
Regenerative Agriculture—A Literature Review on the Practices and Mechanisms Used to Improve Soil Health


{kind=link}
{kind=link}
Abstract
1. Introduction
- Capturing soil carbon through the photosynthesis of high-biomass-producing plants.
- Improving symbiotic soil microbiota–plant interactions.
- Using biological systems to enhance soil structure and water retention.
- Including livestock, with an anticipated positive impact on ecosystem services.
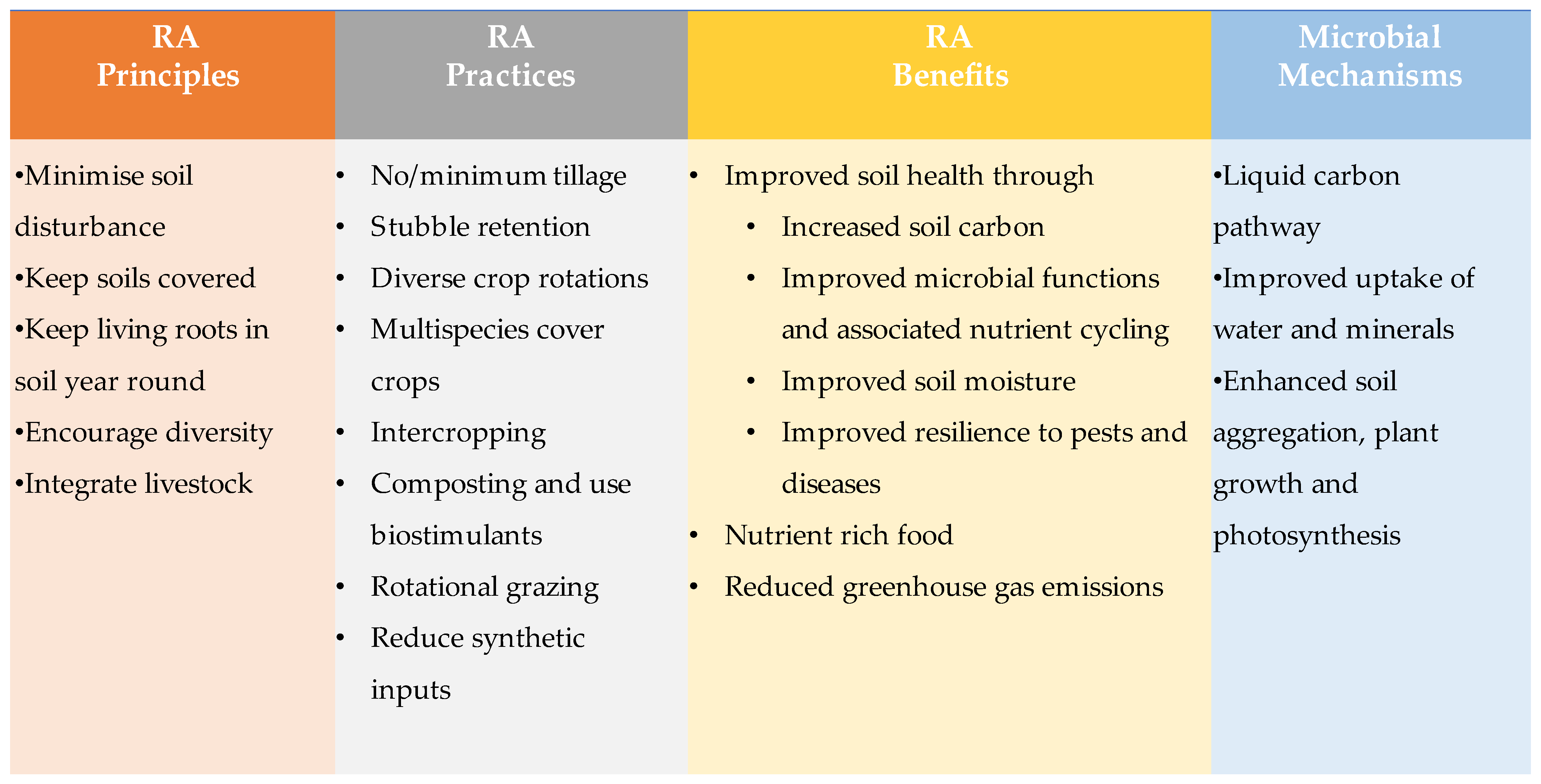
Significance of Regenerative Agriculture to Western Australian Farming Systems
2. Potential Benefits of RA for Soil Health
2.1. Increased Soil Carbon
2.1.1. Minimum/No Tillage
No Tillage and Yield Improvement
No-Tillage and Nitrous Oxide (N2O) Emissions
2.1.2. Cover Crops
2.1.3. Stubble Retention
2.1.4. Crop Rotation and Diversity
2.1.5. Rotational Grazing
2.2. SOM-Mediated Improvement of Soil Moisture/Water Uptake
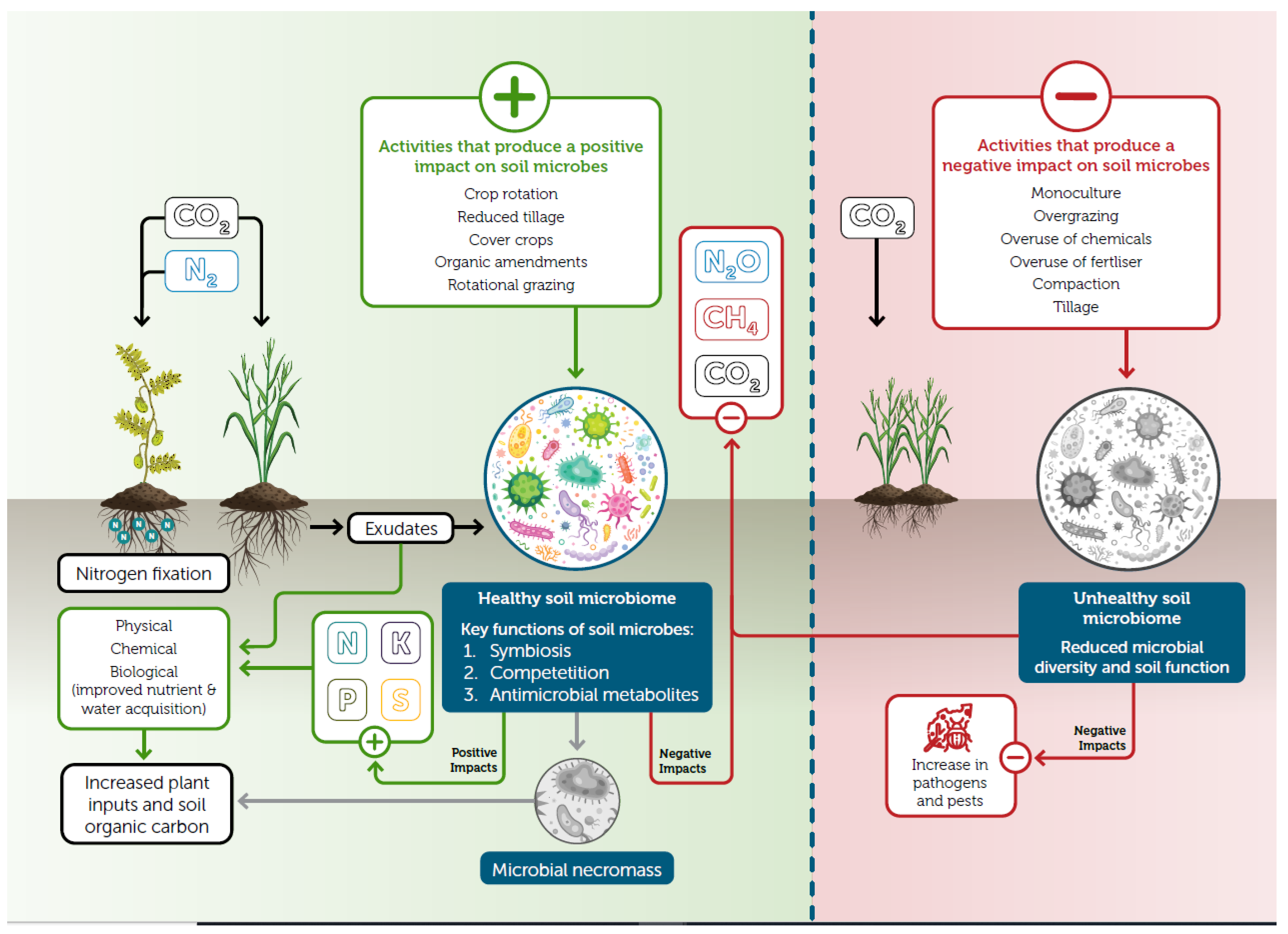
2.3. Increased Soil Biodiversity and Microbial Function
2.3.1. Role of Microbes in Stable Organic Carbon Fraction
2.3.2. Nutrient Cycling and Acquisition
Nitrogen
- 1.
- N2-Fixing Bacteria
- 2.
- Free-Living N2-Fixing Bacteria
Phosphorus (P)
Potassium (K)
Sulphur (S)
3. Pest, Pathogen, and Weed Control/Suppression
4. Mechanisms Involved in Improved Microbial Functions
4.1. Liquid Carbon Pathway
4.2. Improved Uptake of Water
4.2.1. Drought Tolerance
4.2.2. Nutrient Uptake
4.2.3. Role of Soil Microbes in Enhanced Aggregates, Plant Growth, and Photosynthesis
5. Effect of Management Practices on Microbial Activity
5.1. Role of Biofertilisers and Biostimulants
5.2. Effect of Synthetic Inputs on Soil Microbial Activity
6. Nutrient-Dense Food
7. Climate Mitigation
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gabel, M. Ho-Ping: A World Scenario for Food Production; World Game Institute: Philadelphia, PA, USA, 1979. [Google Scholar]
- Rodale, R. Learning to Think Regeneratively. Bull. Sci. Technol. Soc. 1986, 6, 6–13. [Google Scholar] [CrossRef]
- Francis, C.A.; Harwood, R.R.; Parr, J.F. The potential for regenerative agriculture in the developing world. Am. J. Altern. Agric. 1986, 1, 65–74. [Google Scholar] [CrossRef]
- Duchin, F. Drawdown the Most Comprehensive Plan Ever Proposed to Reverse Global Warming. Science 2017, 356, 811. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, S.; Uphoff, N. Soil health: Research, practice and policy for a more regenerative agriculture. Appl. Soil Ecol. 2000, 15, 85–97. [Google Scholar] [CrossRef]
- Rhodes, C.J. The Imperative for Regenerative Agriculture. Sci. Prog. 2017, 100, 80–129. [Google Scholar] [CrossRef]
- Zoveda, F.; Garcia, S.; Pandey, S.; Thomas, G.; Soto, D.; Bianchi, G.; Faures, J.M.; Griffin, J.; Lipper, L.; Vahanen, T.; et al. Building a Common Vision for Sustainable Food and Agriculture; FAO: Rome, Italy, 2014; p. 56. [Google Scholar]
- Burgess, P.J.; Harris, J.G. Regenerative Agriculture: Identifying the Impact, Enabling the Potential. Cranfield, UK. 2019. Available online: http://www.scb.se/statistik/_publikationer/OV9999_2004A01_BR_X97OP0402.pdf (accessed on 5 June 2020).
- Schreefel, L.; Schulte, R.P.; De Boer, I.J.; Schrijver, A.P.; Van Zanten, H.H. Regenerative agriculture–the soil is the base. Glob. Food Secur. 2020, 26, 100404. [Google Scholar] [CrossRef]
- Newton, P.; Civita, N.; Frankel-Goldwater, L.; Bartel, K.; Johns, C. What Is Regenerative Agriculture? A Review of Scholar and Practitioner Definitions Based on Processes and Outcomes. Front. Sustain. Food Syst. 2020, 4, 194. [Google Scholar] [CrossRef]
- Anon. 2020. Available online: https://www.desmog.com/2020/09/11/regenerative-agriculture-criticisms-and-concerns/ (accessed on 18 January 2022).
- Anon. Available online: http://www.australiancollaboration.com.au/pdf/FactSheets/Land-degradation-FactSheet.pdf (accessed on 24 June 2022).
- Kingwell, R.; Carter, C.; Elliott, P.; White, P.; Russia’s Wheat Industry: Implications for Australia. Policy Brief, AEGIC, Australia Department of Agriculture and Food, Perth: Australia Grain Research and Development Corporation GRDC. 2016. Available online: https://aegic.org.au/wp-content/uploads/2021/03/Russia-wheat-industry-Implications-for-Australia.pdf (accessed on 20 January 2022).
- Doran, J.; Sarrantonio, M.; Liebig, M. Soil Health and Sustainability. Adv. Agron. 1996, 56, 1–54. [Google Scholar] [CrossRef]
- Doran, J.W. Soil health and global sustainability: Translating science into practice. Agric. Ecosyst. Environ. 2002, 88, 119–127. [Google Scholar] [CrossRef]
- ITPS 2020. Towards a Definition of Soil Health. Available online: https://www.fao.org/documents/card/fr/c/cb1110en/ (accessed on 13 March 2022).
- Tilman, D.; Fargione, J.; Wolff, B.; D′Antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting Agriculturally Driven Global Environmental Change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G. An underground revolution: Biodiversity and soil ecological engineering for ag-ricultural sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G.A. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef] [PubMed]
- Reeves, D. The role of soil organic matter in maintaining soil quality in continuous cropping systems. Soil Tillage Res. 1997, 43, 131–167. [Google Scholar] [CrossRef]
- Robertson, F.; Armstrong, R.; Partington, D.; Perris, R.; Oliver, I.; Aumann, C.; Crawford, D.; Rees, D. Effect of cropping practices on soil organic carbon: Evidence from long-term field experiments in Victoria, Australia. Soil Res. 2015, 53, 636–646. [Google Scholar] [CrossRef]
- Chabbi, A.; Lehmann, J.; Ciais, P.; Loescher, H.W.; Cotrufo, M.F.; Don, A.; SanClements, M.; Schipper, L.; Six, J.; Smith, P.; et al. Aligning agriculture and climate policy. Nat. Clim. Change 2017, 7, 307–309. [Google Scholar] [CrossRef]
- Poulton, P.; Johnston, J.; Macdonald, A.; White, R.; Powlson, D. Major limitations to achieving “4 per 1000” increases in soil organic carbon stock in temperate regions: Evidence from long-term experiments at Rothamsted Research, United Kingdom. Glob. Change Biol. 2018, 24, 2563–2584. [Google Scholar] [CrossRef]
- De Vries, W. Soil carbon 4 per mille: A good initiative but let′s manage not only the soil but also the expectations: Comment on Minasny et al. Geoderma 292: 59–86. Geoderma 2018, 309, 111–112. [Google Scholar] [CrossRef]
- Lal, R. Promoting “4 Per Thousand” and “Adapting African Agriculture” by south-south cooperation: Conservation agri-culture and sustainable intensification. Soil Tillage Res. 2019, 188, 27–34. [Google Scholar] [CrossRef]
- Rumpel, C.; Amiraslani, F.; Chenu, C.; Garcia Cardenas, M.; Kaonga, M.; Koutika, L.S.; Ladha, J.; Madari, B.; Shirato, Y.; Smith, P.; et al. The 4p1000 initiative: Opportunities, limitations and challenges for implementing soil organic carbon sequestration as a sustainable development strategy. Ambio 2020, 49, 350–360. [Google Scholar] [CrossRef]
- Francaviglia, R.; Di Bene, C.; Farina, R.; Salvati, L.; Vicente-Vicente, J.L. Assessing “4 per 1000” soil organic carbon storage rates under Mediterranean climate: A comprehensive data analysis. Mitig. Adapt. Strat. Glob. Change 2019, 24, 795–818. [Google Scholar] [CrossRef]
- Oldfield, E.E.; Bradford, M.A.; Wood, S.A. Global meta-analysis of the relationship between soil organic matter and crop yields. Soil 2019, 5, 15–32. [Google Scholar] [CrossRef]
- Kane, D.A.; Bradford, M.A.; Fuller, E.; Oldfield, E.E.; Wood, S.A. Soil organic matter protects US maize yields and lowers crop in-surance payouts under drought. Environ. Res. Lett. 2021, 16, 044018. [Google Scholar] [CrossRef]
- Hoyle, F.C.; O’Leary, R.A.; Murphy, D.V. Spatially governed climate factors dominate management in determining the quantity and distribution of soil organic carbon in dryland agricultural systems. Sci. Rep. 2016, 6, 31468. [Google Scholar] [CrossRef] [PubMed]
- Wielemaker, W.G.; Lansu, A.L.E. Land-Use Changes Affecting Classification of a Costa Rican Soil. Soil Sci. Soc. Am. J. 1991, 55, 1621–1624. [Google Scholar] [CrossRef]
- Anderson-Teixeira, K.J.; Davis, S.C.; Masters, M.D.; Delucia, E.H. Changes in soil organic carbon under biofuel crops. GCB Bioenergy 2009, 1, 75–96. [Google Scholar] [CrossRef]
- Nyiraneza, J.; Thompson, B.; Geng, X.; He, J.; Jiang, Y.; Fillmore, S.; Stiles, K. Changes in soil organic matter over 18 year in Prince Edward Island, Canada. Can. J. Soil Sci. 2017, 97, 745–756. [Google Scholar] [CrossRef]
- Sapkota, T.B.; Jat, M.L.; Aryal, J.P.; Jat, R.K.; Khatri-Chhetri, A. Climate change adaptation, greenhouse gas mitigation and economic profitability of conservation agriculture: Some examples from cereal systems of Indo-Gangetic Plains. J. Integr. Agric. 2015, 14, 1524–1533. [Google Scholar] [CrossRef]
- Yang, X.; Drury, C.F.; Wander, M.M. A wide view of no-tillage practices and soil organic carbon sequestration. Acta Agric. Scand. Sect. B Soil Plant Sci. 2013, 63, 523–530. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Chang, S.X.; Cui, S.; Jagadamma, S.; Zhang, Q.; Cai, Y. Residue retention promotes soil carbon accumulation in minimum tillage systems: Implications for conservation agriculture. Sci. Total. Environ. 2020, 740, 140147. [Google Scholar] [CrossRef]
- Haddaway, N.R.; Hedlund, K.; Jackson, L.E.; Kätterer, T.; Lugato, E.; Thomsen, I.K.; Jørgensen, H.B.; Isberg, P.-E. How does tillage intensity affect soil organic carbon? A systematic review. Environ. Evid. 2017, 6, 30. [Google Scholar] [CrossRef]
- Deen, W.; Kataki, P.K. Carbon sequestration in a long-term conventional versus conservation tillage experiment. Soil Tillage Res. 2003, 74, 143–150. [Google Scholar] [CrossRef]
- Chan, K.Y.; Heenan, D.P.; So, H.B. Sequestration of carbon and changes in soil quality under conservation tillage on light-textured soils in Australia: A review. Aust. J. Exp. Agric. 2003, 43, 325–334. [Google Scholar] [CrossRef]
- Young, R.R.; Wilson, B.; Harden, S.; Bernardi, A. Accumulation of soil carbon under zero tillage cropping and perennial vegetation on the Liverpool Plains, eastern Australia. Soil Res. 2009, 47, 273–285. [Google Scholar] [CrossRef]
- Van Rees, H.; Jackman, A.; Baldock, J. Can Soil Organic Carbon Be Increased in a Continuous Cropping System in the Low to Medium Rainfall Zone? 2017. Available online: https://www.hartfieldsite.org.au/media/2017%20TRIAL%20RESULTS/Hart_Trial_Results_2017_Can_soil_carbon_be_increased_in_a_continuous_cropping_system_in_the_low_to_medium_rainfall_zone.pdf (accessed on 23 February 2022).
- Dalal, R.; Strong, W.; Weston, E.; Cooper, J.; Lehane, K.; King, A.; Chicken, C. Sustaining productivity of a Vertisol at Warra, Queensland, with fertilisers, no-tillage, or legumes. 1. Organic matter status. Aust. J. Exp. Agric. 1995, 35, 903–913. [Google Scholar] [CrossRef]
- Cooper, H.V.; Sjögersten, S.; Lark, R.M.; Girkin, N.T.; Vane, C.H.; Calonego, J.C.; Rosolem, C.; Mooney, S.J. Long-term zero-tillage en-hances the protection of soil carbon in tropical agriculture. Eur. J. Soil Sci. 2021, 72, 2477–2492. [Google Scholar] [CrossRef]
- Chivenge, P.; Murwira, H.; Giller, K.; Mapfumo, P.; Six, J. Long-term impact of reduced tillage and residue management on soil carbon stabilization: Implications for conservation agriculture on contrasting soils. Soil Tillage Res. 2007, 94, 328–337. [Google Scholar] [CrossRef]
- Baker, J.M.; Ochsner, T.E.; Venterea, R.T.; Griffis, T.J. Tillage and soil carbon sequestration—What do we really know? Agric. Ecosyst. Environ. 2007, 118, 1–5. [Google Scholar] [CrossRef]
- Martínez, E.; Fuentes, J.P.; Pino, V.; Silva, P.; Acevedo, E. Chemical and biological properties as affected by no-tillage and con-ventional tillage systems in an irrigated Haploxeroll of Central Chile. Soil Tillage Res. 2013, 126, 238–245. [Google Scholar] [CrossRef]
- Powlson, D.S.; Stirling, C.M.; Jat, M.L.; Gerard, B.G.; Palm, C.A.; Sanchez, P.A.; Cassman, K.G. Limited potential of no-till agriculture for climate change mitigation. Nat. Clim. Change 2014, 4, 678–683. [Google Scholar] [CrossRef]
- Baker, J.; Griffis, T. Examining strategies to improve the carbon balance of corn/soybean agriculture using eddy covariance and mass balance techniques. Agric. For. Meteorol. 2005, 128, 163–177. [Google Scholar] [CrossRef]
- Huang, Y.; Ren, W.; Wang, L.; Hui, D.; Grove, J.H.; Yang, X.; Tao, B.; Goff, B. Greenhouse gas emissions and crop yield in no-tillage systems: A meta-analysis. Agric. Ecosyst. Environ. 2018, 268, 144–153. [Google Scholar] [CrossRef]
- Kitonyo, O.M.; Sadras, V.O.; Zhou, Y.; Denton, M.D. Evaluation of historic Australian wheat varieties reveals increased grain yield and changes in senescence patterns but limited adaptation to tillage systems. Field Crops Res. 2017, 206, 65–73. [Google Scholar] [CrossRef]
- Nouri, A.; Lee, J.; Yin, X.; Tyler, D.D.; Saxton, A.M. Thirty-four years of no-tillage and cover crops improve soil quality and increase cotton yield in Alfisols, Southeastern USA. Geoderma 2018, 337, 998–1008. [Google Scholar] [CrossRef]
- Roper, M.M.; Gupta, V.V.S.R.; Murphy, D.V. Tillage practices altered labile soil organic carbon and microbial function without affecting crop yields. Soil Res. 2010, 48, 274–285. [Google Scholar] [CrossRef]
- Pittelkow, C.M.; Liang, X.; Linquist, B.A.; van Groenigen, K.J.; Lee, J.; Lundy, M.E.; van Gestel, N.; Six, J.; Venterea, R.T.; van Kessel, C. Productivity limits and potentials of the principles of conservation agriculture. Nature 2015, 517, 365–368. [Google Scholar] [CrossRef]
- Radford, B.J.; Thornton, C.M. Effects of 27 years of reduced tillage practices on soil properties and crop performance in the semi-arid subtropics of Australia. Int. J. Energy Environ. Econ. 2011, 19, 565. [Google Scholar]
- Sun, W.; Canadell, J.G.; Yu, L.; Yu, L.; Zhang, W.; Smith, P.; Fischer, T.; Huang, Y. Climate drives global soil carbon sequestration and crop yield changes under conservation agriculture. Glob. Change Biol. 2020, 26, 3325–3335. [Google Scholar] [CrossRef]
- Dalal, R.C.; Allen, D.E.; Wang, W.J.; Reeves, S.; Gibson, I. Organic carbon and total nitrogen stocks in a Vertisol following 40 years of no-tillage, crop residue retention and nitrogen fertilisation. Soil Tillage Res. 2011, 112, 133–139. [Google Scholar] [CrossRef]
- Mohammad, W.; Shah, S.M.; Shehzadi, S.; Shah, S.A. Effect of tillage, rotation and crop residues on wheat crop productivity, ferti-lizer nitrogen and water use efficiency and soil organic carbon status in dry area (rainfed) of north-west Pakistan. J. Soil Sci. Plant Nutr. 2012, 12, 715–727. [Google Scholar]
- Page, K.L.; Dang, Y.P.; Dalal, R.C.; Reeves, S.; Thomas, G.; Wang, W.; Thompson, J. Changes in soil water storage with no-tillage and crop residue retention on a Vertisol: Impact on productivity and profitability over a 50 year period. Soil Till. Res. 2019, 194, 104319. [Google Scholar] [CrossRef]
- Lorenz, K.; Lal, R. Biochar application to soil for climate change mitigation by soil organic carbon sequestration. J. Plant Nutr. Soil Sci. 2014, 177, 651–670. [Google Scholar] [CrossRef]
- Lal, R. Regenerative agriculture for food and climate. J. Soil Water Conserv. 2020, 75, 123A–124A. [Google Scholar] [CrossRef]
- Smith, P.; Goulding, K.; Smith, K.; Powlson, D.; Smith, J.; Falloon, P.; Coleman, K. Including trace gas fluxes in estimates of the carbon mitigation potential of UK agricultural land. Soil Use Manag. 2000, 16, 251–259. [Google Scholar] [CrossRef]
- Steinbach, H.S.; Alvarez, R. Changes in Soil Organic Carbon Contents and Nitrous Oxide Emissions after Introduction of No-Till in Pampean Agroecosystems. J. Environ. Qual. 2006, 35, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Burford, J.R.; Bremner, J.M. Relationships between the denitrification capacities of soils and total, water-soluble and readily de-composable soil organic matter. Soil Biol. Biochem. 1975, 7, 389–394. [Google Scholar] [CrossRef]
- Bergstrom, D.W.; Tenuta, M.; Beauchamp, E.G. Increase in nitrous oxide production in soil induced by ammonium and organic carbon. Biol. Fertil. Soils 1994, 18, 1–6. [Google Scholar] [CrossRef]
- Wrage, N.; Velthof, G.L.; van Beusichem, M.L.; Oenema, O. Role of nitrifier denitrification in the production of nitrous oxide. Soil Biol. Biochem. 2001, 33, 1723–1732. [Google Scholar] [CrossRef]
- Thilakarathna, S.K.; Hernandez-Ramirez, G. Primings of soil organic matter and denitrification mediate the effects of moisture on nitrous oxide production. Soil Biol. Biochem. 2021, 155, 108166. [Google Scholar] [CrossRef]
- Weiler, D.A.; Tornquist, C.G.; Parton, W.; dos Santos, H.P.; Santi, A.; Bayer, C. Crop Biomass, Soil Carbon, and Nitrous Oxide as Affected by Management and Climate: A DayCent Application in Brazil. Soil Sci. Soc. Am. J. 2017, 81, 945–955. [Google Scholar] [CrossRef]
- Thilakarathna, S.K.; Hernandez-Ramirez, G.; Puurveen, D.; Kryzanowski, L.; Lohstraeter, G.; Powers, L.; Quan, N.; Tenuta, M. Nitrous oxide emissions and nitrogen use efficiency in wheat: Nitrogen fertilization timing and formulation, soil nitrogen, and weather effects. Soil Sci. Soc. Am. J. 2020, 84, 1910–1927. [Google Scholar] [CrossRef]
- Scott, D.A.; Eckhoff, K.D.; Baer, S.G. Plant diversity decreases potential nitrous oxide emissions from restored agricultural soil. Pedobiologia 2020, 83, 150670. [Google Scholar] [CrossRef]
- Barneze, A.S.; Whitaker, J.; McNamara, N.P.; Ostle, N.J. Legumes increase grassland productivity with no effect on nitrous oxide emissions. Plant Soil 2019, 446, 163–177. [Google Scholar] [CrossRef]
- Yang, Q.; Liu, P.; Dong, S.; Zhang, J.; Zhao, B. Combined application of organic and inorganic fertilizers mitigates ammonia and nitrous oxide emissions in a maize field. Nutr. Cycl. Agroecosyst. 2020, 117, 13–27. [Google Scholar] [CrossRef]
- Cole, C.; Duxbury, J.; Freney, J.; Heinemeyer, O.; Minami, K.; Mosier, A.; Paustian, K.; Rosenberg, N.; Sampson, N.; Sauerbeck, D.; et al. Global estimates of potential mitigation of greenhouse gas emissions by agriculture. Nutr. Cycl. Agroecosyst. 1997, 49, 221–228. [Google Scholar] [CrossRef]
- Swanton, C.J.; Weise, S.F. Integrated Weed Management: The Rationale and Approach. Weed Technol. 1991, 5, 657–663. [Google Scholar] [CrossRef]
- Kumar, H.D.; Aloke, P. Role of biostimulant formulations in crop production: An overview. Int. J. Appl. Res. Vet. M. 2020, 8, 38–46. [Google Scholar]
- Summers, H.; Karsten, H.D.; Curran, W.; Malcolm, G.M. Integrated weed management with reduced herbicides in a no-till dairy rotation. Agron. J. 2021, 113, 3418–3433. [Google Scholar] [CrossRef]
- Finney, D.M.; Murrell, E.G.; White, C.M.; Baraibar, B.; Barbercheck, M.E.; Bradley, B.A.; Cornelisse, S.; Hunter, M.C.; Kaye, J.P.; Mortensen, D.A.; et al. Ecosystem services and disservices are bundled in simple and diverse cover cropping sys-tems. Agric. Environ. Lett. 2017, 2, 170033. [Google Scholar] [CrossRef]
- Kaye, J.P.; Quemada, M. Using cover crops to mitigate and adapt to climate change. A review. Agron. Sustain. Dev. 2017, 37, 4. [Google Scholar] [CrossRef]
- McDaniel, M.D.; Tiemann, L.K.; Grandy, A.S. Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecol. Appl. 2014, 24, 560–570. [Google Scholar] [CrossRef]
- Poeplau, C.; Don, A. Carbon sequestration in agricultural soils via cultivation of cover crops—A meta-analysis. Agric. Ecosyst. Environ. 2015, 200, 33–41. [Google Scholar] [CrossRef]
- Ghimire, R.; Ghimire, B.; Mesbah, A.O.; Idowu, O.J.; O’Neill, M.K.; Angadi, S.V.; Shukla, M.K. Current status, opportunities, and chal-lenges of cover cropping for sustainable dryland farming in the Southern Great Plains. J. Crops Improv. 2018, 32, 579–598. [Google Scholar] [CrossRef]
- Chahal, I.; Vyn, R.J.; Mayers, D.; Van Eerd, L.L. Cumulative impact of cover crops on soil carbon sequestration and profitability in a temperate humid climate. Sci. Rep. 2020, 10, 13381. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, R.; Steinbach, H.S.; De Paepe, J.L. Cover crop effects on soils and subsequent crops in the pampas: A meta-analysis. Soil Tillage Res. 2017, 170, 53–65. [Google Scholar] [CrossRef]
- Hassink, J.; Whitmore, A.P. A model of the physical protection of organic matter in soils. Soil Sci. Soc. Am. J. 1997, 61, 131–139. [Google Scholar] [CrossRef]
- Berhe, A.A.; Harte, J.; Harden, J.W.; Torn, M.S. The Significance of the Erosion-induced Terrestrial Carbon Sink. Bioscience 2007, 57, 337–346. [Google Scholar] [CrossRef]
- Olson, K.; Ebelhar, S.A.; Lang, J.M. Long-Term Effects of Cover Crops on Crop Yields, Soil Organic Carbon Stocks and Sequestration. Open J. Soil Sci. 2014, 4, 284–292. [Google Scholar] [CrossRef]
- Jian, J.; Du, X.; Reiter, M.S.; Stewart, R.D. A meta-analysis of global cropland soil carbon changes due to cover cropping. Soil Biol. Biochem. 2020, 143, 107735. [Google Scholar] [CrossRef]
- Beillouin, D.; Ben-Ari, T.; Malézieux, E.; Seufert, V.; Makowski, D. Positive but variable effects of crop diversification on biodiversity and ecosystem services. Glob. Change Biol. 2021, 27, 4697–4710. [Google Scholar] [CrossRef]
- Miguez, F.E.; Bollero, G.A. Review of Corn Yield Response under Winter Cover Cropping Systems Using Meta-Analytic Methods. Crop Sci. 2005, 45, 2318–2329. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.Y.H.; Chen, C.; Ma, Z.; Searle, E.B.; Yu, Z.; Huang, Z. Effects of plant diversity on soil carbon in diverse ecosystems: A global meta-analysis. Biol. Rev. 2019, 95, 167–183. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, C.F.D.S.; Rodrigues, D.R.; da Silva, G.F.; Echer, F.R.; Calonego, J.C. Soil organic carbon stock is improved by cover crops in a tropical sandy soil. Agron. J. 2022, 114, 1546–1556. [Google Scholar] [CrossRef]
- Seitz, D.; Fischer, L.M.; Dechow, R.; Wiesmeier, M.; Don, A. The potential of cover crops to increase soil organic carbon storage in German croplands. Plant Soil 2022, 1–17. [Google Scholar] [CrossRef]
- Olson, K.R.; Ebelhar, S.A.; Lang, J.M. Cover Crop Effects on Crop Yields and Soil Organic Carbon Content. Soil Sci. 2010, 175, 89–98. [Google Scholar] [CrossRef]
- Nielsen, D.C.; Lyon, D.J.; Higgins, R.K.; Hergert, G.W.; Holman, J.D.; Vigil, M.F. Cover Crop Effect on Subsequent Wheat Yield in the Central Great Plains. Agron. J. 2016, 108, 243–256. [Google Scholar] [CrossRef]
- Flower, K.; Ward, P.; Cordingley, N.; Micin, S.; Craig, N. Rainfall, rotations and residue level affect no-tillage wheat yield and gross margin in a Mediterranean-type environment. Field Crops Res. 2017, 208, 1–10. [Google Scholar] [CrossRef]
- Myers, R.; Watts, C. Progress and perspectives with cover crops: Interpreting three years of farmer surveys on cover crops. J. Soil Water Conserv. 2015, 70, 125A–129A. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Claassen, M.M.; Presley, D.R. Summer cover crops fix nitrogen, increase crop yield, and improve soil–crop relationships. Agron. J. 2012, 104, 137–147. [Google Scholar] [CrossRef]
- Motisi, N.; Montfort, F.; Faloya, V.; Lucas, P.; Doré, T. Growing Brassica juncea as a cover crop, then incorporating its residues provide complementary control of Rhizoctonia root rot of sugar beet. Field Crops Res. 2009, 113, 238–245. [Google Scholar] [CrossRef]
- Duff, J.; Firrell, M. Biofumigation: A Cover Crop Option 12 Months of the Year to Manage Three Soilborne Pathogens Ailing the Australian Vegetable Industry. Glob. J. Agric. Innov. Res. Dev. 2021, 8, 104–116. [Google Scholar] [CrossRef]
- Packer, I.; Hamilton, G.; Koen, T. Runoff, soil loss and soil physical property changes of light textured surface soils from long term tillage treatments. Soil Res. 1992, 30, 789–806. [Google Scholar] [CrossRef]
- Harper, R.; Gilkes, R.; Hill, M.; Carter, D. Wind erosion and soil carbon dynamics in south-western Australia. Aeolian Res. 2010, 1, 129–141. [Google Scholar] [CrossRef]
- Saffigna, P.; Powlson, D.; Brookes, P.; Thomas, G. Influence of sorghum residues and tillage on soil organic matter and soil microbial biomass in an australian vertisol. Soil Biol. Biochem. 1989, 21, 759–765. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via bio-chemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Horwath, W.R.; Kuzyakov, Y. The Potential for Soils to Mitigate Climate Change Through Carbon Sequestration. In Developments in Soil Science; Elsevier: Amsterdam, The Netherlands, 2018; Volume 35, pp. 61–92. [Google Scholar] [CrossRef]
- Jha, P.; Hati, K.; Dalal, R.C.; Dang, Y.P.; Kopittke, P.M.; Menzies, N.W. Soil carbon and nitrogen dynamics in a Vertisol following 50 years of no-tillage, crop stubble retention and nitrogen fertilization. Geoderma 2019, 358, 113996. [Google Scholar] [CrossRef]
- Somasundaram, J.; Reeves, S.; Wang, W.; Heenan, M.; Dalal, R. Impact of 47 years of no tillage and stubble retention on soil ag-gregation and carbon distribution in a vertisol. Land Degrad. Dev. 2017, 28, 1589–1602. [Google Scholar] [CrossRef]
- Hati, K.M.; Jha, P.; Dalal, R.C.; Jayaraman, S.; Dang, Y.P.; Kopittke, P.M.; Kirchhof, G.; Menzies, N.W. 50 years of continuous no-tillage, stubble retention and nitrogen fertilization enhanced macro-aggregate formation and stabilisation in a Vertisol. Soil Tillage Res. 2021, 214, 105163. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Colloff, M.J.; Harvey, P.R.; Marschner, P.; Gregg, A.L.; Rogers, S.L. The effects of stubble retention and nitrogen appli-cation on soil microbial community structure and functional gene abundance under irrigated maize. FEMS Microbiol. Ecol. 2007, 59, 661–670. [Google Scholar] [CrossRef]
- Paustian, K.A.O.J.H.; Andren, O.; Janzen, H.H.; Lal, R.; Smith, P.; Tian, G.; Tiessen, H.; van Noordwijk, M.; Woomer, P.L. Agricultural soils as a sink to mitigate CO2 emissions. Soil Use Manag. 1997, 13, 230–244. [Google Scholar] [CrossRef]
- Dalal, R.C.; Chan, K.Y. Soil organic matter in rainfed cropping systems of the Australian cereal belt. Soil Res. 2001, 39, 435–464. [Google Scholar] [CrossRef]
- Pandey, C. Management of crop residue for sustaining soil fertility and foodgrains production in India. Acta Sci. Agric. 2019, 3, 188–195. [Google Scholar]
- Passaris, N.; Flower, K.; Ward, P.; Cordingley, N. Effect of crop rotation diversity and windrow burning of residue on soil chemical composition under long-term no-tillage. Soil Tillage Res. 2021, 213, 105153. [Google Scholar] [CrossRef]
- Chan, Y. Increasing soil organic carbon of agricultural land. Primefact 2008, 735, 1–5. [Google Scholar]
- Tomar, V.P.S.; Narain, P.; Dadhwal, K.S. Effect of perennial mulches on moisture conservation and soil-building properties through agroforestry. Agrofor. Syst. 1992, 19, 241–252. [Google Scholar] [CrossRef]
- Freibauer, A.; Rounsevell, M.D.; Smith, P.; Verhagen, J. Carbon sequestration in the agricultural soils of Europe. Geoderma 2004, 122, 1–23. [Google Scholar] [CrossRef]
- Liu, D.L.; Anwar, M.R.; O′Leary, G.; Conyers, M.K. Managing wheat stubble as an effective approach to se-quester soil carbon in a semi-arid environment: Spatial modelling. Geoderma 2014, 214, 50–61. [Google Scholar] [CrossRef]
- Campbell, C.A.; Gregorich, E.G.; Zentner, R.P.; Roloff, R.; Janzen, H.H.; Paustian, K.; Smith, W.; Liang, B.C.; McConkey, M.G. Carbon sequestration in the Canadian Prairies: Quantification of Short-Term Dynamics. SSSA Spec. Publ. 2001, 57, 93–114. [Google Scholar]
- Xia, L.; Lam, S.K.; Wolf, B.; Kiese, R.; Chen, D.; Butterbach-Bahl, K. Trade-offs between soil carbon sequestration and reactive nitrogen losses under straw return in global agroecosystems. Glob. Change Biol. 2018, 24, 5919–5932. [Google Scholar] [CrossRef]
- Shi, J.; Wang, S.; Li, S.; Tian, X. Increasing soil organic carbon sequestration and yield stability by no-tillage and straw-returning in wheat–maize rotation. Agron. J. 2022, 114, 1534–1545. [Google Scholar] [CrossRef]
- Di Bene, C.; Marchetti, A.; Francaviglia, R.; Farina, R. Soil organic carbon dynamics in typical durum wheat-based crop rotations of Southern Italy. Ital. J. Agron. 2016, 11, 209–216. [Google Scholar] [CrossRef]
- Jarecki, M.K.; Lal, R. Crop Management for Soil Carbon Sequestration. Crit. Rev. Plant Sci. 2003, 22, 471–502. [Google Scholar] [CrossRef]
- Huang, M.; Shao, M.; Zhang, L.; Li, Y. Water use efficiency and sustainability of different long-term crop rotation systems in the Loess Plateau of China. Soil Tillage Res. 2003, 72, 95–104. [Google Scholar] [CrossRef]
- Li, M.; Guo, J.; Ren, T.; Luo, G.; Shen, Q.; Lu, J.; Guo, S.; Ling, N. Crop rotation history constrains soil biodiversity and multifunctionality relationships. Agric. Ecosyst. Environ. 2021, 319, 107550. [Google Scholar] [CrossRef]
- Blair, N.; Crocker, G.J. Crop rotation effects on soil carbon and physical fertility of two Australian soils. Soil Res. 2000, 38, 71–84. [Google Scholar] [CrossRef]
- Singh, K.; Whelan, B. Soil carbon change across ten New South Wales farms under different farm management regimes in Australia. Soil Use Manag. 2020, 36, 616–632. [Google Scholar] [CrossRef]
- López-Bellido, L.; López-Bellido, R.; Fernández-García, P.; Muñoz-Romero, V.; Lopez-Bellido, F.J. Carbon storage in a rainfed Med-iterranean vertisol: Effects of tillage and crop rotation in a long-term experiment. Eur. J. Soil Sci. 2020, 71, 472–483. [Google Scholar] [CrossRef]
- West, T.O.; Post, W.M. Soil organic carbon sequestration rates by tillage and crop rotation: A global data analysis. Soil Sci. Soc. Am. J. 2002, 66, 1930–1946. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Arshad, M.A. Soil Organic Matter Pools during Early Adoption of Conservation Tillage in Northwestern Canada. Soil Sci. Soc. Am. J. 1996, 60, 1422–1427. [Google Scholar] [CrossRef]
- Lal, R. Carbon Sequestration in Dryland Ecosystems. Environ. Manag. 2003, 33, 528–544. [Google Scholar] [CrossRef]
- Wochesländer, R.; Harper, R.J.; Sochacki, S.R.; Ward, P.R.; Revell, C. Tagasaste (Cytisus proliferus Link. ) reforestation as an option for carbon mitigation in dryland farming systems. Ecol. Eng. 2016, 97, 610–618. [Google Scholar]
- Hoyle, F.C.; D′Antuono, M.; Overheu, T.; Murphy, D.V. Capacity for increasing soil organic carbon stocks in dryland agricultural systems. Soil Res. 2013, 51, 657–667. [Google Scholar] [CrossRef]
- Havlin, J.L.; Kissel, D.E.; Maddux, L.D.; Claassen, M.M.; Long, J.H. Crop Rotation and Tillage Effects on Soil Organic Carbon and Nitrogen. Soil Sci. Soc. Am. J. 1990, 54, 448–452. [Google Scholar] [CrossRef]
- Liu, K.; Bandara, M.; Hamel, C.; Knight, J.D.; Gan, Y. Intensifying crop rotations with pulse crops enhances system productivity and soil organic carbon in semi-arid environments. Field Crops Res. 2019, 248, 107657. [Google Scholar] [CrossRef]
- Sanderman, J. Can management induced changes in the carbonate system drive soil carbon sequestration? A review with particular focus on Australia. Agric. Ecosyst. Environ. 2012, 155, 70–77. [Google Scholar] [CrossRef]
- Seó, H.L.S.; Filho, L.C.P.M.; Brugnara, D. Rationally Managed Pastures Stock More Carbon than No-Tillage Fields. Front. Environ. Sci. 2017, 5, 87. [Google Scholar] [CrossRef]
- Díaz-Solís, H.; Grant, W.; Kothmann, M.; Teague, W.; Díaz-García, J. Adaptive management of stocking rates to reduce effects of drought on cow-calf production systems in semi-arid rangelands. Agric. Syst. 2009, 100, 43–50. [Google Scholar] [CrossRef]
- Teague, R.; Provenza, F.; Norton, B.; Steffens, T.; Barnes, M.; Kothmann, M.; Roath, R. Benefits of Multi-Paddock Grazing Management on Rangelands: Limitations of Experimental Grazing Research and Knowledge Gaps. Grasslands: Ecology, Management and Restoration; Nova Science Publishers: Hauppauge, NY, USA, 2008; pp. 41–80. [Google Scholar]
- Teague, W.; Dowhower, S.; Baker, S.; Haile, N.; DeLaune, P.; Conover, D. Grazing management impacts on vegetation, soil biota and soil chemical, physical and hydrological properties in tall grass prairie. Agric. Ecosyst. Environ. 2011, 141, 310–322. [Google Scholar] [CrossRef]
- Byrnes, R.C.; Eastburn, D.J.; Tate, K.W.; Roche, L.M. A Global Meta-Analysis of Grazing Impacts on Soil Health Indicators. J. Environ. Qual. 2018, 47, 758–765. [Google Scholar] [CrossRef]
- Conant, R.T.; Paustian, K.; Elliott, E.T. Grassland management and conversion into grassland: Effects on soil carbon. Ecol. Appl. 2001, 11, 343–355. [Google Scholar] [CrossRef]
- Follett, R.; Stewart, C.; Bradford, J.; Pruessner, E.; Sims, P.L.; Vigil, M. Long-term pasture management impacts on eolian sand soils in the southern mixed-grass prairie. Quat. Int. 2020, 565, 84–93. [Google Scholar] [CrossRef]
- Chan, K.Y.; Oates, A.; Li, G.D.; Conyers, M.K.; Prangnell, R.J.; Poile, G.; Liu, D.L.; Barchia, I.M. Soil carbon stocks under different pastures and pasture management in the higher rainfall areas of south-eastern Australia. Soil Res. 2010, 48, 7–15. [Google Scholar] [CrossRef]
- Sanderman, J.; Reseigh, J.; Wurst, M.; Young, M.-A.; Austin, J. Impacts of Rotational Grazing on Soil Carbon in Native Grass-Based Pastures in Southern Australia. PLoS ONE 2015, 10, e0136157. [Google Scholar] [CrossRef]
- Mosier, S.; Apfelbaum, S.; Byck, P.; Calderon, F.; Teague, R.; Thompson, R.; Cotrufo, M.F. Adaptive multi-paddock grazing enhances soil carbon and nitrogen stocks and stabilization through mineral association in southeastern U.S. grazing lands. J. Environ. Manag. 2021, 288, 112409. [Google Scholar] [CrossRef] [PubMed]
- Hiernaux, P.; Bielders, C.L.; Valentin, C.; Bationo, A.; Fernández-Rivera, S. Effects of livestock grazing on physical and chemical properties of sandy soils in Sahelian rangelands. J. Arid. Environ. 1999, 41, 231–245. [Google Scholar] [CrossRef]
- Reeder, J.; Schuman, G. Influence of livestock grazing on C sequestration in semi-arid mixed-grass and short-grass rangelands. Environ. Pollut. 2001, 116, 457–463. [Google Scholar] [CrossRef]
- Abdalla, M.; Hastings, A.; Chadwick, D.; Jones, D.; Evans, C.; Jones, M.; Rees, R.; Smith, P. Critical review of the impacts of grazing intensity on soil organic carbon storage and other soil quality indicators in extensively managed grasslands. Agric. Ecosyst. Environ. 2017, 253, 62–81. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Cotrufo, M.F. Grassland soil carbon sequestration: Current understanding, challenges, and solutions. Science 2022, 377, 603–608. [Google Scholar] [CrossRef]
- Maestre, F.T.; Le Bagousse-Pinguet, Y.; Delgado-Baquerizo, M.; Eldridge, D.J.; Saiz, H.; Berdugo, M.; Gozalo, B.; Ochoa, V.; Guirado, E.; García-Gómez, M.; et al. Grazing and ecosystem service delivery in global drylands. Science 2022, 378, 915–920. [Google Scholar] [CrossRef]
- Thomasson, A.J.; Carter, A.D. Current and future uses of the UK soil water retention dataset. In Proceedings of the International Workshop on Indirect Methods for Estimating the Hydraulic Properties of Unsaturated Soils, Riverside, CA, USA, 11 October 1989; pp. 355–358. [Google Scholar]
- Loveland, P.; Webb, J. Is there a critical level of organic matter in the agricultural soils of temperate regions: A review. Soil Tillage Res. 2003, 70, 1–8. [Google Scholar] [CrossRef]
- Lal, R. Physical properties and moisture retention characteristics of some nigerian soils. Geoderma 1978, 21, 209–223. [Google Scholar] [CrossRef]
- Danalatos, N.; Kosmas, C.; Driessen, P.; Yassoglou, N. Estimation of the draining soil moisture characteristic from standard data as recorded in routine soil surveys. Geoderma 1994, 64, 155–165. [Google Scholar] [CrossRef]
- Maynard, A.A. Compost: The process and research. Bull. Conn. Agric. Exp. Stn. 2000, 13. [Google Scholar]
- Hudson, B.D. Soil organic matter and available water capacity. J. Soil Water Conserv. 1994, 49, 189–194. [Google Scholar]
- Rawls, W.; Pachepsky, Y.; Ritchie, J.; Sobecki, T.; Bloodworth, H. Effect of soil organic carbon on soil water retention. Geoderma 2003, 116, 61–76. [Google Scholar] [CrossRef]
- De Jong, R.; Campbell, C.A.; Nicholaichuk, W. Water retention equations and their relationship to soil organic matter and particle size distribution for disturbed samples. Can. J. Soil Sci. 1983, 63, 291–302. [Google Scholar] [CrossRef]
- Haynes, R.; Naidu, R. Influence of lime, fertilizer and manure applications on soil organic matter content and soil physical conditions: A review. Nutr. Cycl. Agroecosyst. 1998, 51, 123–137. [Google Scholar] [CrossRef]
- Wolf, B.; Snyder, G. Sustainable Soils: The Place of Organic Matter in Sustaining Soils and Their Productivity; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Emerson, W.W.; McGarry, D. Organic carbon and soil porosity. Soil Res. 2003, 41, 107–118. [Google Scholar] [CrossRef]
- Libohova, Z.; Seybold, C.; Wysocki, D.; Wills, S.; Schoeneberger, P.; Williams, C.; Lindbo, D.; Stott, D.; Owens, P. Reevaluating the effects of soil organic matter and other properties on available water-holding capacity using the National Cooperative Soil Survey Characterization Database. J. Soil Water Conserv. 2018, 73, 411–421. [Google Scholar] [CrossRef]
- Minasny, B.; McBratney, A.B. Limited effect of organic matter on soil available water capacity. Eur. J. Soil Sci. 2018, 69, 39–47. [Google Scholar] [CrossRef]
- Stewart, C.E.; Paustian, K.; Conant, R.; Plante, A.; Six, J. Soil carbon saturation: Concept, evidence and evaluation. Biogeochemistry 2007, 86, 19–31. [Google Scholar] [CrossRef]
- Williams, A.; Hunter, M.C.; Kammerer, M.; Kane, D.A.; Jordan, N.R.; Mortensen, D.A.; Smith, R.G.; Snapp, S.; Davis, A.S. Soil water holding capacity mitigates downside risk and volatility in US rainfed maize: Time to invest in soil organic matter? PLoS ONE 2016, 11, e0160974. [Google Scholar] [CrossRef] [PubMed]
- Bhadha, J.H.; Capasso, J.M.; Khatiwada, R.; Swanson, S.; LaBorde, C. Raising Soil Organic Matter Content to Improve Water Holding Capacity; University of Florida Institute of Food and Agricultural Sciences: Gainesville, FL, USA, 2017. [Google Scholar]
- Oldfield, E.E.; Wood, S.A.; Bradford, M.A. Direct evidence using a controlled greenhouse study for threshold effects of soil organic matter on crop growth. Ecol. Appl. 2020, 30, e02073. [Google Scholar] [CrossRef]
- Parr, J.; Bertrand, A. Water Infiltration into Soils. Adv. Agron. 1960, 12, 311–363. [Google Scholar] [CrossRef]
- De Vries, F.T.; Thébault, E.; Liiri, M.; Birkhofer, K.; Tsiafouli, M.A.; Bjørnlund, L.; Bracht Jørgensen, H.; Brady, M.V.; Christensen, S.; De Ruiter, P.C.; et al. Soil food web properties explain ecosystem services across European land use systems. Proc. Natl. Acad. Sci. USA 2013, 110, 14296–14301. [Google Scholar] [CrossRef]
- Lehmann, A.; Zheng, W.; Rillig, M.C. Soil biota contributions to soil aggregation. Nat. Ecol. Evol. 2017, 1, 1828–1835. [Google Scholar] [CrossRef] [PubMed]
- Calegari, A.; Darolt, M.R.; Ferro, M. Towards sustainable agriculture with a no-tillage system. Adv. GeoEcol. 1998, 31, 1205–1210. [Google Scholar]
- Eden, M.; Gerke, H.H.; Houot, S. Organic waste recycling in agriculture and related effects on soil water retention and plant available water: A review. Agron. Sustain. Dev. 2017, 37, 11. [Google Scholar] [CrossRef]
- Li, Y.; Shao, M.; Wang, J.; Li, T. Effects of earthworm cast application on water evaporation and storage in loess soil column ex-periments. Sustainability 2020, 12, 3112. [Google Scholar] [CrossRef]
- Hallam, J.; Hodson, M.E. Impact of different earthworm ecotypes on water stable aggregates and soil water holding capacity. Biol. Fertil. Soils 2020, 56, 607–617. [Google Scholar] [CrossRef]
- Liang, C.; Balser, T.C. Microbial production of recalcitrant organic matter in global soils: Implications for productivity and climate policy. Nat. Rev. Microbiol. 2011, 9, 75. [Google Scholar] [CrossRef]
- Lal, R. Soil organic matter and water retention. Agron. J. 2020, 112, 3265–3277. [Google Scholar] [CrossRef]
- Al-Kaisi, M.M.; Douelle, A.; Kwaw-Mensah, D. Soil microaggregate and macroaggregate decay over time and soil carbon change as influenced by different tillage systems. J. Soil Water Conserv. 2014, 69, 574–580. [Google Scholar] [CrossRef]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Wagg, C.; Schlaeppi, K.; Banerjee, S.; Kuramae, E.E.; Van Der Heijden, M.G.A. Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning. Nat. Commun. 2019, 10, 4841. [Google Scholar] [CrossRef]
- Brussaard, L.; de Ruiter, P.C.; Brown, G.G. Soil biodiversity for agricultural sustainability. Agric. Ecosyst. Environ. 2007, 121, 233–244. [Google Scholar] [CrossRef]
- Wagg, C.; van Erk, A.; Fava, E.; Comeau, L.-P.; Mitterboeck, T.F.; Goyer, C.; Li, S.; McKenzie-Gopsill, A.; Mills, A. Full-Season Cover Crops and Their Traits That Promote Agroecosystem Services. Agriculture 2021, 11, 830. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Tiedje, J.M.; Asuming-Brempong, S.; Nüsslein, K.; Marsh, T.L.; Flynn, S.J. Opening the black box of soil microbial diversity. Appl. Soil Ecol. 1999, 13, 109–122. [Google Scholar] [CrossRef]
- Jastrow, J.D.; Amonette, J.E.; Bailey, V.L. Mechanisms controlling soil carbon turnover and their potential application for enhancing carbon sequestration. Clim. Change 2006, 80, 5–23. [Google Scholar] [CrossRef]
- Graham, E.B.; Knelman, J.E.; Schindlbacher, A.; Siciliano, S.; Breulmann, M.; Yannarell, A.; Beman, J.M.; Abell, G.; Philippot, L.; Prosser, J.; et al. Microbes as engines of ecosystem function: When does community structure enhance predictions of ecosystem processes? Front. Microbiol. 2016, 7, 214. [Google Scholar] [CrossRef] [PubMed]
- Lamanna, C.; Blonder, B.; Violle, C.; Kraft, N.J.; Sandel, B.; Šímová, I.; Donoghue, J.C.; Svenning, J.C.; McGill, B.J.; Boyle, B.; et al. Functional trait space and the latitudinal diversity gradient. Proc. Natl. Acad. Sci. USA 2014, 111, 13745–13750. [Google Scholar] [CrossRef]
- Brady, N.C. The Nature and Properties of Soils; Macmillan Publishing Company: New York, NY, USA, 1990. [Google Scholar]
- Duxbury, J.M. Soil organic matter as a source and a sink of plant nutrients. In Dynamics of Soil Organic Matter in Tropical Ecosystems; NifTAL Project, University Hawaii: Maui, HI, USA, 1989. [Google Scholar]
- Sidorova, I.; Voronina, E. Microbiome-Driven Nutrient Fortification in Plants: The Role of Microbiota in Chemical Transfor-mation and Nutrient Mobilization. In The Plant Microbiome in Sustainable Agriculture; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 211–230. [Google Scholar]
- Veresoglou, S.D.; Halley, J.M.; Rillig, M.C. Extinction risk of soil biota. Nat. Commun. 2015, 6, 8862. [Google Scholar] [CrossRef] [PubMed]
- Orgiazzi, A.; Panagos, P.; Yigini, Y.; Dunbar, M.B.; Gardi, C.; Montanarella, L.; Ballabio, C. A knowledge-based approach to estimating the magnitude and spatial patterns of potential threats to soil biodiversity. Sci. Total. Environ. 2016, 545–546, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Tibbett, M.; Fraser, T.D.; Duddigan, S. Identifying potential threats to soil biodiversity. PeerJ 2020, 8, e9271. [Google Scholar] [CrossRef] [PubMed]
- Rees, R.M.; Ball, B.C.; Campbell, C.D.; Watson, C.A. Sustaining soil organic matter. In Sustainable Management of Soil Organic Matter; Cabi Publishing: Wallingford, UK, 2001; pp. 413–425. [Google Scholar]
- Bardgett, R.D.; Hobbs, P.J.; Frostegård, Å. Changes in soil fungal: Bacterial biomass ratios following reductions in the intensity of management of an upland grassland. Biol. Fertil. Soils 1996, 22, 261–264. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Manzoni, S.; Moorhead, D.L.; Richter, A. Carbon use efficiency of microbial communities: Stoichiometry, methodology and modelling. Ecol. Lett. 2013, 16, 930–939. [Google Scholar] [CrossRef]
- Strickland, M.S.; Rousk, J. Considering fungal: Bacterial dominance in soils–methods, controls, and ecosystem implications. Soil Biol. Biochem. 2010, 42, 1385–1395. [Google Scholar] [CrossRef]
- Waring, B.G.; Averill, C.; Hawkes, C.V. Differences in fungal and bacterial physiology alter soil carbon and nitrogen cycling: Insights from meta-analysis and theoretical models. Ecol. Lett. 2013, 16, 887–894. [Google Scholar] [CrossRef]
- Dalal, R.C. Soil microbial biomass—What do the numbers really mean? Aust. J. Exp. Agric. 1998, 38, 649–665. [Google Scholar] [CrossRef]
- Xu, X.; Thornton, P.E.; Post, W.M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2013, 22, 737–749. [Google Scholar] [CrossRef]
- Miltner, A.; Bombach, P.; Schmidt-Brücken, B.; Kästner, M. SOM genesis: Microbial biomass as a significant source. Biodegradation 2012, 111, 41–55. [Google Scholar] [CrossRef]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef]
- Wang, B.; An, S.; Liang, C.; Liu, Y.; Kuzyakov, Y. Microbial necromass as the source of soil organic carbon in global ecosystems. Soil Biol. Biochem. 2021, 162, 108422. [Google Scholar] [CrossRef]
- Prommer, J.; Walker, T.W.N.; Wanek, W.; Braun, J.; Zezula, D.; Hu, Y.; Hofhansl, F.; Richter, A. Increased microbial growth, biomass, and turnover drive soil organic carbon accumulation at higher plant diversity. Glob. Change Biol. 2019, 26, 669–681. [Google Scholar] [CrossRef]
- Kallenbach, C.M.; Frey, S.D.; Grandy, A.S. Direct evidence for microbial-derived soil organic matter formation and its ecophysio-logical controls. Nat. Commun. 2016, 7, 13630. [Google Scholar] [CrossRef]
- Liang, C.; Amelung, W.; Lehmann, J.; Kästner, M. Quantitative assessment of microbial necromass contribution to soil organic matter. Glob. Change Biol. 2019, 25, 3578–3590. [Google Scholar] [CrossRef]
- Whalen, E.D.; Grandy, A.S.; Sokol, N.W.; Keiluweit, M.; Ernakovich, J.; Smith, R.G.; Frey, S.D. Clarifying the evidence for microbial-and plant-derived soil organic matter, and the path toward a more quantitative understanding. Glob. Change Biol. 2022, 28, 7167–7185. [Google Scholar] [CrossRef]
- Buckeridge, K.M.; Mason, K.E.; McNamara, N.P.; Ostle, N.; Puissant, J.; Goodall, T.; Griffiths, R.I.; Stott, A.W.; Whitaker, J. Environmental and microbial controls on microbial necromass recycling, an important precursor for soil carbon stabilization. Commun. Earth Environ. 2020, 1, 36. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [PubMed]
- Finn, D.; Kopittke, P.M.; Dennis, P.G.; Dalal, R.C. Microbial energy and matter transformation in agricultural soils. Soil Biol. Biochem. 2017, 111, 176–192. [Google Scholar] [CrossRef]
- Janus, L.R.; Angeloni, N.L.; McCormack, J.; Rier, S.T.; Tuchman, N.C.; Kelly, J.J. Elevated atmospheric CO2 alters soil microbial communities associated with trembling aspen (Populus tremuloides) roots. Microb. Ecol. 2005, 50, 102–109. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sofi, J.A.; Bhat, A.G.; Kirmai, N.A.; Wani, J.A.; Lone, A.H.; Ganie, M.A.; Dar, G.I.H. Soil quality index as affected by different cropping systems in northwestern Himalayas. Environ. Monit. Assess. 2016, 188, 161. [Google Scholar] [CrossRef] [PubMed]
- Schimel, J.P.; Gulledge, J.M.; Clein-Curley, J.S.; Lindstrom, J.E.; Braddock, J.F. Moisture effects on microbial activity and community structure in decomposing birch litter in the Alaskan taiga. Soil Biol. Biochem. 1999, 31, 831–838. [Google Scholar] [CrossRef]
- Coccina, A.; Cavagnaro, T.R.; Pellegrino, E.; Ercoli, L.; McLaughlin, M.J.; Watts-Williams, S.J. The mycorrhizal pathway of zinc uptake contributes to zinc accumulation in barley and wheat grain. BMC Plant Biol. 2019, 19, 133. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.H.; Kirkegaard, J.A. The agronomic relevance of arbuscular mycorrhizas in the fertility of Australian extensive cropping systems. Agric. Ecosyst. Environ. 2012, 163, 37–53. [Google Scholar] [CrossRef]
- Nyamwange, M.M.; Njeru, E.M.; Mucheru-Muna, M.; Ngetich, F. Soil management practices affect arbuscular mycorrhizal fungi propagules, root colonization and growth of rainfed maize. AIMS Agric. Food 2018, 3, 120–134. [Google Scholar] [CrossRef]
- Li, L.-J.; Zhu-Barker, X.; Ye, R.; Doane, T.A.; Horwath, W.R. Soil microbial biomass size and soil carbon influence the priming effect from carbon inputs depending on nitrogen availability. Soil Biol. Biochem. 2018, 119, 41–49. [Google Scholar] [CrossRef]
- Boyd, E.S.; Peters, J.W. New insights into the evolutionary history of biological nitrogen fixation. Front. Microbiol. 2013, 4, 201. [Google Scholar] [CrossRef]
- Howard, J.B.; Rees, D.C. Structural Basis of Biological Nitrogen Fixation. Chem. Rev. 1996, 96, 2965–2982. [Google Scholar] [CrossRef]
- Jensen, E.S. Role of Grain Legume N₂ Fixation in the Nitrogen Cycling of Temperate Cropping Systems. Ph.D. Thesis, University of Copenhagen, Frederiksberg, Denmark, 1997. [Google Scholar]
- McCallum, M.H.; Peoples, M.B.; Connor, D.J. Contributions of nitrogen by field pea (Pisum sativum L.) in a continuous cropping sequence compared with a lucerne (Medicago sativa L.)-based pasture ley in the Victorian Wimmera. Aust. J. Agric. Res. 2000, 51, 13–22. [Google Scholar] [CrossRef]
- Zheng, M.; Zhou, Z.; Luo, Y.; Zhao, P.; Mo, J. Global pattern and controls of biological nitrogen fixation under nutrient enrichment: A meta-analysis. Glob. Change Biol. 2019, 25, 3018–3030. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, W.; Shehzadi, S.; Shah, S.M.; Shah, Z. Effect of tillage and crop residues management on mung bean (Vigna radiata (L.) Wilczek) crop yield, nitrogen fixation and water use efficiency in rain fed areas. Pak. J. Bot. 2010, 42, 1781–1789. [Google Scholar]
- Hu, H.; Li, H.; Hao, M.; Ren, Y.; Zhang, M.; Liu, R.; Zhang, Y.; Li, G.; Chen, J.; Ning, T.; et al. Nitrogen fixation and crop productivity enhancements co-driven by intercrop root exudates and key rhizosphere bacteria. J. Appl. Ecol. 2021, 58, 2243–2255. [Google Scholar] [CrossRef]
- Smercina, D.N.; Evans, S.E.; Friesen, M.L.; Tiemann, L.K. To fix or not to fix: Controls on free-living nitrogen fixation in the rhizo-sphere. Appl. Environ. Microbiol. 2019, 85, e02546-18. [Google Scholar] [CrossRef] [PubMed]
- Orr, C.H.; James, A.; Leifert, C.; Cooper, J.M.; Cummings, S.P. Diversity and Activity of Free-Living Nitrogen-Fixing Bacteria and Total Bacteria in Organic and Conventionally Managed Soils. Appl. Environ. Microbiol. 2011, 77, 911–919. [Google Scholar] [CrossRef]
- Bergersen, F.J.; Peoples, M.B.; Turner, G.L. A role for poly-β-hydroxybutyrate in bacteroids of soybean root nodules. Proc. R. Soc. B Boil. Sci. 1991, 245, 59–64. [Google Scholar] [CrossRef]
- Vadakattu, G.; Paterson, J. Free-living bacteria lift soil nitrogen supply. Farming Ahead 2006, 169, 40. [Google Scholar]
- Dynarski, K.A.; Houlton, B.Z. Nutrient limitation of terrestrial free-living nitrogen fixation. N. Phytol. 2017, 217, 1050–1061. [Google Scholar] [CrossRef]
- Patel, P.; Panchal, K. Effect of Free-Living Nitrogen Fixing and Phosphate Solubilizing Bacteria on Growth of Gossypium hirsutum L. Asian J. Biol. Life Sci. 2020, 9, 169–176. [Google Scholar] [CrossRef]
- Gupta, V.V.; Roper, M.M. Protection of free-living nitrogen-fixing bacteria within the soil matrix. Soil Tillage Res. 2010, 109, 50–54. [Google Scholar] [CrossRef]
- Kennedy, I.R.; Choudhury, A.T.; Kecskés, M.L. Non-symbiotic bacterial diazotrophs in crop-farming systems: Can their potential for plant growth promotion be better exploited? Soil Biol. Biochem. 2004, 36, 1229–1244. [Google Scholar] [CrossRef]
- Chaudhary, D.; Narula, N.; Sindhu, S.S.; Behl, R.K. Plant growth stimulation of wheat (Triticum aestivum L.) by inoculation of salinity tolerant Azotobacter strains. Physiol. Mol. Biol. Plants 2013, 19, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Köberl, M.; Erlacher, A.; Ramadan, E.M.; El-Arabi, T.F.; Müller, H.; Bragina, A.; Berg, G. Comparisons of diazotrophic communities in native and agricultural desert ecosystems reveal plants as important drivers in diversity. FEMS Microbiol. Ecol. 2015, 92, fiv166. [Google Scholar] [CrossRef] [PubMed]
- Ayangbenro, A.S.; Babalola, O.O. Reclamation of arid and semi-arid soils: The role of plant growth-promoting archaea and bac-teria. Curr. Plant Biol. 2021, 25, 100173. [Google Scholar] [CrossRef]
- Begmatov, S.A.; Berestovskaja, Y.Y.; Vasileva, L.V.; Selitskaya, O.V. Isolation, Screening and Identification of Free-Living Diazotrophic Bacteria from Salinated Arid Soils. Microbiology 2020, 89, 374–377. [Google Scholar] [CrossRef]
- Gupta, V.V.; Zhang, B.; Penton, C.R.; Yu, J.; Tiedje, J.M. Diazotroph diversity and nitrogen fixation in summer active perennial grasses in a Mediterranean region agricultural soil. Front. Mol. Biosci. 2019, 6, 115. [Google Scholar] [CrossRef]
- Six, J.; Elliott, E.T.; Paustian, K. Aggregate and Soil Organic Matter Dynamics under Conventional and No-Tillage Systems. Soil Sci. Soc. Am. J. 1999, 63, 1350–1358. [Google Scholar] [CrossRef]
- Young, I.; Ritz, K. Tillage, habitat space and function of soil microbes. Soil Tillage Res. 2000, 53, 201–213. [Google Scholar] [CrossRef]
- Roper, M.M.; Gupta, V.V.S.R. Enhancing Non-symbiotic N2 Fixation in Agriculture. Open Agric. J. 2016, 10, 7–27. [Google Scholar] [CrossRef]
- Rosenblueth, M.; Ormeño-Orrillo, E.; López-López, A.; Rogel, M.A.; Reyes-Hernandez, B.J.; Martínez-Romero, J.C.; Reddy, P.M.; Martinez-Romero, E. Nitrogen Fixation in Cereals. Front. Microbiol. 2018, 9, 1794. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil Microorganisms Mediating Phosphorus Availability Update on Microbial Phosphorus. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Banik, S.; Dey, B.K. Available phosphate content of an alluvial soil as influenced by inoculation of some isolated phosphate-solubilizing micro-organisms. Plant Soil 1982, 69, 353–364. [Google Scholar] [CrossRef]
- Kucey, R.; Janzen, H.; Leggett, M. Microbially Mediated Increases in Plant-Available Phosphorus. Adv. Agron. 1989, 42, 199–228. [Google Scholar] [CrossRef]
- Gyaneshwar, P.; Kumar, G.N.; Parekh, L.J.; Poole, P.S. Role of soil microorganisms in improving P nutrition of plants. Plant Soil 2002, 245, 83–93. [Google Scholar] [CrossRef]
- Hayman, D.S. Plant growth responses to vesiculararbuscular Mycorrhiza: VI effect of light and temperature. N. Phytol. 1974, 73, 71–80. [Google Scholar] [CrossRef]
- Bolan, N.S. A critical review on the role of mycorrhizal fungi in the uptake of phosphorus by plants. Plant Soil 1991, 134, 189–207. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Rao, N.S. (Ed.) Advances in Agricultural Microbiology; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Goldstein, A.H. Bacterial solubilization of mineral phosphates: Historical perspective and future prospects. Am. J. Altern. Agric. 1986, 1, 51–57. [Google Scholar] [CrossRef]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- De Oliveira Mendes, G.; Moreira de Freitas, A.L.; Liparini Pereira, O.; Ribeiro da Silva, I.; Bojkov Vassilev, N.; Dutra Costa, M. Mechanisms of phosphate solubilization by fungal isolates when exposed to different P sources. Ann. Microbiol. 2014, 64, 239–249. [Google Scholar] [CrossRef]
- Prabhu, N.; Borkar, S.; Garg, S. Phosphate solubilization by microorganisms: Overview, mechanisms, applications and ad-vances. Adv. Biol. Sci. Res. 2019, 161–176. [Google Scholar]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 587. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Naveed, M.; Ali, M.; Zahir, Z.A.; Nadeem, S.M.; Yaseen, M.; Meena, V.S.; Farooq, M.; Singh, R.; Rahman, M.; et al. Perspectives of potassium solubilizing microbes in sustainable food production system: A review. Appl. Soil Ecol. 2018, 133, 146–159. [Google Scholar] [CrossRef]
- Vidyalakshmi, R.; Paranthaman, R.; Bhakyaraj, R. Sulphur Oxidizing Bacteria and Pulse Nutrition—A Review. World J. Agric. Sci. 2009, 5, 270–278. [Google Scholar]
- Tourna, M.; Maclean, P.; Condron, L.; O′Callaghan, M.; Wakelin, S.A. Links between sulphur oxidation and sulphur-oxidising bacteria abundance and diversity in soil microcosms based on soxB functional gene analysis. FEMS Microbiol. Ecol. 2014, 88, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Anandham, R.; Gandhi, P.I.; SenthilKumar, M.; Sridar, R.; Nalayini, P.; Sa, T.-M. Sulfur-oxidizing Bacteria: A Novel Bioinoculant for Sulfur Nutrition and Crop Production. In Bacteria in Agrobiology: Plant Nutrient Management; Springer: Berlin/Heidelberg, Germany, 2011; pp. 81–107. [Google Scholar] [CrossRef]
- Panchal, S.; Chitrakar, R.; Thompson, B.K.; Obulareddy, N.; Roy, D.; Hambright, W.S.; Melotto, M. Regulation of Stomatal Defense by Air Relative Humidity. Plant Physiol. 2016, 172, 2021–2032. [Google Scholar] [CrossRef]
- Huot, B.; Castroverde, C.D.M.; Velásquez, A.C.; Hubbard, E.; Pulman, J.A.; Yao, J.; Childs, K.L.; Tsuda, K.; Montgomery, B.L.; He, S.Y. Dual impact of elevated temperature on plant defence and bacterial virulence in Arabidopsis. Nat. Commun. 2017, 8, 1808–1812. [Google Scholar] [CrossRef]
- Williams, A.; Pétriacq, P.; Schwarzenbacher, R.E.; Beerling, D.J.; Ton, J. Mechanisms of glacial-to-future atmospheric CO 2 effects on plant immunity. N. Phytol. 2018, 218, 752–761. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, M.; Mur, L.A.J.; Shen, Q.; Guo, S. Unravelling the Roles of Nitrogen Nutrition in Plant Disease Defences. Int. J. Mol. Sci. 2020, 21, 572. [Google Scholar] [CrossRef]
- Ruiu, L. Microbial Biopesticides in Agroecosystems. Agronomy 2018, 8, 235. [Google Scholar] [CrossRef]
- Babikova, Z.; Gilbert, L.; Bruce, T.J.A.; Birkett, M.; Caulfield, J.C.; Woodcock, C.; Pickett, J.A.; Johnson, D. Underground signals carried through common mycelial networks warn neighbouring plants of aphid attack. Ecol. Lett. 2013, 16, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Schellhorn, N.A.; Gagic, V.; Bommarco, R. Time will tell: Resource continuity bolsters ecosystem services. Trends Ecol. Evol. 2015, 30, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Birkhofer, K.; Arvidsson, F.; Ehlers, D.; Mader, V.L.; Bengtsson, J.; Smith, H.G. Organic farming affects the biological control of hemipteran pests and yields in spring barley independent of landscape complexity. Landsc. Ecol. 2015, 31, 567–579. [Google Scholar] [CrossRef]
- Dassou, A.G.; Tixier, P. Response of pest control by generalist predators to local-scale plant diversity: A meta-analysis. Ecol. Evol. 2016, 6, 1143–1153. [Google Scholar] [CrossRef]
- Rivers, A.; Barbercheck, M.; Govaerts, B.; Verhulst, N. Conservation agriculture affects arthropod community composition in a rainfed maize–wheat system in central Mexico. Appl. Soil Ecol. 2016, 100, 81–90. [Google Scholar] [CrossRef]
- Favrelière, E.; Ronceux, A.; Pernel, J.; Meynard, J.-M. Nonchemical control of a perennial weed, Cirsium arvense, in arable cropping systems: A review. Agron. Sustain. Dev. 2020, 40, 31. [Google Scholar] [CrossRef]
- Wagner, M.; Mitschunas, N. Fungal effects on seed bank persistence and potential applications in weed biocontrol: A review. Basic Appl. Ecol. 2008, 9, 191–203. [Google Scholar] [CrossRef]
- Harding, D.P.; Raizada, M.N. Controlling weeds with fungi, bacteria and viruses: A review. Front. Plant Sci. 2015, 6, 659. [Google Scholar] [CrossRef]
- Lazzaro, M.; Bàrberi, P.; Dell’Acqua, M.; Pè, M.E.; Limonta, M.; Barabaschi, D.; Cattivelli, L.; Laino, P.; Vaccino, P. Unraveling diversity in wheat competitive ability traits can improve integrated weed management. Agron. Sustain. Dev. 2019, 39, 6. [Google Scholar] [CrossRef]
- Aharon, S.; Peleg, Z.; Argaman, E.; Ben-David, R.; Lati, R. Image-Based High-Throughput Phenotyping of Cereals Early Vigor and Weed-Competitiveness Traits. Remote Sens. 2020, 12, 3877. [Google Scholar] [CrossRef]
- Mwendwa, J.M.; Brown, W.B.; Weidenhamer, J.D.; Weston, P.A.; Quinn, J.C.; Wu, H.; Weston, L.A. Evaluation of Commercial Wheat Cultivars for Canopy Architecture, Early Vigour, Weed Suppression, and Yield. Agronomy 2020, 10, 983. [Google Scholar] [CrossRef]
- Gerhards, R.; Kollenda, B.; Machleb, J.; Möller, K.; Butz, A.; Reiser, D.; Griegentrog, H.-W. Camera-guided Weed Hoeing in Winter Cereals with Narrow Row Distance. Gesunde Pflanz. 2020, 72, 403–411. [Google Scholar] [CrossRef]
- Wang, L.; Ma, B.; Wu, F. Effects of wheat stubble on runoff, infiltration, and erosion of farmland on the Loess Plateau, China, subjected to simulated rainfall. Solid Earth 2017, 8, 281–290. [Google Scholar] [CrossRef]
- Nautiyal, P.; Rajput, R.; Pandey, D.; Arunachalam, K.; Arunachalam, A. Role of glomalin in soil carbon storage and its variation across land uses in temperate Himalayan regime. Biocatal. Agric. Biotechnol. 2019, 21, 101311. [Google Scholar] [CrossRef]
- Villarino, S.H.; Pinto, P.; Jackson, R.B.; Piñeiro, G. Plant rhizodeposition: A key factor for soil organic matter formation in stable fractions. Sci. Adv. 2021, 7, eabd3176. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Jackson, R.D.; DeLucia, E.H.; Tiedje, J.M.; Liang, C. The soil microbial carbon pump: From conceptual insights to em-pirical assessments. Glob. Change Biol. 2020, 26, 6032–6039. [Google Scholar] [CrossRef] [PubMed]
- Gardner, W.K.; Barber, D.A.; Parbery, D.G. The acquisition of phosphorus by Lupinus albus L. Plant Soil 1983, 70, 107–124. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Domanski, G. Carbon input by plants into the soil. Rev. J. Plant Nutr. Soil Sci. 2000, 163, 421–431. [Google Scholar] [CrossRef]
- Sun, H.; Jiang, S.; Jiang, C.; Wu, C.; Gao, M.; Wang, Q. A review of root exudates and rhizosphere microbiome for crop production. Environ. Sci. Pollut. Res. 2021, 28, 54497–54510. [Google Scholar] [CrossRef]
- Neal, A.L.; Ton, J. Systemic defense priming by Pseudomonas putida KT2440 in maize depends on benzoxazinoid exudation from the roots. Plant Signal. Behav. 2013, 8, e22655. [Google Scholar] [CrossRef]
- Lombardi, N.; Vitale, S.; Turrà, D.; Reverberi, M.; Fanelli, C.; Vinale, F.; Marra, R.; Ruocco, M.; Pascale, A.; D’Errico, G.; et al. Root Exudates of Stressed Plants Stimulate and Attract Trichoderma Soil Fungi. Mol. Plant Microbe Interact. 2018, 31, 982–994. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.F. Mycorrhizal Fungi: Highways for Water and Nutrients in Arid Soils. Vadose Zone J. 2007, 6, 291–297. [Google Scholar] [CrossRef]
- Püschel, D.; Bitterlich, M.; Rydlová, J.; Jansa, J. Facilitation of plant water uptake by an arbuscular mycorrhizal fungus: A Gordian knot of roots and hyphae. Mycorrhiza 2020, 30, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Bárzana, G.; Aroca, R.; Paz, J.A.; Chaumont, F.; Martinez-Ballesta, M.C.; Carvajal, M.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal symbiosis increases relative apoplastic water flow in roots of the host plant under both well-watered and drought stress con-ditions. Ann. Bot. 2012, 109, 1009–1017. [Google Scholar] [CrossRef]
- Kaya, C.; Higgs, D.; Kirnak, H.; Tas, I. Mycorrhizal colonisation improves fruit yield and water use efficiency in watermelon (Citrullus lanatus Thunb. ) grown under well-watered and water-stressed conditions. Plant Soil 2003, 253, 287–292. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Aroca, R. Host Response to Osmotic Stresses: Stomatal Behaviour and Water Use Efficiency of Arbuscular Mycorrhizal Plants. In Arbuscular Mycorrhizas: Physiology and Function; Springer: Berlin/Heidelberg, Germany, 2010; pp. 239–256. [Google Scholar] [CrossRef]
- Marulanda, A.; Azcon, R.; Ruiz-Lozano, J.M. Contribution of six arbuscular mycorrhizal fungal isolates to water uptake by Lactuca sativa plants under drought stress. Physiol. Plant. 2003, 119, 526–533. [Google Scholar] [CrossRef]
- Kakouridis, A.; Hagen, J.A.; Kan, M.P.; Mambelli, S.; Feldman, L.J.; Herman, D.J.; Weber, P.K.; Pett-Ridge, J.; Firestone, M.K. Routes to roots: Direct evidence of water transport by arbuscular mycorrhizal fungi to host plants. N. Phytol. 2022, 236, 210–221. [Google Scholar] [CrossRef]
- Lau, J.A.; Lennon, J.T. Rapid responses of soil microorganisms improve plant fitness in novel environments. Proc. Natl. Acad. Sci. USA 2012, 109, 14058–14062. [Google Scholar] [CrossRef]
- Xu, L.; Naylor, D.; Dong, Z.; Simmons, T.; Pierroz, G.; Hixson, K.K.; Kim, Y.-M.; Zink, E.M.; Engbrecht, K.M.; Wang, Y.; et al. Drought delays development of the sorghum root microbiome and enriches for monoderm bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, E4284–E4293. [Google Scholar] [CrossRef]
- De Vries, F.T.; Griffiths, R.I.; Knight, C.G.; Nicolitch, O.; Williams, A. Harnessing rhizosphere microbiomes for drought-resilient crop production. Science 2020, 368, 270–274. [Google Scholar] [CrossRef]
- Naylor, D.; Degraaf, S.; Purdom, E.; Coleman-Derr, D. Drought and host selection influence bacterial community dynamics in the grass root microbiome. ISME J. 2017, 11, 2691–2704. [Google Scholar] [CrossRef] [PubMed]
- Naseem, H.; Bano, A. Role of plant growth-promoting rhizobacteria and their exopolysaccharide in drought tolerance of maize. J. Plant Interact. 2014, 9, 689–701. [Google Scholar] [CrossRef]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F.; Gerbino, R.; et al. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 2014, 17, 316–331. [Google Scholar] [CrossRef]
- Vigani, G.; Rolli, E.; Marasco, R.; Dell′Orto, M.; Michoud, G.; Soussi, A.; Raddadi, N.; Borin, S.; Sorlini, C.; Zocchi, G.; et al. Root bacterial endophytes confer drought resistance and enhance expression and activity of a vacuolar H+-pumping pyro-phosphatase in pepper plants. Environ. Microbiol. 2019, 21, 3212–3228. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, A.; Padmakumar, P.; Nair, I.C.; Radhakrishnan, E.K. Drought tolerant bacterial endophytes with potential plant probiotic effects from Ananas comosus. Biologia 2020, 75, 1769–1778. [Google Scholar] [CrossRef]
- Ortiz, N.; Armada, E.; Duque, E.; Roldán, A.; Azcón, R. Contribution of arbuscular mycorrhizal fungi and/or bacteria to enhancing plant drought tolerance under natural soil conditions: Effectiveness of autochthonous or allochthonous strains. J. Plant Physiol. 2015, 174, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Porcel, R.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal influence on leaf water potential, solute accumulation, and oxidative stress in soybean plants subjected to drought stress. J. Exp. Bot. 2004, 55, 1743–1750. [Google Scholar] [CrossRef] [PubMed]
- Abbaspour, H.; Saeidi-Sar, S.; Afshari, H.; Abdel-Wahhab, M. Tolerance of Mycorrhiza infected Pistachio (Pistacia vera L.) seedling to drought stress under glasshouse conditions. J. Plant Physiol. 2012, 169, 704–709. [Google Scholar] [CrossRef]
- Chareesri, A.; De Deyn, G.B.; Sergeeva, L.; Polthanee, A.; Kuyper, T.W. Increased arbuscular mycorrhizal fungal colonization reduces yield loss of rice (Oryza sativa L.) under drought. Mycorrhiza 2020, 30, 315–328. [Google Scholar] [CrossRef]
- Hu, Y.; Xie, W.; Chen, B. Arbuscular mycorrhiza improved drought tolerance of maize seedlings by altering photosystem II efficiency and the levels of key metabolites. Chem. Biol. Technol. Agric. 2020, 7, 20. [Google Scholar] [CrossRef]
- Darrah, P.R. The rhizosphere and plant nutrition: A quantitative approach. Plant Soil 1993, 155, 3–22. [Google Scholar] [CrossRef]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach; John Wiley & Sons: Hoboken, NJ, USA, 1995. [Google Scholar]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. N. Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Gahoonia, T.S.; Nielsen, N.E. Phosphorus (P) uptake and growth of a root hairless barley mutant (bald root barley, brb) and wild type in low-and high-P. soils. Plant Cell Environ. 2003, 26, 1759–1766. [Google Scholar] [CrossRef]
- Hill, J.O.; Simpson, R.J.; Moore, A.; Chapman, D.F. Morphology and response of roots of pasture species to phosphorus and nitrogen nutrition. Plant Soil 2006, 286, 7–19. [Google Scholar] [CrossRef]
- Lambers, H.; Shane, M.W.; Cramer, M.D.; Pearse, S.J.; Veneklaas, E.J. Root Structure and Functioning for Efficient Acquisition of Phosphorus: Matching Morphological and Physiological Traits. Ann. Bot. 2006, 98, 693–713. [Google Scholar] [CrossRef] [PubMed]
- White, J.F.; Kingsley, K.L.; Verma, S.K.; Kowalski, K.P. Rhizophagy Cycle: An Oxidative Process in Plants for Nutrient Extraction from Symbiotic Microbes. Microorganisms 2018, 6, 95. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.E. Soil microorganisms and phosphorus availability. In Soil Biota: Management in Sustainable Farming Systems; CSIRO Information Services: Melbourne, Australia, 1994; pp. 50–62. [Google Scholar]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The role of soil microorganisms in plant mineral nutrition—Current knowledge and future directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef]
- Abdel-Lateif, K.; Bogusz, D.; Hocher, V. The role of flavonoids in the establishment of plant roots endosymbioses with arbuscular mycorrhiza fungi, rhizobia and Frankia bacteria. Plant Signal. Behav. 2012, 7, 636–641. [Google Scholar] [CrossRef]
- Hassan, S.; Mathesius, U. The role of flavonoids in root–rhizosphere signalling: Opportunities and challenges for improving plant–microbe interactions. J. Exp. Bot. 2012, 63, 3429–3444. [Google Scholar] [CrossRef]
- Fierer, N.; Leff, J.W.; Adams, B.J.; Nielsen, U.N.; Bates, S.T.; Lauber, C.L.; Owens, S.; Gilbert, J.A.; Wall, D.H.; Caporaso, J.G. Cross-biome metagenomic analyses of soil microbial communities and their functional attributes. Proc. Natl. Acad. Sci. USA 2012, 109, 21390–21395. [Google Scholar] [CrossRef]
- Marschener, H. Role of root growth, arbuscular mycorrhiza, and root exudates for the efficiency in nutrient acquisition. Field Crop. Res. 1998, 56, 203–207. [Google Scholar] [CrossRef]
- Degens, B.P. Macro-aggregation of soils by biological bonding and binding mechanisms and the factors affecting these: A review. Soil Res. 1997, 35, 431. [Google Scholar] [CrossRef]
- Li, F.; Xue, C.; Qiu, P.; Liu, Y.; Shi, J.; Shen, B.; Yang, X.; Shen, Q. Soil aggregate size mediates the responses of microbial communities to crop rotation. Eur. J. Soil Biol. 2018, 88, 48–56. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef]
- Tang, J.; Mo, Y.; Zhang, J.; Zhang, R. Influence of biological aggregating agents associated with microbial population on soil aggregate stability. Appl. Soil Ecol. 2011, 47, 153–159. [Google Scholar] [CrossRef]
- Baumert, V.L.; Vasilyeva, N.; Vladimirov, A.A.; Meier, I.C.; Kögel-Knabner, I.; Mueller, C.W. Root Exudates Induce Soil Macroaggregation Facilitated by Fungi in Subsoil. Front. Environ. Sci. 2018, 6, 140. [Google Scholar] [CrossRef]
- Rillig, M.C.; Wright, S.F.; Eviner, V. The role of arbuscular mycorrhizal fungi and glomalin in soil aggregation: Comparing effects of five plant species. Plant Soil 2002, 238, 325–333. [Google Scholar] [CrossRef]
- Egamberdiyeva, D. The effect of plant growth promoting bacteria on growth and nutrient uptake of maize in two different soils. Appl. Soil Ecol. 2007, 36, 184–189. [Google Scholar] [CrossRef]
- Santoyo, G.; Orozco-Mosqueda, M.D.; Govindappa, M. Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: A review. Biocontrol Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Bitas, V.; Kim, H.-S.; Bennett, J.W.; Kang, S. Sniffing on Microbes: Diverse Roles of Microbial Volatile Organic Compounds in Plant Health. Mol. Plant Microbe Interact. 2013, 26, 835–843. [Google Scholar] [CrossRef]
- Knowles, C.J. Microorganisms and cyanide. Bacteriol. Rev. 1976, 40, 652–680. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Uphoff, N. Symbiotic Root-Endophytic Soil Microbes Improve Crop Productivity and Provide Environmental Benefits. Scientifica 2019, 2019, 9106395. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Wang, Y.; Yang, R.; Zheng, J.; Liu, C.; Li, H.; Ma, J.; Zhang, Y.; Wei, C.; Zhang, X. Regulation of Plant Growth, Photosynthesis, Antioxidation and Osmosis by an Arbuscular Mycorrhizal Fungus in Watermelon Seedlings under Well-Watered and Drought Conditions. Front. Plant Sci. 2016, 7, 644. [Google Scholar] [CrossRef]
- Panhwar, Q.A.; Radziah, O.; Zaharah, A.R.; Sariah, M.; Razi, I.M. Role of phosphate solubilizing bacteria on rock phosphate solubility and growth of aerobic rice. J. Environ. Biol. 2011, 32, 607. [Google Scholar] [PubMed]
- Wu, F.; Li, J.; Chen, Y.; Zhang, L.; Zhang, Y.; Wang, S.; Shi, X.; Li, L.; Liang, J. Effects of Phosphate Solubilizing Bacteria on the Growth, Photosynthesis, and Nutrient Uptake of Camellia oleifera Abel. Forests 2019, 10, 348. [Google Scholar] [CrossRef]
- Tsiafouli, M.A.; Thébault, E.; Sgardelis, S.P.; de Ruiter, P.C.; van der Putten, W.H.; Birkhofer, K.; Hemerik, L.; de Vries, F.T.; Bardgett, R.D.; Brady, M.V.; et al. Intensive agriculture reduces soil biodiversity across Europe. Glob. Change Biol. 2014, 21, 973–985. [Google Scholar] [CrossRef]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dy-namics of Gemmatimonadetes in soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef]
- Ashworth, A.; DeBruyn, J.; Allen, F.; Radosevich, M.; Owens, P. Microbial community structure is affected by cropping sequences and poultry litter under long-term no-tillage. Soil Biol. Biochem. 2017, 114, 210–219. [Google Scholar] [CrossRef]
- D’Acunto, L.; Andrade, J.F.; Poggio, S.L.; Semmartin, M. Diversifying crop rotation increased metabolic soil diversity and activity of the microbial community. Agric. Ecosyst. Environ. 2018, 257, 159–164. [Google Scholar] [CrossRef]
- Kim, N.; Zabaloy, M.C.; Guan, K.; Villamil, M.B. Do cover crops benefit soil microbiome? A meta-analysis of current research. Soil Biol. Biochem. 2020, 142, 107701. [Google Scholar] [CrossRef]
- Drost, S.M.; Rutgers, M.; Wouterse, M.; de Boer, W.; Bodelier, P.L. Decomposition of mixtures of cover crop residues increases microbial functional diversity. Geoderma 2019, 361, 114060. [Google Scholar] [CrossRef]
- Dos Santos Cordeiro, C.F.; Echer, F.R.; Araujo, F.F. Cover crops impact crops yields by improving microbiological activity and fertility in sandy soil. J. Soil Sci. Plant Nutr. 2021, 21, 1968–1977. [Google Scholar] [CrossRef]
- Carrera, L.; Buyer, J.; Vinyard, B.; Abdul-Baki, A.; Sikora, L.; Teasdale, J. Effects of cover crops, compost, and manure amendments on soil microbial community structure in tomato production systems. Appl. Soil Ecol. 2007, 37, 247–255. [Google Scholar] [CrossRef]
- Daryanto, S.; Fu, B.; Wang, L.; Jacinthe, P.-A.; Zhao, W. Quantitative synthesis on the ecosystem services of cover crops. Earth Sci. Rev. 2018, 185, 357–373. [Google Scholar] [CrossRef]
- Thapa, V.; Ghimire, R.; Marsalis, M. Cover Crops for Resilience of a Limited-Irrigation Winter Wheat–Sorghum–Fallow Rotation: Soil Carbon, Nitrogen, and Sorghum Yield Responses. Agronomy 2021, 11, 762. [Google Scholar] [CrossRef]
- Wortman, S.E.; Francis, C.A.; Lindquist, J.L. Cover Crop Mixtures for the Western Corn Belt: Opportunities for Increased Productivity and Stability. Agron. J. 2012, 104, 699–705. [Google Scholar] [CrossRef]
- Hagan, J.G.; Vanschoenwinkel, B.; Gamfeldt, L. We should not necessarily expect positive relationships between biodiversity and ecosystem functioning in observational field data. Ecol. Lett. 2021, 24, 2537–2548. [Google Scholar] [CrossRef]
- Finney, D.; Buyer, J.; Kaye, J. Living cover crops have immediate impacts on soil microbial community structure and function. J. Soil Water Conserv. 2017, 72, 361–373. [Google Scholar] [CrossRef]
- Gentsch, N.; Boy, J.; Batalla, J.D.; Heuermann, D.; von Wirén, N.; Schweneker, D.; Feuerstein, U.; Groß, J.; Bauer, B.; Reinhold-Hurek, B.; et al. Catch crop diversity increases rhizosphere carbon input and soil microbial biomass. Biol. Fertil. Soils 2020, 56, 943–957. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, X.; Shen, Y.; Philp, J. Functional diversity of soil microbial communities in response to tillage and crop residue retention in an eroded Loess soil. Soil Sci. Plant Nutr. 2013, 59, 311–321. [Google Scholar] [CrossRef]
- Hoyle, F.; Murphy, D.; Fillery, I. Temperature and stubble management influence microbial CO2–C evolution and gross N transformation rates. Soil Biol. Biochem. 2006, 38, 71–80. [Google Scholar] [CrossRef]
- Govaerts, B.; Mezzalama, M.; Unno, Y.; Sayre, K.D.; Luna-Guido, M.; Vanherck, K.; Dendooven, L.; Deckers, J. Influence of tillage, residue management, and crop rotation on soil microbial biomass and catabolic diversity. Appl. Soil Ecol. 2007, 37, 18–30. [Google Scholar] [CrossRef]
- Nannipieri, P.; Kandeler, E.; Ruggiero, P. Enzyme activities and microbiological and biochemical processes in soil. In Enzymes in the Environment; CRC Press: Boca Raton, FL, USA, 2002; pp. 1–33. [Google Scholar]
- Zhang, P.; Chen, X.; Wei, T.; Yang, Z.; Jia, Z.; Yang, B.; Han, Q.; Ren, X. Effects of straw incorporation on the soil nutrient contents, enzyme activities, and crop yield in a semiarid region of China. Soil Tillage Res. 2016, 160, 65–72. [Google Scholar] [CrossRef]
- Balota, E.L.; Calegari, A.; Nakatani, A.S.; Coyne, M.S. Benefits of winter cover crops and no-tillage for microbial parameters in a Brazilian Oxisol: A long-term study. Agric. Ecosyst. Environ. 2014, 197, 31–40. [Google Scholar] [CrossRef]
- Borase, D.; Nath, C.; Hazra, K.; Senthilkumar, M.; Singh, S.; Praharaj, C.; Singh, U.; Kumar, N. Long-term impact of diversified crop rotations and nutrient management practices on soil microbial functions and soil enzymes activity. Ecol. Indic. 2020, 114, 106322. [Google Scholar] [CrossRef]
- Mendes, I.C.; Bandick, A.K.; Dick, R.P.; Bottomley, P.J. Microbial Biomass and Activities in Soil Aggregates Affected by Winter Cover Crops. Soil Sci. Soc. Am. J. 1999, 63, 873–881. [Google Scholar] [CrossRef]
- Sainju, U.; Whitehead, W.F.; Singh, B.P. Cover crops and nitrogen fertilization effects on soil aggregation and carbon and nitrogen pools. Can. J. Soil Sci. 2003, 83, 155–165. [Google Scholar] [CrossRef]
- Jiang, X.; Wright, A.; Wang, J.; Li, Z. Long-term tillage effects on the distribution patterns of microbial biomass and activities within soil aggregates. Catena 2011, 87, 276–280. [Google Scholar] [CrossRef]
- Nakamoto, T.; Komatsuzaki, M.; Hirata, T.; Araki, H. Effects of tillage and winter cover cropping on microbial substrate-induced respiration and soil aggregation in two Japanese fields. Soil Sci. Plant Nutr. 2012, 58, 70–82. [Google Scholar] [CrossRef]
- Morugán-Coronado, A.; Pérez-Rodríguez, P.; Insolia, E.; Soto-Gómez, D.; Fernández-Calviño, D.; Zornoza, R. The impact of crop diversification, tillage and fertilization type on soil total microbial, fungal and bacterial abundance: A worldwide meta-analysis of agricultural sites. Agric. Ecosyst. Environ. 2022, 329, 107867. [Google Scholar] [CrossRef]
- Andrade, D.S.; Colozzi-Filho, A.; Giller, K. The soil microbial community and soil tillage. In Soil Tillage in Agroecosystems; CRC Press: Boca Raton, FL, USA, 2003; pp. 51–81. [Google Scholar] [CrossRef]
- Vázquez, E.; Benito, M.; Espejo, R.; Teutscherova, N. No-tillage and liming increase the root mycorrhizal colonization, plant biomass and N content of a mixed oat and vetch crop. Soil Tillage Res. 2020, 200, 104623. [Google Scholar] [CrossRef]
- Doran, J.W. Microbial biomass and mineralizable nitrogen distributions in no-tillage and plowed soils. Biol. Fertil. Soils 1987, 5, 68–75. [Google Scholar] [CrossRef]
- Beare, M.H.; Parmelee, R.W.; Hendrix, P.F.; Cheng, W.; Coleman, D.C.; Crossley, D.A., Jr. Microbial and faunal interactions and effects on litter nitrogen and decomposition in agroecosystems. Ecol. Monogr. 1992, 62, 569–591. [Google Scholar] [CrossRef]
- Gupta, V.V.; Roper, M.M.; Kirkegaard, J.A.; Angus, J.F. Changes in microbial biomass and organic matter levels during the first year of modified tillage and stubble management practices on a red earth. Soil Res. 1994, 32, 1339–1354. [Google Scholar] [CrossRef]
- Bell, J.M.; Smith, J.L.; Bailey, V.L.; Bolton, H. Priming effect and C storage in semi-arid no-till spring crop rotations. Biol. Fertil. Soils 2003, 37, 237–244. [Google Scholar] [CrossRef]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and Fungal Contributions to Carbon Sequestration in Agroecosystems. Soil Sci. Soc. Am. J. 2006, 70, 555–569. [Google Scholar] [CrossRef]
- Bailey, V.L.; Smith, J.L.; Bolton, H., Jr. Fungal-to-bacterial ratios in soils investigated for enhanced C sequestration. Soil Biol. Biochem. 2002, 34, 997–1007. [Google Scholar] [CrossRef]
- Pankhurst, C.; Kirkby, C.; Hawke, B.; Harch, B. Impact of a change in tillage and crop residue management practice on soil chemical and microbiological properties in a cereal-producing red duplex soil in NSW, Australia. Biol. Fertil. Soils 2002, 35, 189–196. [Google Scholar]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef]
- Abbott, L.K.; Macdonald, L.M.; Wong, M.T.; Webb, M.J.; Jenkins, S.N.; Farrell, M. Potential roles of biological amendments for profitable grain production—A review. Agric. Ecosyst. Environ. 2018, 256, 34–50. [Google Scholar] [CrossRef]
- Anon. 2021. Available online: https://www.meticulousresearch.com/product/biostimulants-market-5057 (accessed on 7 July 2022).
- Laurent, E.-A.; Ahmed, N.; Durieu, C.; Grieu, P.; Lamaze, T. Marine and fungal biostimulants improve grain yield, nitrogen absorption and allocation in durum wheat plants. J. Agric. Sci. 2020, 158, 279–287. [Google Scholar] [CrossRef]
- Parađiković, N.; Vinković, T.; Vinković Vrček, I.; Žuntar, I.; Bojić, M.; Medić-Šarić, M. Effect of natural biostimulants on yield and nutritional quality: An example of sweet yellow pepper (Capsicum annuum L.) plants. J. Sci. Food Agric. 2011, 91, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- Kocira, S. Effect of amino acid biostimulant on the yield and nutraceutical potential of soybean. Chil. J. Agric. Res. 2019, 79, 17–25. [Google Scholar] [CrossRef]
- Popko, M.; Michalak, I.; Wilk, R.; Gramza, M.; Chojnacka, K.; Górecki, H. Effect of the New Plant Growth Biostimulants Based on Amino Acids on Yield and Grain Quality of Winter Wheat. Molecules 2018, 23, 470. [Google Scholar] [CrossRef]
- Wang, S.; Tian, X.; Liu, Q. The Effectiveness of Foliar Applications of Zinc and Biostimulants to Increase Zinc Concentration and Bioavailability of Wheat Grain. Agronomy 2020, 10, 178. [Google Scholar] [CrossRef]
- Powlson, D.; Cai, Z.; Lemanceau, P. Soil Carbon Dynamics and Nutrient Cycling. Soil Carbon Sci. Manag. Policy Mult. Benefits 2014, 71, 98. [Google Scholar]
- Devi, Y.B.; Meetei, T.T.; Kumari, N. Impact of Pesticides on Soil Microbial Diversity and Enzymes: A Review. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 952–958. [Google Scholar] [CrossRef]
- Karas, P.; Baguelin, C.; Pertile, G.; Papadopoulou, E.; Nikolaki, S.; Storck, V.; Ferrari, F.; Trevisan, M.; Ferrarini, A.; Fornasier, F.; et al. Assessment of the impact of three pesticides on microbial dynamics and functions in a lab-to-field experimental approach. Sci. Total. Environ. 2018, 637–638, 636–646. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F. Indirect Effect of Pesticides on Insects and Other Arthropods. Toxics 2021, 9, 177. [Google Scholar] [CrossRef]
- Kuthubutheen, A.; Pugh, G. The effects of fungicides on soil fungal populations. Soil Biol. Biochem. 1979, 11, 297–303. [Google Scholar] [CrossRef]
- Kosel, K.; Collins, H. Foliar Fungicides Reduce Short Term Non-Target Soil Microbial Activity and Community Structure. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Wang, X.; Lu, Z.; Miller, H.; Liu, J.; Hou, Z.; Liang, S.; Zhao, X.; Zhang, H.; Borch, T. Fungicide azoxystrobin induced changes on the soil microbiome. Appl. Soil Ecol. 2019, 145, 103343. [Google Scholar] [CrossRef]
- Zhang, C.; Zhou, T.; Zhu, L.; Juhasz, A.; Du, Z.; Li, B.; Wang, J.; Wang, J. Response of soil microbes after direct contact with py-raclostrobin in fluvo-aquic soil. Environ. Pollut. 2019, 255, 113164. [Google Scholar] [CrossRef]
- Katsoula, A.; Vasileiadis, S.; Sapountzi, M.; Karpouzas, D.G. The response of soil and phyllosphere microbial communities to repeated application of the fungicide iprodione: Accelerated biodegradation or toxicity? FEMS Microbiol. Ecol. 2020, 96, fiaa056. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yin, Y.; Wang, S.; Yu, Y. Accumulation of chlorothalonil and its metabolite, 4-hydroxychlorothalonil, in soil after repeated applications and its effects on soil microbial activities under greenhouse conditions. Environ. Sci. Pollut. Res. 2013, 21, 3452–3459. [Google Scholar] [CrossRef]
- Schreiner, R.P.; Bethlenfalvay, G.J. Mycorrhizae, biocides, and biocontrol 3. Effects of three different fungicides on developmental stages of three AM fungi. Biol. Fertil. Soils 1997, 24, 18–26. [Google Scholar] [CrossRef]
- Channabasava, A.; Lakshman, H.; Jorquera, M. Effect of fungicides on association of arbuscular mycorrhiza fungus Rhizophagus fasciculatus and growth of Proso millet (Panicum miliaceum L.). J. Soil Sci. Plant Nutr. 2015, 15, 35–45. [Google Scholar] [CrossRef]
- Ding, H.; Zheng, X.; Zhang, J.; Zhang, Y.; Yu, J.; Chen, D. Influence of chlorothalonil and carbendazim fungicides on the transformation processes of urea nitrogen and related microbial populations in soil. Environ. Sci. Pollut. Res. 2019, 26, 31133–31141. [Google Scholar] [CrossRef]
- Wilson, G.; Williamson, M. Topsin-M: The new benomyl for mycorrhizal-suppression experiments. Mycologia 2008, 100, 548–554. [Google Scholar] [CrossRef]
- Madhaiyan, M.; Poonguzhali, S.; Hari, K.; Saravanan, V.; Sa, T. Influence of pesticides on the growth rate and plant-growth promoting traits of Gluconacetobacter diazotrophicus. Pestic. Biochem. Physiol. 2006, 84, 143–154. [Google Scholar] [CrossRef]
- Sarnaik, S.S.; Kanekar, P.P.; Raut, V.M.; Taware, S.P.; Chavan, K.S.; Bhadbhade, B.J. Effect of application of different pesticides to soybean on the soil microflora. J. Environ. Biol. 2006, 27, 423–426. [Google Scholar]
- Tang, T.; Chen, G.; Liu, F.; Bu, C.; Liu, L.; Zhao, X. Effects of transgenic glufosinate-tolerant rapeseed (Brassica napus L.) and the associated herbicide application on rhizospheric bacterial communities. Physiol. Mol. Plant Pathol. 2019, 106, 246–252. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Rizvi, P.Q. Biotoxic Effects of Herbicides on Growth, Nodulation, Nitrogenase Activity, and Seed Production in Chickpeas. Commun. Soil Sci. Plant Anal. 2006, 37, 1783–1793. [Google Scholar] [CrossRef]
- Treseder, K.K.; Allen, M.F. Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi: A model and field test. N. Phytol. 2002, 155, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Zhang, J.-B.; Zhang, L.; Yang, M.; He, J.-Z. Long-term fertilization regimes affect bacterial community structure and diversity of an agricultural soil in northern China. J. Soils Sediments 2008, 8, 43–50. [Google Scholar] [CrossRef]
- Verbruggen, E.; Heijden, M.G.A.; Rillig, M.C.; Kiers, E.T. Mycorrhizal fungal establishment in agricultural soils: Factors determining inoculation success. N. Phytol. 2012, 197, 1104–1109. [Google Scholar] [CrossRef]
- Albizua, A.; Williams, A.; Hedlund, K.; Pascual, U. Crop rotations including ley and manure can promote ecosystem services in conventional farming systems. Appl. Soil Ecol. 2015, 95, 54–61. [Google Scholar] [CrossRef]
- Tian, H.; Drijber, R.; Zhang, J.; Li, X. Impact of long-term nitrogen fertilization and rotation with soybean on the diversity and phosphorus metabolism of indigenous arbuscular mycorrhizal fungi within the roots of maize (Zea mays L.). Agric. Ecosyst. Environ. 2013, 164, 53–61. [Google Scholar] [CrossRef]
- Williams, A.; Börjesson, G.; Hedlund, K. The effects of 55 years of different inorganic fertiliser regimes on soil properties and microbial community composition. Soil Biol. Biochem. 2013, 67, 41–46. [Google Scholar] [CrossRef]
- Wuest, S.B.; Caesar-TonThat, T.; Wright, S.F.; Williams, J.D. Organic matter addition, N, and residue burning effects on infiltration, biological, and physical properties of an intensively tilled silt-loam soil. Soil Tillage Res. 2005, 84, 154–167. [Google Scholar] [CrossRef]
- Feldman, E.V. Impact of Nitrogen Fertilization on Arbuscular Mycorrhizal Fungi Abundance in Association with Panicum Virgatum. Inq. J. 2015, 7. Available online: http://www.inquiriesjournal.com/a?id=1184 (accessed on 30 November 2022).
- Olsson, P.A.; Baath, E.; Jakobsen, I. Phosphorus effects on the mycelium and storage structures of an arbuscular mycorrhizal fungus as studied in the soil and roots by analysis of Fatty Acid signatures. Appl. Environ. Microbiol. 1997, 63, 3531–3538. [Google Scholar] [CrossRef] [PubMed]
- Nagahashi, G.; Douds, D.D. Partial separation of root exudate components and their effects upon the growth of germinated spores of AM fungi. Mycol. Res. 2000, 104, 1453–1464. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Schwenke, G.D.; Van Zwieten, L. Impact of agricultural inputs on soil organisms—a review. Soil Res. 2006, 44, 379–406. [Google Scholar] [CrossRef]
- Cheng, Y.; Ishimoto, K.; Kuriyama, Y.; Osaki, M.; Ezawa, T. Ninety-year-, but not single, application of phosphorus fertilizer has a major impact on arbuscular mycorrhizal fungal communities. Plant Soil 2012, 365, 397–407. [Google Scholar] [CrossRef]
- Williams, A.; Manoharan, L.; Rosenstock, N.P.; Olsson, P.A.; Hedlund, K. Long-term agricultural fertilization alters arbuscular mycorrhizal fungal community composition and barley (Hordeum vulgare) mycorrhizal carbon and phosphorus exchange. N. Phytol. 2017, 213, 874–885. [Google Scholar] [CrossRef]
- Wang, G.M.; Stribley, D.P.; Tinker, P.B.; Walker, C. Effects of pH on arbuscular mycorrhiza I. Field observations on the long-term liming experiments at Rothamsted and Woburn. N. Phytol. 1993, 124, 465–472. [Google Scholar] [CrossRef]
- Rousk, J.; Brookes, P.C.; Bååth, E. The microbial PLFA composition as affected by pH in an arable soil. Soil Biol. Biochem. 2010, 42, 516–520. [Google Scholar] [CrossRef]
- Klironomos, J.N.; Hart, M.M.; Gurney, J.E.; Moutoglis, P. Interspecific differences in the tolerance of arbuscular mycorrhizal fungi to freezing and drying. Can. J. Bot. 2001, 79, 1161–1166. [Google Scholar]
- Gai, J.P.; Cai, X.B.; Feng, G.; Christie, P.; Li, X.L. Arbuscular mycorrhizal fungi associated with sedges on the Tibetan plateau. Mycorrhiza 2006, 16, 151–157. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Kobae, Y.; Ohmori, Y.; Saito, C.; Yano, K.; Ohtomo, R.; Fujiwara, T. Phosphate treatment strongly inhibits new arbuscule de-velopment but not the maintenance of arbuscule in mycorrhizal rice roots. Plant Physiol. 2016, 171, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Corkidi, L.; Rowland, D.L.; Johnson, N.C.; Allen, E.B. Nitrogen fertilization alters the functioning of arbuscular mycorrhizas at two semiarid grasslands. Plant Soil 2002, 240, 299–310. [Google Scholar] [CrossRef]
- Lin, X.; Feng, Y.; Zhang, H.; Chen, R.; Wang, J.; Zhang, J.; Chu, H. Long-term balanced fertilization decreases arbuscular my-corrhizal fungal diversity in an arable soil in North China revealed by 454 pyrosequencing. Environ. Sci. Technol. 2012, 46, 5764–5771. [Google Scholar] [CrossRef]
- Weese, D.J.; Heath, K.D.; Dentinger, B.T.; Lau, J.A. Long-term nitrogen addition causes the evolution of less-cooperative mutu-alists. Evolution 2015, 69, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; McGrath, S.P.; Hirsch, P.R.; Clark, I.M.; Storkey, J.; Wu, L.; Zhou, J.; Liang, Y. Plant–microbe networks in soil are weakened by century-long use of inorganic fertilizers. Microb. Biotechnol. 2019, 12, 1464–1475. [Google Scholar] [CrossRef] [PubMed]
- Riah, W.; Laval, K.; Laroche-Ajzenberg, E.; Mougin, C.; Latour, X.; Trinsoutrot-Gattin, I. Effects of pesticides on soil enzymes: A review. Environ. Chem. Lett. 2014, 12, 257–273. [Google Scholar] [CrossRef]
- Atlas, R.; Pramer, D.; Bartha, R. Assessment of pesticide effects on non-target soil microorganisms. Soil Biol. Biochem. 1978, 10, 231–239. [Google Scholar] [CrossRef]
- Kinney, C.A.; Mandernack, K.W.; Mosier, A.R. Laboratory investigations into the effects of the pesticides mancozeb, chlorothalonil, and prosulfuron on nitrous oxide and nitric oxide production in fertilized soil. Soil Biol. Biochem. 2005, 37, 837–850. [Google Scholar] [CrossRef]
- Edlinger, A.; Garland, G.; Hartman, K.; Banerjee, S.; Degrune, F.; García-Palacios, P.; Hallin, S.; Valzano-Held, A.; Herzog, C.; Jansa, J.; et al. Agricultural management and pesticide use reduce the functioning of beneficial plant symbionts. Nat. Ecol. Evol. 2022, 6, 1145–1154. [Google Scholar] [CrossRef]
- Chumley, H.; Hewlings, S. The effects of elevated atmospheric carbon dioxide [CO2] on micronutrient concentration, specifically iron (Fe) and zinc (Zn) in rice—A systematic review. J. Plant Nutr. 2020, 43, 1571–1578. [Google Scholar] [CrossRef]
- Marler, J.B.; Wallin, J.R. Human Health, the Nutritional Quality of Harvested Food and Sustainable Farming Systems; Nutrition Security Institute: Bellevue, WA, USA, 2006. [Google Scholar]
- Marles, R.J. Mineral nutrient composition of vegetables, fruits and grains: The context of reports of apparent historical declines. J. Food Compos. Anal. 2017, 56, 93–103. [Google Scholar] [CrossRef]
- Mayer, A.-M.B.; Trenchard, L.; Rayns, F. Historical changes in the mineral content of fruit and vegetables in the UK from 1940 to 2019: A concern for human nutrition and agriculture. Int. J. Food Sci. Nutr. 2021, 73, 315–326. [Google Scholar] [CrossRef]
- Cunningham, J.; Milligan, G.; Trevisan, L. Minerals in Australian Fruits and Vegetables; Food Standards Australia New Zealand: Canberra, Australia, 2001. Available online: http://www.foodstandards.gov.au/publications/documents/minerals_report.pdf (accessed on 21 April 2022).
- Fan, M.-S.; Zhao, F.-J.; Fairweather-Tait, S.J.; Poulton, P.R.; Dunham, S.J.; McGrath, S.P. Evidence of decreasing mineral density in wheat grain over the last 160 years. J. Trace Elem. Med. Biol. 2008, 22, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.-S.; Zhao, F.-J.; Poulton, P.R.; McGrath, S. Historical changes in the concentrations of selenium in soil and wheat grain from the Broadbalk experiment over the last 160 years. Sci. Total. Environ. 2008, 389, 532–538. [Google Scholar] [CrossRef]
- McIntyre, B.; Bouldin, D.; Urey, G.; Kizito, F. Modeling cropping strategies to improve human nutrition in Uganda. Agric. Syst. 2001, 67, 105–120. [Google Scholar] [CrossRef]
- Welch, R.M.; Graham, R.D. Agriculture: The real nexus for enhancing bioavailable micronutrients in food crops. J. Trace Elem. Med. Biol. 2005, 18, 299–307. [Google Scholar] [CrossRef]
- Gregory, P.J.; Wahbi, A.; Adu-Gyamfi, J.; Heiling, M.; Gruber, R.; Joy, E.J.; Broadley, M.R. Approaches to reduce zinc and iron deficits in food systems. Glob. Food Secur. 2017, 15, 1–10. [Google Scholar] [CrossRef]
- McConkey, B.G.; Curtin, D.; A Campbell, C.; A Brandt, S.; Selles, F. Crop and soil nitrogen status of tilled and no-tillage systems in semiarid regions of Saskatchewan. Can. J. Soil Sci. 2002, 82, 489–498. [Google Scholar] [CrossRef]
- Mustafa, M.A.; Mabhaudhi, T.; Massawe, F. Building a resilient and sustainable food system in a changing world—A case for climate-smart and nutrient dense crops. Glob. Food Secur. 2020, 28, 100477. [Google Scholar] [CrossRef]
- Delgado, J.A.; Groffman, P.M.; Nearing, M.A.; Goddard, T.; Reicosky, D.; Lal, R.; Kitchen, N.R.; Rice, C.W.; Towery, D.; Salon, P. Conservation practices to mitigate and adapt to climate change. J. Soil Water Conserv. 2011, 66, 118A–129A. [Google Scholar] [CrossRef]
- Albrecht, A.; Kandji, S.T. Carbon sequestration in tropical agroforestry systems. Agric. Ecosyst. Environ. 2003, 99, 15–27. [Google Scholar] [CrossRef]
- Tiefenbacher, A.; Sandén, T.; Haslmayr, H.-P.; Miloczki, J.; Wenzel, W.; Spiegel, H. Optimizing Carbon Sequestration in Croplands: A Synthesis. Agronomy 2021, 11, 882. [Google Scholar] [CrossRef]
- Panchasara, H.; Samrat, N.; Islam, N. Greenhouse Gas Emissions Trends and Mitigation Measures in Australian Agriculture Sector—A Review. Agriculture 2021, 11, 85. [Google Scholar] [CrossRef]
- Olander, L.; Wollenberg, E.; Tubiello, F.; Herold, M. Advancing agricultural greenhouse gas quantification. Environ. Res. Lett. 2013, 8, 011002. [Google Scholar] [CrossRef]
- Dickie, A.; Streck, C.; Roe, S. Strategies for Mitigating Climate Change in Agriculture: Recommendations for Philanthropy—Executive Summary. Climate Focus and California Environmental Associates, Prepared with the Support of the Climate and Land Use Alliance. 2014. Available online: www.agriculturalmitigation.org (accessed on 20 October 2022).
- Paustian, K.; Collier, S.; Baldock, J.; Burgess, R.; Creque, J.; DeLonge, M.; Dungait, J.; Ellert, B.; Frank, S.; Goddard, T.; et al. Quantifying carbon for agricultural soil management: From the current status toward a global soil information system. Carbon Manag. 2019, 10, 567–587. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Urbanski, L.; Hobley, E.; Lang, B.; von Lützow, M.; Marin-Spiotta, E.; van Wesemael, B.; Rabot, E.; Ließ, M.; Garcia-Franco, N.; et al. Soil organic carbon storage as a key function of soils—A review of drivers and indicators at various scales. Geoderma 2018, 333, 149–162. [Google Scholar] [CrossRef]
- Smith, P.; Soussana, J.-F.; Angers, D.; Schipper, L.; Chenu, C.; Rasse, D.P.; Batjes, N.H.; van Egmond, F.; McNeill, S.; Kuhnert, M.; et al. How to measure, report and verify soil carbon change to realize the potential of soil carbon sequestration for atmospheric greenhouse gas removal. Glob. Change Biol. 2020, 26, 219–241. [Google Scholar] [CrossRef]
- Kenne, G.J.; Kloot, R.W. The Carbon Sequestration Potential of Regenerative Farming Practices in South Carolina, USA. Am. J. Clim. Change 2019, 8, 157–172. [Google Scholar] [CrossRef]
- Jordon, M.W.; Smith, P.; Long, P.R.; Bürkner, P.-C.; Petrokofsky, G.; Willis, K.J. Can Regenerative Agriculture increase national soil carbon stocks? Simulated country-scale adoption of reduced tillage, cover cropping, and ley-arable integration using RothC. Sci. Total. Environ. 2022, 825, 153955. [Google Scholar] [CrossRef]
- Bennett, A. A Review of the Economics of Regenerative Agriculture in Western Australia. Available online: https://library.dpird.wa.gov.au/cgi/viewcontent.cgi?article=1171&context=pubns (accessed on 15 February 2022).
- Hayward, C.T. The Financial Implications of Regenerative Agriculture in the Southern Cape and the Subsequent Impact on Future Animal and Winter Cereal Crop Production. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, March 2021. [Google Scholar]
- Brown, K.; Schirmer, J.; Upton, P. Can regenerative agriculture support successful adaptation to climate change and im-proved landscape health through building farmer self-efficacy and wellbeing? Curr. Res. Environ. Sustain. 2022, 4, 100170. [Google Scholar] [CrossRef]
- Schulte, L.A.; Dale, B.E.; Bozzetto, S.; Liebman, M.; Souza, G.M.; Haddad, N.; Richard, T.L.; Basso, B.; Brown, R.C.; Hilbert, J.A.; et al. Meeting global challenges with regenerative agriculture producing food and energy. Nat. Sustain. 2021, 5, 384–388. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khangura, R.; Ferris, D.; Wagg, C.; Bowyer, J. Regenerative Agriculture—A Literature Review on the Practices and Mechanisms Used to Improve Soil Health. Sustainability 2023, 15, 2338. https://doi.org/10.3390/su15032338
Khangura R, Ferris D, Wagg C, Bowyer J. Regenerative Agriculture—A Literature Review on the Practices and Mechanisms Used to Improve Soil Health. Sustainability. 2023; 15(3):2338. https://doi.org/10.3390/su15032338
Chicago/Turabian StyleKhangura, Ravjit, David Ferris, Cameron Wagg, and Jamie Bowyer. 2023. "Regenerative Agriculture—A Literature Review on the Practices and Mechanisms Used to Improve Soil Health" Sustainability 15, no. 3: 2338. https://doi.org/10.3390/su15032338
APA StyleKhangura, R., Ferris, D., Wagg, C., & Bowyer, J. (2023). Regenerative Agriculture—A Literature Review on the Practices and Mechanisms Used to Improve Soil Health. Sustainability, 15(3), 2338. https://doi.org/10.3390/su15032338