
Hyperspectral Imagery Detects Water Deficit and Salinity Effects on Photosynthesis and Antioxidant Enzyme Activity of Three Greek Olive Varieties

 , and
, and 
Abstract
1. Introduction
- Evaluating stress tolerance of three olive varieties with major importance based on photosynthetic rate, water content and enzyme activity alterations in response to water deficit and salinity;
- Estimating the relationship between the results of VIs, PCR, PLSR and LDA and photosynthesis and enzymes of stressed olive plants, aiming at preparing the development of drone-based field scanning for early detection of crop stresses;
- Identifying substantial wavelength ranges with correlation to enzyme activity and create customized vegetation indices for each of the studied antioxidative enzymes to enable high-throughput plant phenotyping.
2. Materials and Methods
2.1. Study Area and Dataset Description
2.2. Spectral Data Acquisition
2.3. Photosynthetic Measurements
2.4. Water Content Measurements
2.5. Determination of Enzyme Activity
2.6. Spectral Data Preprocessing
2.7. Statistical Analysis
3. Results
3.1. Photosynthesis
3.2. Water Content
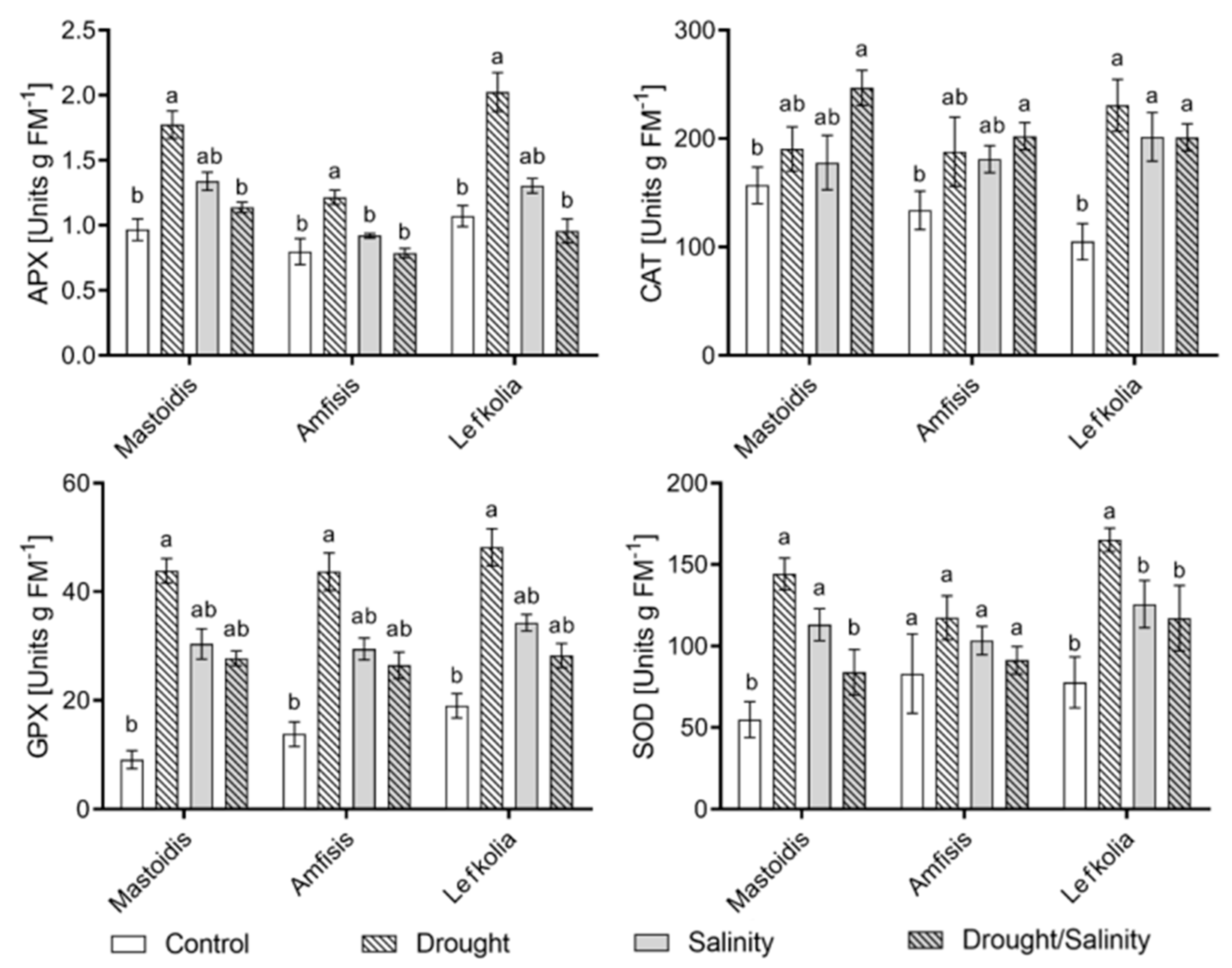
3.3. Enzyme Activity
3.4. Spectral Vegetation Indices
3.5. Correlation between Vegetation Indices and Enzyme Activities
3.6. PCR and PLSR Analyses of Hyperspectral Data and Photosynthesis
3.7. Linear Discriminant Analysis of Hyperspectral Datasets
3.8. Correlation between Single Wavelengths and Enzyme Activity Results
4. Discussion
4.1. Water Content and Photosynthesis
4.2. Enzyme Response to Stress
4.3. Relationship between Plant Health Parameters and Spectral Reflectance
4.4. Methodological Considerations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koubouris, G. Advances in prediction, monitoring and mitigation of climate change effects on water resources and good agricultural practices for crop adaptation to environmental stresses. J. Water Clim. Chang. 2018, 9, 631–632. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, J.; An, P.; Ren, W.; Pan, Z.; Dong, Z.; Han, G.; Pan, Y.; Pan, S.; Tian, H. Enhancing soil drought induced by climate change and agricultural practices: Observational and experimental evidence from the semiarid area of northern China. Agric. For. Meteorol. 2017, 243, 74–83. [Google Scholar] [CrossRef]
- St.Clair, S.B.; Lynch, J.P. The opening of Pandora’s Box: Climate change impacts on soil fertility and crop nutrition in developing countries. Plant Soil 2010, 335, 101–115. [Google Scholar] [CrossRef]
- Tanasijevic, L.; Todorovic, M.; Pereira, L.S.; Pizzigalli, C.; Lionello, P. Impacts of climate change on olive crop evapotranspiration and irrigation requirements in the Mediterranean region. Agric. Water Manag. 2014, 144, 54–68. [Google Scholar] [CrossRef]
- Cortignani, R.; Dell’Unto, D.; Dono, G. Paths of adaptation to climate change in major Italian agricultural areas: Effectiveness and limits in supporting the profitability of farms. Agric. Water Manag. 2021, 244, 106433. [Google Scholar] [CrossRef]
- Corwin, D.L. Climate change impacts on soil salinity in agricultural areas. Eur. J. Soil Sci. 2021, 72, 842–862. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef]
- Kourgialas, N.N.; Koubouris, G.C.; Dokou, Z. Optimal irrigation planning for addressing current or future water scarcity in Mediterranean tree crops. Sci. Total Environ. 2019, 654, 616–632. [Google Scholar] [CrossRef]
- Boshkovski, B.; Tzerakis, C.; Doupis, G.; Zapolska, A.; Kalaitzidis, C.; Koubouris, G. Relationship between physiological and biochemical measurements with spectral reflectance for two Phaseolus vulgaris L. genotypes under multiple stress. Int. J. Remote Sens. 2021, 42, 1230–1249. [Google Scholar] [CrossRef]
- Trabelsi, L.; Gargouri, K.; Ben Hassena, A.; Mbadra, C.; Ghrab, M.; Ncube, B.; Van Staden, J.; Gargouri, R. Impact of drought and salinity on olive water status and physiological performance in an arid climate. Agric. Water Manag. 2019, 213, 749–759. [Google Scholar] [CrossRef]
- Calderón, R.; Navas-Cortés, J.A.; Zarco-Tejada, P.J. Early Detection and Quantification of Verticillium Wilt in Olive Using Hyperspectral and Thermal Imagery over Large Areas. Remote Sens. 2015, 7, 5584–5610. [Google Scholar] [CrossRef]
- Fraga, H.; Moriondo, M.; Leolini, L.; Santos, J.A. Mediterranean Olive Orchards under Climate Change: A Review of Future Impacts and Adaptation Strategies. Agronomy 2020, 11, 56. [Google Scholar] [CrossRef]
- Ponti, L.; Gutierrez, A.P.; Ruti, P.M.; Dell’Aquila, A. Fine-scale ecological and economic assessment of climate change on olive in the Mediterranean Basin reveals winners and losers. Proc. Natl. Acad. Sci. USA 2014, 111, 5598–5603. [Google Scholar] [CrossRef] [PubMed]
- Aragüés, R.; Puy, J.; Royo, A.; Espada, J.L. Three-year field response of young olive trees (Olea europaea L., cv. Arbequina) to soil salinity: Trunk growth and leaf ion accumulation. Plant Soil 2005, 271, 265–273. [Google Scholar] [CrossRef]
- Chartzoulakis, K. Salinity and olive: Growth, salt tolerance, photosynthesis and yield. Agric. Water Manag. 2005, 78, 108–121. [Google Scholar] [CrossRef]
- Centritto, M.; Brilli, F.; Fodale, R.; Loreto, F. Different sensitivity of isoprene emission, respiration and photosynthesis to high growth temperature coupled with drought stress in black poplar (Populus nigra) saplings. Tree Physiol. 2011, 31, 275–286. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T. Diffusive and Metabolic Limitations to Photosynthesis under Drought and Salinity in C 3 Plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef]
- Pan, Y.; Wu, L.J.; Yu, Z.L. Effect of salt and drought stress on antioxidant enzymes activities and SOD isoenzymes of liquorice (Glycyrrhiza uralensis Fisch). Plant Growth Regul. 2006, 49, 157–165. [Google Scholar] [CrossRef]
- Vemanna, R.S.; Babitha, K.; Solanki, J.K.; Reddy, V.A.; Sarangi, S.; Udayakumar, M. Aldo-keto reductase-1 (AKR1) protect cellular enzymes from salt stress by detoxifying reactive cytotoxic compounds. Plant Physiol. Biochem. 2017, 113, 177–186. [Google Scholar] [CrossRef]
- Zaidi, I.; Ebel, C.; Belgaroui, N.; Ghorbel, M.; Amara, I.; Hanin, M. The wheat MAP kinase phosphatase 1 alleviates salt stress and increases antioxidant activities in Arabidopsis. J. Plant Physiol. 2016, 193, 12–21. [Google Scholar] [CrossRef]
- Koubouris, G.; Kavroulakis, N.; Metzidakis, I.T.; Vasilakakis, M.; Sofo, A. Ultraviolet-B radiation or heat cause changes in photosynthesis, antioxidant enzyme activities and pollen performance in olive tree. Photosynthetica 2015, 53, 279–287. [Google Scholar] [CrossRef]
- Tanveer, M.; Shabala, S. Targeting redox regulatory mechanisms for salinity stress tolerance in crops. In Salinity Responses and Tolerance in Plants, 1st ed.; Kumar, V., Wani, S.H., Suprasanna, P., Tran, L.-S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2018; Volume 1, pp. 213–234. [Google Scholar] [CrossRef]
- Yan, H.; Li, Q.; Park, S.-C.; Wang, X.; Liu, Y.-J.; Zhang, Y.-G.; Tang, W.; Kou, M.; Ma, D.-F. Overexpression of CuZnSOD and APX enhance salt stress tolerance in sweet potato. Plant Physiol. Biochem. 2016, 109, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Pessarakli, M. Handbook of Plant and Crop Stress, Fourth Edition. In Handbook of Plant and Crop Stress, Fourth Edition; CRC Press: Boca Raton, FL, USA, 2019; Volume 20102370, ISBN 978-1-4398-1396-6. [Google Scholar]
- Brinkhoff, J.; Schultz, A.; Suarez, L.A.; Robson, A.J. Olive Tree Water Stress Detection Using Daily Multispectral Imagery. In Proceedings of the 2021 IEEE International Geoscience and Remote Sensing Symposium IGARSS, Brussels, Belgium, 11–16 July 2021; pp. 5826–5829. [Google Scholar] [CrossRef]
- Boshkovski, B.; Tzerakis, C.; Doupis, G.; Zapolska, A.; Kalaitzidis, C.; Koubouris, G. Relationships of Spectral Reflectance with Plant Tissue Mineral Elements of Common Bean (Phaseolus Vulgaris L.) Under Drought and Salinity Stresses. Commun. Soil Sci. Plant Anal. 2020, 51, 675–686. [Google Scholar] [CrossRef]
- Sun, P.; Wahbi, S.; Tsonev, T.; Haworth, M.; Liu, S.; Centritto, M. On the Use of Leaf Spectral Indices to Assess Water Status and Photosynthetic Limitations in Olea europaea L. during Water-Stress and Recovery. PLoS ONE 2014, 9, e105165. [Google Scholar] [CrossRef] [PubMed]
- Hernández, E.I.; Melendez-Pastor, I.; Navarro-Pedreño, J.; Gómez, I. Spectral indices for the detection of salinity effects in melon plants. Sci. Agric. 2014, 71, 324–330. [Google Scholar] [CrossRef]
- Gao, Y.; Li, D. Detecting salinity stress in tall fescue based on single leaf spectrum. Sci. Hortic. 2012, 138, 159–164. [Google Scholar] [CrossRef]
- Main, R.; Cho, M.A.; Mathieu, R.; O’Kennedy, M.M.; Ramoelo, A.; Koch, S. An investigation into robust spectral indices for leaf chlorophyll estimation. ISPRS J. Photogramm. Remote Sens. 2011, 66, 751–761. [Google Scholar] [CrossRef]
- Marino, G.; Pallozzi, E.; Cocozza, C.; Tognetti, R.; Giovannelli, A.; Cantini, C.; Centritto, M. Assessing gas exchange, sap flow and water relations using tree canopy spectral reflectance indices in irrigated and rainfed Olea europaea L. Environ. Exp. Bot. 2014, 99, 43–52. [Google Scholar] [CrossRef]
- Abdulridha, J.; Ampatzidis, Y.; Ehsani, R.; de Castro, A.I. Evaluating the performance of spectral features and multivariate analysis tools to detect laurel wilt disease and nutritional deficiency in avocado. Comput. Electron. Agric. 2018, 155, 203–211. [Google Scholar] [CrossRef]
- Yu, K.; Anderegg, J.; Mikaberidze, A.; Karisto, P.; Mascher, F.; McDonald, B.; Walter, A.; Hund, A. Hyperspectral Canopy Sensing of Wheat Septoria Tritici Blotch Disease. Front. Plant Sci. 2018, 9, 1195. [Google Scholar] [CrossRef]
- Mirzaie, M.; Darvishzadeh, R.; Shakiba, A.; Matkan, A.; Atzberger, C.; Skidmore, A. Comparative analysis of different uni- and multi-variate methods for estimation of vegetation water content using hyper-spectral measurements. Int. J. Appl. Earth Obs. Geoinf. 2014, 26, 1–11. [Google Scholar] [CrossRef]
- Li, M.; Chu, R.; Yu, Q.; Islam, A.R.M.T.; Chou, S.; Shen, S. Evaluating Structural, Chlorophyll-Based and Photochemical Indices to Detect Summer Maize Responses to Continuous Water Stress. Water 2018, 10, 500. [Google Scholar] [CrossRef]
- Lu, B.; Dao, P.; Liu, J.; He, Y.; Shang, J. Recent Advances of Hyperspectral Imaging Technology and Applications in Agriculture. Remote Sens. 2020, 12, 2659. [Google Scholar] [CrossRef]
- Gomes, L.; Nobre, T.; Sousa, A.; Rei, F.; Guiomar, N. Hyperspectral Reflectance as a Basis to Discriminate Olive Varieties—A Tool for Sustainable Crop Management. Sustainability 2020, 12, 3059. [Google Scholar] [CrossRef]
- Smigaj, M.; Gaulton, R.; Suárez, J.C.; Barr, S.L. Canopy temperature from an Unmanned Aerial Vehicle as an indicator of tree stress associated with red band needle blight severity. For. Ecol. Manag. 2019, 433, 699–708. [Google Scholar] [CrossRef]
- Atzberger, C.; Jarmer, T.; Schlerf, M.; Werner, W.; Atzberger, C.; Jarmer, T.; Schlerf, M.; Kötz, B.; Werner, W. Spectroradio-metric determination of wheat bio-physical variables. Comparison of different empirical-statistical approaches. In Proceedings of the 23rd, EARSeL Symposium and General Assembly: Remote Sensing in Transition, Ghent, Belgium, 2–5 June 2003; Millpress: Rotterdam, The Netherlands, 2003. [Google Scholar]
- Zapolska, A.; Kalaitzidis, C.; Markakis, E.; Ligoxigakis, E.; Koubouris, G. Linear Discriminant Analysis of spectral measurements for discrimination between healthy and diseased trees of Olea europaea L. artificially infected by Fomitiporia mediterranea. Int. J. Remote Sens. 2020, 41, 5388–5398. [Google Scholar] [CrossRef]
- Santos-Rufo, A.; Mesas-Carrascosa, F.-J.; García-Ferrer, A.; Meroño-Larriva, J. Wavelength Selection Method Based on Partial Least Square from Hyperspectral Unmanned Aerial Vehicle Orthomosaic of Irrigated Olive Orchards. Remote Sens. 2020, 12, 3426. [Google Scholar] [CrossRef]
- Rubio-Delgado, J.; Pérez, C.J.; Vega-Rodríguez, M.A. Predicting leaf nitrogen content in olive trees using hyperspectral data for precision agriculture. Precis. Agric. 2021, 22, 1–21. [Google Scholar] [CrossRef]
- Hueni, A.; Bialek, A. Cause, Effect, and Correction of Field Spectroradiometer Interchannel Radiometric Steps. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2017, 10, 1542–1551. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Bao, Y.; Luo, J.; Jin, X.; Xu, X.; Song, X.; Yang, G. Exploring the Best Hyperspectral Features for LAI Estimation Using Partial Least Squares Regression. Remote Sens. 2014, 6, 6221–6241. [Google Scholar] [CrossRef]
- Moula, I.; Boussadia, O.; Koubouris, G.; Ben Hassine, M.; Boussetta, W.; Van Labeke, M.C.; Braham, M. Ecophysiological and biochemical aspects of olive tree (Olea europaea L.) in response to salt stress and gibberellic acid-induced alleviation. South Afr. J. Bot. 2020, 132, 38–44. [Google Scholar] [CrossRef]
- Angelopoulos, K.; Dichio, B.; Xiloyannis, C. Inhibition of photosynthesis in olive trees (Olea europaea L.) during water stress and rewatering. J. Exp. Bot. 1996, 47, 1093–1100. [Google Scholar] [CrossRef]
- Koubouris, G.C.; Tzortzakis, N.; Kourgialas, N.N.; Darioti, M.; Metzidakis, I. Growth, photosynthesis and pollen performance in saline water treated olive plants under high temperature. Int. J. Plant Biol. 2015, 6, 28–32. [Google Scholar] [CrossRef]
- Mousavi, S.; Regni, L.; Bocchini, M.; Mariotti, R.; Cultrera, N.; Mancuso, S.; Googlani, J.; Chakerolhosseini, M.R.; Guerrero, C.; Albertini, E.; et al. Physiological, epigenetic and genetic regulation in some olive cultivars under salt stress. Sci. Rep. 2019, 9, 1093. [Google Scholar] [CrossRef]
- Proietti, P.; Nasini, L.; Del Buono, D.; D’Amato, R.; Tedeschini, E.; Businelli, D. Selenium protects olive (Olea europaea L.) from drought stress. Sci. Hortic. 2013, 164, 165–171. [Google Scholar] [CrossRef]
- Ben Abdallah, M.; Methenni, K.; Nouairi, I.; Zarrouk, M.; Ben Youssef, N. Drought priming improves subsequent more severe drought in a drought-sensitive cultivar of olive cv. Chétoui. Sci. Hortic. 2017, 221, 43–52. [Google Scholar] [CrossRef]
- Asada, K. THE WATER-WATER CYCLE IN CHLOROPLASTS: Scavenging of Active Oxygens and Dissipation of Excess Photons. Annu. Rev. Plant Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Aganchich, B.; Wahbi, S.; Loreto, F.; Centritto, M. Partial root zone drying: Regulation of photosynthetic limitations and antioxidant enzymatic activities in young olive (Olea europaea) saplings. Tree Physiol. 2009, 29, 685–696. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Santos, A.D.L.; Moutinho-Pereira, A.J.M.; Gonçalves, A.B.C.; Lopes, J.I.; Ferreira, T.C.; Ae, E.A.B.; Santos, D.L.; José, A.E.; Moutinho-Pereira, M.; et al. Physiological behaviour, oxidative damage and antioxidative protection of olive trees grown under different irrigation regimes. Plant Soil 2007, 292, 1–12. [Google Scholar] [CrossRef]
- Guerfel, M.; Ouni, Y.; Boujnah, D.; Zarrouk, M. Photosynthesis parameters and activities of enzymes of oxidative stress in two young ’Chemlali’ and ’Chetoui’ olive trees under water deficit. Photosynth. 2009, 47, 340–346. [Google Scholar] [CrossRef]
- Yildirim, T.; Zhou, Y.; Flynn, K.C.; Gowda, P.H.; Ma, S.; Moriasi, D.N. Evaluating the sensitivity of vegetation and water indices to monitor drought for three Mediterranean crops. Agron. J. 2021, 113, 123–134. [Google Scholar] [CrossRef]
- Serrano, L.; González-Flor, C.; Gorchs, G. Assessing vineyard water status using the reflectance based Water Index. Agric. Ecosyst. Environ. 2010, 139, 490–499. [Google Scholar] [CrossRef]
- Rallo, G.; Minacapilli, M.; Ciraolo, G.; Provenzano, G. Detecting crop water status in mature olive groves using vegetation spectral measurements. Biosyst. Eng. 2014, 128, 52–68. [Google Scholar] [CrossRef]
- Garbulsky, M.F.; Peñuelas, J.; Gamon, J.; Inoue, Y.; Filella, I. The photochemical reflectance index (PRI) and the remote sensing of leaf, canopy and ecosystem radiation use efficiencies: A review and meta-analysis. Remote Sens. Environ. 2011, 115, 281–297. [Google Scholar] [CrossRef]
- Gamon, J.A.; Kovalchuck, O.; Wong, C.Y.S.; Harris, A.; Garrity, S.R. Monitoring seasonal and diurnal changes in photosynthetic pigments with automated PRI and NDVI sensors. Biogeosciences 2015, 12, 4149–4159. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X.; Zhou, G. Automatic Detection of Rice Disease Using Near Infrared Spectra Technologies. J. Indian Soc. Remote Sens. 2017, 45, 785–794. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NDVI | WI | PRI | APX | GPX | CAT | SOD | LeafWC | ShootWC | |
|---|---|---|---|---|---|---|---|---|---|
| NDVI | 1 | 0.550 ** | 0.474 ** | −0.033 | −0.377 * | −0.264 | −0.259 | 0.194 | 0.165 |
| WI | 0.550 ** | 1 | 0.493 ** | −0.313 | −0.519 ** | −0.173 | −0.503 ** | −0.226 | 0.132 |
| PRI | 0.474 ** | 0.493 ** | 1 | −0.200 | −0.169 | −0.013 | −0.211 | −0.251 | 0.063 |
| APX | −0.033 | −0.313 | −0.200 | 1 | 0.638 ** | 0.353 * | 0.492 ** | 0.193 | −0.092 |
| GPX | −0.377 * | −0.519 ** | −0.169 | 0.638 ** | 1 | 0.441 * | 0.712 ** | 0.091 | −0.033 |
| CAT | −0.264 | −0.173 | −0.013 | 0.353 * | 0.441 * | 1 | 0.333 | −0.307 | −0.411 * |
| SOD | −0.259 | −0.503 ** | −0.211 | 0.492 ** | 0.712 ** | 0.333 | 1 | 0.195 | −0.179 |
| LeafWC | 0.194 | −0.226 | −0.251 | 0.193 | 0.091 | −0.307 | 0.195 | 1 | 0.170 |
| ShootWC | 0.165 | 0.132 | 0.063 | −0.092 | −0.033 | −0.411 * | −0.179 | 0.170 | 1 |
| All | Mastoidis | Amfisis | Lefkolia Serron | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NDVI | WI | PRI | NDVI | WI | PRI | NDVI | WI | PRI | NDVI | WI | PRI | |
| 22 March 2016 | 0.868 ** | −0.115 | 0.498 ** | 0.878 ** | 0.165 | 0.242 | 0.900 ** | 0.251 | 0.220 | 0.839 ** | −0.495 | 0.687 ** |
| 27 April 2016 | 0.853 ** | 0.034 | 0.522 ** | 0.840 ** | 0.055 | 0.419 ** | 0.918 ** | 0.087 | 0.578 | 0.866 ** | 0.485 | 0.552 |
| 23 May 2016 | 0.805 ** | 0.113 | 0.351 * | 0.907 ** | 0.074 | 0.168 | 0.777 ** | 0.238 | 0.546 * | 0.961 ** | −0.385 | 0.344 |
| 10 June 2016 | 0.855 ** | 0.404 ** | 0.390 ** | 0.857 ** | 0.491 | 0.613 * | 0.764 ** | 0.381 | 0.448 | 0.936 ** | 0.423 | 0.327 |
| 22 June 2016 | 0.832 ** | 0.425 * | 0.283 | 0.920 ** | 0.569 | 0.154 | 0.680 * | 0.167 | 0.166 | 0.940 ** | 0.627 * | 0.473 |
| Measurement Date | Mastoidis | Amfisis | Lefkolia Serron | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PCR | PLSR | PCR | PLSR | PCR | PLSR | |||||||
| r2 | RMSE | r2 | RMSE | r2 | RMSE | r2 | RMSE | r2 | RMSE | r2 | RMSE | |
| All dates | 0.502 | 2.991 | 0.658 | 2.468 | 0.634 | 3.146 | 0.653 | 3.074 | 0.522 | 2.867 | 0.562 | 2.793 |
| 22 March 2016 | 0.686 | 1.867 | 0.672 | 1.909 | 0.705 | 2.001 | 0.695 | 2.027 | 0.656 | 3.028 | 0.596 | 3.283 |
| 27 April 2016 | 0.556 | 1.953 | 0.514 | 2.044 | 0.632 | 1.491 | 0.604 | 1.545 | 0.540 | 2.561 | 0.513 | 2.635 |
| 23 May 2016 | 0.911 | 0.971 | 0.914 | 0.954 | 0.393 | 4.720 | 0.331 | 4.953 | 0.734 | 1.314 | 0.834 | 1.037 |
| 10 June 2016 | 0.719 | 2.308 | 0.673 | 2.489 | 0.533 | 2.741 | 0.563 | 2.651 | 0.857 | 1.622 | 0.883 | 1.471 |
| 22 June 2016 | 0.841 | 0.684 | 0.834 | 0.699 | 0.288 | 2.264 | 0.938 | 0.608 | 0.812 | 1.163 | 0.872 | 0.960 |
| PCR | PLSR | |||
|---|---|---|---|---|
| r2 | RMSE | r2 | RMSE | |
| Ascorbate peroxidase (APX) | 0.058 | 0.427 | NA | - |
| Guaiacol peroxidase (GPX) | 0.194 | 11.617 | 0.184 | 11.489 |
| Catalase (CAT) | 0.023 | 52.959 | 0.029 | 53.586 |
| Superoxide dismutase (SOD) | 0.048 | 37.214 | 0.090 | 37.101 |
| Leaf water content | 0.192 | 8.718 | NA | - |
| Shoot water content | 0.009 | 10.326 | 0.020 | 10.322 |
| 22 March 2016 | 27 April 2016 | 23 May 2016 | 10 June 2016 | 22 June 2016 | |
|---|---|---|---|---|---|
| Drought | 64.58 | 79.41 | 77.78 | 84.44 | 87.88 |
| Salinity | 70.83 | 76.47 | 73.33 | 80.00 | 87.88 |
| Index | Correlation | |
|---|---|---|
| APX | (1350 − 730)/(1350 + 730) | 0.340 |
| GPX | (1380 − 770)/(1380 + 770) | 0.527 ** |
| CAT | (640 − 780)/(640 + 780) | 0.343 |
| SOD | (1870 − 1380)/(1870 + 1380) | 0.413 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boshkovski, B.; Doupis, G.; Zapolska, A.; Kalaitzidis, C.; Koubouris, G. Hyperspectral Imagery Detects Water Deficit and Salinity Effects on Photosynthesis and Antioxidant Enzyme Activity of Three Greek Olive Varieties. Sustainability 2022, 14, 1432. https://doi.org/10.3390/su14031432
Boshkovski B, Doupis G, Zapolska A, Kalaitzidis C, Koubouris G. Hyperspectral Imagery Detects Water Deficit and Salinity Effects on Photosynthesis and Antioxidant Enzyme Activity of Three Greek Olive Varieties. Sustainability. 2022; 14(3):1432. https://doi.org/10.3390/su14031432
Chicago/Turabian StyleBoshkovski, Blagoja, Georgios Doupis, Anhelina Zapolska, Chariton Kalaitzidis, and Georgios Koubouris. 2022. "Hyperspectral Imagery Detects Water Deficit and Salinity Effects on Photosynthesis and Antioxidant Enzyme Activity of Three Greek Olive Varieties" Sustainability 14, no. 3: 1432. https://doi.org/10.3390/su14031432
APA StyleBoshkovski, B., Doupis, G., Zapolska, A., Kalaitzidis, C., & Koubouris, G. (2022). Hyperspectral Imagery Detects Water Deficit and Salinity Effects on Photosynthesis and Antioxidant Enzyme Activity of Three Greek Olive Varieties. Sustainability, 14(3), 1432. https://doi.org/10.3390/su14031432