
Out of the Dark: Establishing a Large-Scale Field Experiment to Assess the Effects of Artificial Light at Night on Species and Food Webs

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
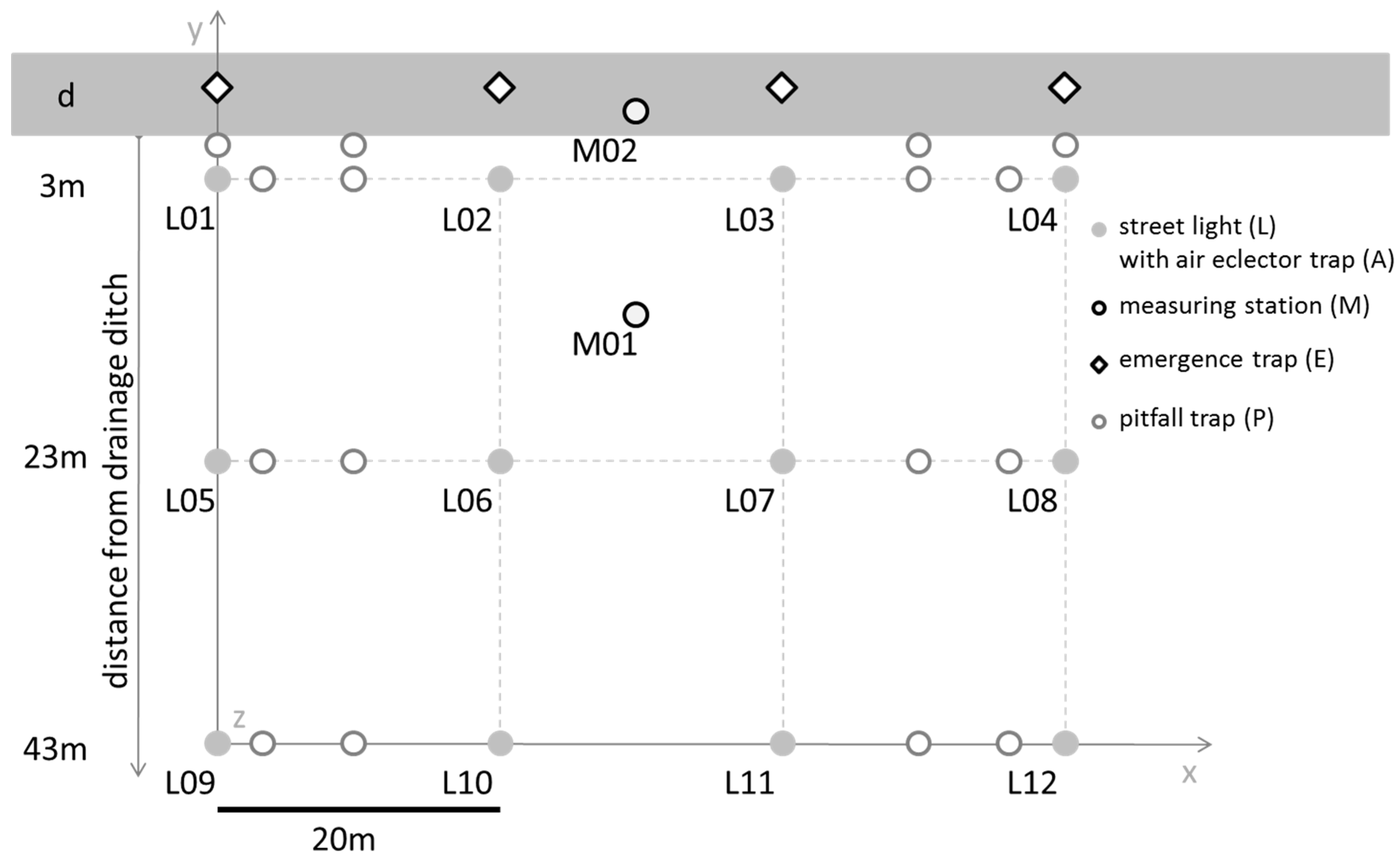
2.1. Study Area
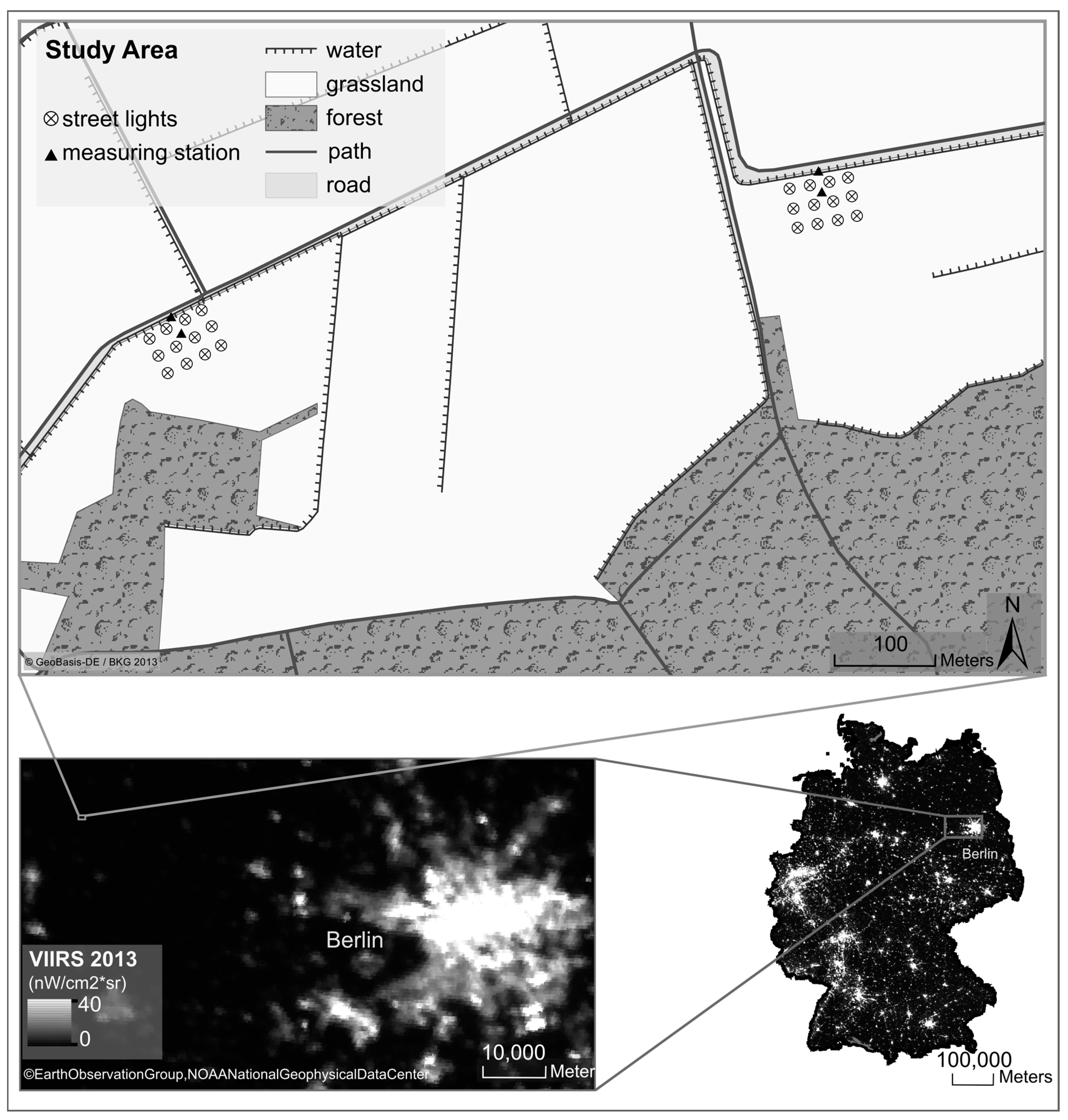
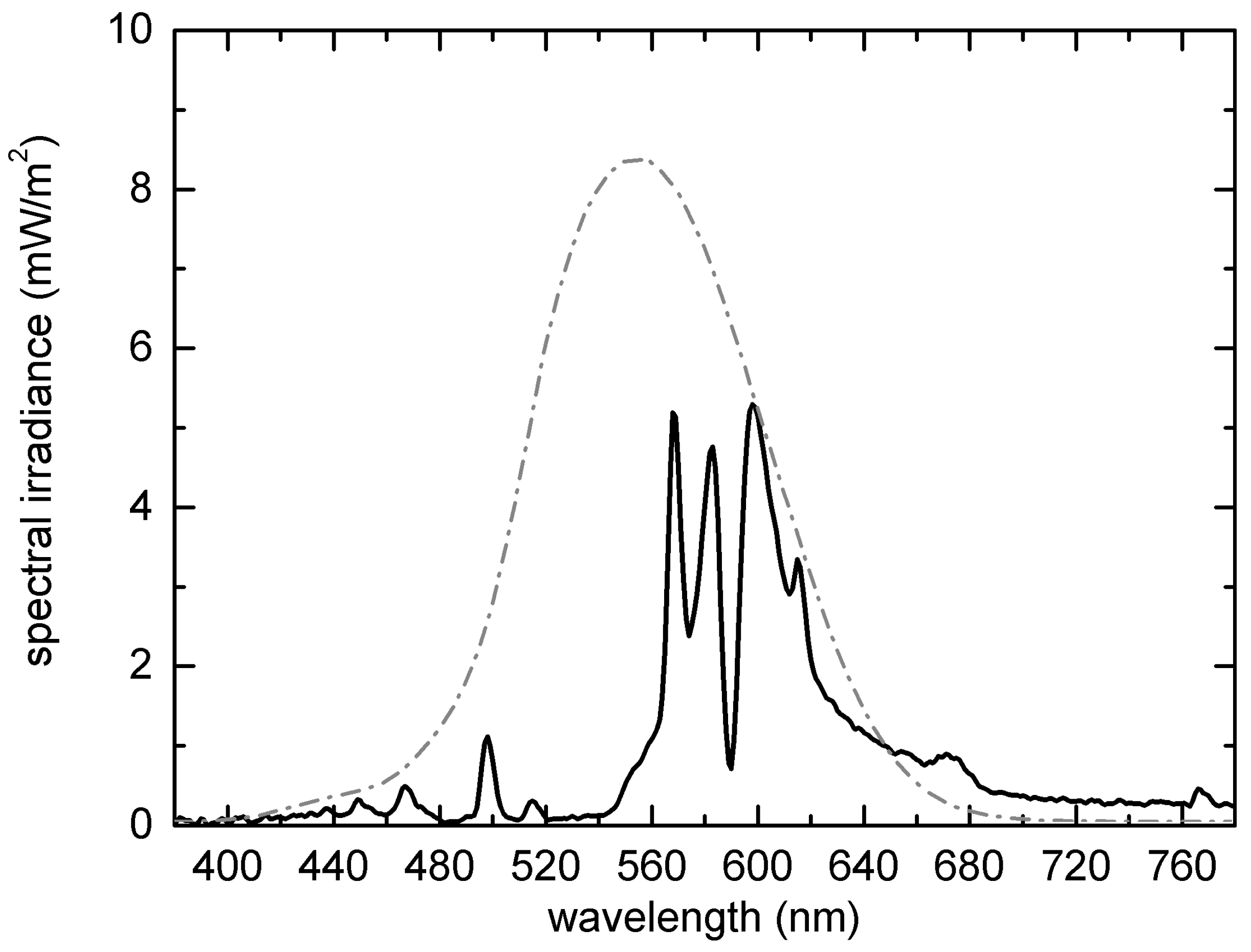
2.2. Street-Light Infrastructure and Sampling of Light and Abiotic Data
2.2.1. Street-Light Infrastructure
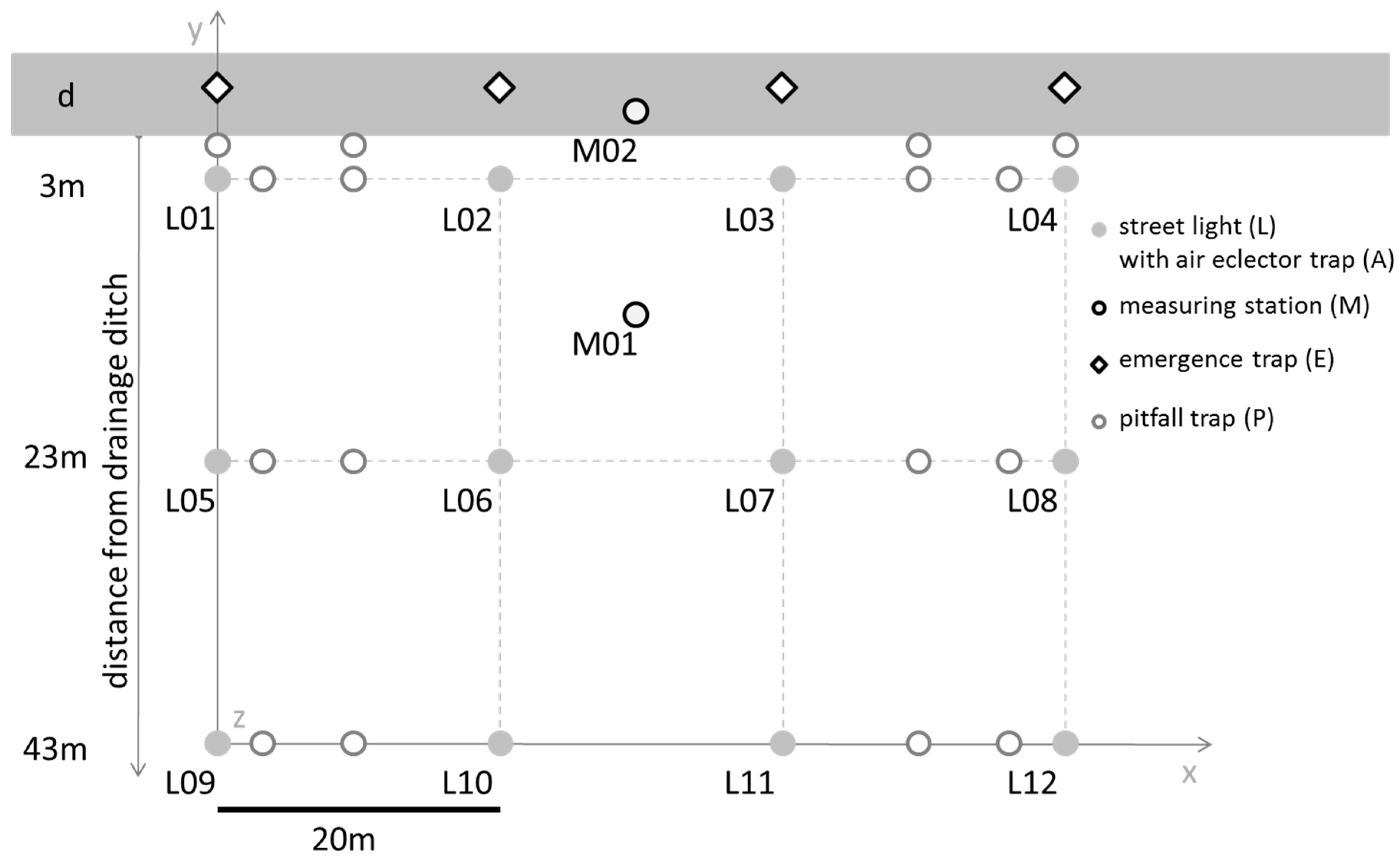
2.2.2. Automated Sampling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property (Unit) | Device | Position Coordinates (m) P(x, y, z) | |
|---|---|---|---|
| Light-Related Parameters | |||
| broadband solar irradiance (W/m2) | CMP3 Pyranometer (300 to 2800 nm), Kipp & Zonen | M01/03: P(30, 30, 2.75) | |
| photosynthetically active radiation (µmol·m−2·s−1) | PQS 1 PAR Quantum Sensor (300–700 nm), Kipp & Zonen | M01/03: P(30, 30, 2.75) M02/04: P(30, 43, −0.80) | |
| relative irradiance (mV); after transformation: illuminance (lux) | Kuffner-Lightmeter Mark 2.4 V03.01 Analog, K2W Lights | M01: P(30, 30, 2.75) M01: P(30, 30, 5.25) M03: P(30, 30, 2.75) M03: P(30, 30, 5.25) | |
| night sky radiance (magSQM/arcsec2) | Sky Quality Meter, Unihedron SQM-LU-DL | L16: P(60, 40, 4.80) | |
| Weather-Related Parameters | |||
| temperature (°C) | Vaisala Weather Transmitter WXT520 | M01/03: P(30, 30, 2.75) | |
| precipitation (mm) | Vaisala Weather Transmitter WXT520 | M01/03: P(30, 30, 2.75) | |
| relative humidity (%) | Vaisala Weather Transmitter WXT520 | M01/03: P(30, 30, 2.75) | |
| barometric pressure (hPa) | Vaisala Weather Transmitter WXT520 | M01/03: P(30, 30, 2.75) | |
| wind intensity (m/s) | Vaisala Weather Transmitter WXT520 | M01/03: P(30, 30, 2.75) | |
| wind direction (degree) (0°–359°) | Vaisala Weather Transmitter WXT520 | M01/03: P(30, 30, 2.75) | |
| precipitation (mm), incl. snow, drizzling rain | Tipping bucket raining gauge (0,2 mm) | M01/03: P(30, 30, 2.75) | |
| wind direction (3D anemometer, vectors x, y, z (m/s), transformation to (°)) | Ultrasonic 3d Anemometer; THIES CLIMA | M01/03: P(30, 30, 5.25) | |
| Water-Related Parameters | |||
| temperature (°C) | YSI 6600 V2 data sonde, YSI | M02/04: P(30, 43, −0.80) | |
| O2 concentration (mg/L) | YSI 6600 V2 data sonde, YSI | M02/04: P(30, 43, −0.80) | |
| O2 saturation (%) | YSI 6600 V2 data sonde, YSI | M02/04: P(30, 43, −0.80) | |
| pH | YSI 6600 V2 data sonde, YSI | M02/04: P(30, 43, −0.80) | |
| conductivity (µS/cm) | YSI 6600 V2 data sonde, YSI | M02/04: P(30, 43, −0.80) | |
| chlorophyll-a concentration (µg/L) | YSI 6600 V2 data sonde, YSI | M02/04: P(30, 43, −0.80) |
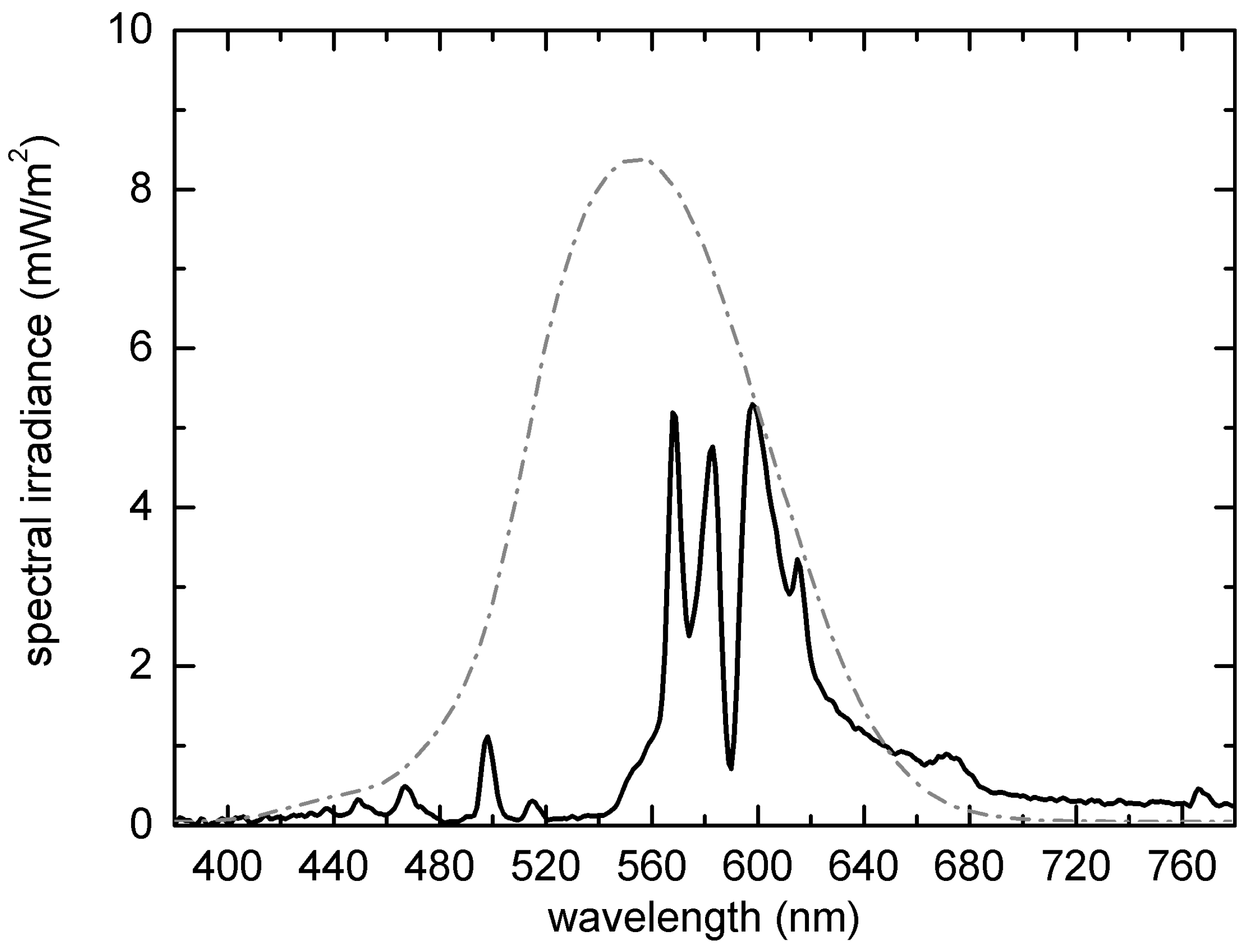
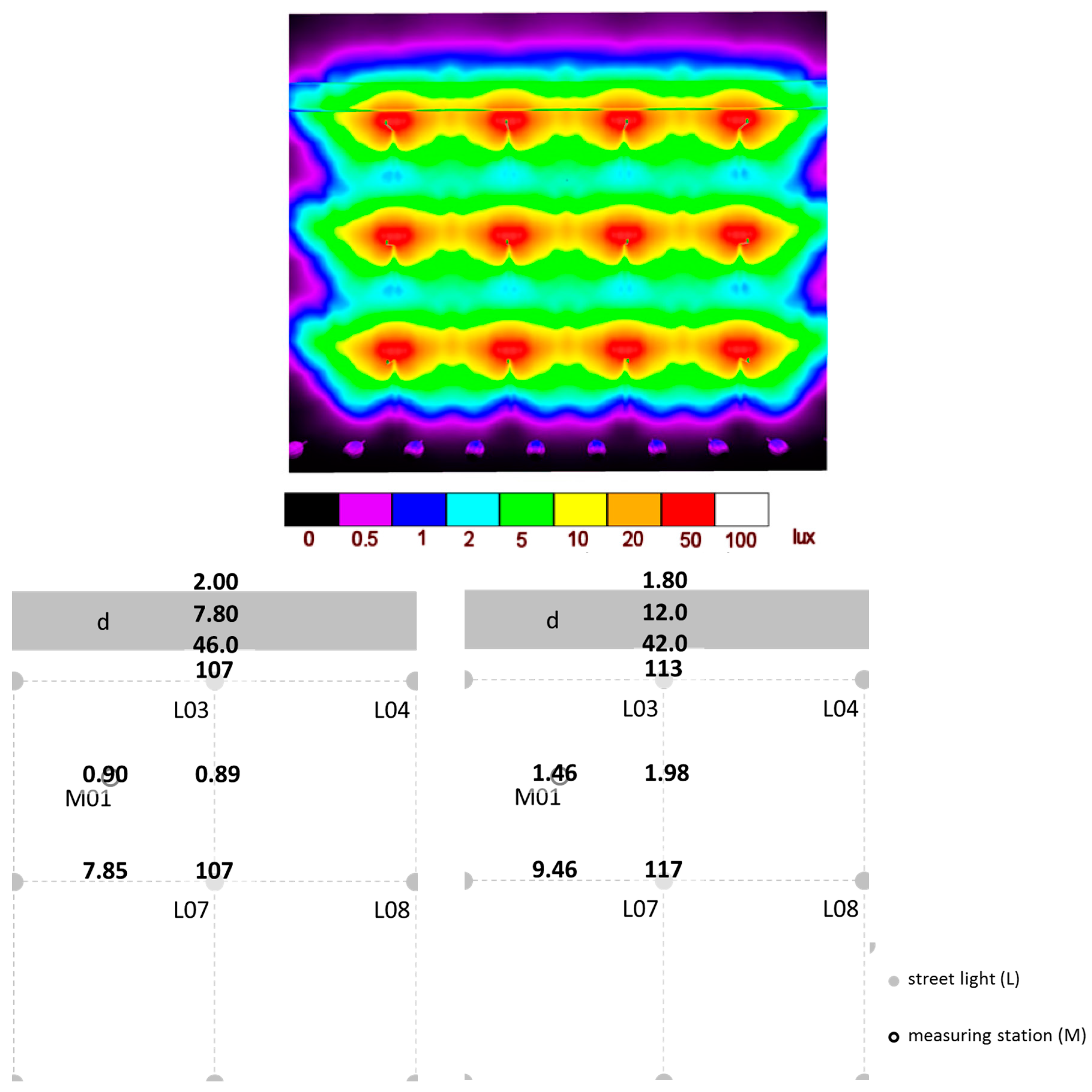
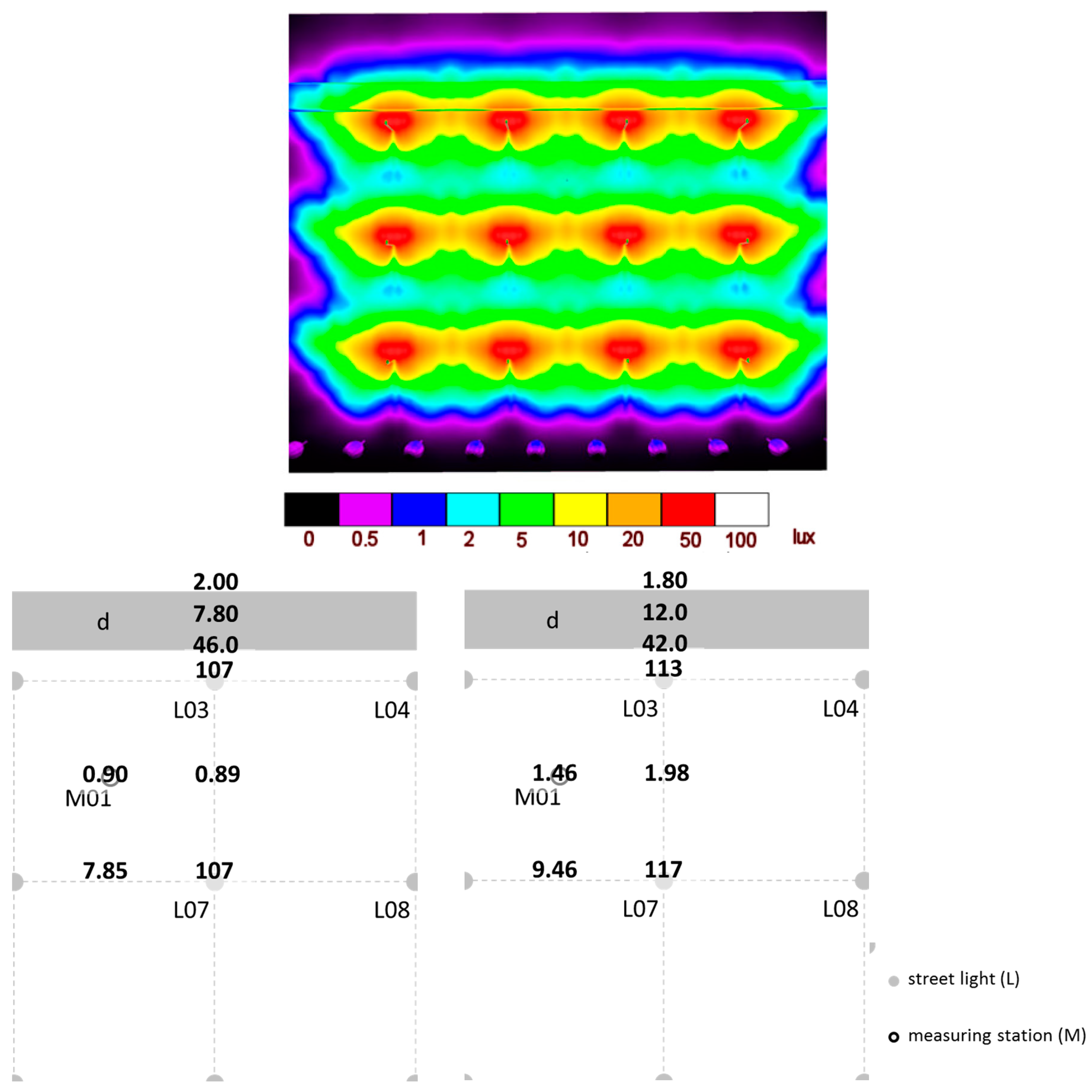
2.3. Photometric Characterization of the Study Area after Installation of the Infrastructure
2.4. Biological Studies
2.4.1. Arthropod Sampling Design
2.4.2. Pilot Study
2.4.3. Monitoring of Bats as a Group of Obligatory Nocturnal Mammals
2.4.4. Monitoring of Birds
2.4.5. Physiological Experiment with Fish
2.4.6. Food Web Dynamics
3. Preliminary Results
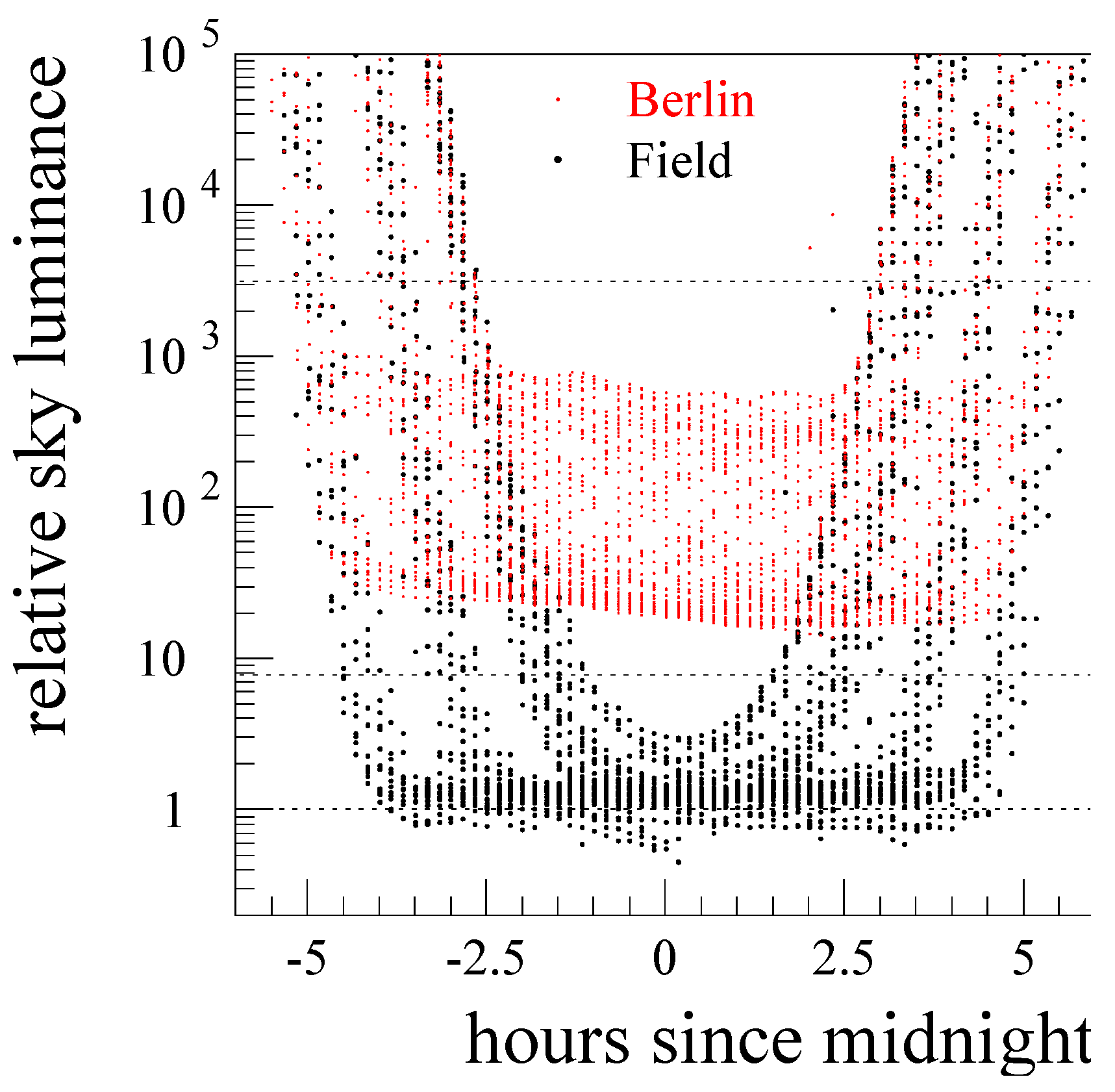
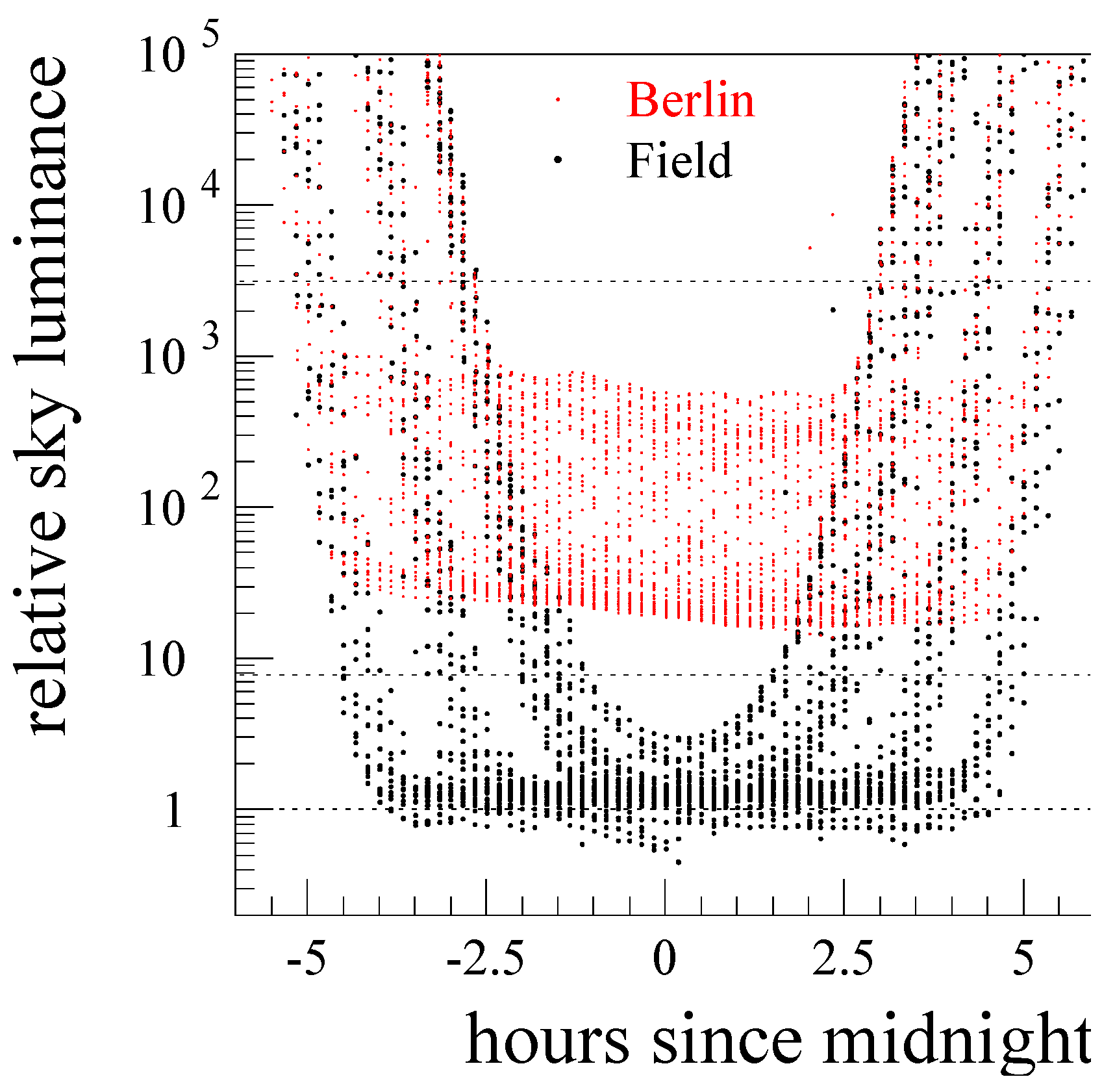
3.1. Comparison of Environmental Parameters between Sites
3.2. Photometric Characterization of the Study Area after Installation of Infrastructure
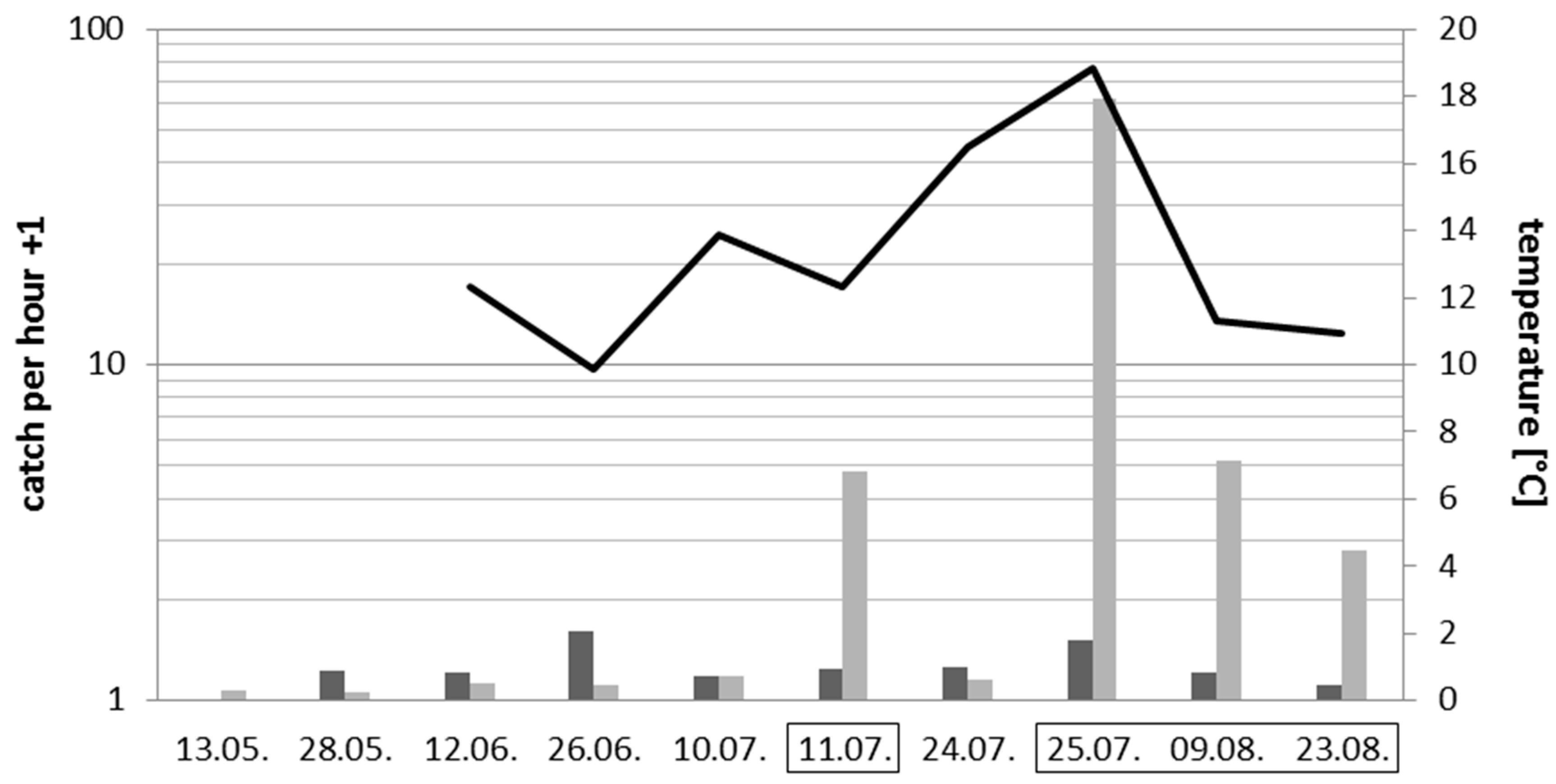
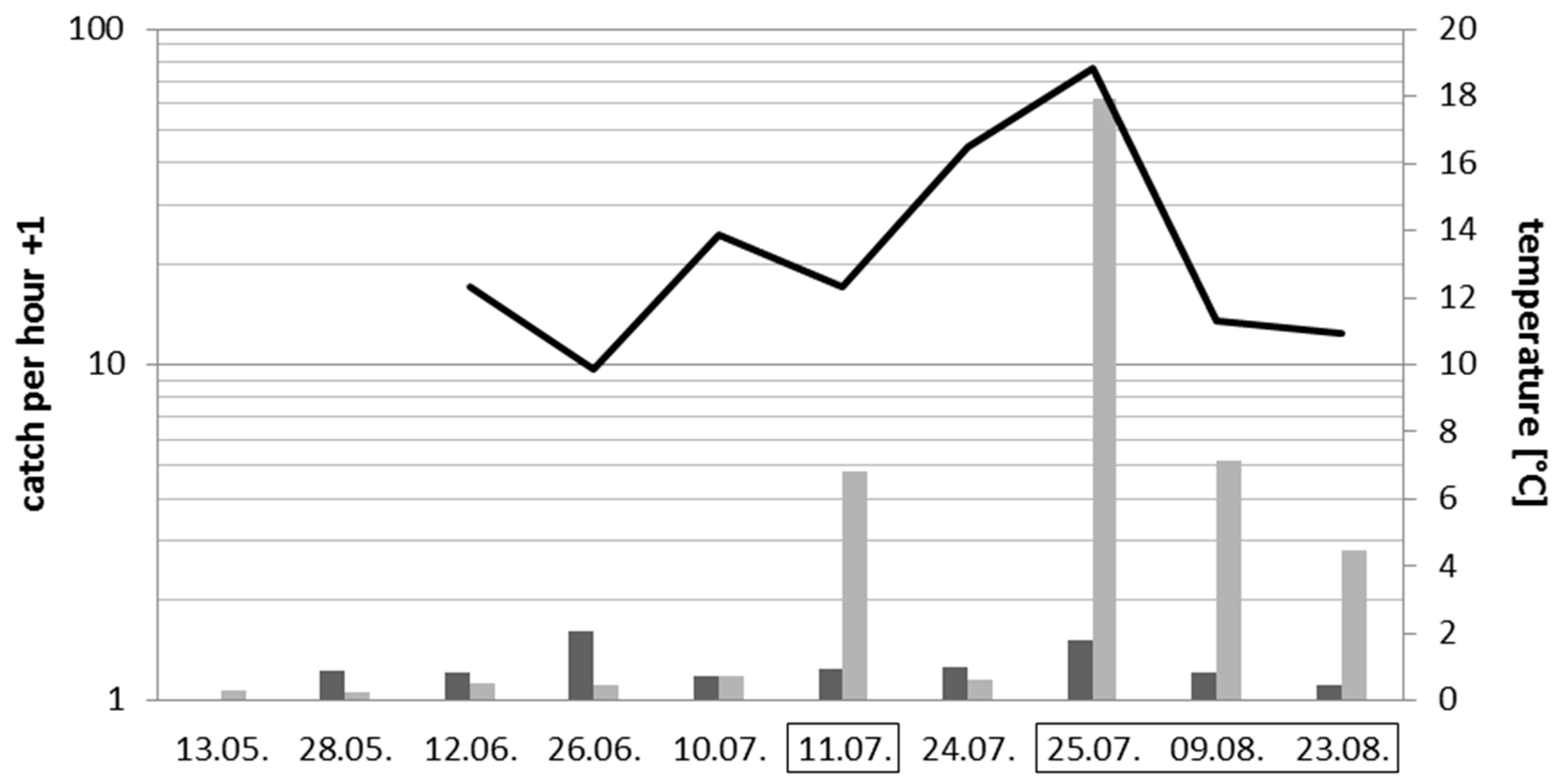
3.3. Effects of Introduced Artificial Light at Night on Insects and Spiders
| Model | Group | χ² | Df | R2m | Comparison |
|---|---|---|---|---|---|
| overnight term | total arthropods | 109.75 *** | 3 | 0.78 | AW/AE *** |
| AW/BE *** | |||||
| AW/BW *** | |||||
| aquatic insects | 148.29 *** | 3 | 0.79 | AW/AE *** | |
| AW/BE *** | |||||
| AW/BW *** | |||||
| period of pilot study | total arthropods | 233.89 *** | 3 | 0.60 | AW/AE *** |
| AW/BE *** | |||||
| AW/BW *** | |||||
| aquatic insects | 278.65 *** | 3 | 0.68 | AW/AE *** | |
| AW/BE *** | |||||
| AW/BW *** |
4. Discussion
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hölker, F.; Moss, T.; Griefahn, B.; Kloas, W.; Voigt, C.C.; Henckel, D.; Hänel, A.; Kappeler, P.M.; Völker, S.; Schwope, A.; et al. The dark side of light: A transdisciplinary research agenda for light pollution policy. Ecol. Soc. 2010, 15. Article 13. [Google Scholar]
- Bennie, J.; Davies, T.; Duffy, J.; Inger, R.; Gaston, K.J. Contrasting trends in light pollution across Europe. Sci. Rep. 2014. [Google Scholar] [CrossRef] [PubMed]
- Gaston, K.J.; Bennie, J.; Davies, T.W.; Hopkins, J. Reducing the ecological consequences of night-time light pollution: Options and developments. J. Appl. Ecol. 2012, 49, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Longcore, T.; Rich, C. Ecological light pollution. Front. Ecol. Environ. 2004, 2, 191–198. [Google Scholar] [CrossRef]
- Hölker, F.; Wolter, C.; Perkin, E.K.; Tockner, K. Light pollution as a biodiversity threat. Trends Ecol. Evol. 2010, 25, 681–682. [Google Scholar] [CrossRef] [PubMed]
- Gaston, K.J.; Visser, M.E.; Hölker, F. The biological impacts of artificial light at night: The research challenge. Philos. Trans. R. Soc. B 2015. [Google Scholar] [CrossRef] [PubMed]
- Gosh, T.; Anderson, S.J.; Elvidge, C.D.; Sutton, P.C. Using nighttime satellite imagery as a proxy measure of human well-being. Sustainability 2013, 5, 4988–5019. [Google Scholar] [CrossRef]
- Rich, C.; Longcore, T. Ecological Consequences of Artificial Night Lighting; Island Press: Washington, DC, USA, 2006. [Google Scholar]
- Moore, M.V.; Kohler, S.J.; Cheers, M.S. Artificial light at night in freshwater habitats and its potential ecological effects. In Ecological Consequences of Artificial Night Lighting; Rich, C., Longcore, T., Eds.; Island Press: Washington, DC, USA, 2006. [Google Scholar]
- Nightingale, B.; Longcore, T.; Simenstad, C.A. Artificial night lighting and fishes. In Ecological Consequences of Artificial Night Lighting; Rich, C., Longcore, T., Eds.; Island Press: Washington, DC, USA, 2006. [Google Scholar]
- Perkin, E.K.; Hölker, F.; Richardson, J.S.; Sadler, J.P.; Wolter, C.; Tockner, K. The influence of artificial light on freshwater and riparian ecosystems: Questions, challenges, and perspectives. Ecosphere 2011, 2, 122. [Google Scholar] [CrossRef]
- Hölker, F.; Wurzbacher, C.; Weißenborn, C.; Monaghan, M.T.; Holzhauer, S.I.J.; Premke, K. Microbial diversity and community respiration in freshwater sediments influenced by artificial light at night. Philos. Trans. R. Soc. B 2015. [Google Scholar] [CrossRef] [PubMed]
- Kummu, M.; de Moel, H.; Ward, P.J.; Varis, O. How close do we live to water? A global analysis of population distance to freshwater bodies. PLoS ONE 2011, 6, e20578. [Google Scholar] [CrossRef] [PubMed]
- Kuechly, H.U.; Kyba, C.C.M.; Ruhtz, T.; Lindemann, C.; Wolter, C.; Fischer, J.; Hölker, F. Aerial survey and spatial analysis of sources of light pollution in Berlin, Germany. Remote Sens. Environ. 2012, 126, 39–50. [Google Scholar] [CrossRef]
- Brüning, A.; Hölker, F.; Franke, S.; Preuer, T.; Kloas, W. Spotlight on fish: Light pollution affects circadian rhythms of European perch but does not cause stress. Sci. Total Environ. 2015, 511, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Perkin, E. K.; Hölker, F.; Tockner, K.; Richardson, J. S. Artificial light as a disturbance to light-naïve streams. Freshw. Biol. 2014, 59, 2235–2244. [Google Scholar] [CrossRef]
- Riley, W.D.; Bendall, B.; Ives, M.J.; Edmonds, N.J.; Maxwell, D.L. Street lighting disrupts the diel migratory pattern of wild Atlantic salmon, Salmo salar L., smolts leaving their natal stream. Aquaculture 2012, 330–333, 74–81. [Google Scholar] [CrossRef]
- Riley, W. D.; Davison, P. I.; Maxwell, D. L.; Newman, R. C.; Ives, M. J. A laboratory experiment to determine the dispersal response of Atlantic salmon (Salmo salar) fry to street light intensity. Freshw. Biol. 2015, 60, 1016–1028. [Google Scholar] [CrossRef]
- Perkin, E. K.; Hölker, F.; Tockner, K. The effects of artificial lighting on adult aquatic and terrestrial insects. Freshw. Biol. 2014, 59, 368–377. [Google Scholar] [CrossRef]
- Meyer, L.A.; Sullivan, S.M.P. Bright lights, big city: influences of ecological light pollution on reciprocal stream-riparian invertebrate fluxes. Ecol. Appl. 2013, 23, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- International Commission on Illumination. CIE 158:2004: Ocular Lighting Effects on Human Physiology and Behavior; International Commission on Illumination: Vienna, Austria, 2004. [Google Scholar]
- International Commission on Illumination. CIE 126:1997: Guidelines for Minimizing Sky Glow; International Commission on Illumination: Vienna, Austria, 1997. [Google Scholar]
- German Institute for Standardisation. B. DIN V 5031–100:2009: Optical Radiation Physics and Illuminating Engineering—Part 100: Non-visual Effects of Ocular Light on Human Beings—Quantities, Symbols and Action Spectra; Beuth: Berlin, Germany, 2009. (In German) [Google Scholar]
- German Institute for Standardisation. B. DIN SPEC 67600:2013: Biologically Effective Illumination—Design Guidelines; Beuth: Berlin, Germany, 2013. (In German) [Google Scholar]
- International Commission on Illumination. CIE 150:2003: Guide on the Limitation of the Effects of Obstrusive Light from Outdoor Lighting Installations; International Commission on Illumination: Vienna, Austria, 2003. [Google Scholar]
- Spoelstra, K.; van Grunsven, R.H.A.; Donners, M.; Gienapp, P.; Huigens, M.E.; Slaterus, R.; Berendse, F.; Visser, M.E.; Veenendaal, E. Experimental illumination of natural habitat—An experimental set-up to assess the direct and indirect ecological consequences of artificial light of different spectral composition. Philos. Trans. R. Soc. B 2015, 370, 20140129. [Google Scholar] [CrossRef] [PubMed]
- Bennie, J.; Davies, T.W.; Cruse, D.; Inger, R.; Gaston, K.J. Cascading effects of artificial light at night: Resource-mediated control of herbivores in a grassland ecosystem. Philos. Trans. R. Soc. B 2015. [Google Scholar] [CrossRef] [PubMed]
- Perkin, E.K.; Hölker, F.; Heller, S.; Berghahn, S. Artificial light and nocturnal activity in gammarids. PeerJ 2014, 2, 279. [Google Scholar] [CrossRef] [PubMed]
- Thurber, W.A.; Komar, O. Turquoise-browed motmot (Eumomota superciliosa) feeds by artificial light. Wilson Bull. 2002, 114, 525–526. [Google Scholar] [CrossRef]
- Eisenbeis, G. Artificial night lighting and insects: Attraction of insects to streetlamps in a rural setting in germany. In Ecological Consequences of Artificial Night Lighting; Rich, C., Longcore, T., Eds.; Island Press: Washington, DC, USA, 2006. [Google Scholar]
- Lebbin, D.J.; Harvey, M.G.; Lenz, T.C.; Andersen, M.J.; Ellis, J.M. Nocturnal migrants foraging at night by artificial light. Wilson J. Ornithol. 2007, 119, 506–508. [Google Scholar] [CrossRef]
- Davies, T.W.; Bennie, J.; Gaston, K.J. Street lighting changes the composition of invertebrate communities. Biol. Lett. 2012, 8, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, L. Logic of experiments in ecology: is pseudoreplication a pseudoissue? Oikos 2001, 94, 27–38. [Google Scholar] [CrossRef]
- Landesamt für Umwelt, G.u.V.L., Brandenburg, Germany. Naturpark Westhavelland. Available online: http://www.lugv.brandenburg.de/cms/detail.php/bb1.c.331126.de (accessed on 13 November 2015).
- International Dark Sky Association (IDA). International Dark Sky Reserves: Westhavelland (Germany). Available online: http://darksky.org/idsp/reserves/westhavelland/ (accessed on 13 November 2015).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1–120. Available online: http://CRAN.R-project.org/package=nlme (accessed on 13 November 2015).
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org (accessed on 13 November 2015).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; Sage Publications: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Fox, J. Applied Regression Analysis and Generalized Linear Models, 2nd ed.; Sage: Thousand Oaks, CA, USA, 2008. [Google Scholar]
- Cinzano, P. Night Sky Photometry with Sky Quality Meter. Available online: http://www.unihedron.com/projects/darksky/sqmreport_v1p4.pdf (accessed on 13 November 2015).
- Schnitt, S.; Ruhtz, T.; Fischer, J.; Hölker, F.; Kyba, C.C.M. Temperature stability of the sky quality meter. Sensors 2013, 13, 12166–12174. [Google Scholar] [CrossRef] [PubMed]
- Kyba, C.C.M.; Ruhtz, T.; Fischer, J.; Hölker, F. Cloud coverage acts as an amplifier for ecological light pollution in urban ecosystems. PLoS ONE 2011, 6, e17307. [Google Scholar] [CrossRef] [PubMed]
- Lolkema, D.E.; Haaima, M.; den Outer, P.N.; Spoelstra, H. Effects of meteorological and atmospheric parameters on night sky brightness. RIVM Rapp. 2011. [Google Scholar] [CrossRef]
- Kyba, C.C.M.; Tong, K.P.; Bennie, J.; Birriel, I.; Birriel, J.J.; Cool, A.; Danielsen, A.; Davies, T.W.; Outer, P.N.D.; Edwards, W.; et al. Worldwide variations in artificial skyglow. Sci. Rep. 2015, 5, 8409. [Google Scholar] [CrossRef] [PubMed]
- Cinzano, P.; Falchi, F.; Elvidge, C.D. The first world atlas of the artificial night sky brightness. Mon. Notices R. Astron. Soc. 2001, 328, 689–707. [Google Scholar] [CrossRef]
- Kyba, C.C.M.; Lolkema, D.E. A community standard for the recording of skyglow data. Astron. Geophys. 2012, 53, 6.17–6.18. [Google Scholar] [CrossRef]
- Schaffrath, U. Zur Käferfauna am Edersee (Insecta, Coleoptera). Philippia 1999, 9, 1–94. (In German) [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B. Lme4: Linear Mixed-Effects Models Using s4 Classes. 2013. Available online: http://www.cran.r-project.org/ (accessed on 13 November 2015).
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models Methods and Classes for S and S-plus. Available online: https://www.biostat.wisc.edu/sites/default/files/tr_89.pdf (accessed on 13 November 2015).
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. A General and Simple Method for Obtaining R2 from Generalized Linear Mixed-Effects Models. Methods Ecol. Evol. 2014, 4, 133–142. [Google Scholar] [CrossRef]
- Barton, K. Mumin: Multi-Model Inference. R Package Version 0.3.10. 2011. Available online: http://R-Forge.R-project.org/projects/mumin (accessed on 13 November 2015).
- De Rosario-Martinez, H. Phia: Post-Hoc Interaction Analysis. R Package Version 0.1–5. 2013. Available online: http://CRAN.R-project.org/package=phia (accessed on 13 November 2015).
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Rydell, J. Bats and their insect prey at street lights. In Ecological Consequences of Artificial Night Lighting; Rich, C., Longcore, T., Eds.; Island Press: Washington, DC, USA, 2006. [Google Scholar]
- Rowse, E.G.; Lewanzik, D.; Stone, E.L.; Harris, S.; Jones, G. Dark matters: the effects of artificial lighting on bats. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer: Berlin, Germany, 2016; in press. [Google Scholar]
- Rydell, J. Exploitation of insects around streetlamps by bats in Sweden. Funct. Ecol. 1992, 6, 744–750. [Google Scholar] [CrossRef]
- Rydell, J.; Racey, P.A. Street lamps and the feeding ecology of insectivorous bats. Symp. Zool. Soc. Lond. 1995, 67, 291–307. [Google Scholar]
- Schnitzler, H.U.; Kalko, E.K.V. Echolocation by insect-eating bats. Bioscience 2001, 51, 557–569. [Google Scholar] [CrossRef]
- Kempenaers, B.; Borgström, P.; Loës, P.; Schlicht, E.; Valcu, M. Artificial night lighting affects dawn song, extra-pair siring success, and lay date in songbirds. Curr. Biol. 2010, 20, 1735–1739. [Google Scholar] [CrossRef] [PubMed]
- Bridge, E.S.; Bonter, D.N. A low-cost radio frequency identification device for ornithological research. Journal of field ornithology. J. Field Ornithol. 2011, 82, 52–59. [Google Scholar] [CrossRef]
- Bonter, D.N.; Bridge, E.S. Applications of radio frequency identification (rfid) in ornithological research: A review. J. Field Ornithol. 2011, 82, 1–10. [Google Scholar] [CrossRef]
- Frank, K. Effects of artificial night lighting on moths. In Ecological Consequences of Artificial Night Lighting; Rich, C., Longcore, T., Eds.; Island Press: Washington, DC, USA, 2006. [Google Scholar]
- Truxa, C.; Fiedler, K. Attraction to light—From how far do moths (Lepidoptera) return to weak artificial sources of light? Eur. J. Entomol. 2012, 109, 77–84. [Google Scholar] [CrossRef]
- Beck, J.; Linsenmair, K.E. Feasibility of light-trapping in community research on moths: Attraction radius of light, completeness of samples, nightly flight times and seasonality of Southeast-Asian hawkmoths (Lepidoptera: Sphingidae). J. Res. Lepidoptera 2006, 39, 18–36. [Google Scholar]
- McCafferty, W.P. Aquatic Entomology: The Fisherman’s and Ecologists’ Illustrated Guide to Insects and Their Relatives; Science Books International, Inc.: Boston, MA, USA, 1981. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holzhauer, S.I.J.; Franke, S.; Kyba, C.C.M.; Manfrin, A.; Klenke, R.; Voigt, C.C.; Lewanzik, D.; Oehlert, M.; Monaghan, M.T.; Schneider, S.; et al. Out of the Dark: Establishing a Large-Scale Field Experiment to Assess the Effects of Artificial Light at Night on Species and Food Webs. Sustainability 2015, 7, 15593-15616. https://doi.org/10.3390/su71115593
Holzhauer SIJ, Franke S, Kyba CCM, Manfrin A, Klenke R, Voigt CC, Lewanzik D, Oehlert M, Monaghan MT, Schneider S, et al. Out of the Dark: Establishing a Large-Scale Field Experiment to Assess the Effects of Artificial Light at Night on Species and Food Webs. Sustainability. 2015; 7(11):15593-15616. https://doi.org/10.3390/su71115593
Chicago/Turabian StyleHolzhauer, Stephanie I. J., Steffen Franke, Christopher C. M. Kyba, Alessandro Manfrin, Reinhard Klenke, Christian C. Voigt, Daniel Lewanzik, Martin Oehlert, Michael T. Monaghan, Sebastian Schneider, and et al. 2015. "Out of the Dark: Establishing a Large-Scale Field Experiment to Assess the Effects of Artificial Light at Night on Species and Food Webs" Sustainability 7, no. 11: 15593-15616. https://doi.org/10.3390/su71115593
APA StyleHolzhauer, S. I. J., Franke, S., Kyba, C. C. M., Manfrin, A., Klenke, R., Voigt, C. C., Lewanzik, D., Oehlert, M., Monaghan, M. T., Schneider, S., Heller, S., Kuechly, H., Brüning, A., Honnen, A.-C., & Hölker, F. (2015). Out of the Dark: Establishing a Large-Scale Field Experiment to Assess the Effects of Artificial Light at Night on Species and Food Webs. Sustainability, 7(11), 15593-15616. https://doi.org/10.3390/su71115593