A.; Kolle, O.; Julitta, T.; et al. Sun-induced chlorophyll fluorescence and photochemical reflectance index improve remote-sensing gross primary production estimates under varying nutrient availability in a typical mediterranean savanna ecosystem. Biogeosciences 2015, 12, 6351–6367. [Google Scholar] [CrossRef]
- Luo, Y.; El-Madany, T.; Filippa, G.; Ma, X.; Ahrens, B.; Carrara, A.; Gonzalez-Cascon, R.; Cremonese, E.; Galvagno, M.; Hammer, T. Using near-infrared-enabled digital repeat photography to track structural and physiological phenology in mediterranean tree–grass ecosystems. Remote Sens. 2018, 10, 1293. [Google Scholar] [CrossRef]
- Spellerberg, I.F.; Fedor, P.J. A tribute to claude shannon (1916-2001) and a plea for more rigorous use of species richness, species diversity and the ‘shannon-wiener’ index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef]
- Brand, W.A.; Coplen, T.B. Stable isotope deltas: Tiny, yet robust signatures in nature. Isot. Environ. Health Stud. 2012, 48, 393–409. [Google Scholar] [CrossRef]
- Coplen, T.B. Guidelines and recommended terms for expression of stable-isotope-ratio and gas-ratio measurement results. Rapid Commun. Mass Spectrom. 2011, 25, 2538–2560. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Labrador, J.; Perez-Priego, O.; El-Madany, T.S.; Julitta, T.; Rossini, M.; Guan, J.; Moreno, G.; Carvalhais, N.; Martín, M.P.; Gonzalez-Cascon, R. Multiple-constraint inversion of scope. Evaluating the potential of gpp and sif for the retrieval of plant functional traits. Remote Sens. Environ. 2019, 234, 111362. [Google Scholar] [CrossRef]
- Seibt, U.; Rajabi, A.; Griffiths, H.; Berry, J.A. Carbon isotopes and water use efficiency: Sense and sensitivity. Oecologia 2008, 155, 441–454. [Google Scholar] [CrossRef] [PubMed]
- van der Tol, C.; Verhoef, W.; Timmermans, J.; Verhoef, A.; Su, Z. An integrated model of soil-canopy spectral radiances, photosynthesis, fluorescence, temperature and energy balance. Biogeosciences 2009, 6, 3109–3129. [Google Scholar] [CrossRef]
- Vina, A.; Gitelson, A.A. New developments in the remote estimation of the fraction of absorbed photosynthetically active radiation in crops. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Li, Z.; Moreau, L. A new approach for remote sensing of canopy-absorbed photosynthetically active radiation. I: Total surface absorption. Remote Sens. Environ. 1996, 55, 175–191. [Google Scholar] [CrossRef]
- Moreau, L.; Li, Z. A new approach for remote sensing of canopy absorbed photosynthetically active radiation. Ii: Proportion of canopy absorption. Remote Sens. Environ. 1996, 55, 192–204. [Google Scholar] [CrossRef]
- Sager, J.; McFarlane, J. Plant growth chamber handbook. Radiation 1997, 1–29. [Google Scholar]
- Moder, K. Alternatives to f-test in one way anova in case of heterogeneity of variances (a simulation study). Psychol. Test. Assess. Model. 2010, 52, 343–353. [Google Scholar]
- Games, P.A.; Howell, J.F. Pairwise multiple comparison procedures with unequal n’s and/or variances: A monte carlo study. J. Educ. Stat. 1976, 1, 113–125. [Google Scholar]
- Groemping, U.; Matthias, L. Relaimpo: Relative Importance of Regressors in Linear Models; R Package Version 1.1-1; Foundation for Open Access Statistics: Los Angeles, CA, USA, 2006. [Google Scholar]
- Sumayao, C.; Kanemasu, E.; Brakke, T. Using leaf temperature to assess evapotranspiration and advection. Agric. Meteorol. 1980, 22, 153–166. [Google Scholar] [CrossRef]
- Rosseel, Y. Lavaan: An r package for structural equation modeling and more. Version 0.5–12 (beta). J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Weis, E.; Berry, J.A. Plants and high temperature stress. Symp. Soc. Exp. Biol. 1988, 42, 329–346. [Google Scholar] [PubMed]
- Hu, L.t.; Bentler, P.M. Cutoff criteria for fit indexes in covariance structure analysis: Conventional criteria versus new alternatives. Struct. Equ. Model. A Multidiscip. J. 1999, 6, 1–55. [Google Scholar] [CrossRef]
- Iacobucci, D. Structural equations modeling: Fit indices, sample size, and advanced topics. J. Consum. Psychol. 2010, 20, 90–98. [Google Scholar] [CrossRef]
- Singh, S.K.; Reddy, V.R. Combined effects of phosphorus nutrition and elevated carbon dioxide concentration on chlorophyll fluorescence, photosynthesis, and nutrient efficiency of cotton. J. Plant Nutr. Soil Sci. 2014, 177, 892–902. [Google Scholar] [CrossRef]
- Tatarko, A.R.; Knops, J.M.H. Nitrogen addition and ecosystem functioning: Both species abundances and traits alter community structure and function. Ecosphere 2018, 9, e02087. [Google Scholar] [CrossRef]
- Jackson, R.D.; Idso, S.; Reginato, R.; Pinter, P. Canopy temperature as a crop water stress indicator. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Boulet, G.; Chehbouni, A.; Gentine, P.; Duchemin, B.; Ezzahar, J.; Hadria, R. Monitoring water stress using time series of observed to unstressed surface temperature difference. Agr. For. Meteorol. 2007, 146, 159–172. [Google Scholar] [CrossRef]
- Diaz, M.; de Haro, V.; Munoz, R.; Quiles, M.J. Chlororespiration is involved in the adaptation of brassica plants to heat and high light intensity. Plant Cell Environ. 2007, 30, 1578–1585. [Google Scholar]
- Sims, D.A.; Rahman, A.F.; Cordova, V.D.; El-Masri, B.Z.; Baldocchi, D.D.; Bolstad, P.V.; Flanagan, L.B.; Goldstein, A.H.; Hollinger, D.Y.; Misson, L. A new model of gross primary productivity for north american ecosystems based solely on the enhanced vegetation index and land surface temperature from modis. Remote Sens. Environ. 2008, 112, 1633–1646. [Google Scholar] [CrossRef]
- Weisser, W.W.; Roscher, C.; Meyer, S.T.; Ebeling, A.; Luo, G.; Allan, E.; Beßler, H.; Barnard, R.L.; Buchmann, N.; Buscot, F. Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: Patterns, mechanisms, and open questions. Basic Appl. Ecol. 2017, 23, 1–73. [Google Scholar] [CrossRef]
- Feng, X.H.; Dietze, M. Scale dependence in the effects of leaf ecophysiological traits on photosynthesis: Bayesian parameterization of photosynthesis models. New Phytol. 2013, 200, 1132–1144. [Google Scholar] [CrossRef] [PubMed]
- Frankenberg, C.; Berry, J. Solar induced chlorophyll fluorescence: Origins, relation to photosynthesis and retrieval. In Comprehensive Remote Sensing; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Peng, Y.; Gitelson, A.A.; Keydan, G.; Rundquist, D.C.; Moses, W. Remote estimation of gross primary production in maize and support for a new paradigm based on total crop chlorophyll content. Remote Sens. Environ. 2011, 115, 978–989. [Google Scholar] [CrossRef]
- Buschmann, C. Variability and application of the chlorophyll fluorescence emission ratio red/far-red of leaves. Photosynth. Res. 2007, 92, 261–271. [Google Scholar] [CrossRef]
- Knyazikhin, Y.; Schull, M.A.; Stenberg, P.; Mõttus, M.; Rautiainen, M.; Yang, Y.; Marshak, A.; Carmona, P.L.; Kaufmann, R.K.; Lewis, P. Hyperspectral remote sensing of foliar nitrogen content. Proc. Nat. Acad. Sci. USA 2013, 110, E185–E192. [Google Scholar] [CrossRef]
- Ollinger, S.V.; Richardson, A.D.; Martin, M.E.; Hollinger, D.Y.; Frolking, S.E.; Reich, P.B.; Plourde, L.C.; Katul, G.G.; Munger, J.W.; Oren, R. Canopy nitrogen, carbon assimilation, and albedo in temperate and boreal forests: Functional relations and potential climate feedbacks. Proc. Nat. Acad. Sci. USA 2008, 105, 19336–19341. [Google Scholar] [CrossRef]
- Migliavacca, M.; El-Madany, T.S.; Perez-Priego, O.; Carrara, A.; Gonzalez-Cascon, R.; Martin Isabel, M.P.; Moreno, G.; Guan, J.; Hammer, T.W.; Henkel, K.; et al. Effects of a large scale stoichiometric imbalance manipulation on the ecosystem functioning of a Mediterranean tree-grass ecosystem: The MANIP experiment. In AGU Fall Meeting Abstracts; American Geophysical Union: Washington, DC, USA, 2018. [Google Scholar]
- Ollinger, S.; Frolking, S.; Richardson, A.; Martin, M.; Hollinger, D.; Reich, P.; Plourde, L. Reply to fisher: Nitrogen–albedo relationship in forests remains robust and thought-provoking. Proc. Nat. Acad. Sci. USA 2009, 106, E17. [Google Scholar] [CrossRef]
- Sullivan, F.B.; Ollinger, S.V.; Martin, M.E.; Ducey, M.J.; Lepine, L.C.; Wicklein, H.F. Foliar nitrogen in relation to plant traits and reflectance properties of new hampshire forests. Can. J. For. Res. 2012, 43, 18–27. [Google Scholar] [CrossRef]
- Sun, Z.; Livingston, N.; Guy, R.; Ethier, G. Stable carbon isotopes as indicators of increased water use efficiency and productivity in white spruce (picea glauca (moench) voss) seedlings. Plant Cell Environ. 1996, 19, 887–894. [Google Scholar]
- Vilfan, N.; van der Tol, C.; Verhoef, W. Estimating photosynthetic capacity from leaf reflectance and chlorophyll fluorescence by coupling radiative transfer to a model for photosynthesis. New Phytol. 2019, 223, 487–500. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Campaign | Fertilization | PAR μmol s−1 m−2 | VPD hPa | Ta °C | SWC % | SZA ° |
|---|---|---|---|---|---|---|---|
| 20-03-2014 | 1 | No | 1604.82 ± 11.33 | 12.59 ± 0.38 | 24.2 ± 0.2 | 19.01 ± 0.27 | 41.86 ± 0.23 |
| 15-04-2014 | 2 | Yes | 1842.92 ± 32.63 | 15.12 ± 0.59 | 30.09 ± 0.55 | 22.58 ± 0.58 | 31.83 ± 0.85 |
| 7-05-2014 | 3 | Yes | 1342.1 ± 93.73 | 22.4 ± 1.98 | 32.1 ± 0.91 | 4.78 ± 0.09 | 25.69 ± 0.6 |
| 27-05-2014 | 4 | Yes | 1417.15 ± 104.4 | 15.83 ± 1.2 | 27.89 ± 0.47 | 6.57 ± 0.09 | 21.4 ± 0.82 |
| 04-03-2015 | 5 | Yes | 1411.29 ± 18.05 | 7.01 ± 0.36 | 23.9 ± 0.48 | 21.49 ± 1.91 | 49.66 ± 0.49 |
| 23-04-2015 | 6 | Yes | 1842.64 ± 25.23 | 16.38 ± 0.84 | 29.98 ± 0.37 | 6.7 ± 0.11 | 31.21 ± 0.98 |
| 27-05-2015 | 7 | Yes | 1955.21 ± 35.25 | 23.2 ± 1.56 | 36.33 ± 0.73 | 1.14 ± 0.02 | 24.26 ± 1.87 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martini, D.; Pacheco-Labrador, J.; Perez-Priego, O.; van der Tol, C.; El-Madany, T.S.; Julitta, T.; Rossini, M.; Reichstein, M.; Christiansen, R.; Rascher, U.; et al. Nitrogen and Phosphorus Effect on Sun-Induced Fluorescence and Gross Primary Productivity in Mediterranean Grassland. Remote Sens. 2019, 11, 2562. https://doi.org/10.3390/rs11212562
Martini D, Pacheco-Labrador J, Perez-Priego O, van der Tol C, El-Madany TS, Julitta T, Rossini M, Reichstein M, Christiansen R, Rascher U, et al. Nitrogen and Phosphorus Effect on Sun-Induced Fluorescence and Gross Primary Productivity in Mediterranean Grassland. Remote Sensing. 2019; 11(21):2562. https://doi.org/10.3390/rs11212562
Chicago/Turabian StyleMartini, David, Javier Pacheco-Labrador, Oscar Perez-Priego, Christiaan van der Tol, Tarek S. El-Madany, Tommaso Julitta, Micol Rossini, Markus Reichstein, Rune Christiansen, Uwe Rascher, and et al. 2019. "Nitrogen and Phosphorus Effect on Sun-Induced Fluorescence and Gross Primary Productivity in Mediterranean Grassland" Remote Sensing 11, no. 21: 2562. https://doi.org/10.3390/rs11212562
APA StyleMartini, D., Pacheco-Labrador, J., Perez-Priego, O., van der Tol, C., El-Madany, T. S., Julitta, T., Rossini, M., Reichstein, M., Christiansen, R., Rascher, U., Moreno, G., Martín, M. P., Yang, P., Carrara, A., Guan, J., González-Cascón, R., & Migliavacca, M. (2019). Nitrogen and Phosphorus Effect on Sun-Induced Fluorescence and Gross Primary Productivity in Mediterranean Grassland. Remote Sensing, 11(21), 2562. https://doi.org/10.3390/rs11212562