The Influence of Different Cover Types on American Robin Nest Success in Organic Agroecosystems

Abstract
:1. Introduction
2. Experimental Section

{kind=link}
 |
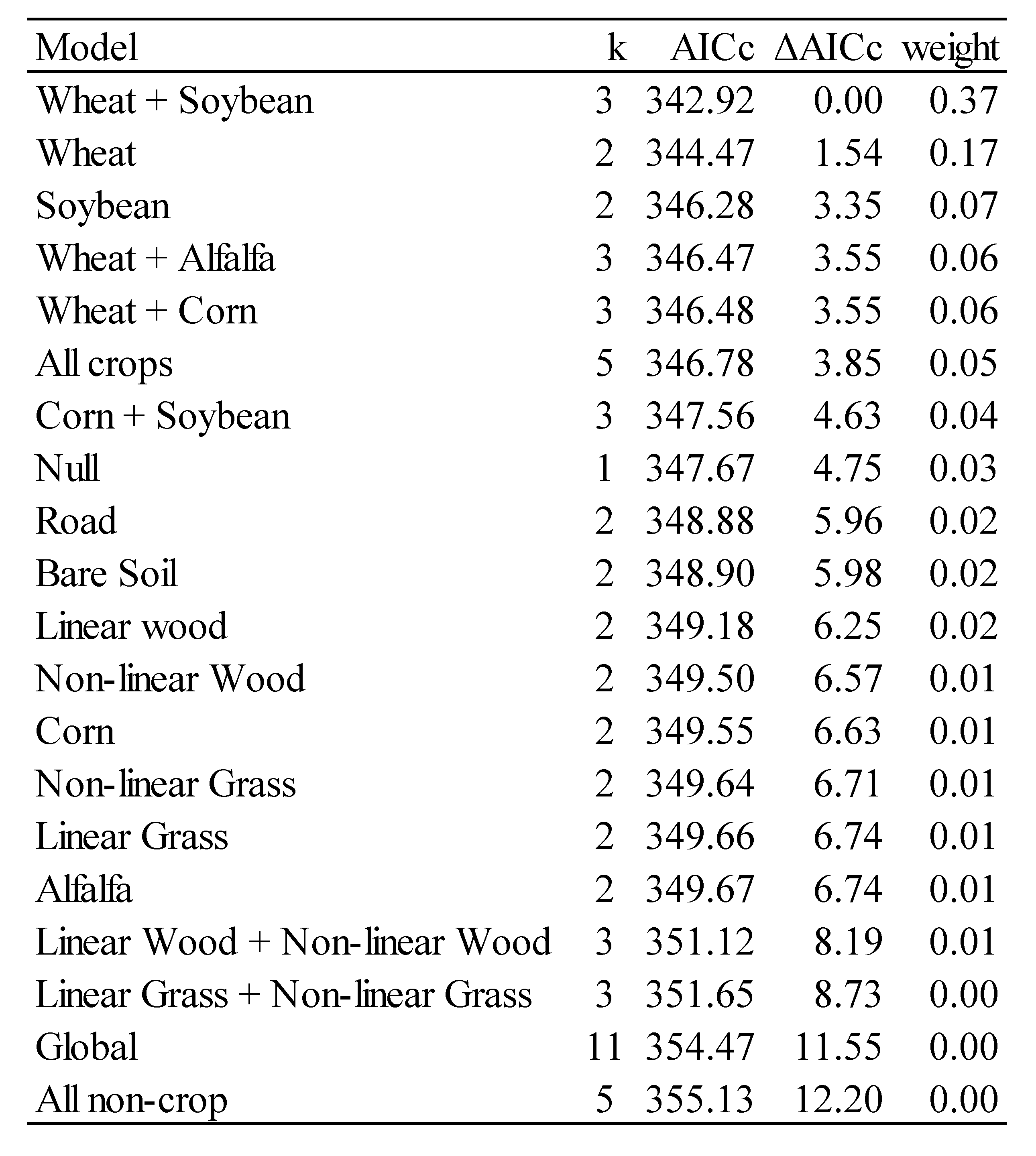 |
3. Results and Discussion
3.2. Model Results
 |
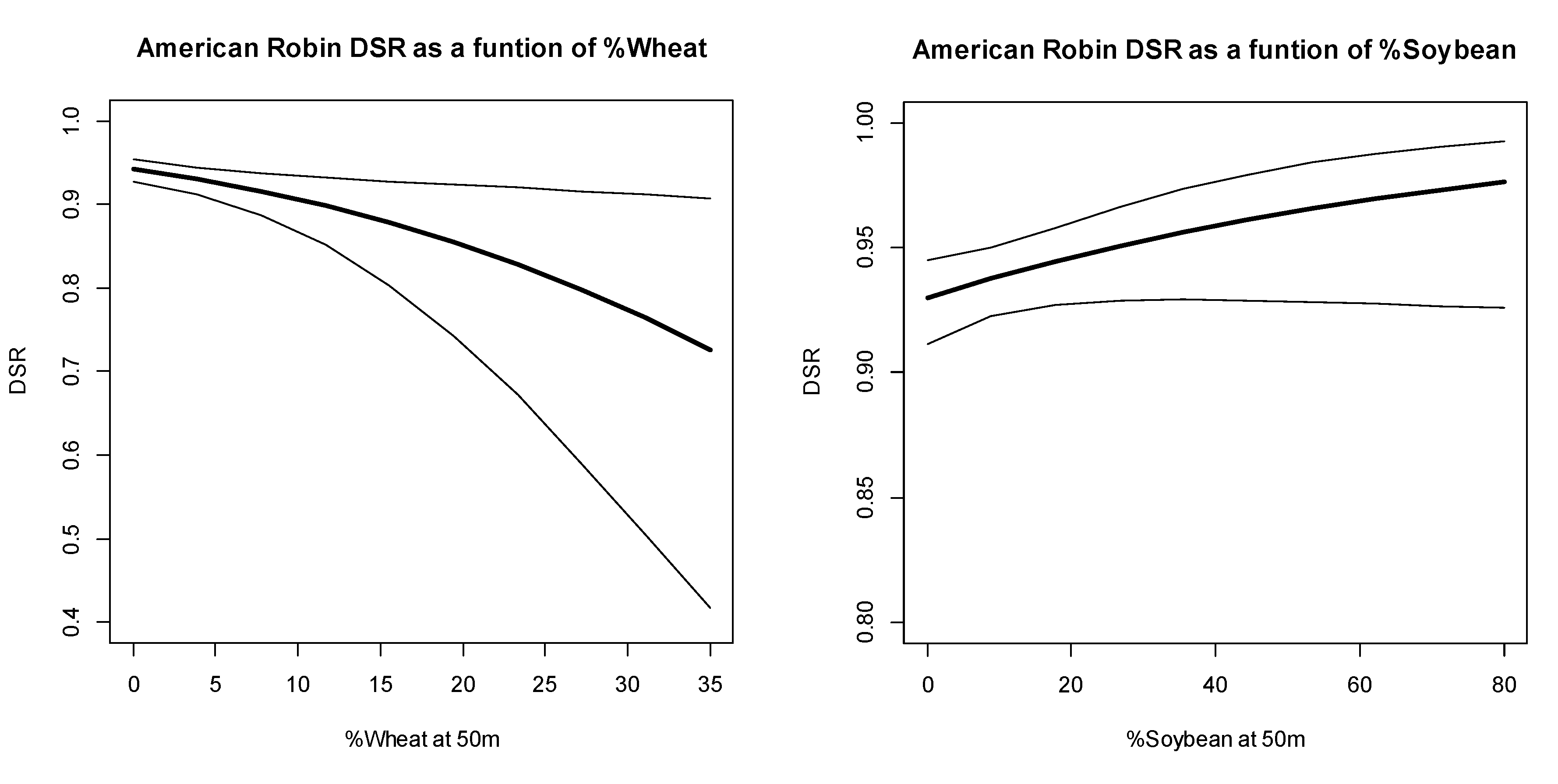
3.2. Discussion
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Butchart, S.H.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.; Almond, R.E.; Baillie, J.E.; Bomhard, B.; Brown, C.; Bruno, J. Global Biodiversity: Indicators of Recent Declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef]
- Martin, L.J.; Blossey, B.; Ellis, E. Mapping Where Ecologists Work: Biases in the Global Distribution of Terrestrial Ecological Observations. Front. Ecol. Environ. 2012, 10, 195–201. [Google Scholar] [CrossRef]
- Fischer, J.; Brosi, B.; Daily, G.C.; Ehrlich, P.R.; Goldman, R.; Goldstein, J.; Lindenmayer, D.B.; Manning, A.D.; Mooney, H.A.; Pejchar, L. Should Agricultural Policies Encourage Land Sparing or Wildlife-Friendly Farming? Front. Ecol. Environ. 2008, 6, 380–385. [Google Scholar] [CrossRef]
- Phalan, B.; Balmford, A.; Green, R.E.; Scharlemann, J.P. Minimising the Harm to Biodiversity of Producing More Food Globally. Food Policy 2011, 36, S62–S71. [Google Scholar] [CrossRef]
- Quinn, J.E. Sharing a Vision for Biodiversity Conservation and Agriculture. Renew. Agr. Food Syst. 2013, 28, 93–96. [Google Scholar] [CrossRef]
- Bengtsson, J.; Ahnström, J.; Weibull, A. The Effects of Organic Agriculture on Biodiversity and Abundance: A Meta-analysis. J. Appl. Ecol. 2005, 42, 261–269. [Google Scholar] [CrossRef]
- Hole, D.; Perkins, A.; Wilson, J.; Alexander, I.; Grice, P.; Evans, A. Does Organic Farming Benefit Biodiversity? Biol. Conserv. 2005, 122, 113–130. [Google Scholar] [CrossRef]
- Fuller, R.; Norton, L.; Feber, R.; Johnson, P.; Chamberlain, D.; Joys, A.; Mathews, F.; Stuart, R.; Townsend, M.; Manley, W. Benefits of Organic Farming to Biodiversity Vary among Taxa. Biol. Lett. UK 2005, 1, 431–434. [Google Scholar] [CrossRef]
- Beecher, N.A.; Johnson, R.J.; Brandle, J.R.; Case, R.M.; Young, L.J. Agroecology of Birds in Organic and Nonorganic Farmland. Conserv. Biol. 2002, 16, 1620–1631. [Google Scholar] [CrossRef]
- Chamberlain, D.; Wilson, J.; Fuller, R. A Comparison of Bird Populations on Organic and Conventional Farm Systems in Southern Britain. Biol. Conserv. 1999, 88, 307–320. [Google Scholar] [CrossRef]
- Dänhardt, J.; Green, M.; Lindström, A.; Rundlöf, M.; Smith, H.G. Farmland as Stopover Habitat for Migrating Birds–Effects of Organic Farming and Landscape Structure. Oikos 2010, 119, 1114–1125. [Google Scholar] [CrossRef]
- Lokemoen, J.T.; Beiser, J.A. Bird Use and Nesting in Conventional, Minimum-Tillage, and Organic Cropland. J. Wildlife Manage. 1997, 61, 644–655. [Google Scholar]
- Kragten, S.; de Snoo, G.R. Nest Success of Lapwings Vanellus vanellus on Organic and Conventional Arable Farms in the Netherlands. Ibis 2007, 149, 742–749. [Google Scholar] [CrossRef]
- Skagen, S.K.; Yackel Adams, A.A. Potential Misuse of Avian Density as a Conservation Metric. Conserv. Biol. 2011, 25, 48–55. [Google Scholar] [CrossRef]
- Best, L.B.; Freemark, K.E.; Dinsmore, J.J.; Camp, M. A Review and Synthesis of Habitat use by Breeding Birds in Agricultural Landscapes of Iowa. Am. Midl. Nat. 1995, 134, 1–29. [Google Scholar] [CrossRef]
- Riddle, J.D.; Moorman, C.E. The Importance of Agriculture-Dominated Landscapes and Lack of Field Border Effect for Early-Succession Songbird Nest Success Importance. Avian Conserv. Ecol. 2010, 5, 9. [Google Scholar]
- Bradbury, R.B.; Kyrkos, A.; Morris, A.J.; Clark, S.C.; Perkins, A.J.; Wilson, J.D. Habitat Associations and Breeding Success of Yellowhammers on Lowland Farmland. J. Appl. Ecol. 2000, 37, 789–805. [Google Scholar] [CrossRef]
- Johnson, R.J.; Jedlicka, J.A.; Quinn, J.E.; Brandle, J.R. Global Perspectives on Birds in Agricultural Landscapes. In Integrating Agriculture, Conservation and Ecotourism: Examples from the Field; Springer: New York, NY, USA, 2011; pp. 55–140. [Google Scholar]
- Francis, C.A. Conventional Research on Controversial Issues: An Exercise in Futility? Renew. Agri. Food Syst. 2010, 25, 3. [Google Scholar] [CrossRef]
- Peak, R.G.; Thompson, F.R., III; Shaffer, T.L.; Stouffer, P. Factors Affecting Songbird Nest Survival in Riparian Forests in a Midwestern Agricultural Landscape. Auk 2004, 121, 726–737. [Google Scholar]
- Balmford, A.; Green, R.; Phalan, B. What Conservationists Need to Know about Farming. P. Roy. Soc. Land B. Bio. 2012, 279, 2714–2724. [Google Scholar] [CrossRef]
- Ellis, E.C.; Ramankutty, N. Putting People in the Map: Anthropogenic Biomes of the World. Front. Ecol. Environ. 2008, 6, 439–447. [Google Scholar] [CrossRef]
- Samson, F.; Knopf, F. Prairie Conservation in North America. Bioscience 1994, 44, 418–421. [Google Scholar] [CrossRef]
- Fahrig, L. How Much Habitat is Enough? Biol. Conserv. 2001, 100, 65–74. [Google Scholar] [CrossRef]
- Koh, L.P.; Ghazoul, J. Spatially Explicit Scenario Analysis for Reconciling Agricultural Expansion, Forest Protection, and Carbon Conservation in Indonesia. Proc. Natl. Acad. Sci. USA 2010, 107, 11140–11144. [Google Scholar] [CrossRef]
- USDA National Agriculture Statistical Service. Available online: http://www.agcensus.usda.gov/Surveys/Organic_Production_Survey/index.asp (accessed on 15 May 2013).
- Willer, H.; Kilcher, L. (Eds.) The World of Organic Agriculture. Statistics and Emerging Trends 2011; IFOAM: Bonn, Germany; FiBL: Frick, Switzerland, 2011; Available online: http://www.organic-world.net/fileadmin/documents/yearbook/2011/world-of-organic-agriculture-2011-page-1-34.pdf (accessed 12 May 2013).
- Quinn, J.E.; Brandle, J.R.; Johnson, R.J. The Effects of Land Sparing and Wildlife-Friendly Practices on Grassland Bird Abundance within Organic Farmlands. Agr. Ecosyst. Environ. 2012, 161, 10–16. [Google Scholar] [CrossRef]
- Meehan, T.D.; Hurlbert, A.H.; Gratton, C. Bird Communities in Future Bioenergy Landscapes of the Upper Midwest. Proc. Natl. Acad. Sci. USA 2010, 107, 18533–18538. [Google Scholar] [CrossRef]
- Ehrlich, P.; Dobkin, D.S.; Wheye, D. Birder’s Handbook: A Field Guide to the Natural History of North American Birds; Touchstone: New York, NY, USA, 1988; p. 785. [Google Scholar]
- Haas, C.A. What Characteristics of Shelterbelts are Important to Breeding Success and Return Rate of Birds? Am. Midl. Nat. 1997, 137, 225–238. [Google Scholar] [CrossRef]
- Puckett, H.L.; Brandle, J.R.; Johnson, R.J.; Blankenship, E.E. Avian Foraging Patterns in Crop Field Edges Adjacent to Woody Habitat. Agr. Ecosyst. Environ. 2009, 131, 9–15. [Google Scholar] [CrossRef]
- Dinsmore, S.J.; White, G.C.; Knopf, F.L. Advanced Techniques for Modeling Avian Nest Survival. Ecology 2002, 83, 3476–3488. [Google Scholar] [CrossRef]
- Rotella, J.J.; Dinsmore, S.J.; Shaffer, T.L. Modeling Nest-Survival Data: A Comparison of Recently Developed Methods that can be Implemented in MARK and SAS. Anim. Biodivers. Conserv. 2004, 27, 187–205. [Google Scholar]
- Dinsmore, S.J.; Dinsmore, J.J. Modeling Avian Nest Survival in Program MARK. Stud. Avian Bio. 2007, 34, 73–83. [Google Scholar]
- Martin, T.; Paine, C.; Conway, C.; Hochachka, W.; Allen, P.; Jenkins, W. BBIRD Field Protocol. Available online: http://www.umt.edu/bbird/docs/BBIRDPROT.pdf (accessed on 6 August 2013).
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland Biodiversity: Is Habitat Heterogeneity the Key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Wilson, J.D.; Evans, J.; Browne, S.J.; King, J.R. Territory Distribution and Breeding Success of Skylarks Alauda arvensis on Organic and Intensive Farmland in Southern England. J. Appl. Ecol. 1997, 34, 1462–1478. [Google Scholar] [CrossRef]
- Shake, C.S.; Moorman, C.E.; Burchell, M.R. Cropland Edge, Forest Succession, and Landscape Affect Shrubland Bird Nest Predation. J. Wildlife Manage. 2011, 75, 825–835. [Google Scholar] [CrossRef]
- Winter, M.; Johnson, D.H.; Faaborg, J. Evidence for Edge Effects on Multiple Levels in Tallgrass Prairie. The Condor 2000, 102, 256–266. [Google Scholar] [Green Version]
- White, G.C.; Burnham, K.P. Program MARK: Survival Estimation from Populations of Marked Animals. Bird Study 1999, 46, S120–S139. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multi–Model Inference: A Practical Information–Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Powell, L.A. Approximating Variance of Demographic Parameters using the Delta Method: A Reference for Avian Biologists. The Condor 2007, 109, 949–954. [Google Scholar] [CrossRef]
- Prevedello, J.A.; Vieira, M.V. Does the Type of Matrix Matter? A Quantitative Review of the Evidence. Biodivers. Conserv. 2010, 19, 1205–1223. [Google Scholar] [CrossRef]
- Richmond, S.; Nol, E.; Burke, D. Local-Versus Landscape-Scale Effects on the Demography of Three Forest-Breeding Songbirds in Ontario, Canada. Can. J. Zool. 2012, 90, 815–828. [Google Scholar] [CrossRef]
- Wheelwright, N.T. The Diet of American Robins: An Analysis of US Biological Survey Records. Auk 1986, 103, 710–725. [Google Scholar]
- Haas, C.A. Dispersal and Use of Corridors by Birds in Wooded Patches on an Agricultural Landscape. Conserv. Biol. 1995, 9, 845–854. [Google Scholar]
- Yahner, R.H. Site-Related Nesting Success of Mourning Doves and American Robins in Shelterbelts. Wilson Bull. 1983, 95, 573–580. [Google Scholar]
- Martin, T.E. Nest Predation among Vegetation Layers and Habitat Types: Revising the Dogmas. Am. Nat. 1993, 141, 897–913. [Google Scholar]
- Latif, Q.S.; Heath, S.K.; Rotenberry, J.T. Effects of Parents and Brown-headed Cowbirds (Molothrus ater) on Nest Predation Risk for a Songbird. Ecol. Evol. 2012, 2, 3079–3097. [Google Scholar] [CrossRef]
- Skutch, A.F. Do Tropical Birds Rear as Many Young as they can Nourish? Ibis 1949, 91, 430–455. [Google Scholar] [CrossRef]
- Bonnington, C.; Gaston, K.J.; Evans, K.L. Fearing the Feline: Domestic Cats Reduce Avian Fecundity through Trait-mediated Indirect Effects that Increase Nest Predation by Other Species. J. Appl. Ecol. 2013, 50, 15–24. [Google Scholar] [CrossRef]
- Askins, R.A.; Chávez-Ramírez, F.; Dale, B.C.; Haas, C.A.; Herkert, J.R.; Knopf, F.L.; Vickery, P.D. Conservation of Grassland Birds in North America: Understanding Ecological Processes in Different Regions. Ornithological Monographs 2007, 64, iii-viii, 1-46. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Quinn, J.E.; Oden, A.; Brandle, J.R. The Influence of Different Cover Types on American Robin Nest Success in Organic Agroecosystems. Sustainability 2013, 5, 3502-3512. https://doi.org/10.3390/su5083502
Quinn JE, Oden A, Brandle JR. The Influence of Different Cover Types on American Robin Nest Success in Organic Agroecosystems. Sustainability. 2013; 5(8):3502-3512. https://doi.org/10.3390/su5083502
Chicago/Turabian StyleQuinn, John E., Amy Oden, and James R. Brandle. 2013. "The Influence of Different Cover Types on American Robin Nest Success in Organic Agroecosystems" Sustainability 5, no. 8: 3502-3512. https://doi.org/10.3390/su5083502
APA StyleQuinn, J. E., Oden, A., & Brandle, J. R. (2013). The Influence of Different Cover Types on American Robin Nest Success in Organic Agroecosystems. Sustainability, 5(8), 3502-3512. https://doi.org/10.3390/su5083502