
A Field Evaluation of Sodium Silicate and Bacillus subtilis on the Growth and Yield of Bananas following Fusarium Wilt Disease Infection

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation and Treatment of Experminetal Materials
2.2. Data Collection
2.2.1. Determination of Crop Growth and Physiological Attributes
2.2.2. Measurement of Leaf Gas Exchange
2.2.3. Biochemical Assay and Nutrient Uptake
2.2.4. Disease Assessment
- Without disease symptoms on the plant canopy = 0;
- Yellowing of lower leaves and necrotic symptom less than 25% = 1;
- Less than 50% of the total number of leaves have a yellowish color or necrosis present = 2;
- A total of 50 to 75% of the total number of leaves have a yellowish color or necrosis present = 3;
- All leaves have a yellowish color or necrosis present (severe wilting), or plant death = 4.
2.2.5. Yield
2.3. Statistical Analysis
3. Results
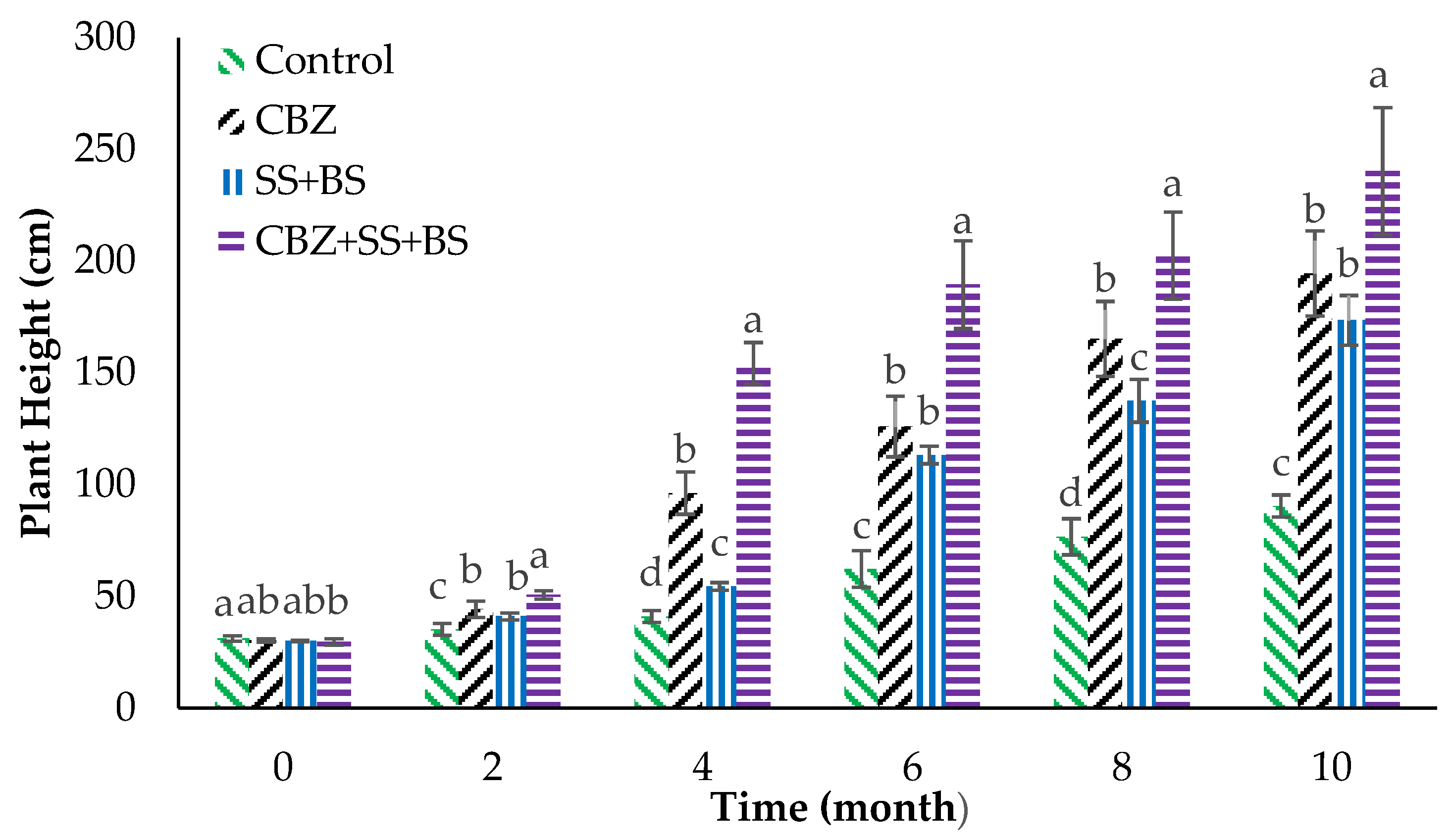
3.1. Crop Growth and Physiological Attributes
3.2. Leaf Gas Exchange
3.3. Biochemical Content and Nutrient Uptake
3.4. Disease Assessment
3.5. Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tiri, R.N.E.; Gulbagca, F.; Aygun, A.; Cherif, A.; Sen, F. Biosynthesis of Ag–Pt bimetallic nanoparticles using propolis extract: Antibacterial effects and catalytic activity on NaBH4 hydrolysis. Environ. Res. 2022, 206, 112622. [Google Scholar] [CrossRef] [PubMed]
- Thangavelu, R.; Saraswathi, M.S.; Uma, S.; Loganathan, M.; Backiyarani, S.; Durai, P.; Swennen, R. Identification of sources resistant to a virulent Fusarium wilt strain (VCG 0124) infecting Cavendish bananas. Sci. Rep. 2021, 11, 1–14. [Google Scholar]
- Blomme, G.; Ocimati, W.; Zum Felde, A.; Amwonya, D.; Kantungeko, D. A literature review on yield gaps of various root, tuber and banana crops as a background for assessing banana yield reductions due to pests and diseases at a field site in western Burundi. Afr. J. Agric. Res. 2020, 16, 1169–1183. [Google Scholar]
- Proboningrum, A.; Widono, S. Effectivity and compatibility of Azotobacter and Bacillus for biological control agents of Fusarium wilt on banana seedlings. IOP Conf. Ser. Earth Environ. Sci. 2019, 250, 012003. [Google Scholar] [CrossRef]
- Dadrasnia, A.; Usman, M.M.; Omar, R.; Ismail, S.; Abdullah, R. Potential use of Bacillus genus to control of bananas diseases: Approaches toward high yield production and sustainable management. J. King Saud Univ. Sci. 2020, 32, 2336–2342. [Google Scholar] [CrossRef]
- Wong, C.K.F.; Zulperi, D.; Vadamalai, G.; Saidi, N.B.; Teh, C.Y. Phylogenetic analysis of Fusarium oxysporum f. sp. cubense associated with Fusarium wilt of bananas from peninsular Malaysia. Sains Malays. 2019, 48, 1593–1600. [Google Scholar] [CrossRef]
- Maryani, N.; Lombard, L.; Poerba, Y.S.; Subandiyah, S.; Crous, P.W.; Kema, G.H.J. Phylogeny and genetic diversity of the banana Fusarium wilt pathogen Fusarium oxysporum f. sp. cubense in the Indonesian centre of origin. Stud. Mycol. 2019, 92, 155–194. [Google Scholar] [CrossRef]
- Dong, H.; Fan, H.; Lei, Z.; Wu, C.; Zhou, D.; Li, H. Histological and gene expression analyses in banana reveals the pathogenic differences between races 1 and 4 of banana Fusarium wilt pathogen. Phytopathology 2019, 109, 1029–1042. [Google Scholar] [CrossRef]
- Clement, W.K.F.; Vadamalai, G.; Saidi, N.B.; Zulperi, D. Research progress, challenges, future perspectives on the management of Fusarium wilt of banana in malaysia: A review. Malays. J. Sci. 2019, 382, 47–66. [Google Scholar] [CrossRef]
- Desai, S.; Dubey, S.C.; Prasad, R.D. Impacts of climate change on Fusarium species vis-à-vis adaptation strategies. Indian Phytopathol. 2020, 73, 593–603. [Google Scholar] [CrossRef]
- Liu, P.; Prada, V. World Banana Forum. Rome: Food and Agriculture Organization of the United Nations, FAO. Available online: http://www.fao.org/economic/worldbananaforum/en/ (accessed on 29 April 2020).
- Matichenkov, V.; Bocharnikova, E.; Romanova, A.; Doullet, P. Growth of Bacillus amyloliquefaciens as influence by Si nutrition. Arch. Microbiol. 2021, 203, 4329–4336. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Jeong, B.R. Importance of silicon in fruit nutrition: Agronomic and physiological implications. Fruit Crops. 2020, 19, 255–277. [Google Scholar]
- Gbongue, L.R.; Lalaymia, I.; Zeze, A.; Delvaux, B.; Declerck, S. Increased silicon acquisition in bananas colonized by Rhizophagus irregularis MUCL 41833 reduces the incidence of Pseudocercospora fijiensis. Front. Plant. Sci. 2019, 9, 1977. [Google Scholar] [CrossRef] [PubMed]
- Putrie, R.F.W.; Aryantha, I.N.P.; Antonius, S. The structure characteristic of IAA n-acetyl-transferase enzyme produced by two species of bacteria (Bacillus subtilis and Bacillus amyloliquefaciens). IOP Conf. Ser. Earth Environ. Sci. 2021, 762, 012054. [Google Scholar] [CrossRef]
- Jayanti, R.M.; Joko, T. Research article plant growth promoting and antagonistic potential of endophytic bacteria isolated from melon in Indonesia. Plant. Pathol. J. 2020, 19, 200–210. [Google Scholar]
- Zulkarnaini, Z.M.; Sakimin, S.Z.; Mohamed, M.T.M.; Jaafar, H.Z.E. Relationship between chlorophyll content and soil plant analytical development values in two cultivars of fig (Ficus carica L.) as brassinolide effect at an open field. IOP Conf. Ser. Earth Environ. Sci. 2019, 250, 012025. [Google Scholar] [CrossRef]
- Hoang, H.L.; de Guzman, C.C.; Cadiz, N.M.; Hoang, T.T.H.; Tran, D.H.; Rehman, H. Salicylic acid and calcium signaling induce physiological and phytochemical changes to improve salinity tolerance in Red Amaranth (Amaranthus tricolor L.). J. Soil Sci. Plant. Nutr. 2020, 20, 1759–1769. [Google Scholar] [CrossRef]
- Wong, C.K.F.; Zulperi, D.; Saidi, N.B.; Vadamalai, G. A Consortium of Pseudomonas aeruginosa and Trichoderma harzianum for improving growth and induced biochemical changes in Fusarium wilt infected bananas. Trop. Life Sci. Res. 2021, 32, 23–45. [Google Scholar] [CrossRef]
- Din, S.N.M.; Sakimin, S.Z.; Sijam, K.; Baghdadi, A.; Zakaria, M.A.T. Potential of Bacillus subtilis inoculated on biorichar amended soil for suppression of Fusarium wilt, biochemical changes and leaf gas exchange under water stress condition of banana (Musa acuminata) cv. Berangan. Fundam. Appl. Agric. 2018, 3, 515–524. [Google Scholar] [CrossRef]
- Yi, U.; Zaharah, S.S.; Ismail, S.I.; Musa, M.H. Effect of Aqueous Neem Leaf Extracts in Controlling Fusarium Wilt, Soil Physicochemical Properties and Growth Performance of Banana (Musa spp.). Sustainability 2021, 13, 12335. [Google Scholar] [CrossRef]
- Damodaran, T.; Rajan, S.; Gopal, R.; Yadav, A.; Yadav, K.; Shukla, P.K.; Mishra, V.K. Successful community-based management of banana wilt caused by Fusarium oxysporum f. sp. cubense Tropical race-4 through ICAR-FUSICONT. J. Appl. Hortic. 2019, 21, 37–41. [Google Scholar] [CrossRef]
- Bubici, G.; Kaushal, M.; Prigigallo, M.I.; Gómez-Lama Cabanás, C.; Mercado-Blanco, J. Biological control agents against Fusarium wilt of banana. Front. Microbiol. 2019, 10, 616. [Google Scholar] [CrossRef] [PubMed]
- Zorrilla-Fontanesi, Y.; Pauwels, L.; Panis, B.; Signorelli, S.; Vanderschuren, H.; Swennen, R. Strategies to revise agrosystems and breeding to control Fusarium wilt of banana. Nat. Food. 2020, 1, 599–604. [Google Scholar] [CrossRef]
- Kashyap, A.; Planas-Marquès, M.; Capellades, M.; Valls, M.; Coll, N.S. Blocking intruders: Inducible physico-chemical barriers against plant vascular wilt pathogens. J. Exp. Bot. 2021, 72, 184–198. [Google Scholar] [CrossRef]
- Shaheen, N.; Khan, U.M.; Azhar, M.T.; Tan, D.K.; Atif, R.M.; Israr, M.; Rana, I.A. Genetics and genomics of Fusarium wilt of chilies: A Review. Agronomy 2021, 11, 2162. [Google Scholar] [CrossRef]
- Niwas, R.; Chand, G.; Azad, C.S. In vitro evaluation of fungicides against growth of Fusarium oxysporum f. sp. cubense causing panama wilt disease of Banana. Int. J. Commun. Syst. 2020, 8, 191–194. [Google Scholar]
- Pant, B.; Manandhar, S.; Manandhar, C.; Baidhya, S. Invitro Evaluation of Fungicides and Botanicals against Fusarium oxysporum f. sp. cubense of Banana. J. Plant Prot. Soc. 2020, 6, 118–126. [Google Scholar]
- Mbasa, W.V.; Nene, W.A.; Kapinga, F.A.; Lilai, S.A.; Tibuhwa, D.D. Characterization and chemical management of cashew Fusarium wilt disease caused by Fusarium oxysporum in Tanzania. Crop. Prot. 2021, 139, 105379. [Google Scholar] [CrossRef]
- Zhou, X.; Shen, Y.; Fu, X.; Wu, F. Application of sodium silicate enhances cucumber resistance to Fusarium wilt and alters soil microbial communities. Front. Plant. Sci. 2018, 9, 624. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Zhang, Y.; Chen, Y.; Tan, X.; Su, P.; Liu, Y. Integrated control of potato late blight with a combination of the photosynthetic bacterium Rhodopseudomonas palustris strain GJ-22 and fungicides. Biocontrol 2020, 65, 635–645. [Google Scholar] [CrossRef]
- Shah, A.A.; Yasin, N.A.; Akram, K.; Ahmad, A.; Khan, W.U.; Akram, W.; Akbar, M. Ameliorative role of Bacillus subtilis FBL-10 and silicon against lead induced stress in Solanum melongena. Plant. Physiol. Biochem. 2021, 158, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Castiglione, A.M.; Mannino, G.; Contartese, V.; Bertea, C.M.; Ertani, A. Microbial biostimulants as response to modern agriculture needs: Composition, role and application of these innovative products. Plants 2021, 10, 1533. [Google Scholar] [CrossRef] [PubMed]
- Fouda, A.; Eid, A.M.; Elsaied, A.; El-Belely, E.F.; Barghoth, M.G.; Azab, E.; Hassan, S.E.D. Plant growth-promoting endophytic bacterial community inhabiting the leaves of Pulicaria incisa (Lam.) DC inherent to arid regions. Plants 2021, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Kharshandi, F.; Khyllep, A.; Kayang, H. Plant growth-promoting rhizobacteria of Curcuma longa L. and their impact on its growth. Proc. Biol. Sci. 2021, 91, 769–776. [Google Scholar] [CrossRef]
- Sorokan, A.; Veselova, S.; Benkovskaya, G.; Maksimov, I. Endophytic strain Bacillus subtilis 26D increases levels of phytohormones and repairs growth of Potato Plants after Colorado Potato Beetle Damage. Plants 2021, 10, 923. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Huang, W.; Huang, S.; Cui, B.; Dong, Y.; Guo, A.; Jin, Y. Recognition of banana Fusarium wilt based on UAV remote sensing. Remote Sens. 2020, 12, 938. [Google Scholar] [CrossRef]
- Raiesi, T.; Golmohammadi, M. Changes in nutrient concentrations and biochemical characteristics of Mexican lime (Citrus aurantifolia) infected by phytoplasma. J. Gen. Plant. Pathol. 2020, 86, 486–493. [Google Scholar] [CrossRef]
- Awan, Z.A.; Shoaib, A. Combating early blight infection by employing Bacillus subtilis in combination with plant fertilizers. Curr. Plant. Biol. 2019, 20, 100125. [Google Scholar]
- Ahmad, Z.; Waraich, E.A.; Akhtar, S.; Anjum, S.; Ahmad, T.; Mahboob, W.; Rizwan, M. Physiological responses of wheat to drought stress and its mitigation approaches. Acta Physiol. Plant. 2018, 40, 1–13. [Google Scholar] [CrossRef]
- Win, K.T.; Tanaka, F.; Okazaki, K.; Ohwaki, Y. The ACC deaminase expressing endophyte Pseudomonas spp. Enhances NaCl stress tolerance by reducing stress-related ethylene production, resulting in improved growth, photosynthetic performance, and ionic balance in tomato plants. Plant. Physiol. Biochem. 2018, 127, 599–607. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, L.; Yan, B.; Zhu, Y.; Ma, H.; Chen, W.; Ruan, S. Comparative transcriptome analysis reveals the transcriptional alterations in growth-and development-related genes in sweet potato plants infected and non-infected by SPFMV, SPV2, and SPVG. Int. J. Mol. Sci. 2019, 20, 1012. [Google Scholar] [CrossRef] [PubMed]
- Tagoe, S.M.A.; Mensah, T.A.; Asare, A.T. Effect of rust (Uromyces Phaseoli var. Vignae) infection on photosynthetic efficiency, growth and yield potentials of cowpea (Vigna Unguiculata L. Walp) in an open field system. GJSFR 2020, 20, 49–61. [Google Scholar] [CrossRef]
- Sperdouli, I.; Mellidou, I.; Moustakas, M. Harnessing chlorophyll fluorescence for phenotyping analysis of wild and cultivated tomato for high photochemical efficiency under water deficit for climate change resilience. Climate 2021, 9, 154. [Google Scholar] [CrossRef]
- Zakaria, M.A.T.; Sakimin, S.Z.; Juraimi, A.S.; Ramlan, M.F.; Jaafar, H.Z.; Baghdadi, A.; Din, S.N.M. Effect of different water regimes and plant growth regulators on growth, physiology and yield of banana (Musa acuminata cv. Berangan) in tropical climate. Fundam. Appl. Agric. 2018, 3, 505–514. [Google Scholar] [CrossRef]
- Singh, P.; Singh, J.; Ray, S.; Rajput, R.S.; Vaishnav, A.; Singh, R.K.; Singh, H.B. Seed biopriming with antagonistic microbes and ascorbic acid induce resistance in tomato against Fusarium wilt. Microbiol. Res. 2020, 237, 126482. [Google Scholar] [CrossRef] [PubMed]
- Wallis, C.M.; Galarneau, E.R.A. Phenolic compound induction in plant-microbe and plant-insect interactions: A meta-analysis. Front. Plant. Sci. 2020, 11, 2034. [Google Scholar] [CrossRef] [PubMed]
- Hamzah, K.A.; Younes, H.A.; Behiry, S.I.; Abdelkhalek, A. Act of malondialdehyde and total phenolic content under bean yellow mosaic virus infection and biostimulants application. Egypt. Acad. J. Biol. Sci. 2021, 12, 39–42. [Google Scholar] [CrossRef]
- Bouzoumita, A.; Metoui, M.; Jemni, M.; Kabaeir, N.; Belhouchette, K.; Ferchichi, A. The efficacy of various bacterial organisms for biocontrol of Fusarium root rot of olive in Tunisia. Pol. J. Environ. Stud. 2019, 29, 11–16. [Google Scholar] [CrossRef]
- Maulidah, N.I.; Tseng, T.S.; Chen, G.H.; Hsieh, H.Y.; Chang, S.F.; Chuang, H.W. Transcriptome analysis revealed cellular pathways associated with abiotic stress tolerance and disease resistance induced by Pseudomonas aeruginosa in banana plants. Plant Gene 2021, 27, 100321. [Google Scholar] [CrossRef]
- Thakral, V.; Bhat, J.A.; Kumar, N.; Myaka, B.; Sudhakaran, S.; Patil, G.; Deshmukh, R. Role of silicon under contrasting biotic and abiotic stress conditions provides benefits for climate smart cropping. Environ. Exp. Bot. 2021, 189, 104545. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Pseudo-Stem Diameter | Canopy Diameter | Total Chlorophyll Content | Root Electrolyte Leakage |
|---|---|---|---|---|
| (cm) | (cm) | (mgcm−2) | (%) | |
| Control | 16.02 c | 188.60 c | 4.33 c | 71.27 a |
| CBZ | 19.75 b | 290.25 b | 5.56 b | 67.03 ab |
| SS + BS | 18.87 b | 291.50 b | 6.98 a | 61.41 b |
| CBZ + SS + BS | 21.87 a | 368.5 a | 6.69 a | 61.27 b |
| LSD p < 0.05 | 2.08 ** | 36.18 *** | 1.03 ** | 7.10 * |
| Treatments | Stomata Conductance | Transpiration Rate | Vapour Pressure Deficit |
|---|---|---|---|
| (mmol m−2s−1) | (mmol H2O m−2s−1) | (mol H2O m−2s−1) | |
| Control | 0.475 a | 2.98 b | 0.704 b |
| CBZ | 0.476 a | 7.85 a | 1.887 a |
| SS + BS | 0.467 a | 7.91 a | 1.925 a |
| CBZ + SS + BS | 0.501 a | 8.19 a | 1.879 a |
| LSD p < 0.05 | NS | 1.43 *** | 0.17 *** |
| Treatments | Total Phenolic Content | Lignin Content | Malondialdehyde Content |
|---|---|---|---|
| (µg/g FW) | (LTGAg−1 Tissue) | (µmol g−1 FW) | |
| Control | 142.59 a | 0.79 b | 0.71 a |
| CBZ | 117.56 b | 1.39 ab | 0.51 b |
| SS + BS | 130.35 ab | 1.91 a | 0.48 b |
| CBZ + SS + BS | 95.60 c | 1.87 a | 0.39 c |
| LSD p < 0.05 | 16.23 *** | 0.85 * | 0.05 *** |
| Treatments | N | P | K | Ca | Mg |
|---|---|---|---|---|---|
| (%) | (%) | (%) | (%) | (%) | |
| Control | 0.49 c | 0.68 ab | 46.2 ab | 0.32 c | 0.56 a |
| CBZ | 0.52 c | 0.69 a | 40.38 b | 0.41 b | 0.44 a |
| SS + BS | 0.68 b | 0.63 c | 46.81 a | 0.45 a | 0.51 a |
| CBZ + SS + BS | 1.27 a | 0.65 bc | 46.3 a | 0.44 a | 0.56 a |
| LSD p < 0.05 | 0.03 *** | 0.04 * | 5.86 ** | 0.02 *** | NS |
| Treatments | DR | AUDPC |
|---|---|---|
| (%) | (Unit2) | |
| Control | 0 c | 294.64 a |
| CBZ | 43.64 b | 166.07 b |
| SS + BS | 45.36 b | 160.71 b |
| CBZ + SS + BS | 63.05 a | 108.92 c |
| LSD p < 0.05 | 5.76 *** | 17.88 *** |
| Chla+b | Ps | EL | MDA | TPC | Lignin | DI | |
|---|---|---|---|---|---|---|---|
| Chla+b | 1.0 | ||||||
| Ps | 0.79 *** | 1.0 | |||||
| EL | −0.51 * | −0.72 ** | 1.0 | ||||
| MDA | −0.70 ** | −0.85 *** | 0.66 ** | 1.0 | |||
| TPC | −0.57 * | −0.67 ** | 0.41 | 0.69 ** | 1.0 | ||
| Lignin | 0.59 * | 0.61 * | −0.42 | −0.67 ** | −0.42 * | 1.0 | |
| DI | −0.75 *** | −0.84 *** | 0.65 ** | 0.94 *** | 0.77 *** | −0.70 ** | 1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakaria, M.A.T.; Sakimin, S.Z.; Ismail, M.R.; Ahmad, K.; Kasim, S. A Field Evaluation of Sodium Silicate and Bacillus subtilis on the Growth and Yield of Bananas following Fusarium Wilt Disease Infection. Sustainability 2023, 15, 3141. https://doi.org/10.3390/su15043141
Zakaria MAT, Sakimin SZ, Ismail MR, Ahmad K, Kasim S. A Field Evaluation of Sodium Silicate and Bacillus subtilis on the Growth and Yield of Bananas following Fusarium Wilt Disease Infection. Sustainability. 2023; 15(4):3141. https://doi.org/10.3390/su15043141
Chicago/Turabian StyleZakaria, Md Aiman Takrim, Siti Zaharah Sakimin, Mohd Razi Ismail, Khairulmazmi Ahmad, and Susilawati Kasim. 2023. "A Field Evaluation of Sodium Silicate and Bacillus subtilis on the Growth and Yield of Bananas following Fusarium Wilt Disease Infection" Sustainability 15, no. 4: 3141. https://doi.org/10.3390/su15043141
APA StyleZakaria, M. A. T., Sakimin, S. Z., Ismail, M. R., Ahmad, K., & Kasim, S. (2023). A Field Evaluation of Sodium Silicate and Bacillus subtilis on the Growth and Yield of Bananas following Fusarium Wilt Disease Infection. Sustainability, 15(4), 3141. https://doi.org/10.3390/su15043141