Projecting Mammal Distributions in Response to Future Alternative Landscapes in a Rapidly Transitioning Region

Abstract

1. Introduction
2. Materials and Methods
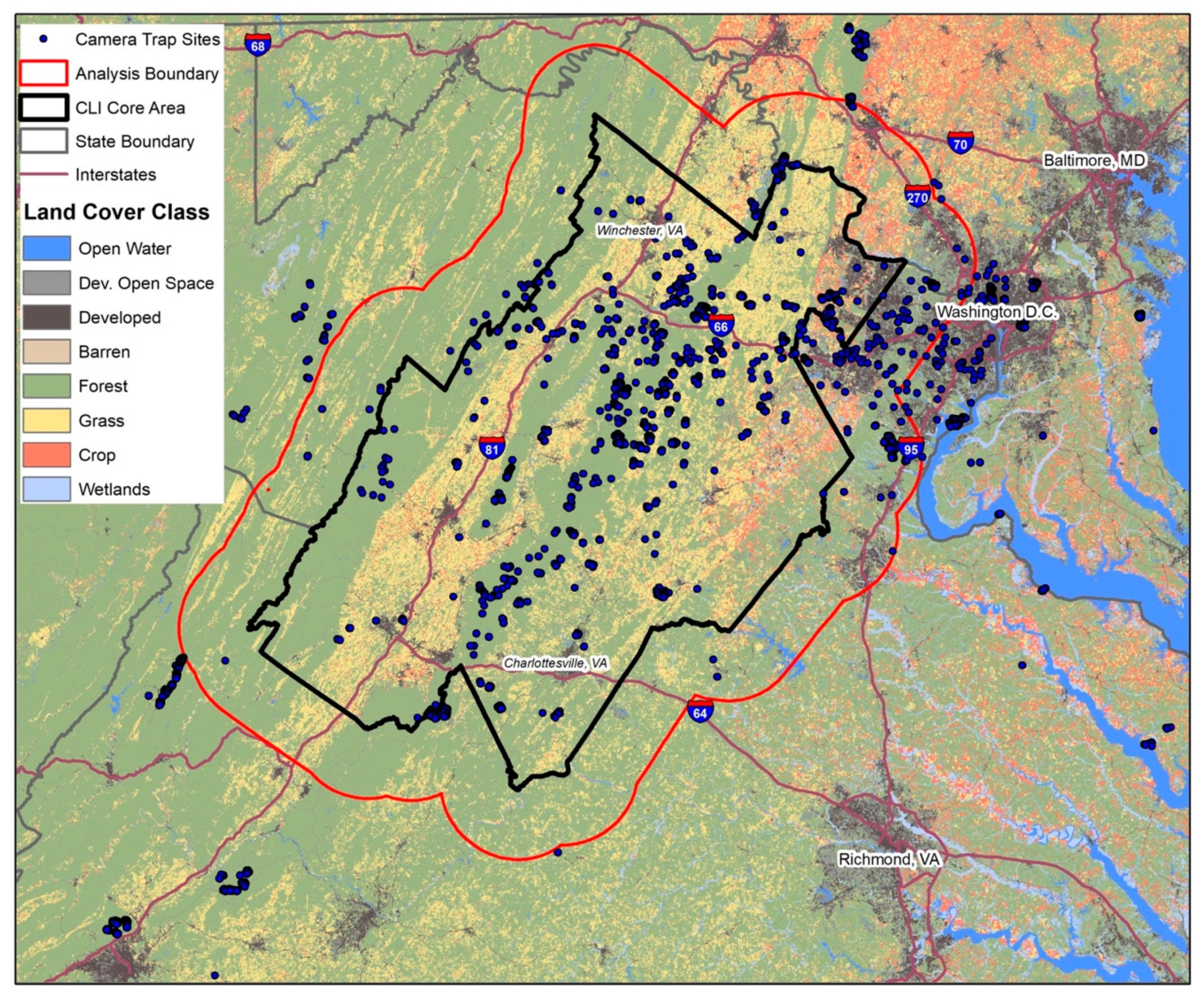
2.1. Study Area
2.2. Mammal Camera Trapping
2.3. Covariates and Models
2.4. Model Selection
2.5. Predicting under Land-Planning Scenarios
2.6. Species’ Habitat Projections
3. Results
3.1. Camera Trap Summary Statistics
3.2. Occupancy Model Results and Habitat Associations
3.3. Predicted Contemporary Distributions
3.4. Projected Land Cover and Suitable Habitat Availability Changes under Different Scenarios
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CLI Land Cover Class | NLCD Land Cover Class |
|---|---|
| Open Water | Open Water |
| Developed Open Space | Developed Open Space |
| Developed | Developed, Low Intensity |
| Developed, Medium Intensity | |
| Developed, High Intensity | |
| Barren | Barren Land |
| Forest | Deciduous Forest |
| Evergreen Forest | |
| Mixed Forest | |
| Shrub/Scrub | |
| Grass | Grassland/Herbaceous |
| Pasture/Hay | |
| Crop | Cultivated Crops |
| Wetland | Woody Wetlands |
| Emergent Herbaceous Wetlands |
References
- Kareiva, P.; Marvier, M. What is conservation science? Bioscience 2012, 62, 962–969. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef] [PubMed]
- Schipper, J.; Chanson, J.S.; Chiozza, F.; Cox, N.A.; Hoffmann, M.; Katariya, V.; Lamoreux, J.; Rodrigues, A.S.L.; Stuart, S.N.; Temple, H.J.; et al. The status of the world’s land and marine mammals: diversity, threat, and knowledge. Science 2008, 322, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Gaynor, K.M.; Hojnowski, C.E.; Carter, N.H.; Brashares, J.S. The influence of human disturbance on wildlife nocturnality. Science 2018, 360, 1232–1235. [Google Scholar] [CrossRef] [PubMed]
- Tucker, M.A.; Bohning-Gaese, K.; Fagan, W.F.; Fryxell, J.M.; Van Moorter, B.; Alberts, S.C.; Ali, A.H.; Allen, A.M.; Attias, N.; Avgar, T.; et al. Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science 2018, 359, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.T.; Burton, A.C. Wildlife winners and losers in an oil sands landscape. Front. Ecol. Environ. 2018, 16, 323–328. [Google Scholar] [CrossRef]
- Lesmeister, D.B.; Nielsen, C.K.; Schauber, E.M.; Hellgren, E.C. Spatial and temporal structure of a mesocarnivore guild in Midwestern North America. Wildl. Monograpghs 2015, 191, 1–61. [Google Scholar] [CrossRef]
- Morin, D.J.; Lesmeister, D.B.; Nielsen, C.K.; Schauber, E.M. The truth about cats and dogs: Landscape composition and human occupation mediate the distribution and potential impact of non-native carnivores. Glob. Ecol. Conserv. 2018, 15, e00413. [Google Scholar] [CrossRef]
- Pardo Vargas, L.E.; Cove, M.V.; Spinola, R.M.; de la Cruz, J.C.; Saenz, J.C. Assessing species traits and landscape relationships of the mammalian carnivore community in a neotropical biological corridor. Biodivers. Conserv. 2016, 25, 739–752. [Google Scholar] [CrossRef]
- Cove, M.V.; Jones, B.M.; Bossert, A.J.; Clever Jr, D.R.; Dunwoody, R.K.; White, B.C.; Jackson, V.L. Use of camera traps to examine the mesopredator release hypothesis in a fragmented Midwestern landscape. Am. Midl. Nat. 2012, 168, 456–465. [Google Scholar] [CrossRef]
- Goad, E.H.; Pejchar, L.; Reed, S.E.; Knight, R.L. Habitat use by mammals varies along an exurban development gradient in northern Colorado. Biol. Conserv. 2014, 176, 172–182. [Google Scholar] [CrossRef]
- Long, R.A.; Donovan, T.M.; Mackay, P.; Zielinski, W.J.; Buzas, J.S. Predicting carnivore occurrence with noninvasive surveys and occupancy modeling. Landsc. Ecol. 2011, 26, 327–340. [Google Scholar] [CrossRef]
- Ordeñana, M.A.; Crooks, K.R.; Boydston, E.E.; Lyren, L.M.; Siudyla, S.; Haas, C.D.; Harris, S.; Hathaway, S.A.; Turschak, G.M.; Miles, A.K.; et al. Effects of urbanization on carnivore species distribution and richness. J. Mamm. 2010, 91, 1322–1331. [Google Scholar] [CrossRef]
- Long, R.A.; MacKay, P.; Ray, J.; Zielinski, W. (Eds.) Noninvasive Survey Methods for Carnivores; Island Press: Washington, DC, USA, 2008. [Google Scholar]
- Steenweg, R.; Hebblewhite, M.; Kays, R.; Ahumada, J.; Fisher, J.T.; Burton, C.; Townsend, S.E.; Carbone, C.; Rowcliffe, J.M.; Whittington, J.; et al. Scaling-up camera traps: Monitoring the planet’s biodiversity with networks of remote sensors. Front. Ecol. Environ. 2017, 15, 26–34. [Google Scholar] [CrossRef]
- Ye, X.; Yu, X.; Yu, C.; Tayibazhaer, A.; Xu, F.; Skidmore, A.K.; Wang, T. Impacts of future climate and land cover changes on threatened mammals in the semi-arid Chinese Altai Mountains. Sci. Tot. Environ. 2018, 612, 775–787. [Google Scholar] [CrossRef]
- Lawler, J.J.; Shafer, S.L.; White, D.; Kareiva, P.; Maurer, E.P.; Blaustein, A.R.; Bartlein, P.J. Projected climate-induced faunal change in the Western Hemisphere. Ecology 2009, 90, 588–597. [Google Scholar] [CrossRef]
- Levinsky, I.; Skov, F.; Svenning, J.C.; Rahbek, C. Potential impacts of climate change on the distributions and diversity patterns of European mammals. Biodivers. Conserv. 2007, 16, 3803–3816. [Google Scholar] [CrossRef]
- Lacher, I.; Akre, T.; McShea, W.J.; McBride, M.; Thompson, J.R.; Fergus, C. Engaging regional stakeholders in scenario planning for the long-term preservation of ecosystem services in Northwestern Virginia. Case Stud. Environ. 2019. [Google Scholar] [CrossRef]
- Bennett, E.M.; Carpenter, S.R.; Peterson, G.D.; Cumming, G.S.; Zurek, M.; Pingali, P. Why global scenarios need ecology. Front. Ecol. Environ. 2003, 1, 322–329. [Google Scholar] [CrossRef]
- Peterson, G.D.; Cumming, G.S.; Carpenter, S.R. Scenario planning: a tool for conservation in an uncertain world. Conserv. Biol. 2003, 17, 358–366. [Google Scholar] [CrossRef]
- Lacher, I.L.; Ahmadisharaf, E.; Fergus, C.; Akre, T.; Mcshea, W.J.; Benham, B.L.; Kline, K.S. Scale-dependent impacts of urban and agricultural land use on nutrients, sediment, and runoff. Sci. Tot. Environ. 2019, 652, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Kays, R.; Parsons, A.W.; Baker, M.C.; Kalies, E.L.; Forrester, T.; Costello, R.; Rota, C.T.; Millspaugh, J.J.; Mcshea, W.J. Does hunting or hiking affect wildlife communities in protected areas? J. Appl. Ecol. 2017, 54, 242–252. [Google Scholar] [CrossRef]
- McShea, W.J.; Forrester, T.; Costello, R.; He, Z.; Kays, R. Volunteer-run cameras as distributed sensors for macrosystem mammal research. Landsc. Ecol. 2016, 31, 55–66. [Google Scholar] [CrossRef]
- Parsons, A.W.; Forrester, T.; Baker-Whatton, M.C.; McShea, W.J.; Rota, C.T.; Schuttler, S.G.; Millspaugh, J.J.; Kays, R. Mammal communities are larger and more diverse in moderately developed areas. ELife 2018, 7, e38012. [Google Scholar] [CrossRef]
- Mackenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modeling, 2nd ed.; Academic Press: Burlington, MA, USA, 2018. [Google Scholar]
- Homer, C.; Dewitz, J.; Yang, L.; Jin, S.; Danielson, P.; Xian, G. Completion of the 2011 National Land Cover Database for the conterminous United States–representing a decade of land cover change information. Photogramm. Eng. Remote Sens. 2015, 81, 345–354. [Google Scholar]
- Jones, B.M.; Cove, M.V.; Lashley, M.A.; Jackson, V.L. Do coyotes Canis latrans influence occupancy of prey in suburban forest fragments? Curr. Zool. 2016, 62, 1–6. [Google Scholar] [CrossRef]
- U.S. Geological Survey (USGS); Gap Analysis Project (GAP). Protected Areas Database of the United States (PAD-US). 2016. U.S. Geological Survey data release. Available online: https://maps.usgs.gov/padus/ (accessed on 1 April 2019).
- Lacher, I.; Akre, T.; McShea, W.J.; Fergus, C. Spatial and temporal patterns of public and private land protection within the Blue Ridge and Piedmont ecoregions of the eastern US. Landsc. Urban Plan. 2019, 186, 91–102. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A.; Ene, E. Fragstats v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps. Computer software program produced by the authors at the University of Massachusetts, Amherst. 2012. Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 1 April 2019).
- Fiske, I.; Chandler, R.; Miller, D.; Royle, J.A.; Kery, M.; Hostetler, J.; Hutchinson, R.; Smith, A.; Kellner, K. Package ‘Unmarked’. 2019. Available online: https://cran.r-project.org/web/packages/unmarked/unmarked.pdf (accessed on 24 October 2019).
- R Development Core Team. R: A Language and Environment for Statistical Computing. 2011. Available online: https://www.r-project.org/ (accessed on 24 October 2019).
- Sing, T.; Sander, O.; Beerenwinkel, N.; Lengauer, T. ROCR: visualizing classifier performance in R. Bioinformatics 2005, 21, 3940–3941. [Google Scholar] [CrossRef]
- Soares-Filho, B.S.; Cerqueira, G.C.; Pennachin, C.L. DINAMICA—a stochastic cellular automata model designed to simulate the landscape dynamics in an Amazonian colonization frontier. Ecol. Model. 2002, 154, 217–235. [Google Scholar] [CrossRef]
- Bradbury, M.; Peterson, M.N.; Liu, J. Long-term dynamics of household size and their environmental implications. Pop. Environ. 2014, 36, 73–84. [Google Scholar] [CrossRef]
- Available online: https://demographics.coopercenter.org/virginia-population-projections (accessed on 1 April 2019).
- Crooks, K.R. Relative sensitivities of mammalian carnivores to habitat fragmentation. Conserv. Biol. 2002, 16, 488–502. [Google Scholar] [CrossRef]
- Evans, M.J.; Rittenhouse, T.A.; Hawley, J.E.; Rego, P.W. Black bear recolonization patterns in a human-dominated landscape vary based on housing: New insights from spatially explicit density models. Landsc. Urban Plan. 2017, 162, 13–24. [Google Scholar] [CrossRef]
- Gray, C.L.; Hill, S.L.; Newbold, T.; Hudson, L.N.; Börger, L.; Contu, S.; Hoskins, A.J.; Ferrier, S.; Purvis, A.; Scharlemann, J.P.W. Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 2016, 7, 12306. [Google Scholar] [CrossRef] [PubMed]
- Cove, M.V.; Gardner, B.; Simons, T.R.; Kays, R.; O’Connell, A.F. Free-ranging cats (Felis catus) on public lands: estimating density, movement, activity and diet. Biol. Invasions. 2018, 20, 333–344. [Google Scholar] [CrossRef]
- Hernandez, S.M.; Loyd, K.A.T.; Newton, A.N.; Carswell, B.L.; Abernathy, K.J. The use of point-of-view cameras (Kittycams) to quantify predation by colony cats (Felis catus) on wildlife. Wildl. Res. 2018, 45, 357–365. [Google Scholar] [CrossRef]
- Loss, S.R.; Will, T.; Marra, P.P. The impact of free-ranging domestic cats on wildlife of the United States. Nat. Commun. 2013, 4, 1396. [Google Scholar] [CrossRef]
- Parsons, A.W.; Rota, C.T.; Forrester, T.; Baker-Whatton, M.C.; McShea, W.J.; Schuttler, S.G.; Millspaugh, J.J.; Kays, R. Urbanization focuses carnivore activity in remaining natural habitats, increasing species interactions. J. Appl. Ecol. 2019. [Google Scholar] [CrossRef]
- Kays, R.; Costello, R.; Forrester, T.; Baker, M.C.; Parsons, A.W.; Kalies, E.L.; Hess, G.; Millspaugh, J.J.; Mcshea, W. Cats are rare where coyotes roam. J. Mamm. 2015, 96, 981–987. [Google Scholar] [CrossRef]
- Bragina, E.V.; Kays, R.; Hody, A.; Moorman, C.E.; Deperno, C.S.; Mills, L.S. Effects on white-tailed deer following eastern coyote colonization. J. Wildl. Manag. 2019. [Google Scholar] [CrossRef]
- Cove, M.V.; Niva, L.M.; Jackson, V.L. Use of probability of detection when conducting analyses of surveys of mesopredators: A Case Study from the Ozark Highlands of Missouri. Southwest Nat. 2012, 57, 257–262. [Google Scholar] [CrossRef]
- Demeny, K.; McLoon, M.; Winesett, B.; Fastner, J.; Hammerer, E.; Pauli, J.N. Food subsidies of Raccoons in anthropogenic landscapes. Can. J. Zool. 2019. [Google Scholar] [CrossRef]
- Kays, R.; Parsons, A.W. Mammals in and around suburban yards, and the attraction of chicken coops. Urban Ecosyst. 2014, 17, 691–705. [Google Scholar] [CrossRef]
- Moritz, C.; Patton, J.L.; Conroy, C.J.; Parra, J.L.; White, G.C.; Beissinger, S.R. Impact of a century of climate change on small-mammal communities in Yosemite National Park, USA. Science 2008, 322, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.J.; Thorne, J.H.; Moritz, C. Synchronicity in elevation range shifts among small mammals and vegetation over the last century is stronger for omnivores. Ecography 2015, 38, 556–568. [Google Scholar] [CrossRef]




| Species | β | SE | LCI | UCI | p-Value | a priori |
|---|---|---|---|---|---|---|
| Model | ||||||
| Parameter | ||||||
| American Black Bear | ||||||
| Occupancy | ||||||
| Intercept | −0.80 | 0.17 | −1.13 | −0.46 | 0.00 | |
| Forest 1000 | 1.73 | 0.22 | 1.30 | 2.16 | 0.00 | + |
| Grass 1000 | 1.19 | 0.15 | 0.89 | 1.49 | 0.00 | + |
| Dist Dev | 0.48 | 0.10 | 0.29 | 0.66 | 0.00 | + |
| Dist Road | 0.28 | 0.08 | 0.12 | 0.44 | 0.00 | + |
| Dist Core | 0.12 | 0.13 | −0.13 | 0.37 | 0.35 | − |
| Dist Crop | 0.29 | 0.09 | 0.11 | 0.47 | 0.00 | + |
| Protected | −0.33 | 0.18 | −0.67 | 0.02 | 0.06 | + |
| Detection | ||||||
| Intercept | −2.14 | 0.05 | −2.25 | −2.04 | 0.00 | |
| Spring | −0.25 | 0.08 | −0.41 | −0.08 | 0.00 | + |
| Summer | 0.23 | 0.07 | 0.10 | 0.37 | 0.00 | + |
| Bobcat | ||||||
| Occupancy | ||||||
| Intercept | −2.47 | 0.33 | −3.11 | −1.84 | 0.00 | + |
| Forest 1000 | 2.22 | 0.47 | 1.29 | 3.14 | 0.00 | + |
| Grass 1000 | 0.98 | 0.36 | 0.29 | 1.68 | 0.01 | + |
| Dist Dev | 0.12 | 0.11 | −0.09 | 0.33 | 0.28 | + |
| Dist Road | 0.20 | 0.10 | 0.00 | 0.40 | 0.05 | + |
| Dist Core | 0.17 | 0.33 | −0.47 | 0.81 | 0.61 | − |
| Dist Crop | 0.43 | 0.12 | 0.19 | 0.67 | 0.00 | + |
| Protected | −0.14 | 0.28 | −0.68 | 0.40 | 0.62 | + |
| Detection | ||||||
| Intercept | −3.01 | 0.10 | −3.21 | −2.82 | 0.00 | |
| Spring | 0.37 | 0.14 | 0.10 | 0.65 | 0.01 | + |
| Species | β | SE | LCI | UCI | p-Value | a priori |
|---|---|---|---|---|---|---|
| Model | ||||||
| Parameter | ||||||
| Domestic Cat | ||||||
| Occupancy | ||||||
| Intercept | −3.567 | 0.1981 | −3.96 | −3.18 | 0.00 | |
| Dev 500 | 0.302 | 0.0813 | 0.14 | 0.46 | 0.00 | + |
| Dist Dev | −0.343 | 0.2163 | −0.77 | 0.08 | 0.11 | − |
| Dist Road | −0.718 | 0.2955 | −1.30 | −0.14 | 0.02 | − |
| Core 500 | −0.084 | 0.1809 | −0.44 | 0.27 | 0.64 | − |
| Detection | ||||||
| Intercept | −1.72 | 0.0902 | −1.90 | −1.54 | 0.00 | |
| Fall | −0.454 | 0.1875 | −0.82 | −0.09 | 0.02 | − |
| Red Fox | ||||||
| Occupancy | ||||||
| Intercept | −1.386 | 0.0759 | −1.53 | −1.24 | 0.00 | |
| Dev 1000 | 0.257 | 0.0799 | 0.10 | 0.41 | 0.00 | + |
| Dist Dev | −0.204 | 0.0987 | −0.40 | −0.01 | 0.04 | − |
| Dist Road | −0.174 | 0.1096 | −0.39 | 0.04 | 0.11 | − |
| Core 1000 | −0.572 | 0.0945 | −0.76 | −0.39 | 0.00 | − |
| Detection | ||||||
| Intercept | −2.104 | 0.0492 | −2.20 | −2.01 | 0.00 | |
| Fall | 0.256 | 0.074 | 0.11 | 0.40 | 0.00 | − |
| White-tailed Deer | ||||||
| Occupancy | ||||||
| Intercept | 2.508 | 0.227 | 2.06 | 2.95 | 0.00 | |
| For 1000 | −0.27 | 0.217 | −0.70 | 0.16 | 0.22 | + |
| Grass 1000 | 0.185 | 0.166 | −0.14 | 0.51 | 0.26 | + |
| Dist Dev | −0.296 | 0.111 | −0.51 | −0.08 | 0.01 | − |
| Dist Road | −0.188 | 0.107 | −0.40 | 0.02 | 0.08 | − |
| Dist Core | −0.497 | 0.112 | −0.72 | −0.28 | 0.00 | + |
| Dist Crop | −0.337 | 0.125 | −0.58 | −0.09 | 0.01 | − |
| Protected | 0.524 | 0.295 | −0.05 | 1.10 | 0.08 | − |
| Detection | ||||||
| Intercept | −0.618 | 0.0132 | −0.64 | −0.59 | 0.00 | |
| Spring | −0.17 | 0.0267 | −0.22 | −0.12 | 0.00 | + |
| Species | Scenario | Status | Core Habitat | Suitable Habitat | ||
|---|---|---|---|---|---|---|
| Area (km2) | Percent Change | Area (km2) | Percent Change | |||
| Bear | 2011 | Not Protected | 1753.3 | 6439.8 | ||
| Protected | 2314.4 | 4107.7 | ||||
| HS | Not Protected | 1619.3 | −7.6 | 6262.3 | −2.8 | |
| Protected | 2285.3 | −1.3 | 4109.8 | 0.0 | ||
| HR | Not Protected | 1591.4 | −9.2 | 6070.4 | −5.7 | |
| Protected | 2284.2 | −1.3 | 4091.1 | −0.4 | ||
| LR | Not Protected | 1607.2 | −8.3 | 6200.1 | −3.7 | |
| Protected | 2290.9 | −1.0 | 4110.3 | 0.1 | ||
| LS | Not Protected | 1637.7 | −6.6 | 6331.6 | −1.7 | |
| Protected | 2338.1 | 1.0 | 4147.0 | 1.0 | ||
| Bobcat | 2011 | Not Protected | 555.5 | 2317.6 | ||
| Protected | 1713.3 | 3295.6 | ||||
| HS | Not Protected | 423.7 | −23.7 | 1930.3 | −16.7 | |
| Protected | 1649.0 | −3.8 | 3220.7 | −2.3 | ||
| HR | Not Protected | 429.7 | −22.6 | 1927.1 | −16.8 | |
| Protected | 1682.3 | −1.8 | 3230.8 | −2.0 | ||
| LR | Not Protected | 426.6 | −23.2 | 1911.1 | −17.5 | |
| Protected | 1695.7 | −1.0 | 3231.7 | −1.9 | ||
| LS | Not Protected | 442.6 | −20.3 | 1967.3 | −15.1 | |
| Protected | 1743.9 | 1.8 | 3271.6 | −0.7 | ||
| Domestic Cat | 2011 | Not Protected | 5404.4 | 13484.4 | ||
| Protected | 255.7 | 1099.0 | ||||
| HS | Not Protected | 6709.7 | 24.2 | 14028.9 | 4.0 | |
| Protected | 322.7 | 26.2 | 1177.1 | 7.1 | ||
| HR | Not Protected | 7529.6 | 39.3 | 14547.1 | 7.9 | |
| Protected | 353.5 | 38.2 | 1223.1 | 11.3 | ||
| LR | Not Protected | 7075.5 | 30.9 | 14257.1 | 5.7 | |
| Protected | 336.7 | 31.7 | 1194.1 | 8.7 | ||
| LS | Not Protected | 6336.7 | 17.3 | 13866.3 | 2.8 | |
| Protected | 305.4 | 19.5 | 1161.2 | 5.7 | ||
| Red Fox | 2011 | Not Protected | 1874.2 | 11140.8 | ||
| Protected | 53.6 | 858.8 | ||||
| HS | Not Protected | 3190.5 | 70.2 | 11710.7 | 5.1 | |
| Protected | 104.5 | 95.0 | 957.9 | 11.5 | ||
| HR | Not Protected | 3532.0 | 88.5 | 12531.7 | 12.5 | |
| Protected | 107.5 | 100.6 | 1038.0 | 20.9 | ||
| LR | Not Protected | 3129.5 | 67.0 | 12198.3 | 9.5 | |
| Protected | 90.9 | 69.6 | 992.3 | 15.5 | ||
| LS | Not Protected | 2810.8 | 50.0 | 11454.4 | 2.8 | |
| Protected | 91.8 | 71.4 | 926.8 | 7.9 | ||
| White-tailed Deer | 2011 | Not Protected | 5160.2 | 18848.9 | ||
| Protected | 1707.6 | 4364.5 | ||||
| HS | Not Protected | 5005.2 | −3.0 | 18445.1 | −2.1 | |
| Protected | 1769.7 | 3.6 | 4385.5 | 0.5 | ||
| HR | Not Protected | 5096.0 | −1.2 | 18576.7 | −1.4 | |
| Protected | 1806.5 | 5.8 | 4379.0 | 0.3 | ||
| LR | Not Protected | 5231.9 | 1.4 | 18925.4 | 0.4 | |
| Protected | 1787.0 | 4.6 | 4376.9 | 0.3 | ||
| LS | Not Protected | 5067.3 | −1.8 | 18649.0 | −1.1 | |
| Protected | 1752.3 | 2.6 | 4324.9 | −0.9 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cove, M.V.; Fergus, C.; Lacher, I.; Akre, T.; McShea, W.J. Projecting Mammal Distributions in Response to Future Alternative Landscapes in a Rapidly Transitioning Region. Remote Sens. 2019, 11, 2482. https://doi.org/10.3390/rs11212482
Cove MV, Fergus C, Lacher I, Akre T, McShea WJ. Projecting Mammal Distributions in Response to Future Alternative Landscapes in a Rapidly Transitioning Region. Remote Sensing. 2019; 11(21):2482. https://doi.org/10.3390/rs11212482
Chicago/Turabian StyleCove, Michael V., Craig Fergus, Iara Lacher, Thomas Akre, and William J. McShea. 2019. "Projecting Mammal Distributions in Response to Future Alternative Landscapes in a Rapidly Transitioning Region" Remote Sensing 11, no. 21: 2482. https://doi.org/10.3390/rs11212482
APA StyleCove, M. V., Fergus, C., Lacher, I., Akre, T., & McShea, W. J. (2019). Projecting Mammal Distributions in Response to Future Alternative Landscapes in a Rapidly Transitioning Region. Remote Sensing, 11(21), 2482. https://doi.org/10.3390/rs11212482