
Responses of Carbon Isotope Composition of Common C3 and C4 Plants to Climatic Factors in Temperate Grasslands



Abstract
:1. Introduction
2. Materials and Methods
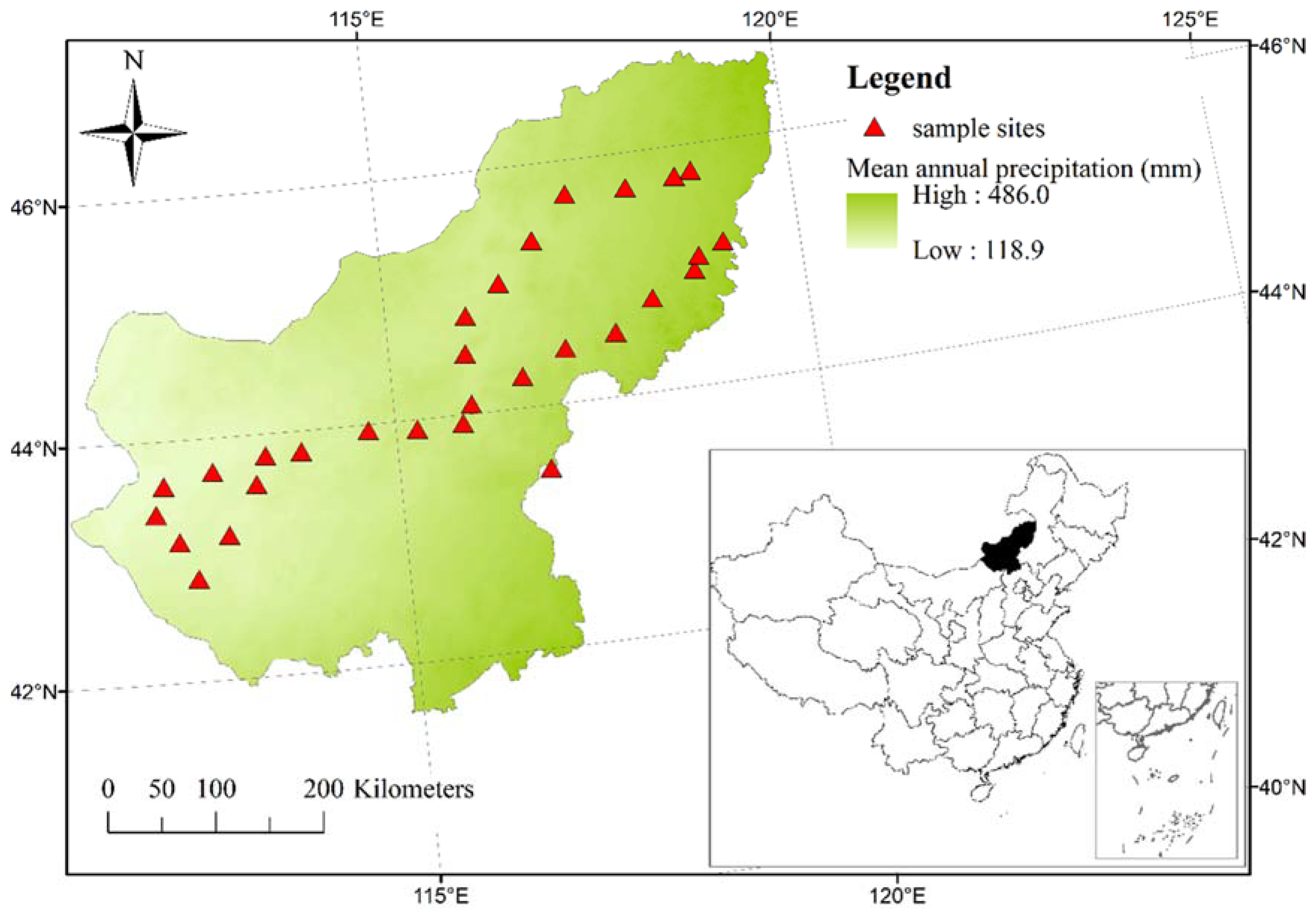
2.1. Study Area
2.2. Sampling
2.3. Sample Analysis
2.4. Meteorological Data
2.5. Data Analysis
3. Results
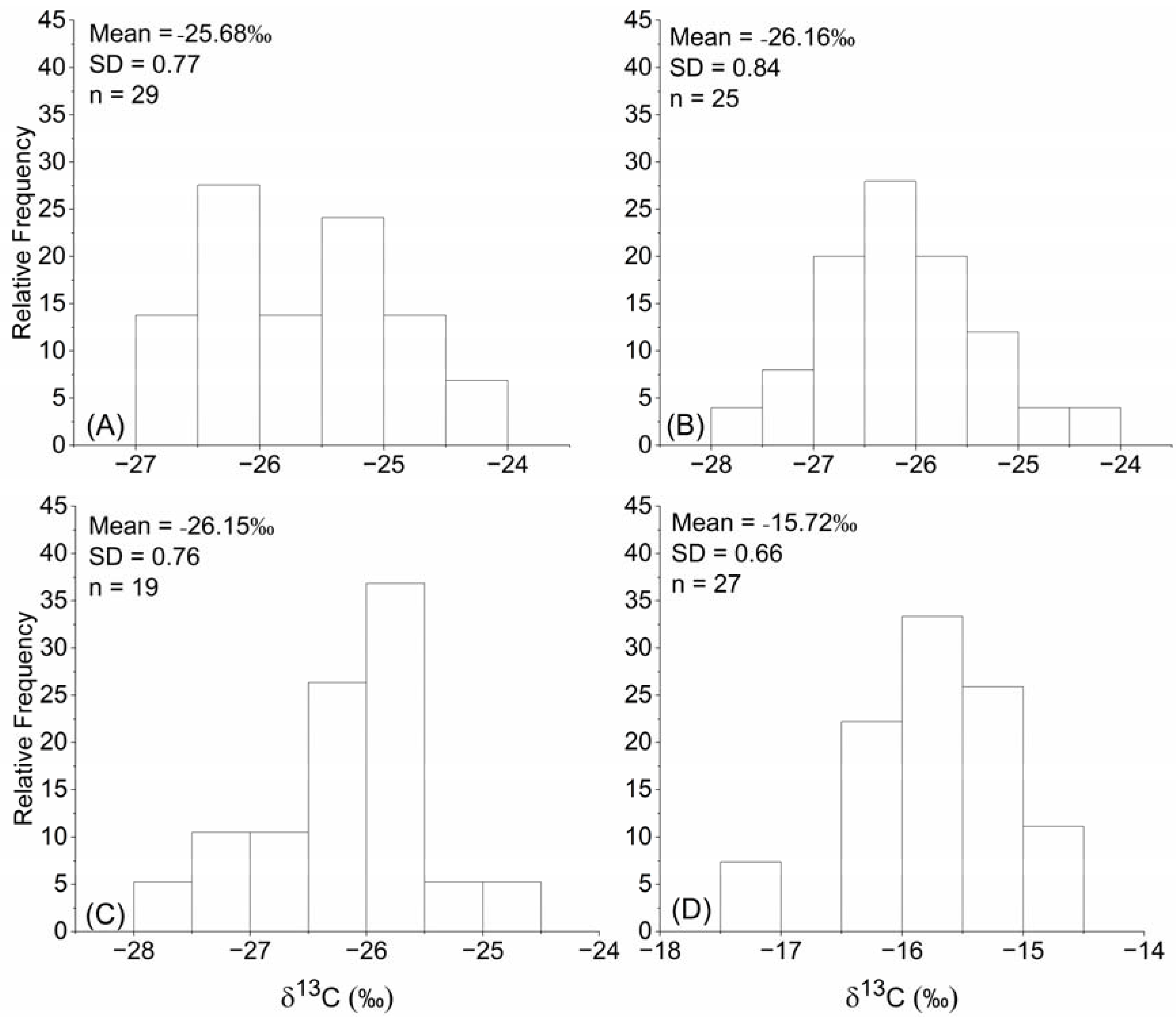
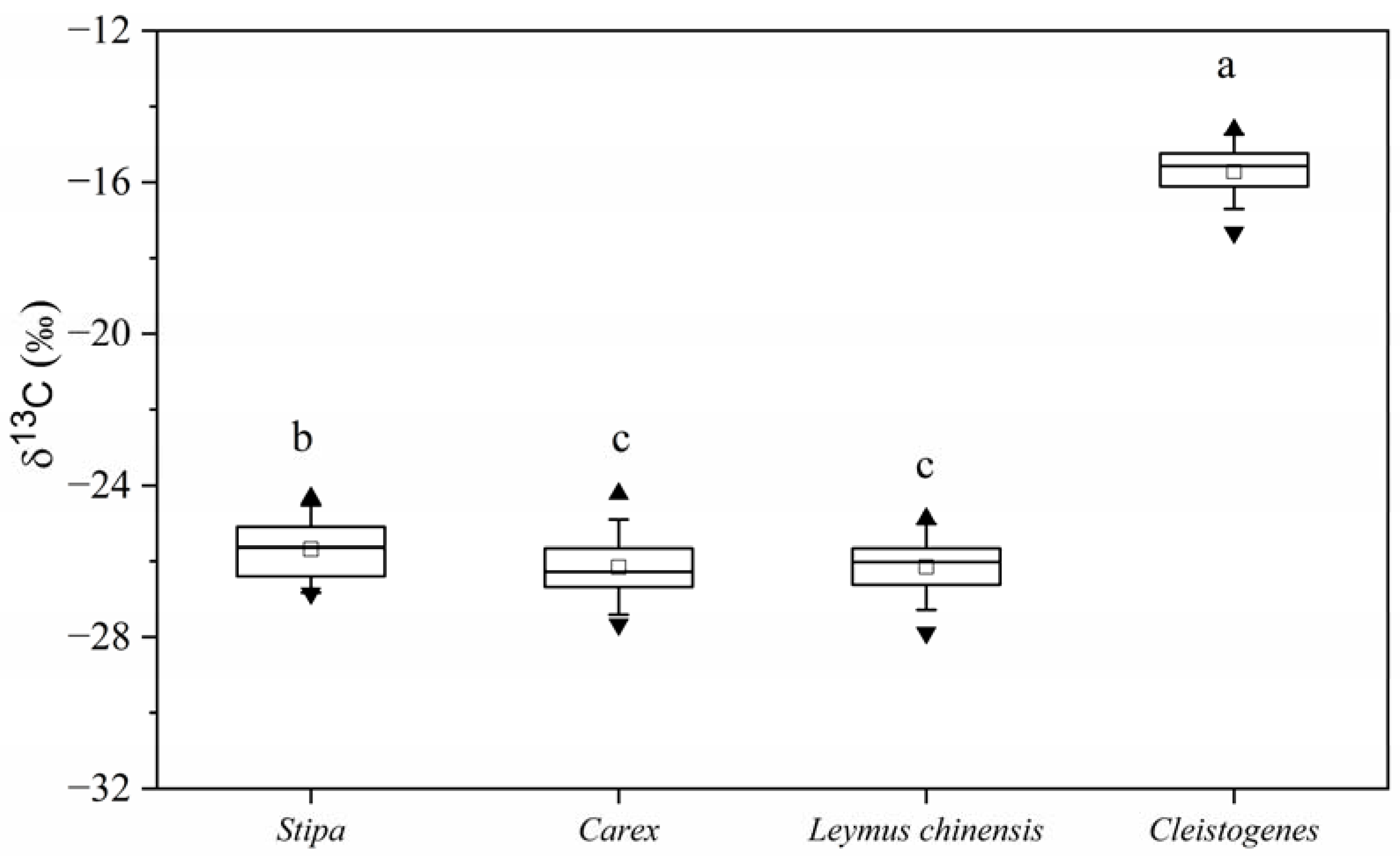
3.1. Plant δ13C of Common C3 and C4 Species in Temperate Grasslands
3.2. Relationships between Plant δ13C and Climatic Factors
3.3. Integrative Effects of Climatic Factors on Plant δ13C
4. Discussion
4.1. Characteristics of the Plant δ13C of C3 and C4 Species in Temperate Grasslands
4.2. The Relationships between Plant δ13C and Temperature
4.3. The Relationships between Plant δ13C and Precipitation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spangenberg, J.E.; Schweizer, M.; Zufferey, V. Carbon and nitrogen stable isotope variations in leaves of two grapevine cultivars (Chasselas and Pinot noir): Implications for ecophysiological studies. Plant Physiol. Biochem. 2021, 163, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.E.; Mambelli, S.; Plamboeck, A.H.; Templer, P.H.; Tu, K.P. Stable isotopes in plant ecology. Annu. Rev. Ecol. Syst. 2002, 33, 507–559. [Google Scholar] [CrossRef]
- Song, W.; Zhou, Y. Linking leaf δ15N and δ13C with soil fungal biodiversity, ectomycorrhizal and plant pathogenic abundance in forest ecosystems of China. Catena 2021, 200, 105176. [Google Scholar] [CrossRef]
- Song, W.; Liu, Y. Survival strategy of the endangered tree Acer catalpifolium Rehd., based on 13C fractionation. Ecol. Evol. 2020, 10, 8532–8537. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, W.; Cheng, X.; Harris, W.; Schaeffer, S.M.; Xu, X.; Zhao, B. Factors affecting 13C enrichment of vegetation and soil in temperate grasslands in Inner Mongolia, China. J. Soils Sediments 2019, 19, 2190–2199. [Google Scholar] [CrossRef]
- Yan, G.; Han, S.; Zhou, M.; Sun, W.; Huang, B.; Wang, H.; Xing, Y.; Wang, Q. Variations in the natural 13C and 15N abundance of plants and soils under long-term N addition and precipitation reduction: Interpretation of C and N dynamics. For. Ecosyst. 2020, 7, 49. [Google Scholar] [CrossRef]
- Murphy, B.P.; Bowman, D.M.J.S. The carbon and nitrogen isotope composition of Australian grasses in relation to climate. Funct. Ecol. 2009, 23, 1040–1049. [Google Scholar] [CrossRef]
- Zhou, Y.; Fan, J.; Zhang, W.; Harris, W.; Zhong, H.; Hu, Z.; Song, L. Factors influencing altitudinal patterns of C3 plant foliar carbon isotope composition of grasslands on the Qinghai-Tibet Plateau, China. Alp. Bot. 2011, 121, 79–90. [Google Scholar] [CrossRef]
- Ma, F.; Liang, W.; Zhou, Z.; Xiao, G.; Liu, J.; He, J.; Jiao, B.; Xu, T. Spatial variation in leaf stable carbon isotope composition of three Caragana species in Northern China. Forests 2018, 9, 297. [Google Scholar] [CrossRef] [Green Version]
- Loader, N.; Hemming, D. Spatial variation in pollen δ13C correlates with temperature and seasonal development timing. Holocene 2001, 11, 587–592. [Google Scholar] [CrossRef]
- Zheng, S.; Shangguan, Z. Spatial patterns of foliar stable carbon isotope compositions of C3 plant species in the Loess Plateau of China. Ecol. Res. 2007, 22, 342–353. [Google Scholar] [CrossRef]
- Morecroft, M.D.; Woodward, F.I.; Marrs, R.H. Altitudinal trends in leaf nutrient contents, leaf size and δ13C of Alchemilla alpina. Funct. Ecol. 1992, 6, 730–740. [Google Scholar] [CrossRef]
- Liu, W.-G.; Feng, X.-H.; Ning, Y.-F.; Zhang, Q.-L.; Cao, Y.-N.; An, Z.-S. δ13C variation of C3 and C4 plants across an Asian monsoon rainfall gradient in arid northwestern China. Glob. Chang. Biol. 2005, 11, 1094–1100. [Google Scholar] [CrossRef]
- Buchmann, N.; Brooks, J.R.; Rapp, K.D.; Ehleringer, J.R. Carbon isotope composition of C4 grasses is influenced by light and water supply. Plant Cell Environ. 1996, 19, 392–402. [Google Scholar] [CrossRef]
- Wang, G.; Han, J.; Zhou, L.; Xiong, X.; Wu, Z. Carbon isotope ratios of plants and occurrences of C4 species under different soil moisture regimes in arid region of Northwest China. Physiol. Plant. 2005, 125, 74–81. [Google Scholar] [CrossRef]
- Yang, Q.; Li, X. Investigation of the controlled factors influencing carbon isotope composition of foxtail and common millet on the Chinese Loess Plateau. Sci. China Earth Sci. 2015, 58, 2296–2308. [Google Scholar] [CrossRef]
- Sonawane, B.V.; Cousins, A.B. Mesophyll CO2 conductance and leakiness are not responsive to short- and long-term soil water limitations in the C4 plant Sorghum bicolor. Plant J. 2020, 103, 1590–1602. [Google Scholar] [CrossRef]
- Luo, W.; Wang, X.; Auerswald, K.; Wang, Z.; Bird, M.I.; Still, C.J.; Lu, X.-T.; Han, X. Effects of plant intraspecific variation on the prediction of C3/C4 vegetation ratio from carbon isotope composition of topsoil organic matter across grasslands. J. Plant Ecol. 2021, 14, 628–637. [Google Scholar] [CrossRef]
- Yang, H.; Auerswald, K.; Bai, Y.; Wittmer, M.H.; Schnyder, H. Variation in carbon isotope discrimination in Cleistogenes squarrosa (Trin.) Keng: Patterns and drivers at tiller, local, catchment, and regional scales. J. Exp. Bot. 2011, 62, 4143–4152. [Google Scholar] [CrossRef] [Green Version]
- Eggels, S.; Blankenagel, S.; Schn, C.C.; Avramova, V. The carbon isotopic signature of C4 crops and its applicability in breeding for climate resilience. Theor. Appl. Genet. 2021, 134, 1663–1675. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Richards, R.A. Isotope composition of plant carbon correlates with warter-use efficiency of wheat genotypes. Aust. J. Plant Physiol. 1984, 11, 539–552. [Google Scholar] [CrossRef]
- Murphy, B.P.; Bowman, D.M. Seasonal water availability predicts the relative abundance of C3 and C4 grasses in Australia. Glob. Ecol. Biogeogr. 2007, 16, 160–169. [Google Scholar] [CrossRef]
- Department of Animal Husbandry and Veterinary, Institute of Grasslands, Chinese Academy of Agricultural Sciences (DAHV); Commission for Integrated Survey of Natural Resources, Chinese Academy of Sciences (CISNR). Rangeland Resources of China; China Agricultural Science and Technology Press: Beijing, China, 1996. [Google Scholar]
- Fu, Y.; Yu, Y.; Yao, H. Changes of soil organic carbon of grassland in the Xilinguole, Inner Mongolia from 2000 to 2007. Pr. Sci. 2011, 28, 1589–1597. [Google Scholar]
- O’Leary, M. Carbon isotope fractionation in plants. Phytochemistry 1981, 20, 553–567. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Song, L.; Fan, J.; Harris, W.; Wu, S.; Zhong, H.; Zhou, Y.; Wang, N.; Zhu, X. Adaptive characteristics of grassland community structure and leaf traits along an altitudinal gradient on a subtropical mountain in Chongqing, China. Plant Ecol. 2012, 213, 89–101. [Google Scholar] [CrossRef]
- Yang, H.; Yu, Q.; Sheng, W.; Li, S.; Tian, J. Determination of leaf carbon isotope discrimination in C4 plants under variable N and water supply. Sci. Rep. 2017, 7, 351. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-Y.; Wu, C.-C.; Duan, B.-L.; Korpelainen, H.; Luukkanen, O. Age-related nutrient content and carbon isotope composition in the leaves and branches of Quercus aquifolioides along an altitudinal gradient. Trees 2009, 23, 1109–1121. [Google Scholar] [CrossRef]
- Körner, C.; Farquhar, G.; Wong, S. Carbon isotope discrimination by plants follows latitudinal and altitudinal trends. Oecologia 1991, 88, 30–40. [Google Scholar] [CrossRef]
- Grace, J.; Berninger, F.; Nagy, L. Impacts of climate change on the tree line. Ann. Bot. 2002, 90, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Schleser, G.; Helle, G.; Lucke, A.; Vos, H. Isotope signals as climate proxies: The role of transfer functions in the study of terrestrial archives. Quat. Sci. Rev. 1999, 18, 927–943. [Google Scholar] [CrossRef]
- Wang, G.; Li, J.; Liu, X.; Li, X. Variations in carbon isotope ratios of plants across a temperature gradient along the 400 mm isoline of mean annual precipitation in north China and their relevance to paleovegetation reconstruction. Quat. Sci. Rev. 2013, 63, 83–90. [Google Scholar] [CrossRef]
- Wang, G.; Han, J.; Zhou, L. Relationship between δ13C value of C3 plant and the annual average temperature in northern China. Geol. China 2002, 29, 55–57. [Google Scholar]
- Zhang, C.; Chen, F.J.M. Study on modern plant C-13 in western China and its significance. Chin. J. Geochem. 2003, 22, 97–106. [Google Scholar]
- Wang, L.; Li, X.; Guo, L. The distribution of δ13C value of C3 plant and its responses to climate in arid and semiarid Central East Asia. Quat. Sci. 2006, 26, 955–961. [Google Scholar]
- Zhou, Y.-C.; Cheng, X.-L.; Fan, J.-W.; Xu, X.-Y. Spatial pattern of foliar δ13C in C3-dominated grasslands and its responses to climatic factors in Inner Mongolia, China. J. Northeast. Univ. Nat. Sci. 2016, 37, 273–279. [Google Scholar] [CrossRef]
- Li, J.; Wang, G.; Liu, X.; Han, J.; Liu, M.; Liu, X. Variations in carbon isotope ratios of C3 plants and distribution of C4 plants along an altitudinal transect on the eastern slope of Mount Gongga. Sci. China Ser. D Earth Sci. 2009, 52, 1714–1723. [Google Scholar] [CrossRef]
- Wittmer, M.; Auerswald, K.; Tungalag, R.; Bai, Y.F.; Schaufele, R.; Schnyder, H. Carbon isotope discrimination of C3 vegetation in Central Asian grassland as related to long-term and short-term precipitation patterns. Biogeosciences 2008, 5, 913–924. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.-A.; Han, J.-M.; Liu, D.-S. The carbon isotope composition of C3 herbaceous plants in loess area of northern China. Sci. China Ser. D Earth Sci. 2003, 46, 1069–1076. [Google Scholar] [CrossRef]
- Van de Water, P.; Leavitt, S.; Betancourt, J. Trends in stomatal density and 13C/12C ratios of Pinus flexilis needles during last glacial-interglacial cycle. Science 1994, 264, 239. [Google Scholar] [CrossRef]
- Farquhar, G. On the nature of carbon isotope discrimination in C4 species. Aust. J. Plant Physiol. 1983, 10, 205–226. [Google Scholar] [CrossRef]
- Henderson, S.; Von Caemmerer, S.; Farquhar, G. Short-term measurements of carbon isotope discrimination in several C4 species. Aust. J. Plant Physiol. 1992, 19, 263–285. [Google Scholar] [CrossRef]
- Wang, G.; Han, J.; Zhou, L.; Xiong, X.; Tan, M.; Wu, Z.; Peng, X. The carbon isotope composition of C4 herbaceous plants in loess area of northern China. Sci. China Ser. D Earth Sci. 2005, 35, 1174–1179. [Google Scholar]
- Saliendra, N.Z.; Meinzer, F.C.; Perry, M.; Thom, M. Associations between partitioning of carboxylase activity and bundle sheath leakiness to CO2, carbon isotope discrimination, photosynthesis, and growth in sugarcane. J. Exp. Bot. 1996, 47, 907–914. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhu, Q. Intercellular CO2 concentration, CO2 leakage from bundle sheath cell and stable carbon isotope in leaf of summer maize under water stress. Acta Pedol. Sin. 2009, 46, 1040–1049. [Google Scholar] [CrossRef]
- Lightfoot, E.; Ustunkaya, M.C.; Przelomska, N.; O’Conne, T.C.; Hunt, H.V.; Jones, M.K.; Petrie, C.A. Carbon and nitrogen isotopic variability in foxtail millet (Setaria italica) with watering regime. Rapid Commun. Mass Spectrom. 2020, 34, e8615. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.Y.; Schaeufele, R.; Schnyder, H. Bundle-sheath leakiness and intrinsic water use efficiency of a perennial C-4 grass are increased at high vapour pressure deficit during growth. J. Exp. Bot. 2017, 68, 321–333. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Stipa | Carex | Leymus Chinensis | Cleistogenes | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MS | SS (%) | p Value | MS | SS (%) | p Value | MS | SS (%) | p Value | MS | SS (%) | p Value | |
| GSP | 9.88 | 60.02 | <0.001 | 12.97 | 77.06 | <0.001 | 3.98 | 38.6 | 0.001 | 2.12 | 18.83 | 0.018 |
| GSP Break | 0.33 | 2.93 | 0.32 | |||||||||
| GSP × GSP Break | 2.27 | 20.16 | 0.014 | |||||||||
| MAP | 0.79 | 4.80 | 0.09 | 0.58 | 3.45 | 0.048 | 2.50 | 24.25 | 0.006 | 0.20 | 1.78 | 0.43 |
| GST | 0.01 | 0.06 | 0.84 | 0.01 | 0.06 | 0.85 | 0.48 | 4.66 | 0.17 | 0.41 | 3.64 | 0.27 |
| GST Break | 0.20 | 1.78 | 0.43 | |||||||||
| GST × GST Break | 0.08 | 0.71 | 0.61 | |||||||||
| MAT | 0.03 | 0.18 | 0.75 | 0.01 | 0.06 | 0.74 | 0.02 | 0.19 | 0.77 | 0.42 | 3.73 | 0.26 |
| GSP × GST | 0.04 | 0.24 | 0.71 | 0.81 | 4.81 | 0.02 | 0.28 | 2.72 | 0.30 | 0.04 | 0.36 | 0.73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Li, H.; Xu, X.; Li, Y. Responses of Carbon Isotope Composition of Common C3 and C4 Plants to Climatic Factors in Temperate Grasslands. Sustainability 2022, 14, 7311. https://doi.org/10.3390/su14127311
Zhou Y, Li H, Xu X, Li Y. Responses of Carbon Isotope Composition of Common C3 and C4 Plants to Climatic Factors in Temperate Grasslands. Sustainability. 2022; 14(12):7311. https://doi.org/10.3390/su14127311
Chicago/Turabian StyleZhou, Yongchun, Haibo Li, Xinyang Xu, and Yinghua Li. 2022. "Responses of Carbon Isotope Composition of Common C3 and C4 Plants to Climatic Factors in Temperate Grasslands" Sustainability 14, no. 12: 7311. https://doi.org/10.3390/su14127311
APA StyleZhou, Y., Li, H., Xu, X., & Li, Y. (2022). Responses of Carbon Isotope Composition of Common C3 and C4 Plants to Climatic Factors in Temperate Grasslands. Sustainability, 14(12), 7311. https://doi.org/10.3390/su14127311