


Tomato and Pepper Seeds as Pathways for the Dissemination of Phytopathogenic Bacteria: A Constant Challenge for the Seed Industry and the Sustainability of Crop Production




Abstract
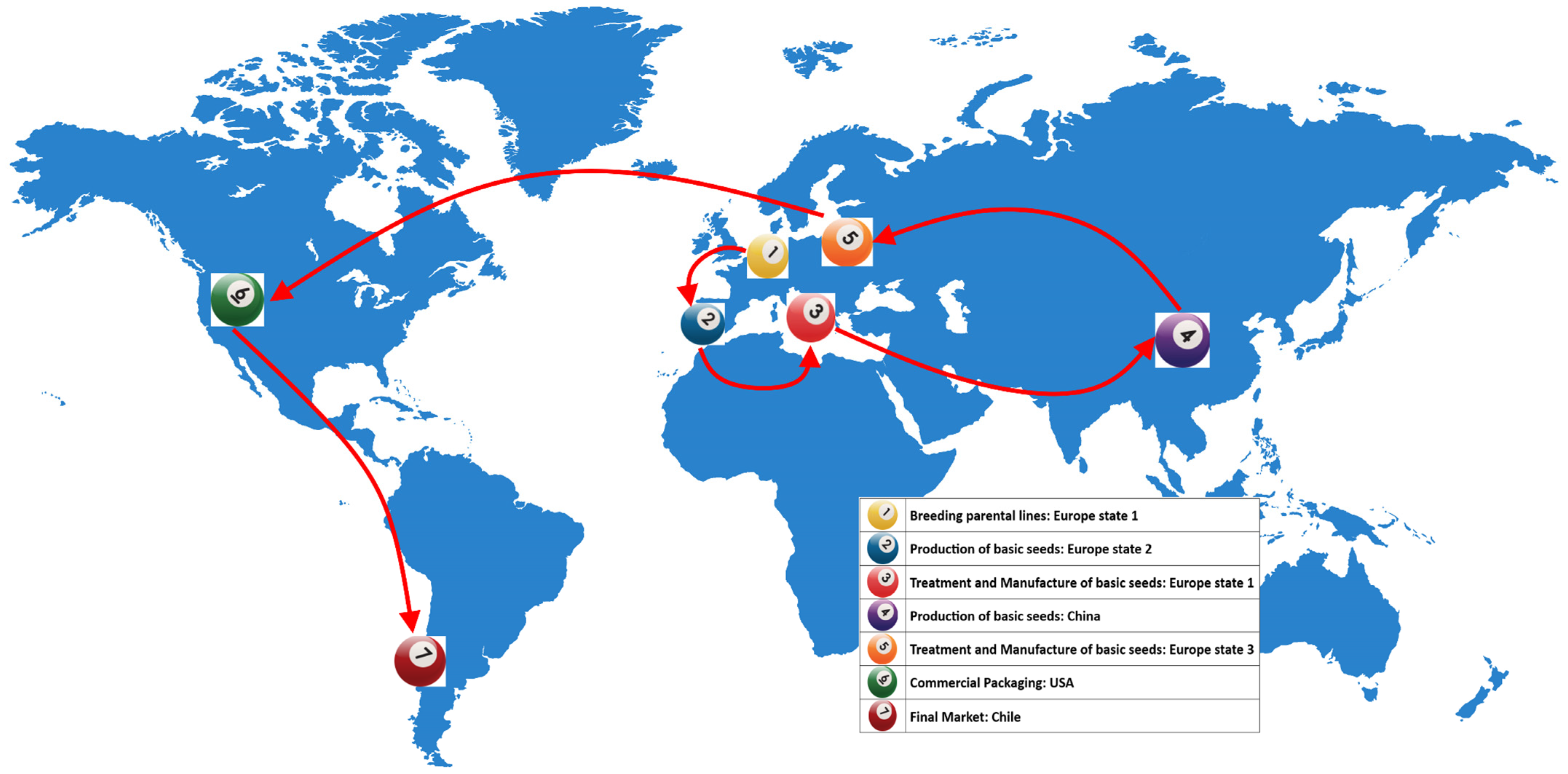
1. Introduction

2. Seed Endophytes: Recruitment and Role
3. Phytopathogenic Bacteria in Tomato and Pepper Seeds
3.1. Clavibacter michiganensis subsp. michiganensis (Smith) Davis et al. and Clavibacter capsici (Oh et al.) Li et al.
3.2. Pseudomonas syringae pv. tomato (Okabe) Young, Dye, and Wilkie
3.3. Xanthomonads: Xanthomonas vesicatoria (Doidge) Vauterin, Hoste, Kersters, and Swings, X. euvesicatoria pv. euvesicatoria (Jones et al.) Constantin et al.; X. euvesicatoria pv. perforans (Jones et al.) Constantin et al., and X. hortorum pv. gardneri (Jones et al.) Morinière et al.
4. How Phytopathogenic Clavibacters, Pseudomonas syringae pv. Tomato, and Xanthomonads Colonize Tomato and Pepper Seeds
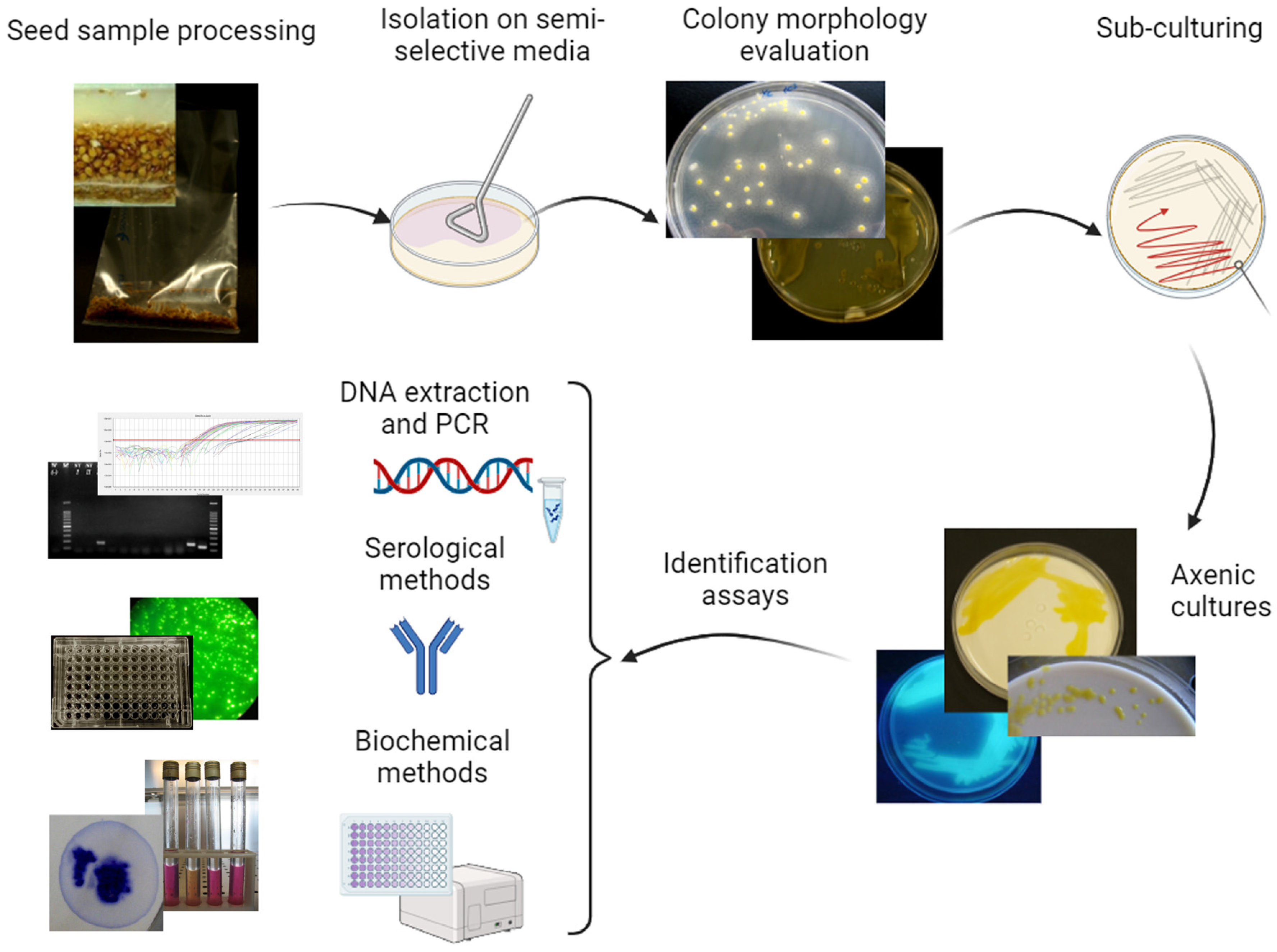
5. Detection of Phytopathogenic Bacteria in Tomato and Pepper Seeds
5.1. Direct Isolation on Agar Media
5.2. Serological Detection
5.3. Molecular Detection
6. Seed Treatments
6.1. Tomato and Pepper Seed Extraction Procedures
6.2. Seed Sanitation Methods and Procedures
6.2.1. Chemical Seed Treatments
6.2.2. Physical Seed Treatments
Heat Treatments
Ozone
UV-C Light Irradiation
6.2.3. Microorganisms and Natural Products
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Louwaars, N.P.; Manicad, G. Seed Systems Resilience—An Overview. Seeds 2022, 1, 340–356. [Google Scholar] [CrossRef]
- FAO Glossary of Phytosanitary Terms, ISPM 5. Available online: https://www.fao.org/3/mc891e/mc891e.pdf (accessed on 19 December 2023).
- Seed and GM Crop Market Analysis. Available online: https://www.spglobal.com/ratings/en/research-insights/topics/outlook-2023 (accessed on 17 July 2023).
- Dongyu, Q.U. A Statement by FAO Director-General of FAO. Available online: https://www.fao.org/director-general/speeches/detail/en/c/1321173/ (accessed on 4 November 2023).
- Malhotra, B. Global Vegetable Seeds Market Is Increasingly Fragmented and Diversified; S&P Global—Commodity Insights: London, UK, 2021; Available online: https://www.spglobal.com/commodityinsights/en/ci/research-analysis/global-vegetable-seeds-market.html (accessed on 3 November 2023).
- OECD Seed Schemes. Rules and Regulations; Organization for Economic Co-Operation and Development (OECD). 2023. Available online: https://www.oecd.org/agriculture/seeds/rules-regulations/ (accessed on 14 February 2024).
- Liang, D.; Tang, W.; Fu, Y. Sustainable Modern Agricultural Technology Assessment by a Multistakeholder Transdisciplinary Approach. IEEE Trans. Eng. Manag. 2023, 70, 1061–1075. [Google Scholar] [CrossRef]
- The Business Research Company. Tomato Seed Global Market Report; The Business Research Company: Dublin, Ireland, 2023; Available online: https://www.thebusinessresearchcompany.com/report/tomato-seeds-global-market-report (accessed on 6 November 2023).
- FAO. International Standard for Phytosanitary Measures n°38. International Movement of Seeds. 2018. Available online: http://www.fao.org/3/i7219en/i7219en.pdf (accessed on 11 December 2023).
- Oh, E.J.; Bae, C.; Lee, H.B.; Hwang, I.S.; Lee, H.I.; Yea, M.C.; Yim, K.O.; Lee, S.; Heu, S.; Cha, J.S.; et al. Clavibacter michiganensis subsp. capsici subsp. Nov., Causing Bacterial Canker Disease in Pepper. Int. J. Syst. Evol. Microbiol. 2016, 66, 4065–4070. [Google Scholar] [CrossRef]
- Wilson, D. Endophyte: The Evolution of a Term, and Clarification of Its Use and Definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Chee-Sanford, J.C.; Williams, M.M.; Davis, A.S.; Sims, J.K. Do Microorganisms Influence Seed-Bank Dynamics? Weed Sci. 2006, 54, 575–587. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; van Themaat, E.V.L.; Ahmadinejad, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E.; et al. Revealing Structure and Assembly Cues for Arabidopsis Root-Inhabiting Bacterial Microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.B. Microbial Dynamics and Interactions in the Spermosphere. Ann. Rev. Phytopathol. 2004, 42, 271–309. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.K.; Sinclair, J.B. Principles of Seed Pathology; eBook; CRC Press: Boca Raton, FL, USA, 1996; pp. 1–560. ISBN 9780429152856. [Google Scholar] [CrossRef]
- Malfanova, N.; Lugtenberg, B.J.J.; Berg, G. Molecular Microbial Ecology of the Rhizosphere; Wiley-Blackwell: Hoboken, NJ, USA, 2013. [Google Scholar]
- Santoyo, G. How Plants Recruit Their Microbiome? New Insights into Beneficial Interactions. J. Adv. Res. 2021, 40, 45–58. [Google Scholar] [CrossRef]
- Herre, E.A.; Knowlton, N.; Mueller, U.G.; Rehner, S.A. The Evolution of Mutualisms: Exploring the Paths between Conflict and Cooperation. Trends Ecol. Evol. 1999, 14, 49–53. [Google Scholar] [CrossRef]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic Potential of Endophytic Bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Brader, G.; Compant, S.; Vescio, K.; Mitter, B.; Trognitz, F.; Ma, L.-J.; Sessitsch, A. Ecology and Genomic Insights into Plant-Pathogenic and Plant-Nonpathogenic Endophytes. Ann. Rev. Phytopathol. 2017, 55, 61–83. [Google Scholar] [CrossRef] [PubMed]
- Bashir, M.A.; Silvestri, C.; Ahmad, T.; Hafiz, I.A.; Abbasi, N.A.; Manzoor, A.; Cristofori, V.; Rugini, E. Osmotin: A Cationic Protein Leads to Improve Biotic and Abiotic Stress Tolerance in Plants. Plants 2020, 9, 992. [Google Scholar] [CrossRef] [PubMed]
- Samreen, T.; Naveed, M.; Nazir, M.Z.; Asghar, H.N.; Khan, M.I.; Zahir, Z.A.; Kanwal, S.; Jeevan, B.; Sharma, D.; Meena, V.S.; et al. Seed Associated Bacterial and Fungal Endophytes: Diversity, Life Cycle, Transmission, and Application Potential. Appl. Soil Ecol. 2021, 168, 104–161. [Google Scholar] [CrossRef]
- Bergna, A.; Cernava, T.; Rändler, M.; Grosch, R.; Zachow, C.; Berg, G. Tomato Seeds Preferably Transmit Plant Beneficial Endophytes. Phytobiome J. 2018, 2, 183–193. [Google Scholar] [CrossRef]
- Yildirim, K.C.; Orel, D.C.; Okyay, H.; Gursan, M.M.; Demir, I. Quality of Immature and Mature Pepper (Capsicum annuum L.) Seeds in Relation to Bio-Priming with Endophytic Pseudomonas and Bacillus spp. Horticulturae 2021, 7, 75. [Google Scholar] [CrossRef]
- Corrigendum—PM 7/42 (3) Clavibacter michiganensis subsp. michiganensis. EPPO Bull. 2022, 53, 148. [CrossRef]
- PM 7/110 (2) Xanthomonas spp. (Xanthomonas euvesicatoria pv. euvesicatoria, Xanthomonas hortorum pv. gardneri, Xanthomonas euvesicatoria pv. perforans, Xanthomonas vesicatoria) Causing Bacterial Spot of Tomato and Sweet Pepper. EPPO Bull. 2023, 53, 558–579. [CrossRef]
- PM 7/21 (2) Ralstonia solanacearum, R. pseudosolanacearum and R. syzygii (Ralstonia solanacearum species Complex). EPPO Bull. 2018, 48, 32–63. [CrossRef]
- Sen, Y.; van der Wolf, J.; Visser, R.G.F.; van Heusden, S. Bacterial Canker of Tomato: Current Knowledge of Detection, Management, Resistance, and Interactions. Plant Dis. 2015, 99, 4–13. [Google Scholar] [CrossRef]
- Peritore-Galve, F.C.; Tancos, M.A.; Smart, C.D. Bacterial Canker of Tomato: Revisiting a Global and Economically Damaging Seedborne Pathogen. Plant Dis. 2021, 105, 1581–1595. [Google Scholar] [CrossRef]
- Osdaghi, E.; Rahimi, T.; Taghavi, S.M.; Ansari, M.; Zarei, S.; Portier, P.; Briand, M.; Jacques, M.-A. Comparative Genomics and Phylogenetic Analyses Suggest Several Novel Species within the Genus Clavibacter, Including Nonpathogenic Tomato-Associated Strains. Appl. Environ. Microbiol. 2020, 86, e02873-19. [Google Scholar] [CrossRef]
- Yim, K.-O.; Lee, H.-I.; Kim, J.-H.; Lee, S.-D.; Cho, J.-H.; Cha, J.-S. Characterization of Phenotypic Variants of Clavibacter michiganensis subsp. michiganensis Isolated from Capsicum annuum. Eur. J. Plant Pathol. 2012, 133, 559–575. [Google Scholar] [CrossRef][Green Version]
- Nandi, M.; MacDonald, J.; Liu, P.; Weselowski, B.; Yuan, Z. Clavibacter michiganensis subsp. michiganensis: Bacterial Canker of Tomato, Molecular Interactions and Disease Management. Mol. Plant Pathol. 2018, 19, 2036–2050. [Google Scholar] [CrossRef]
- Chang, R.J.; Ries, S.M.; Pataky, J.K. Dissemination of Clavibacter michiganensis subsp. michiganensis by Practices Used to Produce Tomato Transplants. Phytopathology 1991, 81, 1276–1281. [Google Scholar] [CrossRef]
- Jones, J.B.; Zitter, T.A.; Momol, T.M.; Miller, S.A. Compendium of Tomato Diseases and Pests, 2nd ed.; APS Press: St. Paul, MN, USA, 2014; pp. 1–168. ISBN 978-0-89054-434-1. [Google Scholar]
- Quesada-Ocampo, L.M.; Landers, N.A.; Lebeis, A.C.; Fulbright, D.W.; Hausbeck, M.K.; Sen, Y.; Aysan, Y.; Mirik, M.; Ozdemir, D.; Meijer-Dekens, F.; et al. Genetic Structure of Clavibacter michiganensis subsp. michiganensis Populations in Michigan Commercial Tomato Fields. Plant Dis. 2012, 96, 788–796. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Plant Health. Scientific Opinion on the Pest Categorisation of Clavibacter michiganensis subsp. michiganensis (Smith) Davis et al. EFSA J. 2014, 12, 3721. [Google Scholar] [CrossRef]
- El-Fatah, B.A.; Imran, M.; Abo-Elyousr, K.; Mahmoud, A. Isolation of Pseudomonas syringae Pv. Tomato Strains Causing Bacterial Speck Disease of Tomato and Marker-Based Monitoring for Their Virulence. Mol. Biol. Rep. 2023, 50, 4917–4930. [Google Scholar] [CrossRef]
- Wilson, M.; Campbell, H.L.; Ji, P.; Jones, J.B.; Cuppels, D.A. Biological Control of Bacterial Speck of Tomato under Field Conditions at Several Locations in North America. Phytopathology 2002, 92, 1284–1292. [Google Scholar] [CrossRef]
- Vasileva, K.; Ganeva, D.; Bogatzevska, N. Species Composition of the Bacterial Population Colonizing Tomato Flowers. Bulg. J. Agr. Sci. 2022, 28, 677–690. [Google Scholar]
- Cement, A.; Saygili, H.; Horuz, S.; Aysan, Y. Potential of Bacteriophages to Control Bacterial Speck of Tomato (Pseudomonas syringae pv. tomato). Fresenius Env. Bull. 2018, 27, 9366–9373. [Google Scholar]
- Preston, G.M. Pseudomonas syringae pv. tomato: The Right Pathogen, of the Right Plant, at the Right Time. Mol. Plant Pathol. 2000, 1, 263–275. [Google Scholar] [CrossRef]
- Santamaría-Hernando, S.; López-Maroto, A.; Galvez-Roldán, C.; Munar-Palmer, M.; Monteagudo-Cascales, E.; Rodríguez-Herva, J.-J.; Krell, T.; López-Solanilla, E. Pseudomonas syringae pv. tomato Infection of Tomato Plants Is Mediated by GABA and l-Pro Chemoperception. Mol. Plant Pathol. 2022, 23, 1433–1445. [Google Scholar] [CrossRef] [PubMed]
- Devash, Y.; Bashan, Y.; Okon, Y.; Henis, Y. Survival of Pseudomonas tomato in Soil and Seeds. J. Phytopathol. 1979, 60, 597–601. [Google Scholar] [CrossRef]
- Bashan, Y.; Diab, S.; Okon, Y. Survival of Xanthomonas campestris pv. vesicatoria in pepper seeds and roots in symptomless and dry leaves in non-host plants and in the soil. Plant Soil 1982, 68, 161–170. [Google Scholar] [CrossRef]
- Scortichini, M.; Stefani, E.; Elphinstone, J.G.; Vlami, M.B. PM 7/110 (1) Xanthomonas spp. (Xanthomonas euvesicatoria, Xanthomonas gardneri, Xanthomonas perforans, Xanthomonas vesicatoria) Causing Bacterial Spot of Tomato and Sweet Pepper. EPPO Bull. 2013, 43, 7–20. [Google Scholar] [CrossRef]
- Barak, J.D.; Vancheva, T.; Lefeuvre, P.; Jones, J.B.; Timilsina, S.; Minsavage, G.V.; Vallad, G.E.; Koebnik, R. Whole-Genome Sequences of Xanthomonas euvesicatoria Strains Clarify Taxonomy and Reveal a Stepwise Erosion of Type 3 Effectors. Front. Plant Sci. 2016, 7, 1805. [Google Scholar] [CrossRef]
- Zitter, T.A. Pepper Disease Control It Starts with the Seed; Department of Plant Pathology, Cornell University: Ithaca, NY, USA, 2004; p. 14853. Available online: https://www.northeastipm.org/ipm-in-action/ipmresources/resource-detail/?id=4345 (accessed on 28 October 2023).
- Jones, J.B.; Bouzar, H.; Stall, R.E.; Almira, E.C.; Roberts, P.D.; Bowen, B.W.; Sudberry, J.; Strickler, P.M.; Chun, J. Systematic Analysis of Xanthomonads (Xanthomonas Spp.) Associated with Pepper and Tomato Lesions. Int. J. Syst. Evol. Microbiol. 2000, 50, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Dutta, B.; Gitaitis, R.; Smith, S.; Langston, D.B. Interactions of Seedborne Bacterial Pathogens with Host and Non-Host Plants in Relation to Seed Infestation and Seedling Transmission. PLoS ONE 2014, 9, e99215. [Google Scholar] [CrossRef] [PubMed]
- Giovanardi, D.; Biondi, E.; Ignjatov, M.; Jevtic, R.; Stefani, E. Impact of Bacterial Spot Outbreaks on the Phytosanitary Quality of Tomato and Pepper Seeds. Plant Pathol. 2018, 67, 1168–1176. [Google Scholar] [CrossRef]
- Osdaghi, E. Xanthomonas euvesicatoria pv. euvesicatoria (Bacterial Spot of Tomato and Pepper). CABI Compend. 2022. [Google Scholar] [CrossRef]
- Frank, A.C.; Guzmán, J.P.S.; Shay, J.E. Transmission of Bacterial Endophytes. Microorganisms 2017, 5, 70. [Google Scholar] [CrossRef] [PubMed]
- Barret, M.; Guimbaud, J.F.; Darrasse, A.; Jacques, M.A. Plant Microbiota Affects Seed Transmission of Phytopathogenic Microorganisms. Mol. Plant Pathol. 2016, 17, 791–795. [Google Scholar] [CrossRef]
- Bashan, Y. Long-Term Survival of Pseudomonas syringae pv. tomato and Xanthomonas campestris pv. vesicatoria in Tomato and Pepper Seeds. Phytopathology 1982, 72, 1143–1144. [Google Scholar] [CrossRef]
- Bashan, Y. Detection of Cutinases and Pectic Enzymes During Infection of Tomato by Pseudomonas syringae pv. tomato. Phytopathology 1985, 75, 940. [Google Scholar] [CrossRef]
- Tancos, M.A.; Chalupowicz, L.; Barash, I.; Manulis-Sasson, S.; Smart, C.D. Tomato Fruit and Seed Colonization by Clavibacter Michiganensis Subsp. Michiganensis through External and Internal Routes. Appl. Environ. Microbiol. 2013, 79, 6948–6957. [Google Scholar] [CrossRef]
- Dutta, B.; Avci, U.; Hahn, S.K.; Hahn, M.G.; Walcott, R. Location of Acidovorax citrulli in Infested Watermelon Seeds Is Influenced by the Pathway of Bacterial Invasion. Phytopathology 2012, 102, 461–468. [Google Scholar] [CrossRef]
- Dutta, B.; Ha, Y.; Lessl, J.T.; Avci, U.; Sparks, A.C.; Johnson, K.L.; Walcott, R.R. Pathways of Bacterial Invasion and Watermelon Seed Infection by Acidovorax citrulli. Plant Pathol. 2015, 64, 537–544. [Google Scholar] [CrossRef]
- Gitaitis, R.D.; Walcott, R. The Epidemiology and Management of Seedborne Bacterial Diseases. Ann. Rev. Phytopathol. 2007, 45, 371–397. [Google Scholar] [CrossRef]
- Koike, H. The Aluminum-Cap Method for Testing Sugarcane Varieties against Leaf Scald Disease. Phytopathology 1965, 55, 317–319. [Google Scholar]
- Schaad, N.W.; Franken, A.A.J.M. ISTA Handbook on Seed Health Testing, Working Sheet No. 50, 2nd ed.; ISTA: Zurich, Switzerland, 1996. [Google Scholar]
- Sijam, K.; Chang, C.J.; Gitaitis, R.D. A Medium for Differentiating Tomato and Pepper Strains of Xanthomonas campestris pv. vesicatoria. Can. J. Plant Pathol. 1992, 14, 182–184. [Google Scholar] [CrossRef]
- Alvarez, A.M. Integrated Approaches for Detection of Plant Pathogenic Bacteria And Diagnosis Of Bacterial Diseases. Ann. Rev. Phytopathol. 2004, 42, 339–366. [Google Scholar] [CrossRef]
- Schaad, N.W. Detection of Seedborne Bacterial Plant Pathogens. Plant Dis. 1982, 66, 885–890. [Google Scholar] [CrossRef]
- Chu, P.; Waterhouse, P.; Martin, R.; Gerlach, W. New Approaches to the Detection of Microbial Plant Pathogens. BGER 1989, 7, 45–112. [Google Scholar] [CrossRef]
- De León, L.; Rodríguez, A.; López, M.M.; Siverio, F. Evaluation of the Efficacy of Immunomagnetic Separation for the Detection of Clavibacter michiganensis subsp. michiganensis in Tomato Seeds. J. Appl. Microbiol. 2008, 104, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Ovod, V.V.; Rudolph, K.; Krohn, K. Serological Classification of Pseudomonas syringae Pathovars Based on Mononoclonal Antibodies Towards the Lipopolysaccharide O-Chains. J. Bacteriol. 1997, 9, 526–531. [Google Scholar] [CrossRef]
- Pastrik, K.H.; Rainey, F.A. Identification and Differentiation of Clavibacter michiganensis Subspecies by Polymerase Chain Reaction-Based Techniques. J. Phytopathol. 1999, 147, 687–693. [Google Scholar] [CrossRef]
- Oosterhof, J.; Berendsen, S. The Development of a Specific Real-Time TaqMan for the Detection of Clavibacter michiganensis subsp. michiganensis. In Proceedings of the APS-IPPC Meeting, Honolulu, HI, USA, 6–10 August 2011; Available online: https://www.apsnet.org/meetings/Documents/2011_Meeting_Abstracts/a11ma777.htm (accessed on 12 December 2023).
- Wu, Y.-D.; Chen, L.-H.; Wu, X.-J.; Shang, S.; Lou, J.; Du, L.-Z.; Zhao, Z. Gram Stain-Specific-Probe-Based Real-Time PCR for Diagnosis and Discrimination of Bacterial Neonatal Sepsis. J. Clin. Microbiol. 2008, 46, 2613–2619. [Google Scholar] [CrossRef]
- Yasuhara-Bell, J.; Baysal-Gurel, F.; Miller, S.A.; Alvarez, A.M. Utility of a Loop-Mediated Amplification Assay for Detection of Clavibacter michiganensis subsp. michiganensis in Seeds and Plant Tissues. Can. J. Plant Pathol. 2015, 37, 260–266. [Google Scholar] [CrossRef]
- Yasuhara-Bell, J.; Kubota, R.; Jenkins, D.M.; Alvarez, A.M. Loop-Mediated Amplification of the Clavibacter michiganensis subsp. michiganensis MicA Gene Is Highly Specific. Phytopathology 2013, 103, 1220–1226. [Google Scholar] [CrossRef]
- Morcia, C.; Piazza, I.; Ghizzoni, R.; Terzi, V.; Carrara, I.; Bolli, G.; Chiusa, G. Molecular Diagnostics in Tomato: Chip Digital PCR Assays Targeted to Identify and Quantify Clavibacter michiganensis subsp. michiganensis and Ralstonia solanacearum in Planta. Horticulturae 2023, 9, 553. [Google Scholar] [CrossRef]
- Han, S.; Jiang, N.; Lv, Q.; Kan, Y.; Hao, J.; Li, J.; Luo, L. Detection of Clavibacter michiganensis subsp. michiganensis in Viable but Nonculturable State from Tomato Seed Using Improved qPCR. PLoS ONE 2018, 13, e0196525. [Google Scholar] [CrossRef] [PubMed]
- Koenraadt, H.; Van Betteray, B.; Germain, R.; Hiddink, G.; Jones, J.B.; Oosterhof, J. Development Of Specific Primers For The Molecular Detection Of Bacterial Spot Of Pepper And Tomato. Acta Hortic. 2009, 808, 99–102. [Google Scholar] [CrossRef]
- Strayer, A.L.; Jeyaprakash, A.; Minsavage, G.V.; Timilsina, S.; Vallad, G.E.; Jones, J.B.; Paret, M.L. A Multiplex Real-Time PCR Assay Differentiates Four Xanthomonas Species Associated with Bacterial Spot of Tomato. Plant Dis. 2016, 100, 1660–1668. [Google Scholar] [CrossRef] [PubMed]
- Strayer-Scherer, A.; Jones, J.B.; Paret, M.L. Recombinase Polymerase Amplification Assay for Field Detection of Tomato Bacterial Spot Pathogens. Phytopathology 2019, 109, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Zaccardelli, M.; Spasiano, A.; Bazzi, C.; Merighi, M. Identification and in Planta Detection of Pseudomonas syringae pv. tomato Using PCR Amplification of hrpZPst. Eur. J. Plant Pathol. 2005, 111, 85–90. [Google Scholar] [CrossRef]
- Fanelli, V.; Cariddi, C.; Finetti-Sialer, M. Selective Detection of Pseudomonas syringae pv. tomato Using Dot Blot Hybridization and Real-time PCR. Plant Pathol. 2007, 56, 683–691. [Google Scholar] [CrossRef]
- Jiang, N.; Lv, Q.; Xu, X.; Walcott, R.; Li, J.; Luo, L. Induction of the Viable but Nonculturable State in Clavibacter michiganensis subsp. michiganensis and in Planta Resuscitation of the Cells on Tomato Seedlings. Plant Pathol. 2016, 65, 826–836. [Google Scholar] [CrossRef]
- Wang, H.; Wagnon, R.; Moreno, D.; Timilsina, S.; Jones, J.; Vallad, G.; Turechek, W.W. A Long-Amplicon Viability-qPCR Test for Quantifying Living Pathogens That Cause Bacterial Spot in Tomato Seed. Plant Dis. 2022, 106, 1474–1485. [Google Scholar] [CrossRef]
- Denancé, N.; Grimault, V. Seed Pathway for Pest Dissemination: The ISTA Reference Pest List, a Bibliographic Resource in Non-vegetable Crops. EPPO Bull. 2022, 52, 434–445. [Google Scholar] [CrossRef]
- Strider, D.L. Bacterial Canker of Tomato Caused by Corynebacterium michiganense: A Literature Review and Bibliography. Technical Bull. N. Carol. Agric. Exp. Stn. 1969, 193, 1–110. [Google Scholar]
- GSPP Standard for Tomato Seed and Young Plant Production Sites (Valid from 1st June 2022). 2022. Available online: https://www.Gspp.Eu/Images/Documents/GSPP_Standard_V3.3.pdf (accessed on 9 December 2023).
- Management of Seed-Borne Diseases: An Integrated Approach. In Seed-Borne Diseases of Agricultural Crops: Detection, Diagnosis & Management; Kumar, R., Gupta, A., Eds.; Springer: Singapore, 2020; pp. 717–745. ISBN 978-981-329-045-7. [Google Scholar]
- Jones, J.B.; Pohronezny, K.L.; Stall, R.E.; Jones, J.P. Survival of Xanthomonas campestris pv. vesicatoria in Florida on Tomato Crop Residue, Weeds, Seeds, and Volunteer Tomato Plants. Phytopathology 1986, 76, 430–434. [Google Scholar] [CrossRef]
- European Commission. Commission Implementing Regulation (EU) 2018/1981 of 13 December 2018; OJ L 317, 14.12.2018; European Commission: Brussels, Belgium, 2018; pp. 16–20. [Google Scholar]
- Cooksey, D.A. Genetics of Bactericide Resistance in Plant Pathogenic Bacteria. Ann. Rev. Phytopathol. 1990, 28, 201–219. [Google Scholar] [CrossRef]
- Jones, J.B.; Lacy, G.H.; Bouzar, H.; Stall, R.E.; Schaad, N.W. Reclassification of the Xanthomonads Associated with Bacterial Spot Disease of Tomato and Pepper. Syst. Appl. Microbiol. 2004, 27, 755–762. [Google Scholar] [CrossRef]
- Griffin, K.; Gambley, C.F.; Brown, P.H.; Li, Y. Copper-Tolerance in Pseudomonas syringae pv. tomato and Xanthomonas spp. and the Control of Diseases Associated with These Pathogens in Tomato and Pepper. A Systematic Literature Review. Crop Prot. 2017, 96, 144–150. [Google Scholar] [CrossRef]
- Stall, R.E.; Jones, J.B.; Minsavage, G.V. Durability of resistance in tomato and pepper to Xanthomonads causing bacterial spot. Ann. Rev. Phytopathol. 2009, 47, 265–284. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Gao, Z.; Yang, W. Breeding for resistance to tomato bacterial diseases in China: Challenges and prospects. Hort. Plant J. 2018, 4, 193–207. [Google Scholar] [CrossRef]
- Adhikari, P.; Adhikari, T.B.; Louws, F.J.; Panthee, D.R. Advances and challenges in bacterial spot resistance breeding in tomato (Solanum lycopersicum L.). Int. J. Mol. Sci. 2020, 21, 1734. [Google Scholar] [CrossRef] [PubMed]
- Van Der Plank, J.E. Plant Diseases: Epidemics and Control. Soil Sci. 1964, 98, 279. [Google Scholar] [CrossRef]
- Bryan, M.K. Studies on Bacterial Canker of Tomato. J. Agric. Res. 1930, 41, 825–851. [Google Scholar]
- Shoemaker, P.B.; Echandi, E. Seed and Plant Bed Treatments for Bacterial Canker of Tomato. Plant Dis. Rep. 1976, 60, 163–166. [Google Scholar]
- Pyke, N.B.; Milne, K.S.; Neilson, H.F. Tomato Seed Treatments for the Control of Bacterial Speck. N. Z. J. Exp. Agr. 1984, 12, 161–164. [Google Scholar] [CrossRef]
- Murata, A.; Numata, I. Heat Endurance of Corynebacterium michiganense and Tomato Seeds to Dry Seeds. Proc. Kanto Pl. Protocol Soc. 1970, 17, 55–56. [Google Scholar]
- Cho, M.; Kim, J.; Kim, J.Y.; Yoon, J.; Kim, J.-H. Mechanisms of Escherichia Coli Inactivation by Several Disinfectants. Water Res. 2010, 44, 3410–3418. [Google Scholar] [CrossRef]
- Mbega, E.R.; Mortensen, C.N.; Mabagala, R.B.; Wulff, E.G. The Effect of Plant Extracts as Seed Treatments to Control Bacterial Leaf Spot of Tomato in Tanzania. J. Gen. Plant Pathol. 2012, 78, 277–286. [Google Scholar] [CrossRef]
- Kotan, R.; Dadasoğlu, F.; Karagoz, K.; Cakir, A.; Ozer, H.; Kordali, S.; Cakmakci, R.; Dikbas, N. Antibacterial Activity of the Essential Oil and Extracts of Satureja hortensis against Plant Pathogenic Bacteria and Their Potential Use as Seed Disinfectants. Sci. Hortic. 2013, 153, 34–41. [Google Scholar] [CrossRef]
- Karabuyuk, F.; Aysan, Y. Aqueous Plant Extracts as Seed Treatments on Tomato Bacterial Speck Disease. Acta Hortic. 2018, 1207, 193–196. [Google Scholar] [CrossRef]
- Umesha, S.; Kavitha, R. Prevalence of Bacterial Spot in Tomato Fields of Karnataka and Effect of Biological Seed Treatment on Disease Incidence. Crop Prot. 2006, 25, 375–381. [Google Scholar] [CrossRef]
- Bashan, Y.; de-Bashan, L.E. Protection of Tomato Seedlings against Infection by Pseudomonas syringae Pv. Tomato by Using the Plant Growth-Promoting Bacterium Azospirillum brasilense. Appl. Environ. Microbiol. 2002, 68, 2637–2643. [Google Scholar] [CrossRef] [PubMed]
- Kritzman, G.A. Chemi-Thermal Treatment for Control of Seedborne Bacterial Pathogens of Tomato. Phytoparasitica 1993, 21, 101–109. [Google Scholar] [CrossRef]
- Bashan, Y.; de-Bashan, L.E. Reduction of Bacterial Speck (Pseudomonas syringae pv. tomato) of Tomato by Combined Treatments of Plant Growth-Promoting Bacterium, Azospirillum brasilense, Streptomycin Sulfate, and Chemo-Thermal Seed Treatment. Eur. J. Plant Pathol. 2002, 108, 821–829. [Google Scholar] [CrossRef]
- Sanogo, S.; Clary, M. Bacterial Leaf Spot of Chile Pepper: A Short Guide for Growers. New Mexico State University, College of Agriculture and Home Economics: Las Cruces, NM, USA. 2008. Available online: https://pubs.nmsu.edu/research/horticulture/NMCA30/index.html (accessed on 21 December 2023).
- McFarquhar, J. Organic Seed Treatments for the Reduction of Xanthomonas euvesicatoria on Tomato Seed. Master’s Thesis, University of Georgia, Athens, GA, USA, 2015. Available online: http://getd.libs.uga.edu/pdfs/mcfarquhar_judith_201505_ms.pdf (accessed on 12 December 2023).
- Mtui, H.D.; Bennett, M.A.; Maerere, A.P.; Miller, S.A.; Kleinhenz, M.D.; Sibuga, K.P. Effect of Seed Treatments and Mulch on Seedborne Bacterial Pathogens and Yield of Tomato (Solanum lycopersicum Mill.) in Tanzania. J. Anim. Plant Sci. 2010, 8, 1006–1015. [Google Scholar]
- Lordon, M. Sweet Pepper Breeding and Seed Saving Guide, Department of Agriculture: USDA, Minnesota. 2022. Available online: https://seedalliance.org/crops/peppers/ (accessed on 25 November 2023).
- Welbaum, G. Tomato. Publisher: VCE Publication, Department of Horticulture, Virginia Tech: Blacksburg, VA. 2021. Available online: https://pubs.ext.vt.edu/426/426-418/mobile-version.html (accessed on 25 November 2023).
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; ISBN 9780120445653. [Google Scholar]
- Ryan, R.P.; Vorhölter, F.-J.; Potnis, N.; Jones, J.B.; Van Sluys, M.-A.; Bogdanove, A.J.; Dow, J.M. Pathogenomics of Xanthomonas: Understanding Bacterium-Plant Interactions. Nat. Rev. Microbiol. 2011, 9, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Sauer, D.B.; Burroughs, R. Disinfection of Seed Surfaces with Sodium Hypochlorite. Phytopathology 1986, 76, 745–749. [Google Scholar] [CrossRef]
- Raval, A.; Sasidharan, N.; Rao, K. Effect of seed extraction procedures on seed quality parameters in tomato. Adv. Life Sci. 2016, 5, 9020–9024. [Google Scholar]
- Degwale, A.; Tesfa, T.; Meseret, B.; Fantaw, S. Seed Extraction Methods Affect the Physiological Quality of Tomato Seed and Developing Seedlings. Int. J. Veg. Sci. 2022, 29, 16–24. [Google Scholar] [CrossRef]
- Agarwal, V.K.; Sinclair, J.B. Principles of Seed Pathology, 2nd ed.; Lewis Publisher: Boca Raton, FL, USA, 1997; ISBN 978-0-87371-670-3. [Google Scholar]
- Ercolani, G.L. Effettività e Misura Della Trasmissione Di Xanthomonas vesicatoria e Di Corynebacterium michiganense Attraverso Il Seme Del Pomodoro. Ind. Conserve 1968, 43, 14–22. [Google Scholar]
- Chambers, S.; Merriman, P. Perennation and Control of Pseudomonas Tomato in Victoria. Crop Pasture Sci. 1975, 26, 657–663. [Google Scholar] [CrossRef]
- Pradhanang, P.M.; Collier, G. How Effective Is Hydrochloric Acid Treatment to Control Clavibacter michiganensis subsp. michiganensis Contamination in Tomato Seed. Acta Hortic. 2009, 808, 81–85. [Google Scholar] [CrossRef]
- McCormack, J. Pepper Seed Production. 2005. Available online: https://www.carolinafarmstewards.org/wp-content/uploads/2012/05/PepperSeedProductionVer1_2.pdf (accessed on 10 January 2024).
- Carisse, O.; Ouimet, A.; Toussaint, V.; Philion, V. Philion Evaluation of the Effect of Seed Treatments, Bactericides, and Cultivars on Bacterial Leaf Spot of Lettuce Caused by Xanthomonas campestris pv. vitians. Plant Dis. 2000, 84, 295–299. [Google Scholar] [CrossRef]
- Kasselaki, A.-M.; Goumas, D.E.; Tamm, L.; Fuchs, J.; Fuchs, J.G.; Cooper, J.; Leifert, C. Effect of Alternative Strategies for the Disinfection of Tomato Seed Infected with Bacterial Canker (Clavibacter michiganensis subsp. michiganensis). NJAS Wagening. J. Life Sci. 2011, 58, 145–147. [Google Scholar] [CrossRef]
- Cardarelli, M.; Woo, S.L.; Rouphael, Y.; Colla, G. Seed Treatments with Microorganisms Can Have a Biostimulant Effect by Influencing Germination and Seedling Growth of Crops. Plants 2022, 11, 259. [Google Scholar] [CrossRef]
- Ayesha, M.S.; Suryanarayanan, T.S.; Nataraja, K.N.; Prasad, S.R.; Shaanker, R.U. Seed Treatment with Systemic Fungicides: Time for Review. Front. Plant Sci. 2021, 12, 654512. [Google Scholar] [CrossRef]
- Vasanthakumari, M.M.; Shridhar, J.; Madhura, R.J.; Nandhitha, M.; Kasthuri, C.; Janardhana, B.; Nataraja, K.N.; Ravikanth, G.; Shaanker, R.U. Role of endophytes in early seedling growth of plants: A test using systemic fungicide seed treatment. Plant Physiol. Rep. 2019, 24, 86–95. [Google Scholar] [CrossRef]
- van der Wolf, J.M.; Birnbaum, Y.E.; van der Zouwen, P.S.; Groot, S.P.C.; Groot, S.P.C. Disinfection of Vegetable Seed by Treatment with Essential Oils, Organic Acids and Plant Extract. Seed Sci. Technol. 2008, 36, 76–88. [Google Scholar] [CrossRef]
- Maude, R.B. Seedborne Diseases and Their Control: Principles and Practice; CAB International: Wallingford, UK, 1996; pp. 1–280. [Google Scholar]
- Chun, S.-C.; Schneider, R.W.; Cohn, M.A. Sodium Hypochlorite: Effect of Solution pH on Rice Seed Disinfestation and Its Direct Effect on Seedling Growth. Plant Dis. 1997, 81, 821–824. [Google Scholar] [CrossRef]
- Fatmi, M.; Schaad, N.W.; Bolkan, H.A. Seed Treatments for Eradicating Clavibacter Michiganensis Subsp. Michiganensis from Naturally Infected Tomato Seeds. Plant Dis. 1991, 75, 383–385. [Google Scholar] [CrossRef]
- Black & Veatch Corporation. White’s Handbook of Chlorination and Alternative Disinfectants, 5th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 1–1062. [Google Scholar]
- Ritchie, D.F.; Averre, C.W. Bacterial Spot of Pepper and Tomato. Phytopathology 1996, 86, 952–958. [Google Scholar] [CrossRef]
- Khah, E.M.; Passam, H.C. Sodium Hypochlorite Concentration, Temperature, and Seed Age Influence Germination of Sweet Pepper. Hortscience 1992, 27, 821–823. [Google Scholar] [CrossRef]
- Rutala, W.A.; Weber, D.J. Guideline for Disinfection and Sterilization in Healthcare Facilities, 2008. Inf. Contr. Hosp. Epidemiol. 2008, 18, 240–264. [Google Scholar]
- Le Buanec, B. Organic Seed; APSA Technological Report No. 39; APSA: Seoul, Republic of Korea, 2004. [Google Scholar]
- Borgen, A.; Nielsen, B. Effect of seed treatment with acetic acid in control of seed borne diseases. In Proceedings of the BCPC Symposium No. 76: “Seed Treatment: Challenges & Opportunities”, Farnham, 76, British Crop Protection Council, Wishaw, UK, 26–27 February 2001; Biddle, A.J., Ed.; Available online: http://orgprints.org/1116/1/acidBCPC.htm (accessed on 21 December 2023).
- Kuhar, T.P.; Rideout, S.L.; Reiter, M.S. Southeastern U.S. Vegetable Crop Handbook; Highland AG: Mulberry, FL, USA, 2021. [Google Scholar] [CrossRef]
- Scott, G.; Almasrahi, A.; Mansoorkhani, F.M.; Rupar, M.; Dickinson, M.; Shama, G. Hormetic UV-C Seed Treatments for the Control of Tomato Diseases. Plant Pathol. 2019, 68, 700–707. [Google Scholar] [CrossRef]
- Schaad, N.W. Emerging Plant Pathogenic Bacteria and Global Warming. In Pseudomonas syringae Pathovars and Related Pathogens–Identification, Epidemiology and Genomics; Fatmi, M., Collmer, A., Iacobellis, N.S., Mansfield, J.W., Murillo, J., Schaad, N.W., Ullrich, M., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 369–379. [Google Scholar] [CrossRef]
- McGee, D.C. Seed Pathology: Its Place in Modern Seed Production. Plant Dis. 1981, 65, 638–642. [Google Scholar] [CrossRef]
- Goode, M.J.; Sasser, M. Prevention--the Key to Controlling Bacterial Spot and Bacterial Speck of Tomato. Plant Dis. 1980, 64, 831–834. [Google Scholar] [CrossRef]
- Nega, E.; Ulrich, R.; Werner, S.; Jahn, M. Hot Water Treatment of Vegetable Seed—An Alternative Seed Treatment Method to Control Seed Borne Pathogens in Organic Farming. J. Plant Dis. Prot. 2003, 110, 220–234. [Google Scholar]
- du Toit, L.J.; Hernandez-Perez, P. Efficacy of Hot Water and Chlorine for Eradication of Cladosporium variabile, Stemphylium botryosum, and Verticillium dahliae from Spinach Seed. Plant Dis. 2005, 89, 1305–1312. [Google Scholar] [CrossRef] [PubMed]
- McGrath, M.T. Hot Water Seed Treatment Protocols. 2013. Available online: https://cpb-us-e1.wpmucdn.com/blogs.cornell.edu/dist/1/7446/files/2021/04/Hot-Water-Seed-Treatment-Tables.pdf (accessed on 13 December 2023).
- Marinescu, G. Recherches Sur La Desinfection Des Semences de Tomates Contaminees Par Corynebacterium Michiganense (E. F. Smith). Jensen. Bull. Acad. Sci. Agric. For. 1975, 5, 101–109. [Google Scholar]
- Rui-yun, L.; Lin, Y.; Hua, C.; Wei, L.; Shu-peng, Y.; Fu-xin, G. Effect of Dry-Heat Treatment of Different Temperature on Germination of Vegetable Seed. China Veg. 2011, 1, 67–71. [Google Scholar]
- Akman, Z. Comparison of High Temperature Tolerance in Maize, Rice and Sorghum Seeds by Plant Growth Regulators. J. Anim. Vet. Adv. 2009, 8, 358–361. [Google Scholar]
- Pérez-Calvo, M. Chapter 9—Sanitation with Ozone. In Gases Agro-Food Process, 1st ed.; Cachon, R., Girardon, P., Voilley, A., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Mohan, N.; Patel, K.; Padmanabhan, K.; Ananthi, S. Ozone for Plant Pathological Applications. Ozone Sci. Eng. 2005, 27, 499–502. [Google Scholar] [CrossRef]
- Bocci, V.; Borrelli, E.; Travagli, V.; Zanardi, I. The Ozone Paradox: Ozone Is a Strong Oxidant as Well as a Medical Drug. Med. Res. Rev. 2009, 29, 646–682. [Google Scholar] [CrossRef]
- Çetinkaya, N.; Pazarlar, S.; Paylan, İ.C. Ozone Treatment Inactivates Common Bacteria and Fungi Associated with Selected Crop Seeds and Ornamental Bulbs. Saudi J. Biol. Sci. 2022, 29, 103480. [Google Scholar] [CrossRef]
- Czarnek, K.; Tatarczak-Michalewska, M.; Dreher, P.; Rajput, V.D.; Wójcik, G.; Gierut-Kot, A.; Szopa, A.; Blicharska, E. UV-C Seed Surface Sterilization and Fe, Zn, Mg, Cr Biofortification of Wheat Sprouts as an Effective Strategy of Bioelement Supplementation. Int. J. Mol. Sci. 2023, 24, 10367. [Google Scholar] [CrossRef]
- Falconí, C.E.; Yánez-Mendizábal, V. Efficacy of UV-C Radiation to Reduce Seedborne Anthracnose (Colletotrichum acutatum) from Andean Lupin (Lupinus mutabilis). Plant Pathol. 2018, 67, 831–838. [Google Scholar] [CrossRef]
- Brown, J.E.; Lu, T.Y.; Stevens, C.; Khan, V.A.; Lu, J.Y.; Wilson, C.L.; Collins, D.J.; Wilson, M.A.; Igwegbe, E.C.K.; Chalutz, E.; et al. The Effect of Low Dose Ultraviolet Light-C Seed Treatment on Induced Resistance in Cabbage to Black Rot (Xanthomonas campestris pv. campestris). Crop Prot. 2001, 20, 873–883. [Google Scholar] [CrossRef]
- Shude, S.P.N.; Yobo, K.S.; Mbili, N.C. Progress in the Management of Fusarium Head Blight of Wheat: An Overview. S. Afr. J. Sci. 2020, 116, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Maude, R.B.; Kyle, A.M. Seed Treatments with Benomyl and Other Fungicides for the Control of Ascochyta pisi on Peas. Ann. Appl. Biol. 1970, 66, 37–41. [Google Scholar] [CrossRef]
- European Commission. Implementing Regulation (EU) 2018/1500 of 9 October 2018 Concerning the Non-Renewal of Approval of the Active Substance Thiram, and Prohibiting the Use and Sale of Seeds Treated with Plant Protection Products Containing Thiram, in Accordance with Regulation (EC) No 1107/2009 of the European Parliament and of the Council Concerning the Placing of Plant Protection Products on the Market, and Amending Commission Implementing Regulation (EU) No. 540/2011; OJ L 254, 10.10.2018; European Commission: Brussels, Belgium, 2018; pp. 1–3. [Google Scholar]
- Sain, S.K.; Pandey, A.K. Biological Spectrum of Trichoderma harzianum Rifai Isolates to Control Fungal Diseases of Tomato (Solanum lycopersicon L.). Arch. Phytopathol. Plant Prot. 2016, 49, 507–521. [Google Scholar] [CrossRef]
- Chandel, S.; Allan, E.J.; Woodward, S. Biological Control of Fusarium oxysporum f. sp. lycopersici on Tomato by Brevibacillus brevis. J. Phytopathol. 2010, 158, 470–478. [Google Scholar] [CrossRef]
- Jambhulkar, P.P.; Sharma, P. Promotion of Rice Seedling Growth Characteristics by Development and Use of Bioformulation of Pseudomonas fluorescens. Indian J. Agric. Sci. 2013, 83, 136–142. [Google Scholar]
- Paravar, A.; Piri, R.; Balouchi, H.; Ma, Y. Microbial Seed Coating: An Attractive Tool for Sustainable Agriculture. Biotechnol. Rep. 2023, 37, e00781. [Google Scholar] [CrossRef]
- Bejarano, A.; Puopolo, G. Bioformulation of Microbial Biocontrol Agents for a Sustainable Agriculture. In Progress in Biological Control; Springer: Cham, Switzerland, 2020; pp. 275–293. [Google Scholar] [CrossRef]
- Jambhulkar, P.P.; Sharma, P.; Yadav, R. Delivery Systems for Introduction of Microbial Inoculants in the Field. In Microbial Inoculants in Sustainable Agricultural Productivity; Springer: New Delhi, India, 2016; pp. 199–218. [Google Scholar] [CrossRef]
- Mandiriza, G.; Kritzinger, Q.; Aveling, T.A.S. The Evaluation of Plant Extracts, Biocontrol Agents and Hot Water as Seed Treatments To Control Black Rot Of Rape In South Africa. Crop Prot. 2018, 114, 129–136. [Google Scholar] [CrossRef]
- Grimaldi, A.; Galy, R. An Overview of European Regulatory of Biopesticides. 4th International Symposium on Biological Control of Bacterial Plant Diseases. J. Plant Pathol. 2019, 101, 849–883. [Google Scholar] [CrossRef]
- Rocha, I.; Ma, Y.; Souza-Alonso, P.; Vosátka, M.; Freitas, H.; Oliveira, R.S. Seed Coating: A Tool for Delivering Beneficial Microbes to Agricultural Crops. Frontiers in Plant Sci. 2019, 10, 1357. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Farm to Fork Strategy; For a Fair, healthy and Environmentally-Friendly Food System; 23 Pages; European Commission: Brussels, Belgium, 2020; Available online: https://food.ec.europa.eu/system/files/2020-05/f2f_action-plan_2020_strategy-info_en.pdf (accessed on 22 January 2024).



{kind=link}
{kind=link}
{kind=link}
| Nature of Seed Treatment | Principle of the Method | Substance/Antimicrobial Compound | Operating Conditions | Target Pathogen | Crop Plants | Efficacy and Additional Notes | Ref. |
|---|---|---|---|---|---|---|---|
| Physical | Hot water | / | Soaking infected seeds. | Cmm | Tomato | Complete seed disinfection. | [96] |
| Physical | Hot water | / | Soaking infected seeds. | Cmm | Tomato | Reduced disease quantity observed in the field; seed germination slightly reduced. | [97] |
| Physical | Hot water | / | Soaking infected seeds. | Pst | Tomato | No disease observed under greenhouse conditions; seed germination not affected. | [44] |
| Physical | Hot water | / | Soaking infected seeds. | Pst | Tomato | Pathogen infecting seeds reduced, as evidenced by agar plating. | [98] |
| Physical | Steam-air | / | Treating infected seeds at 55 °C for 30 min. | Pst | Tomato | Pathogen infecting seeds reduced, as evidenced by agar plating. | [98] |
| Physical | Dry heating | / | Heating at 70 °C for 4 to 6 days. | Cmm | Tomato | Complete seed disinfection, as evidenced by agar plating. | [99] |
| Physical | Ozone | Gaseous O3 | Gaseous O3 treatment. | Cmm and Pst | Tomato | Complete seed disinfection, as evidenced by agar plating. | [100] |
| Biological | Plant extracts | Plant extracts from Aloe vera, Coffea arabica, Glycyrrhiza uralensis, and Yucca schidigera | Soaking infected seeds. | Xep | Tomato | Complete seed disinfection, as evidenced by in vitro and in planta observations; germination performance increased and promition of seedlings growth. | [101] |
| Biological | Plant extracts | Hexane–methanol extracts from Satureja hortensis | Soaking infected seeds in extract dilutions on a rotary shaker. | Cmm and Xv | Tomato | Disease severity reductionunder controlled conditions; germination performance decreased | [102] |
| Biological | Plant extracts | Aqueous plant extracts from coriander (Coriandrum sativum), eucalyptus (Eucalyptus sp.), Kastamonu garlic (Allium sativum ‘Kastamonu’), ginger (Zingiber officinale), Istanbul thyme (Origanum vulgare subsp. Hirtum) and Izmir thyme (Origanum onites) | Soaking infected seeds in the extract’s dilutions on a rotary shaker. | Pst | Tomato | Reduced disease incidence and severity on seedlings in controlled conditions. | [103] |
| Biological | Microorganisms (BCAs) | Pseudomonas fluorescens | Soaking infected seeds both in (i) a bacterial suspension and in (ii) a bacterial formulation (in purified talcum powderand carboxy methyl cellulose). | Cmm | Tomato | Disease incidence reduced, as observed in the field. | [104] |
| Biological | Microorganisms (BCAs) | Azospirillum brasilense | Soaking infected seeds in a bacterial suspension. | Pst | Tomato | No disease observed on seedlings under greenhouse conditions; seed germination not affected. | [105] |
| Chemical and physical | Chemi-thermal Treatment | Cupric acetate (2.0 g L−1). Glacial acetic acid (1.0 mL L−1). Mixed solution of 23.2% pentachloronitrobenzene and 5.8% 5-methoxy 3(trichloromethyl)-l,2,4-thiadiazole (4.5 mL L−1) Triton x-100 (0.2 mL L−1) | Soaking infected seeds in the chemical solutions at increasing temperatures. | Cmm, Xv, and Pst | Tomato | Complete seed disinfection with chemi-thermal treatment, as evidenced in vitro and in planta under controlled conditions; seed germination and seedlings vigour were not affected. | [106] |
| Chemical and biological | Acidified nitrite/copper hydroxide/Bacillus spp. Strains | Acidified nitrite solution (300 mmol L−1, pH 2). Kocide 101 (copper hydroxide 50% WP) at the rate of 3 g L−1 Bacillus spp. strains | Soaking infected seeds into prepared solutions. | Cmm | Tomato | Complete seed disinfection by copper hydroxide and Bacillus spp.; pathogen infecting seeds reduced using acidified nitrite solution, as observed under controlled conditions. | [107] |
| Chemical and physical | Chemical treatment/Hot water | NaHCl | Not available. | Xanthomonads | Pepper | Reduction in bacterial populations on seed surface. | [108] |
| Physical, chemical, and biological | Hot water, Chemical, Plant extracts | Hot water, NaHCl, oxidate 2.0, and thyme oil | Soaking infected seeds in: (i)hot water; (ii) NaHCl; (iii) Oxidate 2.0; (iv) Thyme oil. | Xe | Pepper | Complete seed disinfection by hot water and NaHCl; pathogen infecting seeds reduced using Oxidate 2.0 and thyme oil, as evidenced by agar plating. | [109] |
| Physical, chemical | Hot water, Chemical | NaHCl and metalaxyl-M | Soaking infected seeds in: (i) hot water; (ii) NaHCl; (iii) Metalaxyl-M (Ridomil). | Cmm, Xv, and Pst | Tomato | Hot water and Chlorine treatment: reduction in seed contamination as evidenced by agar plating. No disinfection observed using Metalaxyl-M. | [110] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xhemali, B.; Giovanardi, D.; Biondi, E.; Stefani, E. Tomato and Pepper Seeds as Pathways for the Dissemination of Phytopathogenic Bacteria: A Constant Challenge for the Seed Industry and the Sustainability of Crop Production. Sustainability 2024, 16, 1808. https://doi.org/10.3390/su16051808
Xhemali B, Giovanardi D, Biondi E, Stefani E. Tomato and Pepper Seeds as Pathways for the Dissemination of Phytopathogenic Bacteria: A Constant Challenge for the Seed Industry and the Sustainability of Crop Production. Sustainability. 2024; 16(5):1808. https://doi.org/10.3390/su16051808
Chicago/Turabian StyleXhemali, Bekri, Davide Giovanardi, Enrico Biondi, and Emilio Stefani. 2024. "Tomato and Pepper Seeds as Pathways for the Dissemination of Phytopathogenic Bacteria: A Constant Challenge for the Seed Industry and the Sustainability of Crop Production" Sustainability 16, no. 5: 1808. https://doi.org/10.3390/su16051808
APA StyleXhemali, B., Giovanardi, D., Biondi, E., & Stefani, E. (2024). Tomato and Pepper Seeds as Pathways for the Dissemination of Phytopathogenic Bacteria: A Constant Challenge for the Seed Industry and the Sustainability of Crop Production. Sustainability, 16(5), 1808. https://doi.org/10.3390/su16051808