
Biochemical and Quality Parameter Changes of Wheat Grains during One-Year Storage under Different Storage Conditions

 ,
,  ,
, 
 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Wheat Grain Storage
2.2. Determination of Germination Percentage
2.3. Moisture and Protein Content of Grains
2.4. Extractable Protein Content, Peptidase and Oxidoreductase Activities
2.5. Advanced Glycation End Products Content
2.6. Wet Gluten Content
2.7. Falling Number
2.8. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination; Springer: Boston, MA, USA, 1985; ISBN 978-1-4613-5703-2. [Google Scholar]
- McDonald, M.B. Seed Deterioration: Physiology, Repair and Assessment. Seed Sci. Technol. 1999, 27, 177–237. [Google Scholar]
- Farrant, C.W.V.; Jill, M. Acquisition and Loss of Desiccation Tolerance. In Seed Development and Germination; Routledge: London, UK, 1995; ISBN 978-0-203-74007-1. [Google Scholar]
- Staniek, K.; Nohl, H. Are Mitochondria a Permanent Source of Reactive Oxygen Species? Biochim. Biophys. Acta-Bioenerg. 2000, 1460, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Kristal, B.; Yu, B. An Emerging Hypothesis—Synergistic Induction of Aging by Free-Radicals and Maillard Reactions. J. Gerontol. 1992, 47, B107–B114. [Google Scholar] [CrossRef] [PubMed]
- Walters, C. Understanding the Mechanisms and Kinetics of Seed Aging. Seed Sci. Res. 1998, 8, 223–244. [Google Scholar] [CrossRef]
- Ashok, B.T.; Ali, R. The Aging Paradox: Free Radical Theory of Aging. Exp. Gerontol. 1999, 34, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R. Role of Oxidant Species in Aging. Curr. Med. Chem. 2004, 11, 1105–1112. [Google Scholar] [CrossRef]
- Beckman, K.B.; Ames, B.N. The Free Radical Theory of Aging Matures. Physiol. Rev. 1998, 78, 547–581. [Google Scholar] [CrossRef]
- Levine, R.L.; Stadtman, E.R. Oxidative Modification of Proteins during Aging. Exp. Gerontol. 2001, 36, 1495–1502. [Google Scholar] [CrossRef]
- Berlett, B.S.; Stadtman, E.R. Protein Oxidation in Aging, Disease, and Oxidative Stress. J. Biol. Chem. 1997, 272, 20313–20316. [Google Scholar] [CrossRef]
- Oliver, C.; Ahn, B.; Moerman, E.; Goldstein, S.; Stadtman, E. Age-Related-Changes in Oxidized Proteins. J. Biol. Chem. 1987, 262, 5488–5491. [Google Scholar] [CrossRef]
- Viteri, G.; Carrard, G.; Birlouez-Aragón, I.; Silva, E.; Friguet, B. Age-Dependent Protein Modifications and Declining Proteasome Activity in the Human Lens. Arch. Biochem. Biophys. 2004, 427, 197–203. [Google Scholar] [CrossRef]
- Thorpe, S.R.; Baynes, J.W. Maillard Reaction Products in Tissue Proteins: New Products and New Perspectives. Amino Acids 2003, 25, 275–281. [Google Scholar] [CrossRef]
- Monnier, V. Nonenzymatic Glycosylation, the Maillard Reaction and the Aging Process. J. Gerontol. 1990, 45, B105–B111. [Google Scholar] [CrossRef]
- Dills, W. Protein Fructosylation—Fructose and the Maillard Reaction. Am. J. Clin. Nutr. 1993, 58, 779–787. [Google Scholar] [CrossRef]
- Strelec, I. Aktivnost Aminopeptidaza i Sastav Proteina Sorti Ječma; Prirodoslovno-Matematički Fakultet: Zagreb, Croatia, 2004. [Google Scholar]
- Murat Karaoğlu, M.; Aydeniz, M.; Kotancilar, H.G.; Gerçelaslan, K.E. A Comparison of the Functional Characteristics of Wheat Stored as Grain with Wheat Stored in Spike Form. Int. J. Food Sci. Technol. 2010, 45, 38–47. [Google Scholar] [CrossRef]
- Mezei, Z.; Sipos, P.; Győri, Z. Variations in Quality Parameters of Forage and Medium Quality Winter Wheat Varieties in Storage. Agric. Conspec. Sci. 2007, 72, 221–225. [Google Scholar]
- Calucci, L.; Capocchi, A.; Galleschi, L.; Ghiringhelli, S.; Pinzino, C.; Saviozzi, F.; Zandomeneghi, M. Antioxidants, Free Radicals, Storage Proteins, Puroindolines, and Proteolytic Activities in Bread Wheat (Triticum aestivum) Seeds during Accelerated Aging. J. Agric. Food Chem. 2004, 52, 4274–4281. [Google Scholar] [CrossRef] [PubMed]
- Galleschi, L.; Capocchi, A.; Ghiringhelli, S.; Saviozzi, F.; Calucci, L.; Pinzino, C.; Zandomeneghi, M. Antioxidants, Free Radicals, Storage Proteins, and Proteolytic Activities in Wheat (Triticum durum) Seeds during Accelerated Aging. J. Agric. Food Chem. 2002, 50, 5450–5457. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Biswas, S.K.; Jimenez, L.A.; Torres, M.; Forman, H.J. Glutathione, Stress Responses, and Redox Signaling in Lung Inflammation. Antioxid. Redox Signal. 2005, 7, 42–59. [Google Scholar] [CrossRef] [PubMed]
- Das, G.; Senmandi, S. Triphenyl Tetrazolium Chloride Staining Pattern of Differentially Aged Wheat Seed Embryos. Seed Sci. Technol. 1992, 20, 367–373. [Google Scholar]
- Ganguli, S.; Senmandi, S. Some Physiological Differences between Naturally and Artificially Aged Wheat Seeds. Seed Sci. Technol. 1990, 18, 507–514. [Google Scholar]
- Dell’Aquila, A. Wheat Seed Ageing and Embryo Protein Degradation. Seed Sci. Res. 1994, 4, 293–298. [Google Scholar] [CrossRef]
- Bernal-Lugo, I.; Rodriguez, M.; Gavilanes-Ruiz, M.; Hamabata, A. Reduced Aleurone α-Amylase Production in Aged Wheat Seeds Is Accompanied by Lower Levels of High-Pl α-Amylase Transcripts and Reduced Response to Gibberellic Acid. J. Exp. Bot. 1999, 50, 311–317. [Google Scholar] [CrossRef]
- Strelec, I.; Popović, R.; Ivanišić, I.; Jurković, V.; Jurković, Z.; Ugarčić-Hardi, Ž.; Sabo, M. Influence of Temperature and Relative Humidity on Grain Moisture, Germination and Vigour of Three Wheat Cultivars during One Year Storage. Poljoprivreda 2010, 16, 20–24. [Google Scholar]
- Strelec, I.; Šarkanj, B.; Mrša, V.; Ugarčić-Hardi, Ž. Chemical Composition, Quality Parameters, Exopeptidase and Oxidoreductase Activity Changes During Temporal Development of Wheat Grain Infestation by Sitophilus granarius. J. Food Biochem. 2014, 38, 175–183. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nagatsu, I.; Nagatsu, T.; Yamamoto, T.; Glenner, G.G.; Mehl, J.W. Purification of Aminopeptidase a in Human Serum and Degradation of Angiotensin II by the Purified Enzyme. Biochim. Biophys. Acta BBA-Enzymol. 1970, 198, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Strelec, I.; Vukelić, B.; Vitale, L. Aminopeptidases of Germinated and Non-Germinated Barley. Food Technol. Biotechnol. 2009, 47, 296–303. [Google Scholar]
- Waters, S.P.; Peoples, M.B.; Simpson, R.J.; Dalling, M.J. Nitrogen Redistribution during Grain Growth in Wheat (Triticum aestivum L.). Planta 1980, 148, 422–428. [Google Scholar] [CrossRef]
- Voigt, G.; Biehl, B.; Heinrichs, H.; Voigt, J. Aspartic Proteinase Levels in Seeds of Different Angiosperms. Phytochemistry 1997, 44, 389–392. [Google Scholar] [CrossRef]
- Scebba, F.; Sebastiani, L.; Vitagliano, C. Activities of Antioxidant Enzymes during Senescence of Prunus Armeniaca Leaves. Biol. Plant. 2001, 44, 41–46. [Google Scholar] [CrossRef]
- Okot-Kotber, M.; Liavoga, A.; Yong, K.-J.; Bagorogoza, K. Activation of Polyphenol Oxidase in Extracts of Bran from Several Wheat (Triticum aestivum) Cultivars Using Organic Solvents, Detergents, and Chaotropes. J. Agric. Food Chem. 2002, 50, 2410–2417. [Google Scholar] [CrossRef]
- Ugarčić-Hardi, Ž.; Strelec, I. Accumulation of Amadori and Maillard Products in wheat seeds aged under different storage conditions. Croat. Chem. Acta 2008, 81, 131–137. [Google Scholar]
- 155 Determination of Wet Gluten Quantity and Quality (Gluten Index Ac. to Perten) of Whole Wheat Meal and Wheat Flour (Triticum aestivum). Available online: https://icc.or.at/publications/icc-standards/standards-overview/155-standard-method (accessed on 19 December 2023).
- 107/1 Determination of the Falling Number According to Hagberg—As a Measure of the Degree of Alpha-Amylase Activity in Grain and Flour. Available online: https://icc.or.at/publications/icc-standards/standards-overview/107-1-standard-method (accessed on 19 December 2023).
- Murthy, U.M.N.; Kumar, P.P.; Sun, W.Q. Mechanisms of Seed Ageing under Different Storage Conditions for Vigna radiata (L.) Wilczek: Lipid Peroxidation, Sugar Hydrolysis, Maillard Reactions and Their Relationship to Glass State Transition. J. Exp. Bot. 2003, 54, 1057–1067. [Google Scholar] [CrossRef]
- Murthy, U.M.N.; Liang, Y.H.; Kumar, P.P.; Sun, W.Q. Non-Enzymatic Protein Modification by the Maillard Reaction Reduces the Activities of Scavenging Enzymes in Vigna radiata. Physiol. Plant. 2002, 115, 213–220. [Google Scholar] [CrossRef]
- Ganguli, S.; Senmandi, S. Effects of Aging on Amylase Activity and Scutellar Cell Structure during Imbibition in Wheat Seed. Ann. Bot. 1993, 71, 411–416. [Google Scholar] [CrossRef]
- Dragicevic, V.D.; Sredojevic, S.; Spasic, M.B.; Vrvic, M.M. Ageing-Induced Changes of Reduced and Oxidised Glutathione in Fragments of Maize Seedlings. J. Serb. Chem. Soc. 2003, 68, 911–917. [Google Scholar] [CrossRef]
- Basavarajappa, B.; Shetty, H.; Prakash, H. Membrane Deterioration and Other Biochemical-Changes, Associated with Accelerated Aging of Maize Seeds. Seed Sci. Technol. 1991, 19, 279–286. [Google Scholar]
- Heath, R.; Packer, L. Photoperoxidation in Isolated Chloroplasts. i. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Sun, W.; Leopold, A. The Maillard Reaction and Oxidative Stress during Aging of Soybean Seeds. Physiol. Plant. 1995, 94, 94–104. [Google Scholar] [CrossRef]
- Torres, M.; De Paula, M.; Pérez-Otaola, M.; Darder, M.; Frutos, G.; Martínez-Honduvilla, C.J. Ageing-Induced Changes in Glutathione System of Sunflower Seeds. Physiol. Plant. 1997, 101, 807–814. [Google Scholar] [CrossRef]
- Siegenthaler, P.; Douetorhant, V. Relationship between the Atp Content Measured at 3 Imbibition Times and Germination of Onion Seeds during Storage at 3, 15 and 30 °C. J. Exp. Bot. 1994, 45, 1365–1371. [Google Scholar] [CrossRef]
- Bailly, C.; Benamar, A.; Corbineau, F.; Come, D. Changes in Malondialdehyde Content and in Superoxide Dismutase, Catalase and Glutathione Reductase Activities in Sunflower Seeds as Related to Deterioration during Accelerated Aging. Physiol. Plant. 1996, 97, 104–110. [Google Scholar] [CrossRef]
- Zhang, M.; Yoshiyama, M.; Nagashima, T.; Nakagawa, Y.; Yoshioka, T.; Esashi, Y. Aging of Soybean Seeds in Relation to Metabolism at Different Relative Humidities. Plant Cell Physiol. 1995, 36, 1189–1195. [Google Scholar]
- Begnami, C.N.; Cortelazzo, A.L. Cellular Alterations during Accelerated Aging of French Bean Seeds. Seed Sci. Technol. 1996, 24, 295–303. [Google Scholar]
- Sammour, R.H. Effect of Ageing on the Major Reserve Molecules and Their Related Enzyme in Natural Aged Seeds of Flax. J. Islam. Acad. Sci. 1989, 2, 247–251. [Google Scholar]
- Petruzzelli, L.; Taranto, G. Wheat Aging—The Contribution of Embryonic and Non-Embryonic Lesions to Loss of Seed Viability. Physiol. Plant. 1989, 76, 289–294. [Google Scholar]
- Ravikumar, R.; Ananthakrishnan, G.; Ganapathi, A.; Appasamy, T. Biochemical Changes Induced by Accelerated Ageing in Bambusa Bambos Seeds. Biol. Plant. 1998, 40, 459–464. [Google Scholar] [CrossRef]
- Ravikumar, R.; Ananthakrishnan, G.; Girija, S.; Ganapathi, A. Seed Viability and Biochemical Changes Associated with Accelerated Ageing in Dendrocalamus Strictus Seeds. Biol. Plant 2002, 45, 153–156. [Google Scholar] [CrossRef]
- Dhaliwal, Y.; Sekhon, K.; Nagi, H. Enzymatic-Activities and Rheological Properties of Stored Rice. Cereal Chem. 1991, 68, 18–21. [Google Scholar]
- Zhou, Z.; Robards, K.; Helliwell, S.; Blanchard, C. Ageing of Stored Rice: Changes in Chemical and Physical Attributes. J. Cereal Sci. 2002, 35, 65–78. [Google Scholar] [CrossRef]
- Strelec, I.; Vitale, L. Changes of aminopeptidase activities in barley grain during storage. In Proceedings of the 3rd International Congress ‘Flour-Bread 05’ and 5th Croatian Congress of Cereal Technologists, Opatija, Croatia, 26–29 October 2005; CABI: Wallingford, UK, 2006; p. 63. [Google Scholar]
- Matsukura, U.; Kaneko, S.; Momma, M. Method for measuring the freshness of individual rice grains by means of a color reaction of catalase activity. J. Jpn. Soc. Food Sci. Technol.-Nippon Shokuhin Kagaku Kogaku Kaishi 2000, 47, 523–528. [Google Scholar] [CrossRef]
- Chen, T.F.; Chen, C.L. Analysing the Freshness of Intact Rice Grains by Colour Determination of Peroxidase Activity. J. Sci. Food Agric. 2003, 83, 1214–1218. [Google Scholar] [CrossRef]
- Barrett, A.J.; Rawlings, N.D.; Woessner, J.F. Handbook of Proteolytic Enzymes; Academic Press: Cambridge, MA, USA, 2012; ISBN 978-0-12-382220-8. [Google Scholar]
- Kruger, J.E.; Lineback, D.R. Enzymes and Their Role in Cereal Technology; Stauffer, C.E., Ed.; Amer Assn of Cereal Chemists: St. Paul, MN, USA, 1987; ISBN 978-0-913250-46-4. [Google Scholar]
- Every, D.; Simmons, L.D.; Ross, A.P. Distribution of Redox Enzymes in Millstreams and Relationships to Chemical and Baking Properties of Flour. Cereal Chem. 2006, 83, 315. [Google Scholar] [CrossRef]
- Garcia, R.; Kaid, N.; Vignaud, C.; Nicolas, J. Purification and Some Properties of Catalase from Wheat Germ (Triticum aestivum L.). J. Agric. Food Chem. 2000, 48, 1050–1057. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M. Polyphenol Oxidases in Plants and Fungi: Going Places? A Review. Phytochemistry 2006, 67, 2318–2331. [Google Scholar] [CrossRef]
- Baik, B.-K.; Donelson, T. Postharvest and Postmilling Changes in Wheat Grain and Flour Quality Characteristics. Cereal Chem. 2018, 95, 141–148. [Google Scholar] [CrossRef]
- Hrušková, M.; Machová, D. Changes of Wheat Flour Properties during Short Term Storage. Czech J. Food Sci. 2002, 20, 125–130. [Google Scholar] [CrossRef]
- Wang, L.F.; Flores, R.A. The Effects of Storage on Flour Quality and Baking Performance. Food Rev. Int. 1999, 15, 215–234. [Google Scholar] [CrossRef]
- Hrušková, M.; Škodová, V.; Blažek, J. Wheat Sedimentation Values and Falling Number. Czech J. Food Sci. 2004, 22, 51–57. [Google Scholar] [CrossRef]
- Kibar, H. Influence of Storage Conditions on the Quality Properties of Wheat Varieties. J. Stored Prod. Res. 2015, 62, 8–15. [Google Scholar] [CrossRef]
- Ibanoglu, S. Wheat Washing with Ozonated Water: Effects on Selected Flour Properties. Int. J. Food Sci. Technol. 2002, 37, 579–584. [Google Scholar] [CrossRef]
- Curic, D.; Dugum, J.; Bauman, I. The Influence of Fungal α-Amylase Supplementation on Amylolytic Activity and Baking Quality of Flour. Int. J. Food Sci. Technol. 2002, 37, 673–680. [Google Scholar] [CrossRef]
- Mailhot, W.C.; Patton, J.C. Criteria of Flour Quality. In Wheat Chemistry and Technology; American Association of Cereal Chemists Inc.: St. Paul, MN, USA, 1988; pp. 69–90. [Google Scholar]
- Shibuya, N.; Iwasaki, T. Effect of Cell-Wall Degrading Enzymes on the Cooking Properties of Milled Rice and the Texture of Cooked Rice. J. Jpn. Soc. Food Sci. Technol.-Nippon Shokuhin Kagaku Kogaku Kaishi 1984, 31, 656–660. [Google Scholar] [CrossRef]
- Teo, C.H.; Abd, A.; Cheah, P.B.; Norziah, M.H.; Seow, C.C. On the Roles of Protein and Starch in the Aging of Non-Waxy Rice Flour. Food Chem. 2000, 69, 229–236. [Google Scholar] [CrossRef]
- Wilkes, M.; Copeland, L. Storage of Wheat Grains at Elevated Temperatures Increases Solubilization of Glutenin Subunits. Cereal Chem. 2008, 85, 335–338. [Google Scholar] [CrossRef]
- Salman, H.; Copeland, L. Effect of Storage on Fat Acidity and Pasting Characteristics of Wheat Flour. Cereal Chem. 2007, 84, 600–606. [Google Scholar] [CrossRef]
- Loney, D.; Meredith, P. Note on Amylograph Viscosities of Wheat Flours and Their Starches during Storage. Cereal Chem 1974, 51, 702–705. [Google Scholar]
- Zhou, Z.K.; Robards, K.; Helliwell, S.; Blanchard, C.; Baxterb, G. Rice Ageing. I. Effect of Changes in Protein on Starch Behaviour. Starch-Starke 2003, 55, 162–169. [Google Scholar] [CrossRef]
- Weegels, P.; Verhoek, J.; Degroot, A.; Hamer, R. Effects on Gluten of Heating at Different Moisture Contents. 1. Changes in Functional-Properties. J. Cereal Sci. 1994, 19, 31–38. [Google Scholar] [CrossRef]
- Gonzalez-Torralba, J.; Arazuri, S.; Jaren, C.; Arregui, L.M. Influence of Temperature and r.h. during Storage on Wheat Bread Making Quality. J. Stored Prod. Res. 2013, 55, 134–144. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
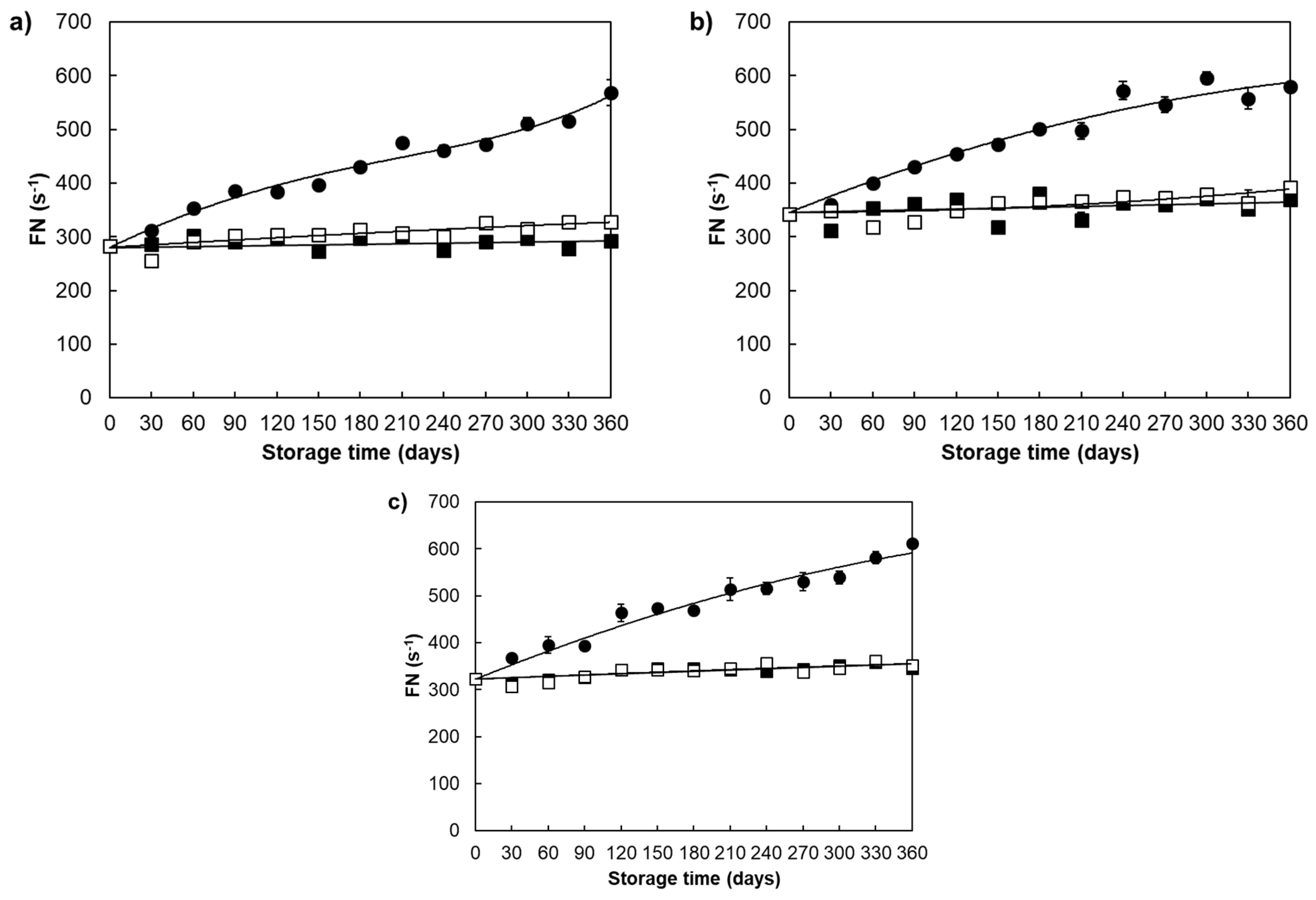{kind=link}
| Parameters Variety | n | 40 ± 1.06 °C 45 ± 3% RH | 4 ± 1.05 °C 45 ± 4% RH | 15 ± 8.51 °C 67 ± 4% RH |
|---|---|---|---|---|
| Germination (%) | ||||
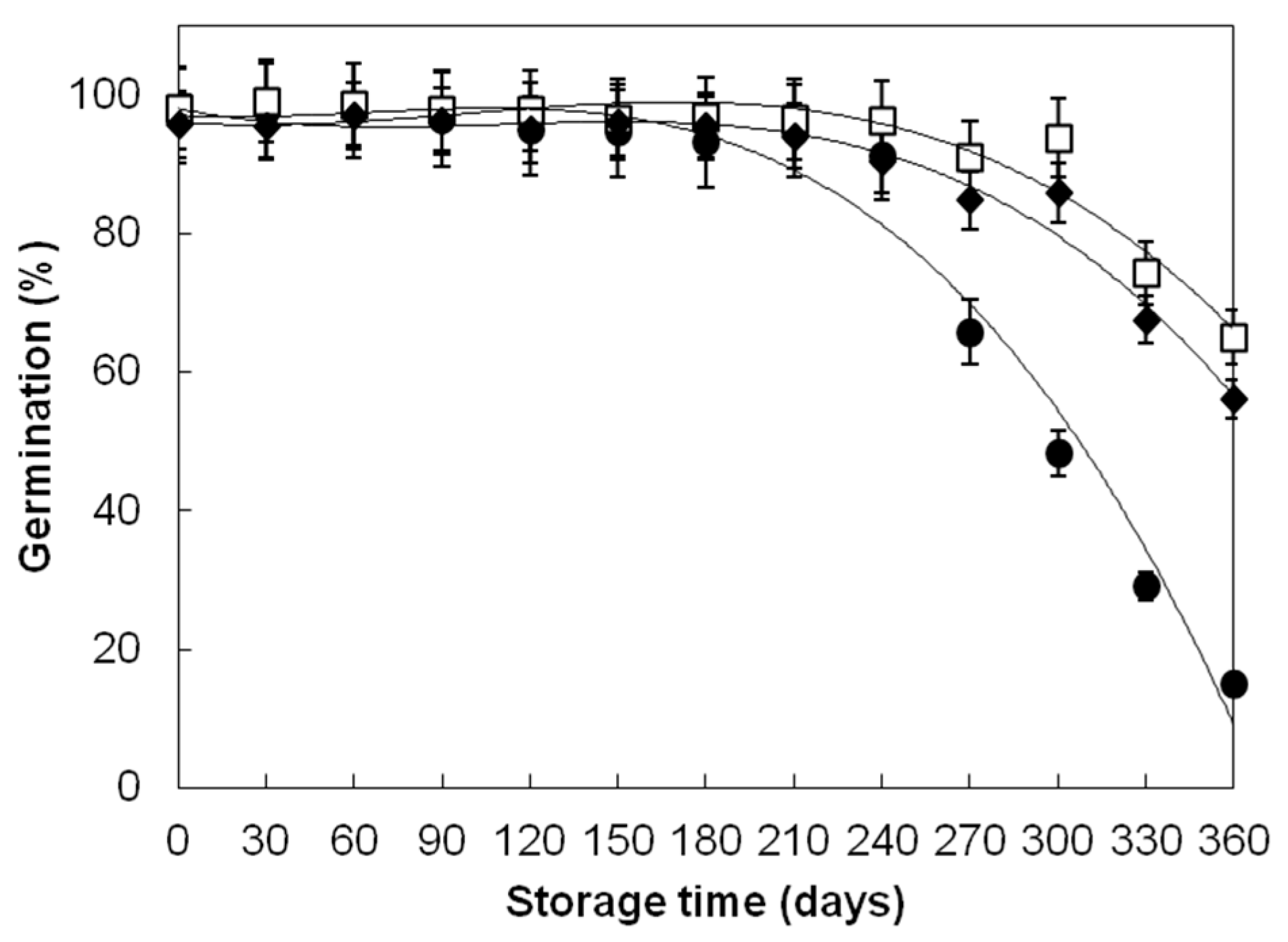
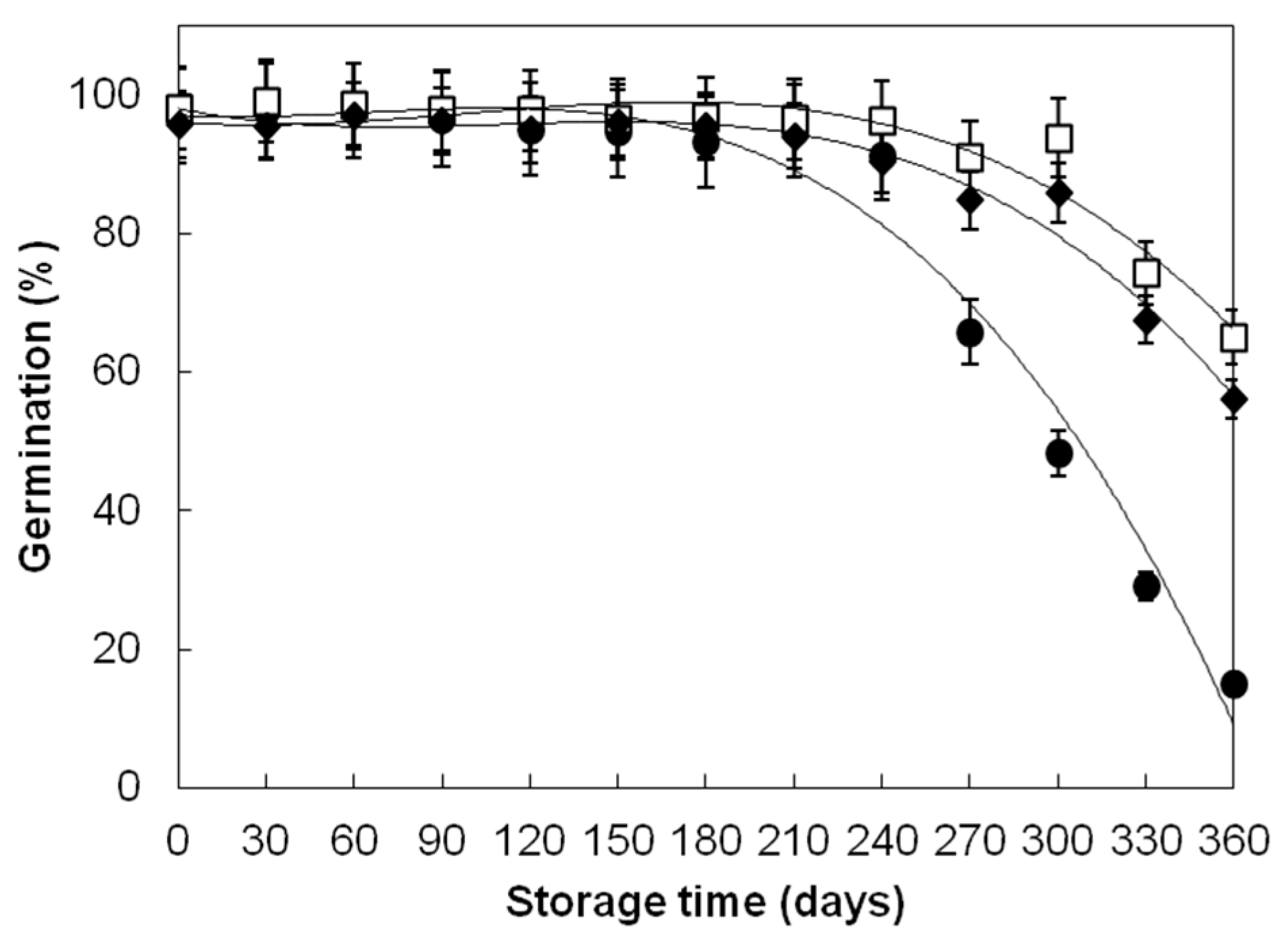
| Divana | 52 | 78.15 ± 28.31 b | 97.35 ± 1.55 a | 97.33 ± 1.42 a |
| Žitarka | 52 | 92.37 ± 10.46 b | 98.46 ± 1.21 a | 98.48 ± 1.24 a |
| Srpanjka | 52 | 88.40 ± 12.68 b | 96.77 ± 1.53 a | 96.75 ± 1.67 a |
| Dry matter content (g/g) | ||||
| Divana | 39 | 0.90 ± 0.02 a | 0.87 ± 0.01 b | 0.86 ± 0.00 c |
| Žitarka | 39 | 0.90 ± 0.02 a | 0.87 ± 0.01 b | 0.86 ± 0.00 c |
| Srpanjka | 39 | 0.90 ± 0.02 a | 0.87 ± 0.00 b | 0.86 ± 0.00 c |
| Proteins A (mg/ggrain dry weight) | ||||
| Divana | 78 | 10.83 ± 0.69 | 11.21 ± 0.77 | 11.29 ± 0.86 |
| Žitarka | 78 | 9.11 ± 0.56 | 9.47 ± 0.50 | 9.55 ± 0.61 |
| Srpanjka | 78 | 7.97 ± 0.77 | 8.35 ± 0.66 | 8.39 ± 0.71 |
| Proteins B (mg/ggrain dry weight) | ||||
| Divana | 78 | 14.70 ± 1.12 | 15.19 ± 1.61 | 15.34 ± 1.36 |
| Žitarka | 78 | 12.63 ± 0.89 | 12.89 ± 0.90 | 13.13 ± 0.96 |
| Srpanjka | 78 | 11.75 ± 0.71 | 12.18 ± 0.78 | 12.30 ± 0.78 |
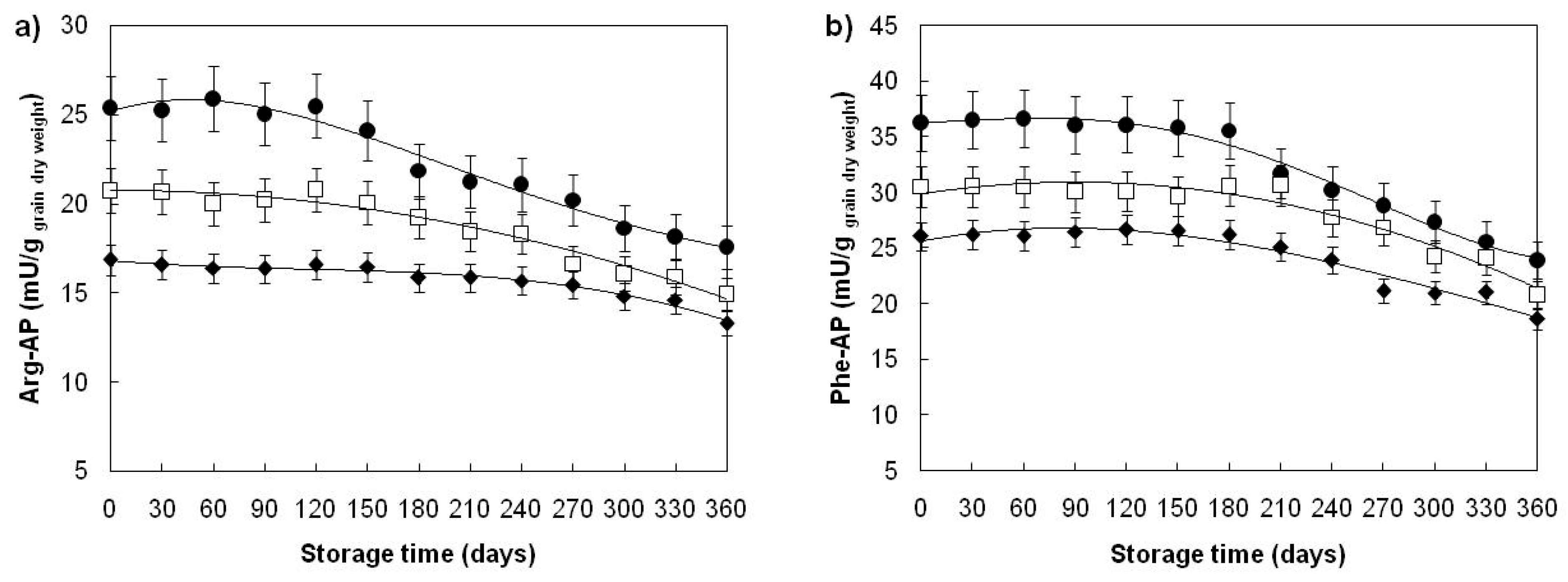
| Arginyl-aminopeptidase (mU/ggrain dry weight) | ||||
| Divana | 78 | 22.39 ± 3.20 b | 25.60 ± 1.43 a | 25.39 ± 1.36 a |
| Žitarka | 78 | 18.69 ± 2.24 b | 20.55 ± 1.36 a | 20.60 ± 1.23 a |
| Srpanjka | 78 | 15.80 ± 1.51 b | 16.64 ± 1.16 a | 16.45 ± 0.91 a |
| Phenylalanyl-aminopeptidase (mU/ggrain dry weight) | ||||
| Divana | 78 | 32.47 ±4.97 b | 36.52 ±1.72 a | 36.50 ±1.68 a |
| Žitarka | 78 | 28.26 ±3.53 b | 30.67 ±2.07 a | 30.82 ±1.89 a |
| Srpanjka | 78 | 24.30 ±3.03 b | 26.42 ±1.57 a | 26.34 ±1.76 a |
| Carboxypeptidase (mU/ggrain dry weight) | ||||
| Divana | 78 | 20.67 ± 1.87 | 20.43 ± 2.41 | 20.54 ± 2.37 |
| Žitarka | 78 | 25.41 ± 3.17 | 25.38 ± 3.12 | 25.23 ± 2.38 |
| Srpanjka | 78 | 30.62 ± 2.99 | 30.57 ± 2.38 | 30.59 ± 1.63 |
| Aspartic proteinase (U/ggrain dry weight) | ||||
| Divana | 78 | 0.40 ±0.03 | 0.41 ±0.03 | 0.41 ±0.03 |
| Žitarka | 78 | 0.45 ±0.03 | 0.46 ±0.03 | 0.46 ±0.03 |
| Srpanjka | 78 | 0.40 ±0.03 | 0.41 ±0.03 | 0.41 ±0.03 |
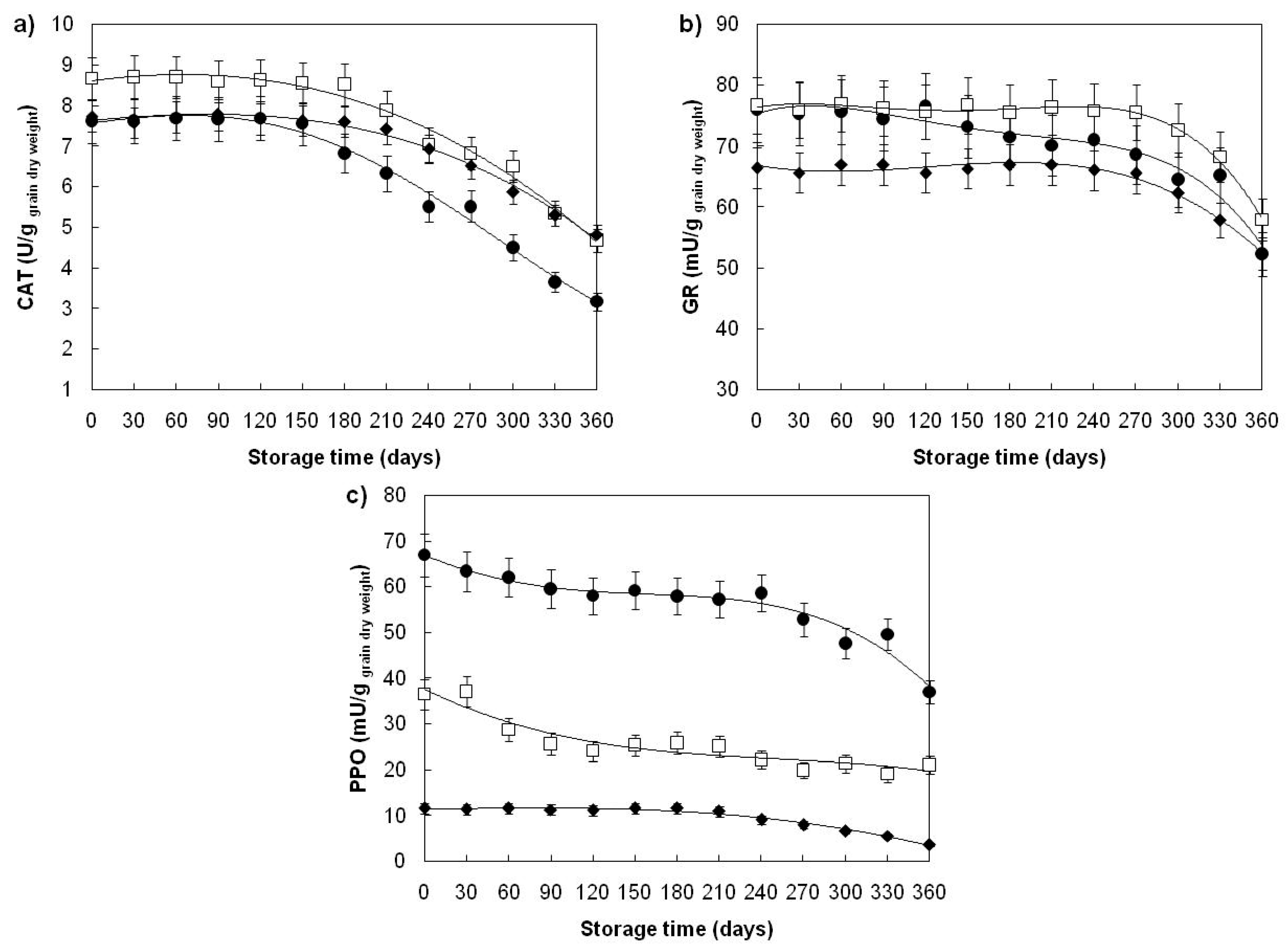
| Catalase (U/ggrain dry weight) | ||||
| Divana | 78 | 6.30 ± 1.61 b | 7.62 ± 0.32 a | 7.63 ± 0.30 a |
| Žitarka | 78 | 7.62 ± 1.38 b | 8.65 ± 0.29 a | 8.63 ± 0.34 a |
| Srpanjka | 78 | 6.99 ± 1.02 b | 7.74 ± 0.32 a | 7.70 ± 0.32 a |
| Glutathione reductase (mU/ggrain dry weight) | ||||
| Divana | 78 | 70.47 ± 7.13 b | 75.33 ± 2.93 a | 75.59 ± 2.93 a |
| Žitarka | 78 | 73.90 ± 5.93 b | 76.31 ± 2.99 a | 76.37 ± 3.33 a |
| Srpanjka | 78 | 64.32 ± 5.16 b | 66.61 ± 3.06 a | 66.54 ± 3.43 a |
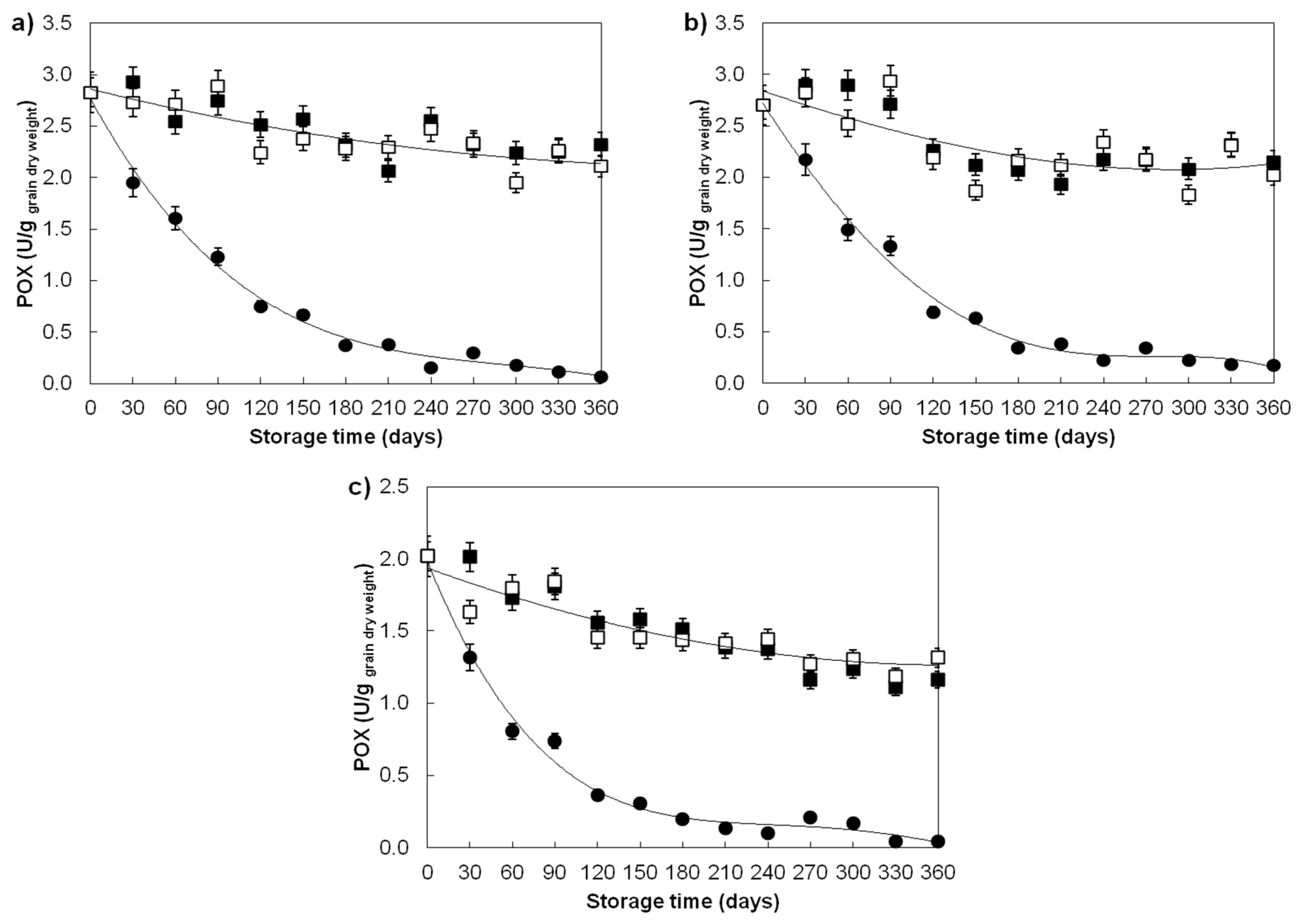
| Guaiacol peroxidase (U/ggrain dry weight) | ||||
| Divana | 78 | 0.82 ± 0.83 b | 2.48 ± 0.28 a | 2.42 ± 0.31 a |
| Žitarka | 78 | 0.84 ± 0.81 b | 2.35 ± 0.36 a | 2.31 ± 0.36 a |
| Srpanjka | 78 | 0.50 ± 0.57 b | 1.51 ± 0.32 a | 1.50 ± 0.26 a |
| Polyphenol oxidase (mU/ggrain dry weight) | ||||
| Divana | 78 | 56.14 ± 8.10 b | 67.22 ± 2.93 a | 67.21 ± 2.93 a |
| Žitarka | 78 | 25.52 ± 6.05 b | 36.61 ± 3.21 a | 36.95 ± 3.05 a |
| Srpanjka | 78 | 9.50 ± 2.77 b | 11.44 ± 0.76 a | 11.40 ± 0.89 a |
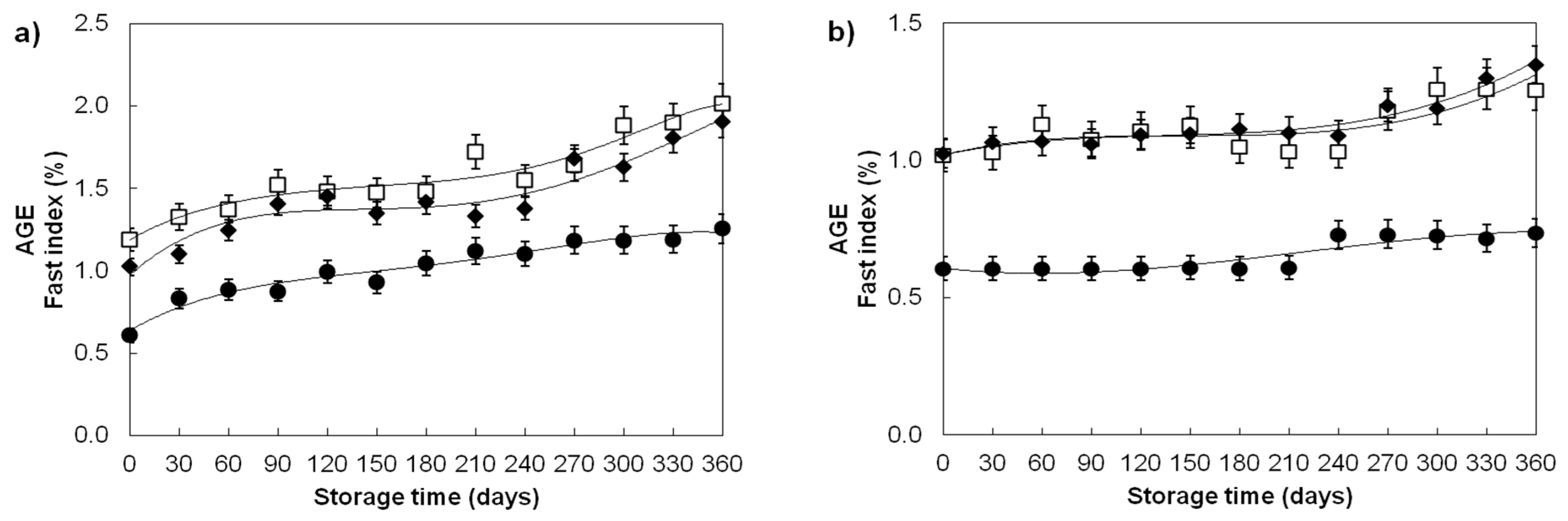
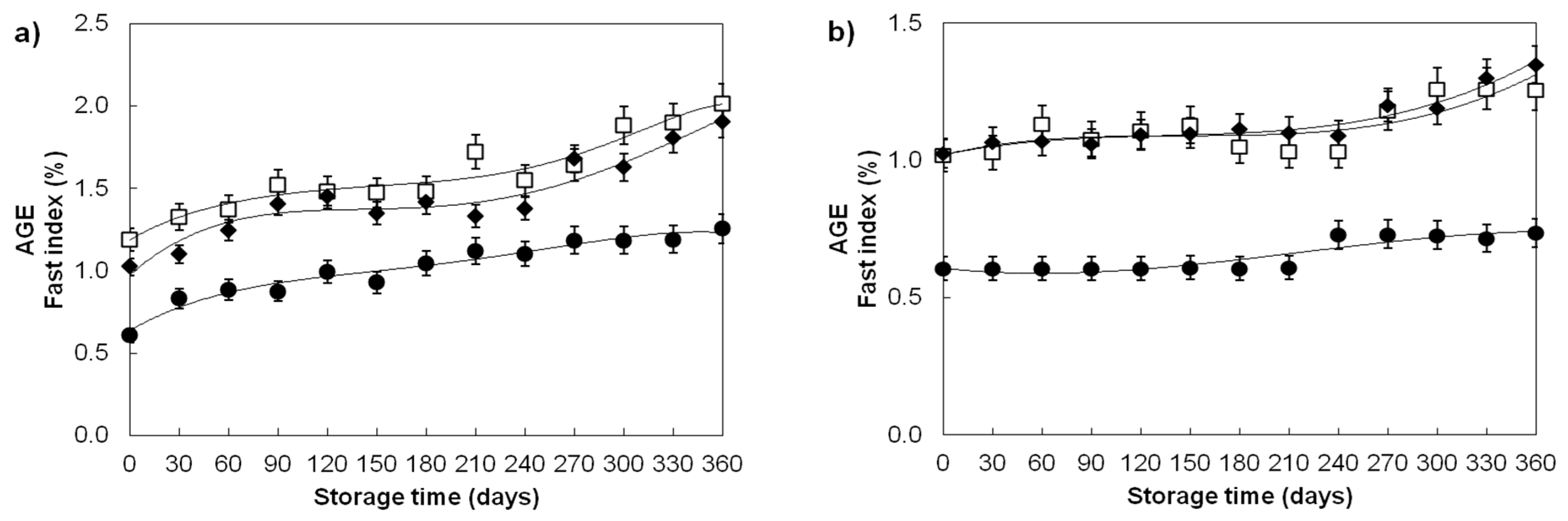
| Advanced glycation end products/Fast index (%) | ||||
| Divana | 78 | 1.02 ± 0.17 a | 0.59 ± 0.03 c | 0.65 ± 0.06 b |
| Žitarka | 78 | 1.56 ± 0.25 a | 1.06 ± 0.04 c | 1.13 ± 0.10 b |
| Srpanjka | 78 | 1.42 ± 0.27 a | 1.08 ± 0.06 c | 1.14 ± 0.12 b |
| Wet gluten content (%) | ||||
| Divana | 78 | 34.55 ± 1.28 | 35.10 ± 0.55 | 34.96 ± 0.49 |
| Žitarka | 78 | 31.18 ± 1.41 | 32.33 ± 0.38 | 32.43 ± 0.45 |
| Srpanjka | 78 | 20.93 ± 2.07 b | 23.17 ± 0.49 a | 23.83 ± 0.69 a |
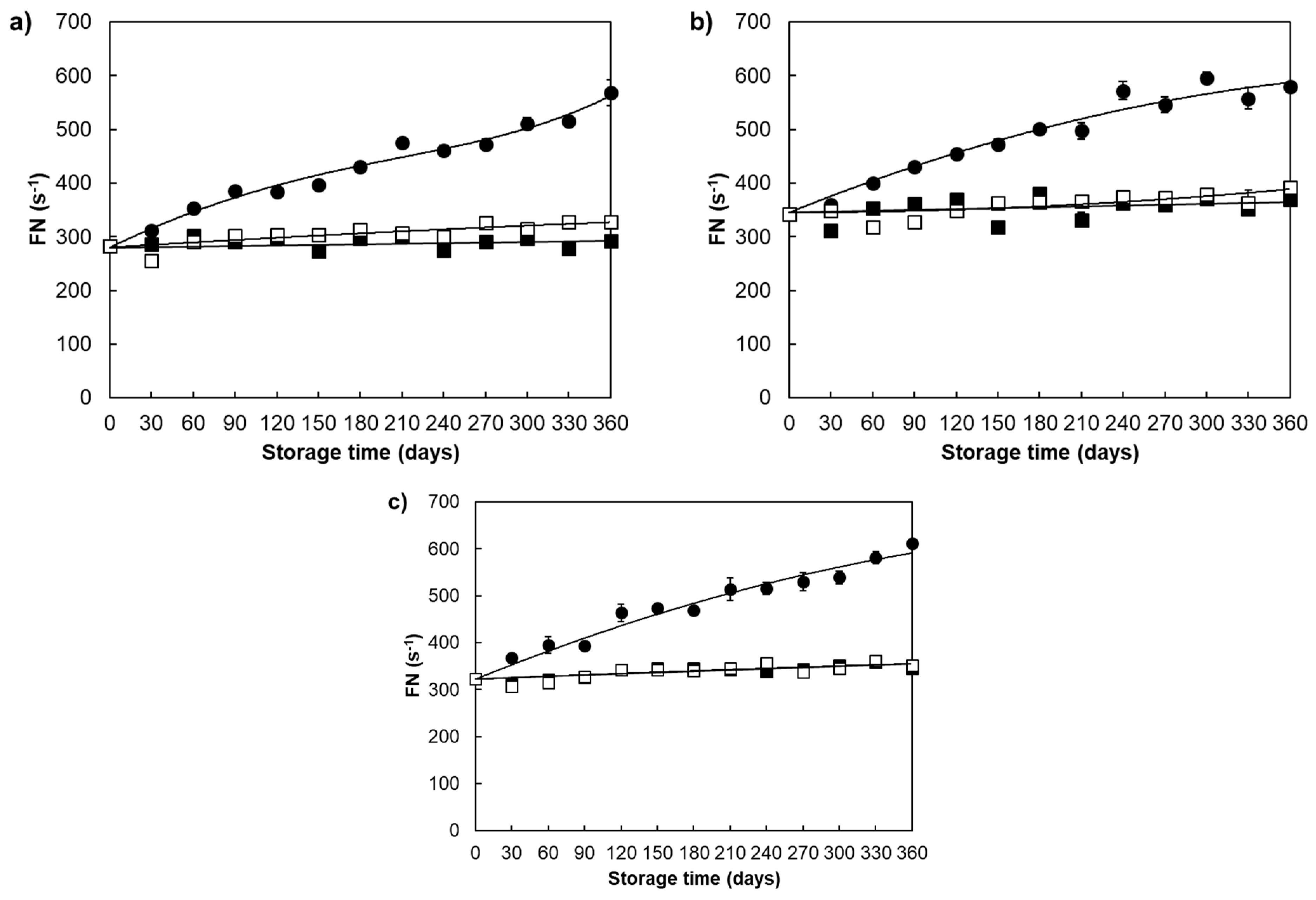
| Falling number (s) | ||||
| Divana | 78 | 425.84 ± 80.23 a | 289.29 ± 11.44 b | 304.02 ± 20.27 b |
| Žitarka | 78 | 485.12 ± 82.67 a | 351.92 ± 21.42 b | 358.48 ± 21.80 b |
| Srpanjka | 78 | 475.67 ± 83.79 a | 335.66 ± 15.80 b | 336.29 ± 16.89 b |
| Parameters | Advanced Glycation End Products | Germination | ||||
|---|---|---|---|---|---|---|
| Divana | Žitarka | Srpanjka | Divana | Žitarka | Srpanjka | |
| Phe-AP | −0.73 | −0.73 | −0.69 | 0.79 | 0.69 | 0.55 |
| Arg-AP | −0.81 | −0.84 | −0.51 | 0.83 | 0.75 | 0.77 |
| POX | −0.91 | −0.71 | −0.83 | 0.55 | 0.44 | 0.44 |
| PPO | −0.78 | −0.68 | −0.81 | 0.86 | 0.46 | 0.88 |
| CAT | −0.78 | −0.83 | −0.8 | 0.91 | 0.84 | 0.90 |
| GR | −0.68 | −0.66 | −0.56 | 0.83 | 0.79 | 0.76 |
| FN | 0.92 | 0.81 | 0.83 | −0.78 | −0.54 | −0.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strelec, I.; Mrša, V.; Simović, D.Š.; Petrović, J.; Zahorec, J.; Budžaki, S. Biochemical and Quality Parameter Changes of Wheat Grains during One-Year Storage under Different Storage Conditions. Sustainability 2024, 16, 1155. https://doi.org/10.3390/su16031155
Strelec I, Mrša V, Simović DŠ, Petrović J, Zahorec J, Budžaki S. Biochemical and Quality Parameter Changes of Wheat Grains during One-Year Storage under Different Storage Conditions. Sustainability. 2024; 16(3):1155. https://doi.org/10.3390/su16031155
Chicago/Turabian StyleStrelec, Ivica, Vladimir Mrša, Dragana Šoronja Simović, Jovana Petrović, Jana Zahorec, and Sandra Budžaki. 2024. "Biochemical and Quality Parameter Changes of Wheat Grains during One-Year Storage under Different Storage Conditions" Sustainability 16, no. 3: 1155. https://doi.org/10.3390/su16031155
APA StyleStrelec, I., Mrša, V., Simović, D. Š., Petrović, J., Zahorec, J., & Budžaki, S. (2024). Biochemical and Quality Parameter Changes of Wheat Grains during One-Year Storage under Different Storage Conditions. Sustainability, 16(3), 1155. https://doi.org/10.3390/su16031155