Mechanisms of Forest Restoration in Landslide Treatment Areas


Abstract
:1. Introduction
2. Material and Methods
2.1. Sites
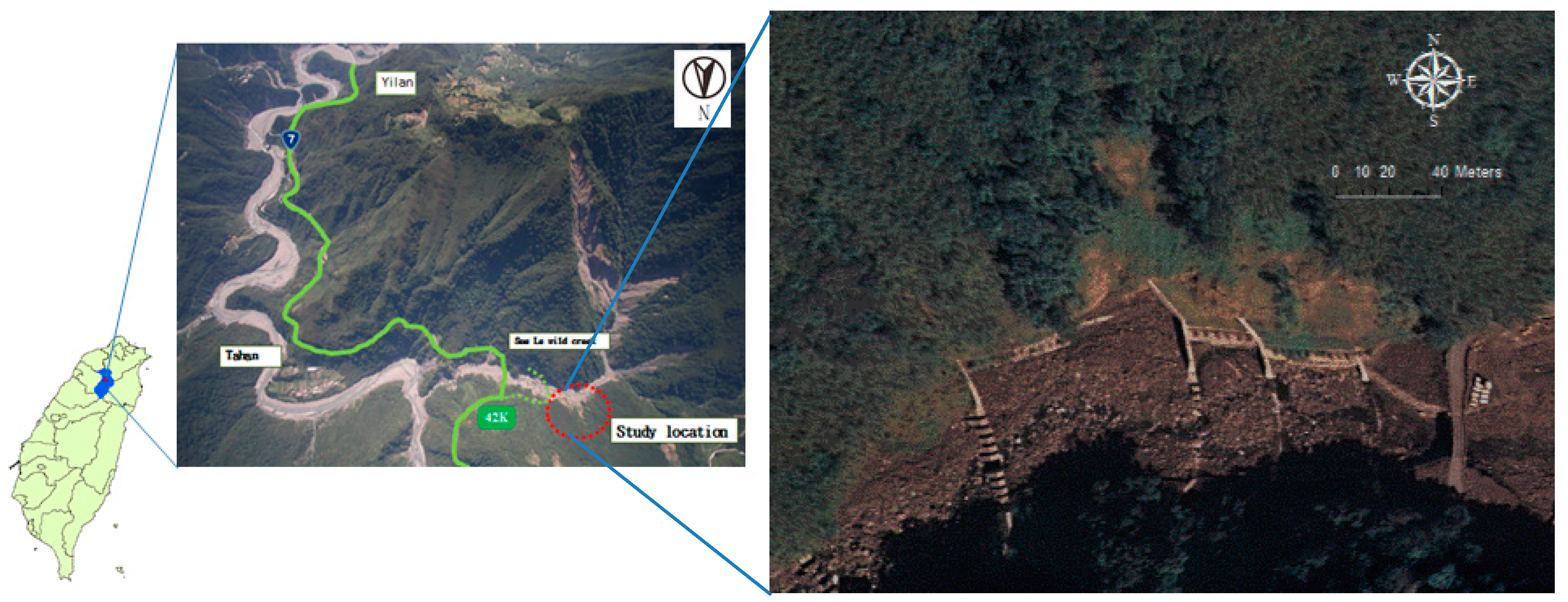
2.2. Relative Illuminance and Biomass of Rhodes Grass


2.3. Germination of Soil-Seed Bank
2.4. Analyses of Aerial Images and Summed Dominance Ratio
3. Results
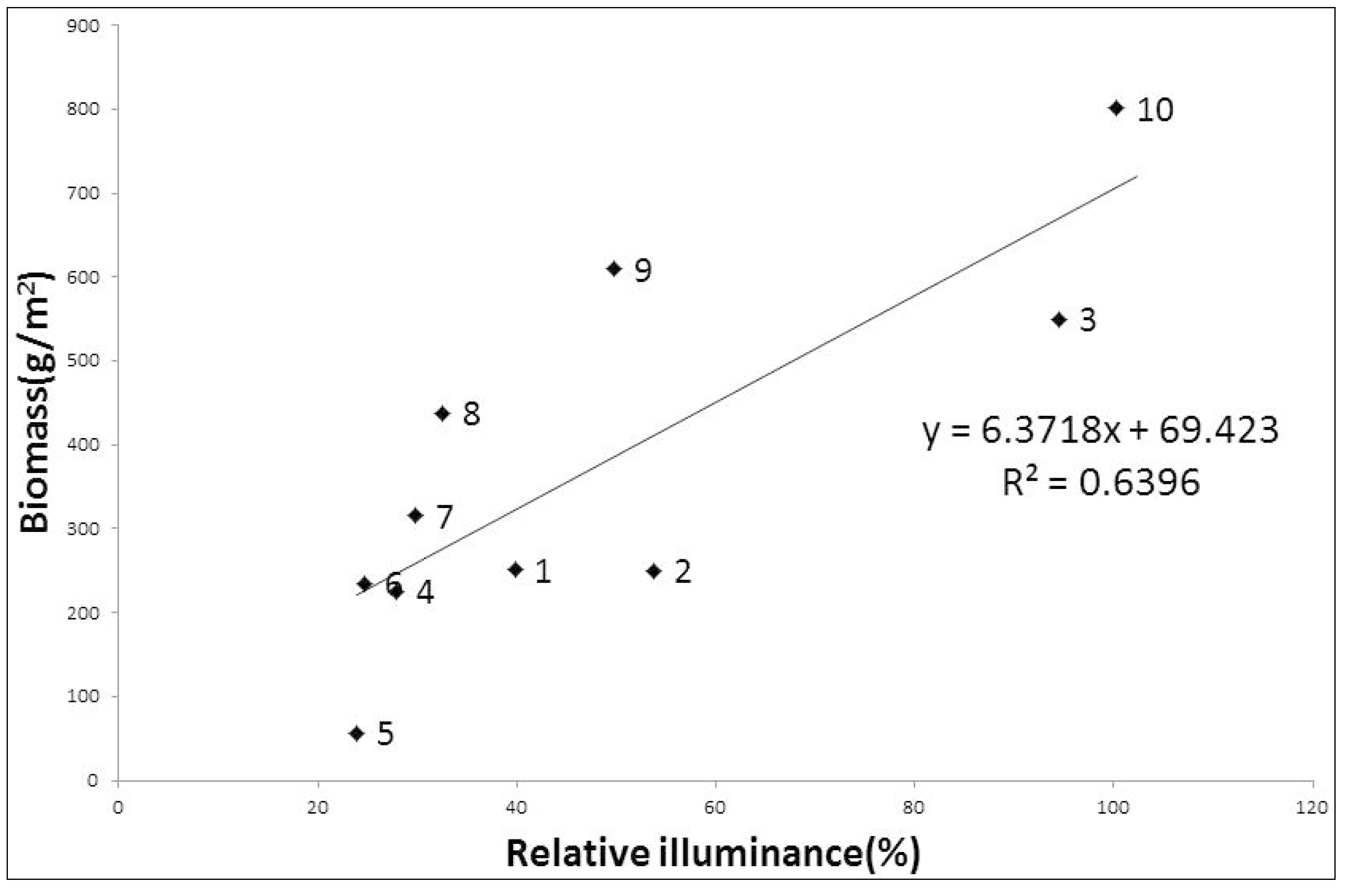
3.1. Relative Illuminance and Biomass of Rhodes Grass
3.2. Soil-Seed Bank

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Names of Plants | Soil Depth: 0–5 cm | Soil Depth: 5–10 cm | Germinate Quantity | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | A | B | C | D | E | ||
| Boehmeria nivea | 91 | 24 | 20 | 18 | 91 | 35 | 36 | 33 | 72 | 420 * | |
| Pouzolzia elegans | 43 | 28 | 26 | 20 | 1 | 32 | 2 | 152 * | |||
| Bidens pilosa | 14 | 2 | 1 | 6 | 1 | 3 | 3 | 2 | 2 | 1 | 35 |
| Crassocephalum crepidioides | 1 | 2 | 1 | 2 | 1 | 2 | 9 | ||||
| Conyza canadensis | 1 | 3 | 2 | 6 | |||||||
| Solanum nigrum | 2 | 2 | 4 | ||||||||
| Cynodon dactylon | 1 | 1 | 1 | 3 | |||||||
| Chloris gayana | 1 | 2 | 3 | ||||||||
| Solanum americanum | 2 | 2 | |||||||||
| Celtis sinensis | 1 | 1 * | |||||||||
| Trema orientalis | 1 | 1 * | |||||||||
| Boehmeria densiflora | 1 | 1 * | |||||||||
| Polygonum multiflorum | 1 | 1 | |||||||||
| Oplismenus hirtellus | 1 | 1 | |||||||||
| Commelina communis | 1 | 1 | |||||||||
| Poaceae | 1 | 1 | |||||||||
| Germinating Quantity of each zone | 106 | 74 | 50 | 52 | 2 | 98 | 61 | 46 | 75 | 75 | 639 |
| species of each zone | 3 | 8 | 4 | 4 | 2 | 4 | 6 | 8 | 8 | 3 | |

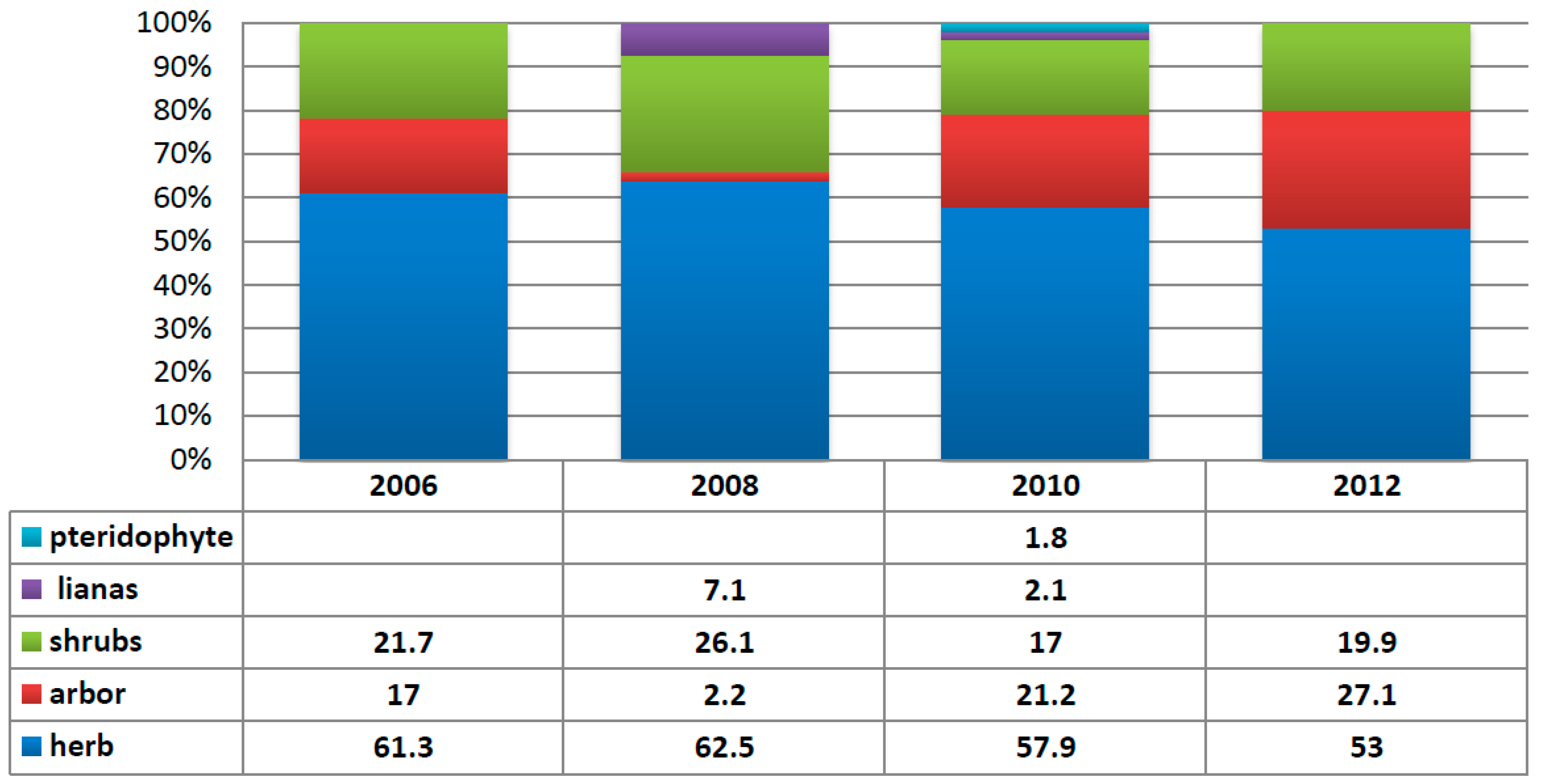
3.3. Annual Variation in the Region Containing the Tree Island and Summed Dominance Ratio
| Year, Month | Orthophotos | Invasion of Region | Site Photos |
|---|---|---|---|
| 2005, January |  | 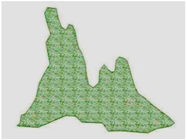 |  |
| 2006, June |  |  |  |
| 2007, January |  | 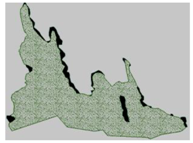 |  |
| 2008, July |  | 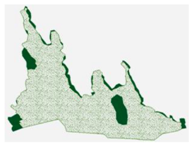 |  |
| 2010, October |  | 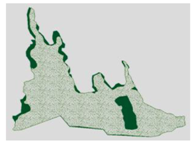 |  |
| 2012, December | - | - |  |
| Y/M | Landslide | Treating | Treated | 2007/1 | 2008/7 | 2010/10 | |
|---|---|---|---|---|---|---|---|
| 2004/10 | 2005/9 | 2006/6 | |||||
| Grassland | - | - | 5494.58 | 5376.91 | 5074.41 | 4974.93 | |
| Trees island | - | - | 18.32 | 57.6 | 186.7 | 280.24 | |
| Tree invasion from forest edge | - | - | 0 | 39.19 | 106.29 | 201.6 | |
| 2006 | 2008 | 2010 | 2012 | ||||
|---|---|---|---|---|---|---|---|
| Names of Plants | SDR | Names of Plants | SDR | Names of Plants | SDR | Names of Plants | SDR |
| Chloris gayana | 47.75 | Chloris gayana | 33.4 | Chloris gayana | 32.4 | Chloris gayana | 27.20 |
| Broussonetia papyrifera | 17 | Miscanthus floridulus | 22 | Broussonetia papyrifera | 13.6 | Bidens pilosa | 14.75 |
| Sesbania cannabina | 6.85 | Boehmeria nivea | 12.85 | Miscanthus floridulus | 7.2 | Broussonetia papyrifera | 10.43 |
| Miscanthus floridulus | 6.7 | Boehmeria densiflora | 10.2 | Neonotonia wightii | 6.7 | Boehmeria nivea | 8.15 |
| Pouzolzia elegans | 6.7 | Macroptilium atropurpureum | 7.1 | Bidens pilosa | 5.2 | Miscanthus floridulus | 5.90 |
| Eupatorium formosanum | 3.1 | Ampelopsis brevipedunculata | 7.1 | Pouzolzia elegans | 4.1 | Koelreuteria henryi | 5.03 |
| Boehmeria wattersii | 3.1 | Callicarpa formosana | 3 | Koelreuteria henryi | 3.9 | Neonotonia wightii | 4.64 |
| Boehmeria densiflora | 3.1 | Broussonetia papyrifera | 2.15 | Setaria sphacelata | 2.6 | Callicarpa pilosissima | 3.97 |
| Buddleja asiatica | 2.85 | Macroptilium atropurpureum | 2.6 | Hibiscus taiwanensis | 3.86 | ||
| Sambucus chinensis | 2.85 | Panicum maximum | 2.4 | Callicarpa formosana | 3.43 | ||
| Callicarpa pilosissima | 2.3 | Buddleja asiatica | 3.10 | ||||
| Ampelopsis brevipedunculata | 2.1 | Fraxinus griffithii | 2.47 | ||||
| Boehmeria nivea | 2.1 | Crassocephalum crepidioides | 1.93 | ||||
| Phyllostachys makinoi | 2.0 | Trema orientalis | 1.93 | ||||
| Solanum americanum | 1.9 | Setaria sphacelata | 1.71 | ||||
| Pterocypsela indica | 1.8 | Macroptilium atropurpureum | 1.50 | ||||
| Crassocephalum crepidioides | 1.8 | ||||||
| Glochidion rubrum | 1.8 | ||||||
| Nephrolepis auriculata | 1.8 | ||||||
| Callicarpa formosana | 1.8 | ||||||

4. Discussion
4.1. Relative Illuminance, Rhodes Grass Biomass and Soil-Seed Bank
4.2. Forest Restoration Processes
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lin, S.H. Vegetation Engineering of Slopeland; Wu-Nan Culture Enterprise: Taipei, Taiwan, 2013. [Google Scholar]
- Charles, M.L. A study of the vegetation in the Florida Everglades. Ecology 1959, 40, 1–9. [Google Scholar]
- Resler, L.M.; Stine, M.B. Patterns and processes of trees islands in two transitional environments: Alpine tree line and bog forest-meadow ecotones. Geogr. Compass 2009, 3, 1–26. [Google Scholar]
- Simpson, R.L.; Leck, M.A.; Parker, V.T. Ecology of Soil Seed Banks; Academic Press: New York, NY, USA, 1989; pp. 3–8. [Google Scholar]
- Chen, Y.C.; Su, Y.T.; Luo, E.C.R. Fertility effects of retained trees with vegetation development on landslide area. J. Eng. Res. Appl. 2014, 4, 77–81. [Google Scholar]
- Lai, K.S. Study on Plant Primarily Growth and Succession Mechanism following Hydroseeding Treatment of Landslide Areas. Master’s Thesis, National Chung-Hsing University, Taichung, Taiwan, 2009. [Google Scholar]
- Lin, S.H.; Su, Y.T. Vegetation development on landslide area: The fertility effect of existing tree island. J. Landsc. 2014, 19, 1–13. [Google Scholar]
- Alvarez, H.; Steinbüchel, A. Triacylglycerols in prokaryotic microorganisms. Appl. Microbiol. Biotechnol. 2002, 60, 367–376. [Google Scholar] [PubMed]
- D’Antonio, C.M.; Vitousek, P.M. Biological invasions by exotic grasses, the grass-fire cycle, and global change. Annu. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar]
- Lin, Y.S. Under Canopy Light Environment and Its Effect on Growth and Survival of Seedlings in a Broadleaf Forest at Fushan Experimental Forest. Master’s Thesis, Natural Resource Management Institute, Kent, UK, 2007. [Google Scholar]
- Lin, W.C.; Kuo, Y.L.; Chen, Y.H.; Chang, N.H.; Horng, F.W.; Ma, F.C. Soil seed bank and forest Regeneration in a Broadleaf-Coniferous Forest of Dona, Southern Taiwan. Taiwan J. For. Sci. 2004, 19, 33–42. [Google Scholar]
- Major, J.; Pyott, W.T. Buried viable seeds in two California bunchgrass sites and their bearing on the definition of a flora. Vegetation 1966, 13, 253–282. [Google Scholar]
- Nyland, R.D. Patterns of lodge pole pine regeneration following the 1988 Yellow stone fires. For. Ecol. Manag. 1998, 111, 23–33. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Adams, G.A. Effects of mobile tree islands on alpine tundra soils. Ecological 2001, 82, 8–17. [Google Scholar]
- Liao, D.C. Woody Floristic Composition of the Secondary Forest Permanent Plot of Nanjenshan Area. Master’s Thesis, National Sun Yat-sen University, Kaohsiung, Taiwan, 2000. [Google Scholar]
- Marr, J.W. The development and movement of tree islands near the upper limit of tree growth in the southern Rocky Mountains. Ecology 1977, 58, 1159–1164. [Google Scholar]
- Arno, S.F.; Hammerly, R.P. Timberline: Mountain and Arctic Forest Frontiers; The Mountaineers: Seattle, WA, USA, 1984. [Google Scholar]
- Zahawi, R.A.; Holl, K.D.; Cole, R.J.; Reid, J.L. Testing applied nucleation as a strategy to facilitate tropical forest recover. J. Appl. Ecol. 2013, 50, 88–96. [Google Scholar]
- Wetzel, P.R.; Valk, A.G.; Newman, S.; Gawlik, D.E.; Coronado-Molina, T.T.G.C.; Childers, D.L.; Sklar, F.H. Maintaining trees islands in the Florida Everglades: Nutrient redistribution is the key. Front. Ecol. Environ. 2005, 3, 370–376. [Google Scholar]
- Holl, K.D.; Zahawi, R.A.; Cole, R.J.; Ostertag, R.; Cordell, S. Planting seedlings in trees islands versus plantations as a large-scale tropical forest restoration strategy. Restor. Ecol. 2010, 19, 470–479. [Google Scholar]
- Miyawaki, A. Restoration of living environment based on vegetation ecology: Theory and practice. Ecol. Res. 2004, 19, 83–90. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-C.; Wu, C.-F.; Lin, S.-H. Mechanisms of Forest Restoration in Landslide Treatment Areas. Sustainability 2014, 6, 6766-6780. https://doi.org/10.3390/su6106766
Chen Y-C, Wu C-F, Lin S-H. Mechanisms of Forest Restoration in Landslide Treatment Areas. Sustainability. 2014; 6(10):6766-6780. https://doi.org/10.3390/su6106766
Chicago/Turabian StyleChen, Yi-Chang, Chen-Fa Wu, and Shin-Hwei Lin. 2014. "Mechanisms of Forest Restoration in Landslide Treatment Areas" Sustainability 6, no. 10: 6766-6780. https://doi.org/10.3390/su6106766
APA StyleChen, Y.-C., Wu, C.-F., & Lin, S.-H. (2014). Mechanisms of Forest Restoration in Landslide Treatment Areas. Sustainability, 6(10), 6766-6780. https://doi.org/10.3390/su6106766