
Impact on Grape Juice Quality and Phenolic Composition of Greek Autochthonous Grapevine Variety Mouhtaro Under Abscisic Acid Biostimulation

 ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
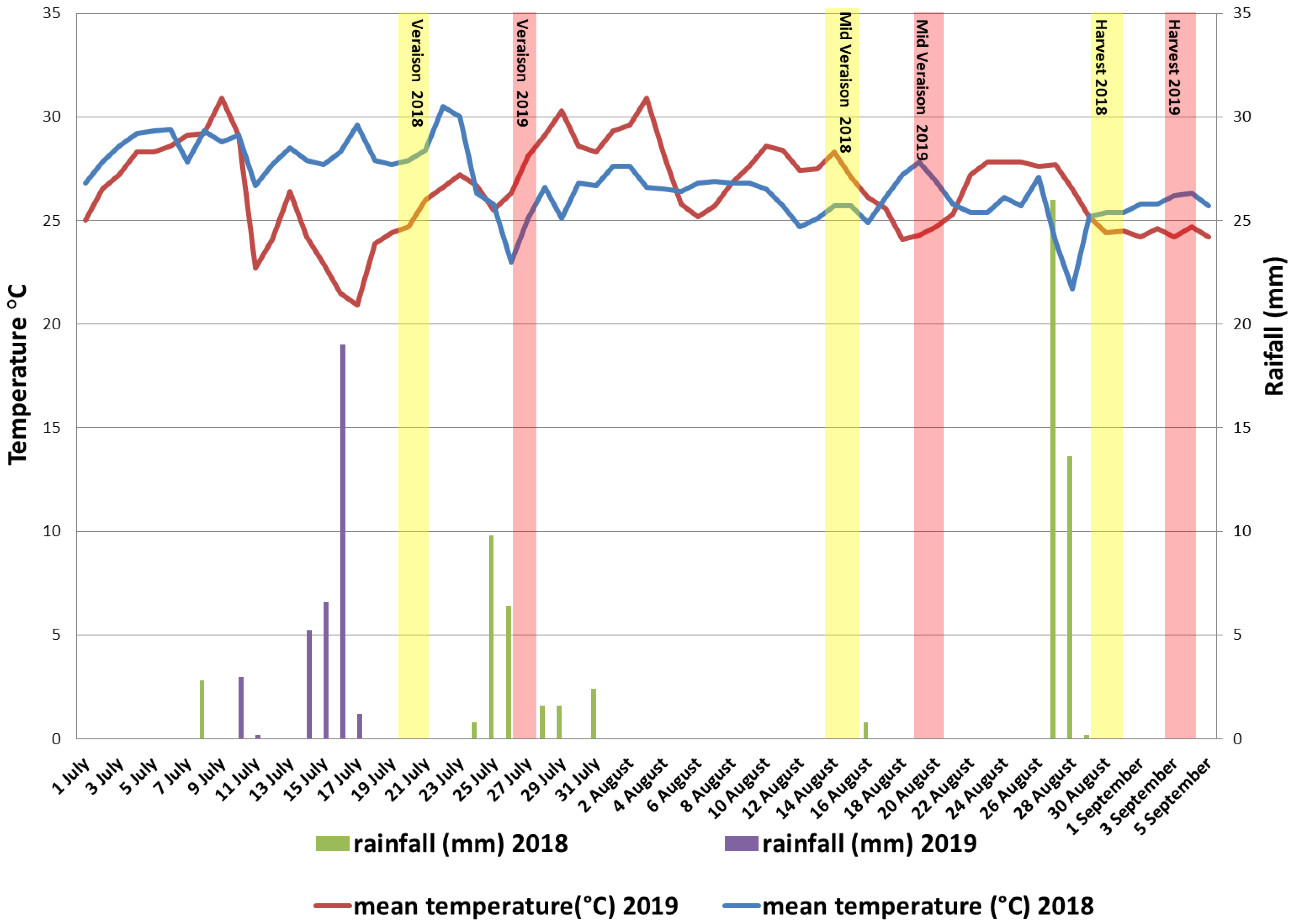
2.2. Meteorological Data
2.3. Grape Sampling and Physicochemical Must Analysis
2.4. UPLC–MS-Based Metabolic Profiling
2.5. Statistical Analysis
3. Results
3.1. Meteorological Conditions
3.2. Grape Weight and Must Conventional Analysis
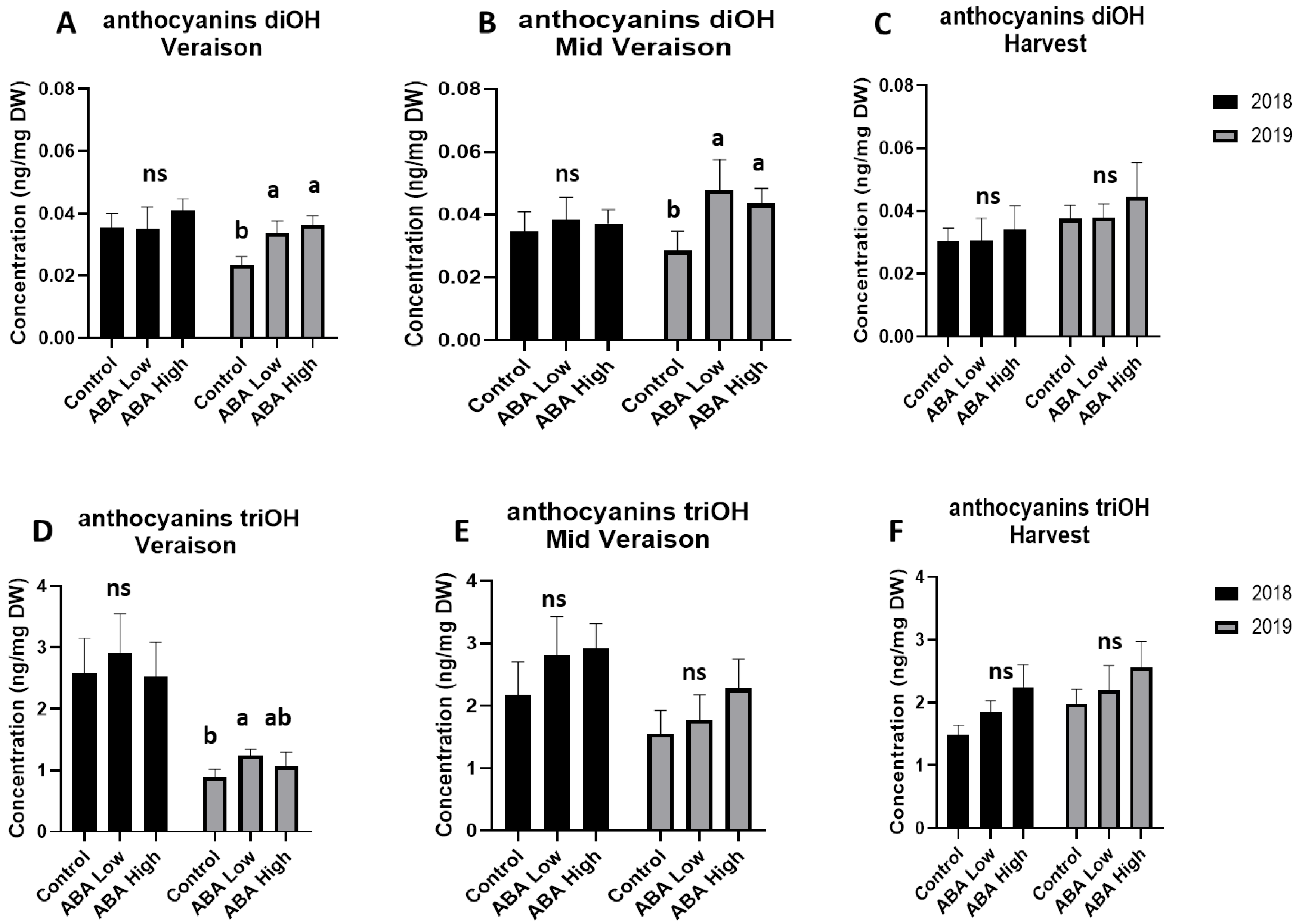
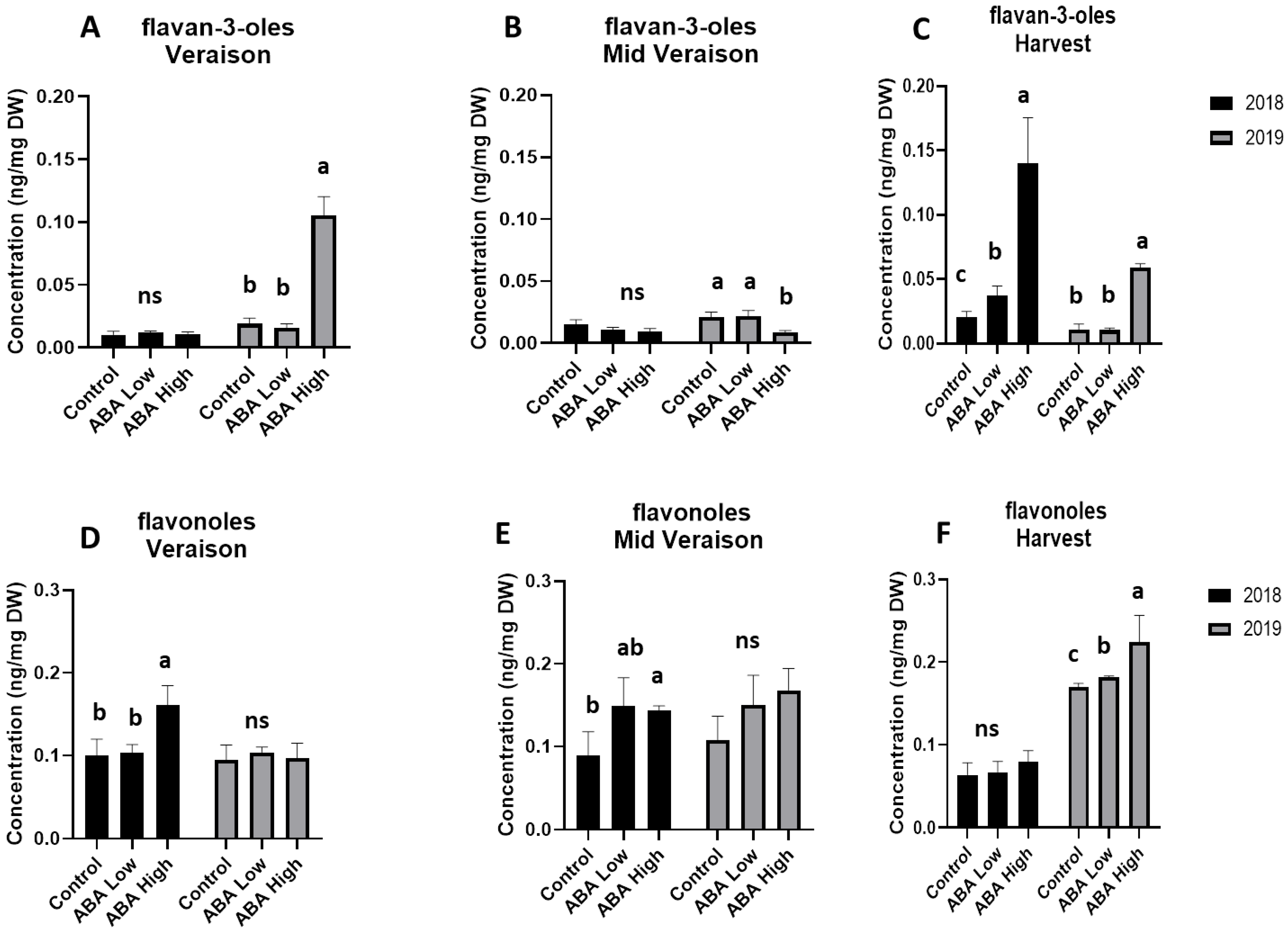
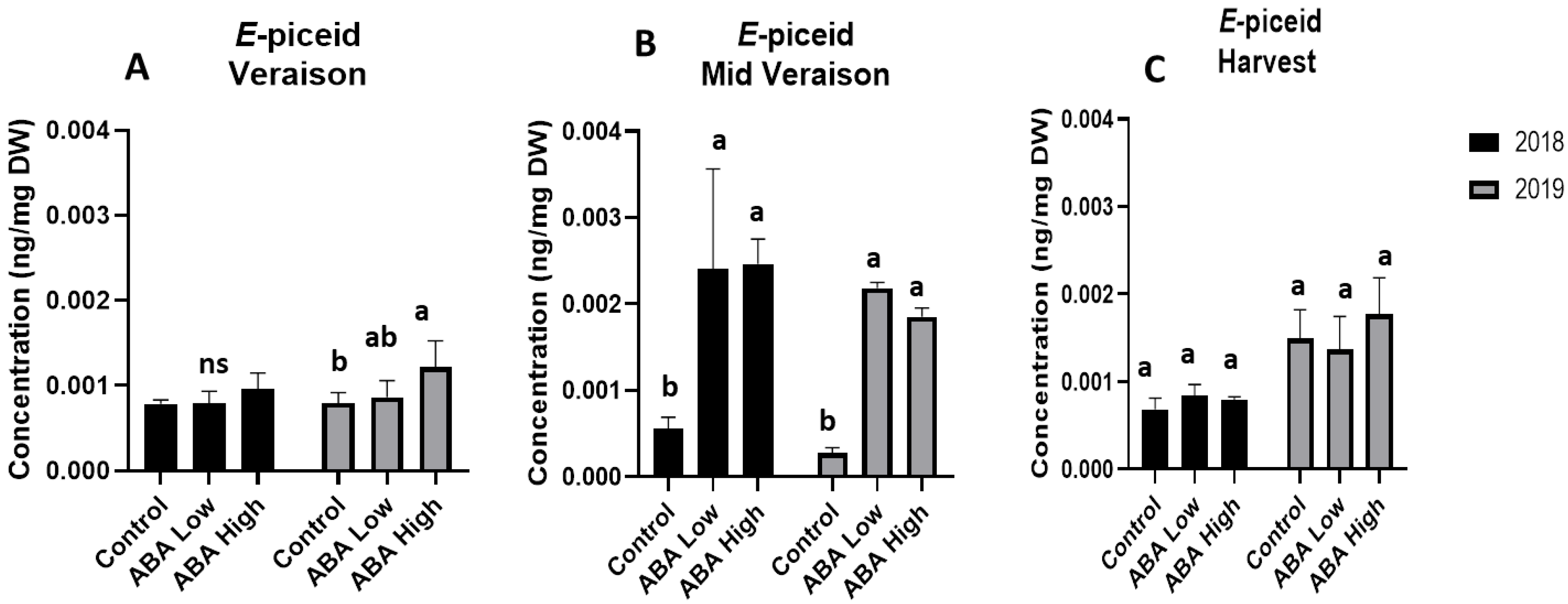
3.3. UPLC_MS Analysis of Grape Metabolites Univariate Statistics
3.4. Multivariate Statistics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cabre, F.; Mario, N.; Nuñez, M. Impacts of climate change on viticulture in Argentina. Reg. Environ. Change 2020, 20, 12. [Google Scholar] [CrossRef]
- Droulia, F.; Charalampopoulos, I. Future climate change impacts on European viticulture: A review on recent scientific advances. Atmosphere 2021, 12, 495. [Google Scholar] [CrossRef]
- Jones, G.V.; Edwards, E.J.; Bonada, M.; Sadras, V.O.; Krstic, M.; Herderich, M. Climate change and its consequences for viticulture. In Managing Wine Quality, 2nd ed.; Woodhead Publishing: Cambridge, UK, 2022; pp. 727–778. [Google Scholar] [CrossRef]
- Hannah, L.; Roehrdanz, P.R.; Ikegami, M.; Shepard, A.V.; Shaw, M.R.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R.J. Climate change, wine, and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar] [CrossRef] [PubMed]
- Drappier, J.; Thibon, C.; Rabot, A.; Geny-Denis, L. Relationship between wine composition and temperature: Impact on Bordeaux wine typicity in the context of global warming. Crit. Rev. Food Sci. Nutr. 2017, 59, 14–30. [Google Scholar] [CrossRef]
- Santos, J.A.; Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Dinis, L.-T.; Correia, C.; Moriondo, M.; Leolini, L.; Dibari, C.; Costafreda-Aumedes, S.; et al. A review of the potential climate change impacts and adaptation options for European viticulture. Appl. Sci. 2020, 10, 3092. [Google Scholar] [CrossRef]
- Ratnakumar, P.; Khan, M.L.R.; Minhas, P.S.; Farooq, M.; Sultana, R.; Per, T.S.; Deokate, P.P.; Khan, N.A.; Rane, J. Can plant bio-regulators minimize crop productivity losses caused by drought, heat and salinity stress? An integrated review. J. Appl. Bot. Food Qual. 2016, 89, 113–125. [Google Scholar] [CrossRef]
- Bernardo, S.; Dinis, L.-T.; Machado, N.; Moutinho-Pereira, J. Grapevine abiotic stress assessment and search for sustainable adaptation strategies in Mediterranean-like climates. A review. Agron. Sustain. Dev. 2018, 38, 66. [Google Scholar] [CrossRef]
- Small, C.C.; Degenhardt, D. Plant growth regulators for enhancing revegetation success in reclamation: A review. Ecol. Eng. 2018, 118, 43–51. [Google Scholar] [CrossRef]
- Dufour, M.C.; Lambert, C.; Bouscauta, J.; Merillon, J.M.; Corio-Costet, M.F. Benzothiadiazole-primed defence responses and enhanced differential expression of defence genes in Vitis vinifera infected with biotrophic pathogens Erysiphe necator and Plasmopara viticola. Plant Pathol. 2013, 62, 370–382. [Google Scholar] [CrossRef]
- Gil-Muñoz, R.; Bautista-Ortín, A.B.; Ruiz-García, Y.; Fernández-Fernández, J.I.; Gómez-Plaza, E. Improving phenolic and chromatic characteristics of Monastrell, Merlot and Syrah wines by using methyl jasmonate and benzothiadiazole. J. Int. Sci. Vigne Vin 2017, 51, 17–27. [Google Scholar] [CrossRef]
- Koyama, R.; Roberto, S.R.; de Souza, R.T.; Borges, W.F.S.; Anderson, M.; Waterhouse, A.L.; Cantu, D.; Fidelibus, M.W.; Blanco-Ulate, B. Exogenous Abscisic Acid Promotes Anthocyanin Biosynthesis and Increased Expression of Flavonoid Synthesis Genes in Vitis vinifera X Vitis labrusca Table Grapes in a Subtropical Region. Front. Plant Sci. 2018, 9, 323. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Soares, B.; Goufo, P.; Castro, I.; Cosme, F.; Pinto-Sintra, A.L.; Inês, A.; Oliveira, A.A.; Falco, V. Chitosan Upregulates the Genes of the ROS Pathway and Enhances the Antioxidant Potential of Grape (Vitis vinifera L. ‘Touriga Franca’ and ‘Tinto Cão’) Tissues. Antioxidants 2019, 8, 525. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Martins, V.; Soares, B.; Castro, I.; Falco, V. Chitosan Application in Vineyards (Vitis vinifera L. cv. Tinto Cão) Induces Accumulation of Anthocyanins and Other Phenolics in Berries, Mediated by Modifications in the Transcription of Secondary Metabolism Genes. Int. J. Mol. Sci. 2020, 21, 306. [Google Scholar] [CrossRef] [PubMed]
- Martins, V.; Unlubayir, M.; Teixeira, A.; Gerós, H.; Lanoue, A. Calcium and methyl jasmonate cross-talk in the secondary metabolism of grape cells. Plant Physiol. Biochem. 2021, 165, 228–238. [Google Scholar] [CrossRef]
- Sabir, A.; Kucukbasmaci, A.; Taytak, M.; Bilgin, O.F.; Jawshle, A.I.M.; Mohammed, O.J.M.; Gayretli, Y. Sustainable viticulture practices on the face of climate change. Agric. Res. Technol. 2018, 17, 133–137. [Google Scholar] [CrossRef]
- Koyama, L.; Sadamatsu, K.; Goto-Yamamoto, N. Abscisic acid stimulated ripening and gene expression in berry skins of the Cabernet Sauvignon grape. Funct. Integr. Genom. 2010, 10, 367–381. [Google Scholar] [CrossRef]
- Ferrandino, A.; Lovisolo, C. Abiotic stress effects on grapevine (Vitis vinifera L.): Focus on abscisic acid-mediated consequences on secondary metabolism and berry quality. Environ. Exp. Bot. 2014, 103, 138–147. [Google Scholar] [CrossRef]
- Muhammad Aslam, M.; Waseem, M.; Jakada, B.H.; Okal, E.J.; Lei, Z.; Saqib, H.S.A.; Yuan, W.; Xu, W.; Zhang, Q. Mechanisms of Abscisic Acid-Mediated Drought Stress Responses in Plants. Int. J. Mol. Sci. 2022, 23, 1084. [Google Scholar] [CrossRef]
- Peppi, M.C.; Walker, M.A.; Fidelibus, M.W. Application of abscisic acid rapidly upregulated UFGT gene expression and improved color of grape berries. Vitis J. Grapevine Res. 2008, 47, 11–14. [Google Scholar]
- Villalobos-González, L.; Peña-Neira, A.; Ibáñez, F.; Pastenes, C. Long-term effects of abscisic acid (ABA) on the grape berry phenylpropanoid pathway: Gene expression and metabolite content. Plant Physiol. Biochem. 2016, 105, 213–223. [Google Scholar] [CrossRef]
- Alonso, R.; Berli, F.J.; Fontana, A.; Piccoli, P.; Bottini, R. Abscisic acid’s role in the modulation of compounds that contribute to wine quality. Plants 2021, 10, 938. [Google Scholar] [CrossRef] [PubMed]
- Neto, F.J.D.; Junior, A.P.; Borges, C.V.; Cunha, S.R.; Callili, D.; Lima, G.P.P.; Roberto, S.R.; Leonel, S.; Tecchio, M.A. The exogenous application of abscisic acid induce accumulation of anthocyanins and phenolic compounds of the ‘Rubi’ grape. Am. J. Plant Sci. 2017, 8, 2422–2432. [Google Scholar] [CrossRef]
- Karimi, R.; Ebrahimi, M.; Amerian, M. Abscisic acid mitigates NaCl toxicity in grapevine by influencing phytochemical compounds and mineral nutrients in leaves. Sci. Hortic. 2021, 288, 110366. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Romanazzi, G.; Garde-Cerdán, T.; Pérez-Álvarez, E.P. A review of the use of biostimulants in the vineyard for improved grape and wine quality: Effects on prevention of grapevine diseases. J. Sci. Food Agric. 2019, 99, 1001–1009. [Google Scholar] [CrossRef]
- Monteiro, E.; Concalves, B.; Cortez, I.; Castro, I. The Role of biostimulants as alleviators of biotic and abiotic stresses in grapevine: A review. Plants 2022, 11, 396. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, D.H.; Eichhorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Growth Stages of the Grapevine: Phenological growth stages of the grapevine (Vitis vinifera L. ssp. vinifera)—Codes and descriptions according to the extended BBCH scale. Aust. J. Grape Wine Res. 1995, 1, 100–103. [Google Scholar] [CrossRef]
- Miliordos, D.E.; Alatzas, A.; Kontoudakis, N.; Kouki, A.; Unlubayir, M.; Gémin, M.-P.; Tako, A.; Hatzopoulos, P.; Lanoue, A.; Kotseridis, Y. Abscisic Acid and Chitosan Modulate Polyphenol Metabolism and Berry Qualities in the Domestic White-Colored Cultivar Savvatiano. Plants 2022, 11, 1648. [Google Scholar] [CrossRef]
- Lagouvardos, K.; Kotroni, V.; Bezes, A.; Koletsis, I.; Kopania, T.; Lykoudis, S.; Mazarakis, N.; Papagiannaki, K.; Vougioukas, S. The automatic weather stations NOANN network of the National Observatory of Athens: Operation and database. Geosci. Data J. 2017, 4, 4–16. [Google Scholar] [CrossRef]
- OIV. Compendium of International Methods of Analysis of Wines and Musts, Methods, OIVMA–AS313–01; OIV–MA–AS2–10; International Organisation of Vine and Wine: Paris, France, 2018; Volume 2, Available online: https://www.oiv.int/public/medias/3731/oiv-ma-as313-01.pdf (accessed on 31 March 2025).
- Billet, K.; Delanoue, G.; Arnault, I.; Besseau, S.; Oudin, A.; Courdavault, V.; Marchand, P.A.; Giglioli-Guivarc’h, N.; Guérin, L.; LaNoue, A. Vineyard evaluation of stilbenoid-rich grape cane extracts against downy mildew: A large-scale study. Pest Manag. Sci. 2018, 75, 1252–1257. [Google Scholar] [CrossRef]
- Miliordos, D.-E.; Tsiknia, M.; Kontoudakis, N.; Zaharias, N.; Kotseridis, Y. A holistic approach to using biostimulants on the red grapevine variety, Mouhtaro—from grape to wine: Sourced from the research article: “Impact of Application of Abscisic Acid, Benzothiadiazole and Chitosan on Berry Quality Characteristics and Plant Associated Microbial Communities of Vitis vinifera L var. Mouhtaro Plants.” (Sustainability, 2021). Original language of the article: English. IVES Tech. Rev. Vine Wine 2022. [Google Scholar] [CrossRef]
- Jeong, S.; Goto-Yamamoto, N.; Kobayashi, S.; Esaka, M. Effects of plant hormones and shading on the accumulation of anthocyanins and the expression of anthocyanin biosynthetic genes in grape berry skins. Plant Sci. 2004, 167, 247–252. [Google Scholar] [CrossRef]
- Cocco, M.; Mercenaro, L.; Lo Cascio, M.; Nieddu, G. Effects of Vine Water Status and Exogenous Abscisic Acid on Berry Composition of Three Red Wine Grapes Grown under Mediterranean Climate. Horticulturae 2020, 6, 12. [Google Scholar] [CrossRef]
- Miliordos, D.E.; Alatzas, A.; Kontoudakis, N.; Unlubayir, M.; Hatzopoulos, P.; Lanoue, A.; Kotseridis, Y. Benzothiadiazole Affects Grape Polyphenol Metabolism and Wine Quality in Two Greek Cultivars: Effects during Ripening Period over Two Years. Plants 2023, 12, 1179. [Google Scholar] [CrossRef] [PubMed]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef]
- Malinowska, M.; Billet, K.; Drouet, S.; Munsch, T.; Unlubayir, M.; Tungmunnithum, D.; Giglioli-Guivarch, N.; Hano, C.; Lanoue, A. Grape cane extracts as multifunctional rejuvenating cosmetic ingredient: Evaluation of sirtuin activity, tyrosinase inhibition and bioavailability potential. Molecules 2020, 25, 2203. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.; Silva, A.M. Plant flavonoids: Chemical characteristics and biological activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Ruiz-García, Y.; Gil-Muñoz, R.; López-Roca, J.M.; Martínez-Cutillas, A.; Romero-Cascales, I.; Gómez-Plaza, E. Increasing the Phenolic Compound Content of Grapes by Preharvest Application of Abcisic Acid and a Combination of Methyl Jasmonate and Benzothiadiazole. J. Agric. Food Chem. 2013, 61, 3978–3983. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Q.; Xi, B.; Dai, H. Study on the regulation of anthocyanin biosynthesis by exogenous abscisic acid in grapevine. Sci. Hortic. 2019, 250, 294–301. [Google Scholar] [CrossRef]
- Alenazi, M.M.; Shafiq, M.; Alobeed, R.S.; Alsdon, A.A.; Abbasi, N.A.; Ali, I.; Mubushar, M.; Javed, I. Application of abscisic acid at véraison improves red pigmentation and accumulation of dietary antioxidants in red table grapes cv. Red Globe at harvest. Sci. Hortic. 2019, 257, 108672. [Google Scholar] [CrossRef]
- He, L.; Ren, Z.-Y.; Wang, Y.; Fu, Y.-Q.; Li, Y.; Meng, N.; Pan, Q.-H. Variation of Growth-to-Ripening Time Interval Induced by Abscisic Acid and Synthetic Auxin Affecting Transcriptome and Flavor Compounds in Cabernet Sauvignon Grape Berry. Plants 2020, 9, 630. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, Y.; Wang, S.; Wang, W.; Xu, X.; Wu, J.; Fang, Y.; Ju, Y. Effects of strigolactone and abscisic acid on the quality and antioxidant activity of grapes (Vitis vinifera L.) and wines. Food Chem. X 2022, 16, 100496. [Google Scholar] [CrossRef] [PubMed]
- Malovini, E.; Arancibia, C.; Durán, M.; Fontana, A.; de Rosas, M.I.; Deis, L.; Gargantini, R.; Bottini, R.; Cavagnaro, B.; Martínez, L. Abscisic acid and methyl jasmonic acid module anthocyanins and trans-resveratrol accumulation in berry skin of five red Vitis vinifera cultivars in two contrasting viticultural regions of mendoza-Argentina. Rev. Fac. Cienc. Agrar. 2019, 51, 451–460. [Google Scholar]
- Paladines-Quezada, D.F.; Moreno-Olivares, J.D.; Fernández-Fernández, J.I.; Bleda-Sánchez, J.A.; Martínez-Moreno, A.; Gil-Muñoz, R. Elicitors and Pre-Fermentative Cold Maceration: Effects on Polyphenol Concentration in Monastrell Grapes and Wines. Biomolecules 2019, 9, 671. [Google Scholar] [CrossRef] [PubMed]
- Luan, L.Y.; Zhang, Z.W.; Xi, Z.M.; Huo, S.S.; Ma, L.N. Comparing the effects of exogenous abscisic acid on the phenolic composition of Yan 73 and Cabernet Sauvignon (Vitis vinifera L.) wines. Eur. Food Res. Technol. 2014, 239, 203–213. [Google Scholar] [CrossRef]
- Xi, Z.M.; Meng, J.F.; Huo, S.S.; Luan, L.Y.; Ma, L.N.; Zhang, Z.W. Exogenously applied abscisic acid to Yan73 (V. vinifera) grapes enhances phenolic content and antioxidant capacity of its wine. Int. J. Food Sci. Nutr. 2013, 64, 444–451. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, M.; Feng, M.; Liu, G.; Torregrosa, L.; Tao, X.; Xu, T. miR156b-targeted VvSBP8/13 functions downstream of the abscisic acid signal to regulate anthocyanins biosynthesis in grapevine fruit under drought. Hortic. Res. 2024, 11, uhad293. [Google Scholar] [CrossRef]
- Katayama-Ikegami, A.; Sakamoto, T.; Shibuya, K.; Katayama, T.; Gao-Takai, M. Effects of abscisic acid treatment on berry coloration and expression of flavonoid biosynthesis genes in grape. Am. J. Plant Sci. 2016, 7, 1325. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2018 | |||||
|---|---|---|---|---|---|
| Stage | Treatment | Berry Weight (mg/berry) | Total Soluble Solids (°Brix) | pH | Total Acidity (g/L of Tartaric Acid) |
| Véraison | Control | 1.14 ± 0.15 a | 14.33 ± 0.16 a | 2.96 ± 0.05 a | 15.32 ± 1.32 a |
| ABA Low | 1.13 ± 0.09 a | 14 ± 0.07 a | 2.97 ± 0.09 a | 15.67 ± 1.12 a | |
| ABA High | 1.17 ± 0.10 a | 14.16 ± 0.22 a | 2.90 ± 0.10 a | 15 ± 0.98 a | |
| Middle Véraison | Control | 1.76 ± 0.18 a | 19.70 ± 1.62 a | 3.61 ± 0.22 ab | 11.1 ± 1.08 a |
| ABA Low | 1.73 ± 0.11 a | 18.33 ± 0.85 b | 3.33 ± 0.16 b | 12.8 ± 0.91 a | |
| ABA High | 1.74 ± 0.08 a | 16.83 ± 1.15 c | 3.71 ± 0.21 b | 12.5 ± 1.20 a | |
| Harvest | Control | 1.98 ± 0.04 a | 21.90 ± 0.77 a | 3.71 ± 0.12 b | 6.9 ± 0.21 b |
| ABA Low | 1.90 ± 0.10 ab | 19.79 ± 1.01 b | 3.69 ± 0.17 b | 7.80 ± 0.14 a | |
| ABA High | 1.89 ± 0.03 b | 17.23 ± 1.06 c | 3.90 ± 0.11 a | 7.5 ± 0.25 a | |
| 2019 | |||||
| Véraison | Control | 1.27 ± 0.08 a | 10.00 ± 0.20 a | 2.42 ± 0.01 a | 12.35 ± 0.48 a |
| ABA Low | 1.37 ± 0.09 a | 10.40 ± 0.17 a | 2.43 ± 0.03 a | 13.47 ± 0.31 a | |
| ABA High | 1.33 ± 0.10 a | 9.83 ± 0.66 a | 2.42 ± 0.03 a | 13.27 ± 0.44 a | |
| Middle Véraison | Control | 1.91 ± 0.34 a | 19.6 ± 0.50 a | 3.50 ± 0.10 a | 8.98 ± 0.20 a |
| ABA Low | 1.82 ± 0.06 a | 19.13 ± 0.85 a | 3.60 ± 0.06 a | 8.99 ± 0.35 a | |
| ABA High | 1.81 ± 0.26 a | 18.13 ± 1.05 a | 3.43 ± 0.17 a | 9.95 ± 0.79 a | |
| Harvest | Control | 2.24 ± 0.05 a | 23.36 ± 0.28 a | 3.31 ± 0.03 ab | 6.25 ± 0.08 c |
| ABA Low | 2.19 ± 0.01 ab | 22.82 ± 0.65 ab | 3.34 ± 0.06 a | 6.88 ± 0.07 a | |
| ABA High | 2.15 ± 0.05 b | 22.95 ± 0.05 b | 3.24 ± 0.01 b | 6.69 ± 0.08 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miliordos, D.-E.; Alatzas, A.; Kontoudakis, N.; Unlubayir, M.; Nikolakis, K.; Hatzopoulos, P.; Lanoue, A.; Kotseridis, Y. Impact on Grape Juice Quality and Phenolic Composition of Greek Autochthonous Grapevine Variety Mouhtaro Under Abscisic Acid Biostimulation. Sustainability 2025, 17, 4385. https://doi.org/10.3390/su17104385
Miliordos D-E, Alatzas A, Kontoudakis N, Unlubayir M, Nikolakis K, Hatzopoulos P, Lanoue A, Kotseridis Y. Impact on Grape Juice Quality and Phenolic Composition of Greek Autochthonous Grapevine Variety Mouhtaro Under Abscisic Acid Biostimulation. Sustainability. 2025; 17(10):4385. https://doi.org/10.3390/su17104385
Chicago/Turabian StyleMiliordos, Dimitrios-Evangelos, Anastasios Alatzas, Nikolaos Kontoudakis, Marianne Unlubayir, Konstantinos Nikolakis, Polydefkis Hatzopoulos, Arnaud Lanoue, and Yorgos Kotseridis. 2025. "Impact on Grape Juice Quality and Phenolic Composition of Greek Autochthonous Grapevine Variety Mouhtaro Under Abscisic Acid Biostimulation" Sustainability 17, no. 10: 4385. https://doi.org/10.3390/su17104385
APA StyleMiliordos, D.-E., Alatzas, A., Kontoudakis, N., Unlubayir, M., Nikolakis, K., Hatzopoulos, P., Lanoue, A., & Kotseridis, Y. (2025). Impact on Grape Juice Quality and Phenolic Composition of Greek Autochthonous Grapevine Variety Mouhtaro Under Abscisic Acid Biostimulation. Sustainability, 17(10), 4385. https://doi.org/10.3390/su17104385