
When Scent Becomes a Weapon—Plant Essential Oils as Potent Bioinsecticides




Abstract
:1. Introduction
2. Insects as the Super-Pests of Modern Agriculture
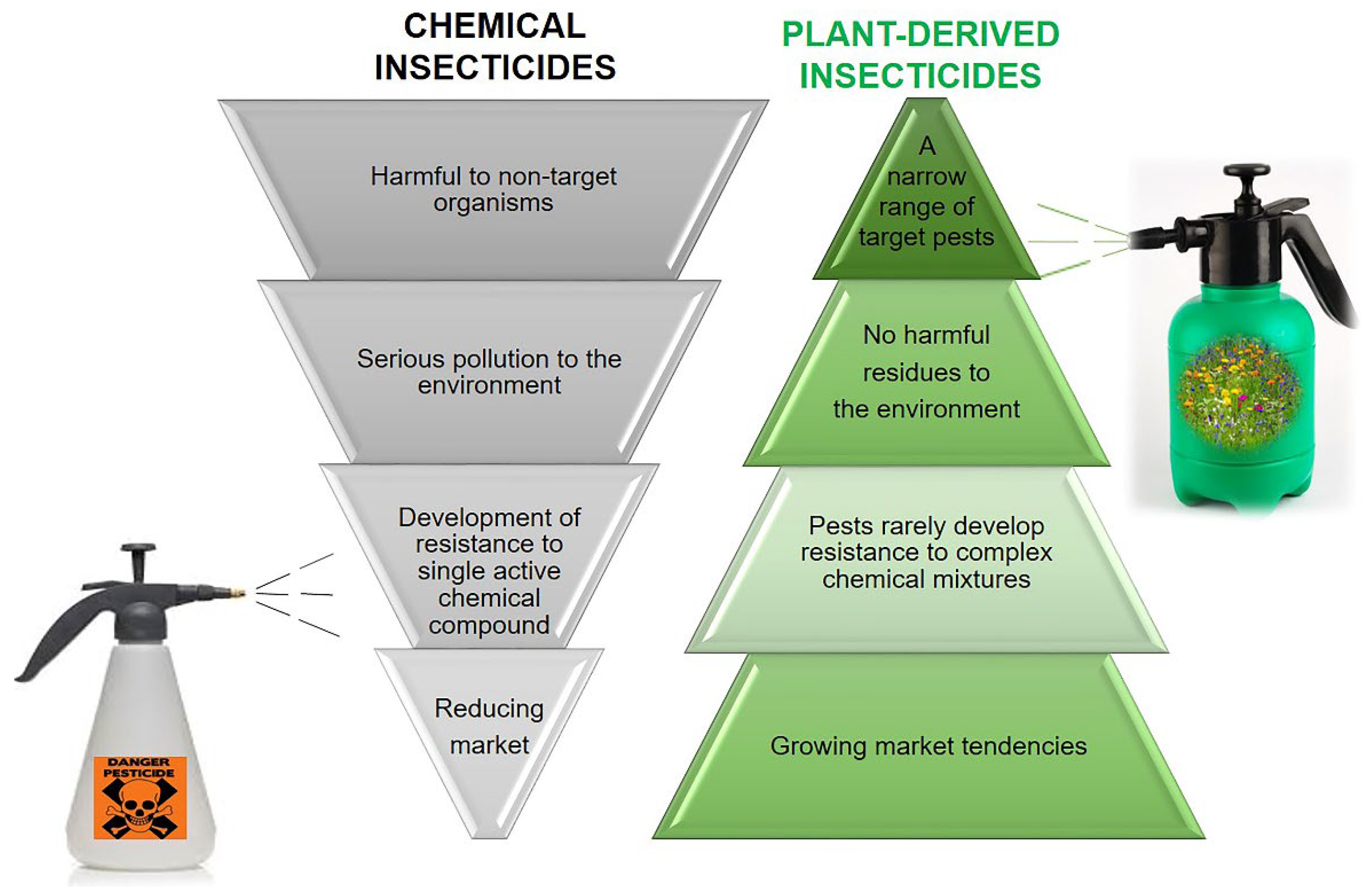
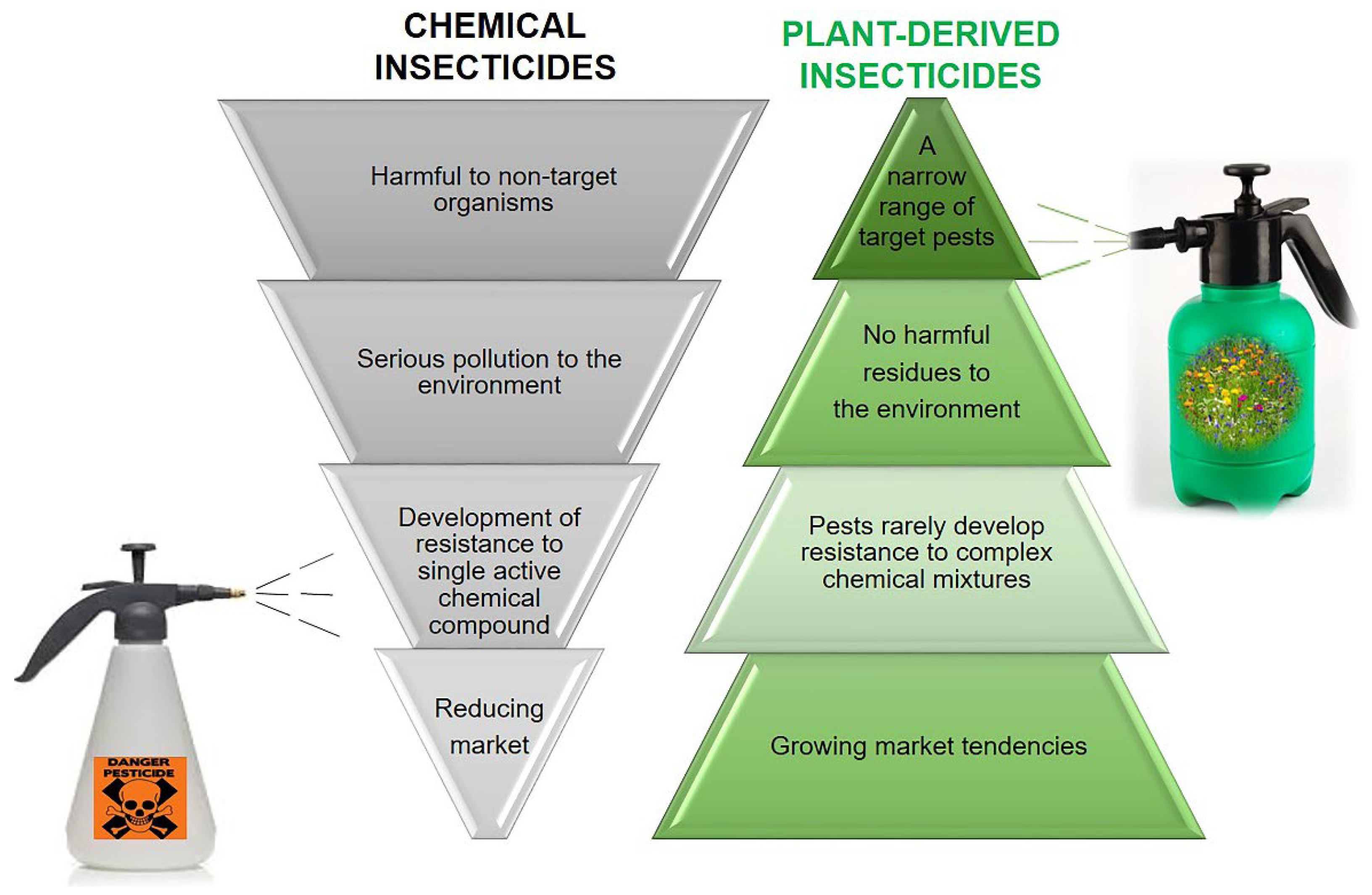
3. The Chemical Teeter: Plant-Based Alternatives for Insecticidal Chemicals
4. Scent That Can Hurt—Essential Oils as Insecticides
5. Mechanisms of Neurotoxic Insecticidal Activity of the Essential Oils—How EOs Conquer the Battlefield in the Targeted Insect Pests
5.1. Inhibitors of Acetylcholinesterase
5.2. Alternant of GABA Receptors
5.3. Ligands of Octopamine Receptors
6. The Future Is Here—New Technologies for EO-Based Insecticide Application
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Moore, D. Blights, Rusts, Bunts, and Mycoses. In Slayers, Saviors, Servants and Sex; Springer: New York, NY, USA, 2001; pp. 19–37. [Google Scholar]
- Mercader, J. Mozambican grass seed consumption during the Middle Stone Age. Science 2009, 326, 1680–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeder, A.M. The Origins of Agriculture in the Near East. Curr. Anthropol. 2011, 52, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations; International Fund for Agricultural Development; World Food Programme; World Health Organization; The United Nations Children’s Fund. The State of Food Security and Nutrition in the World 2020. Transforming Food Systems for Affordable Healthy Diets; FAO: Rome, Italy, 2020. [Google Scholar]
- Boedeker, W.; Watts, M.; Clausing, P.; Marquez, E. The global distribution of acute unintentional pesticide poisoning: Estimations based on a systematic review. BMC Public Health 2020, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Carriger, J.F.; Rand, G.M.; Gardinali, P.R.; Perry, W.B.; Tompkins, M.S.; Fernandez, A.M. Pesticides of potential ecological concern in sediment from South Florida canals: An ecological risk prioritization for aquatic arthropods. Soil Sediment Contam. 2006, 15, 21–45. [Google Scholar] [CrossRef]
- Katagi, T.; Tanaka, H. Metabolism, bioaccumulation, and toxicity of pesticides in aquatic insect larvae. J. Pestic. Sci. 2016, 41, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Tongo, I.; Onokpasa, A.; Emerure, F.; Balogun, P.T.; Enuneku, A.A.; Erhunmwunse, N.; Asemota, O.; Ogbomida, E.; Ogbeide, O.; Ezemonye, L. Levels, bioaccumulation and biomagnification of pesticide residues in a tropical freshwater food web. Int. J. Environ. Sci. Technol. 2022, 19, 1467–1482. [Google Scholar] [CrossRef]
- Koul, O.; Walia, S.; Dhaliwal, G.S. Essential oils as green pesticides: Potential and constraints. Biopestic. Int. 2008, 4, 63–84. [Google Scholar]
- Cingel, A.; Savić, J.; Lazarević, J.; Ćosić, T.; Raspor, M.; Smigocki, A.; Ninković, S. Extraordinary adaptive plasticity of Colorado potato beetle: “Ten-Striped Spearman” in the era of biotechnological warfare. Int. J. Mol. Sci. 2016, 17, 1538. [Google Scholar] [CrossRef]
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Change 2013, 3, 985–988. [Google Scholar] [CrossRef]
- García-Lara, S.; Serna Saldivar, S.O. Insect Pests. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: London, UK, 2016; pp. 432–436. [Google Scholar]
- Willis, K.J. State of the World’s Plants 2017; Report; Royal Botanic Gardens: Kew, UK, 2017. [Google Scholar]
- Whalon, M.E.; Mota-Sanchez, D.; Hollingworth, R.M. Analysis of global pesticide resistance in arthropods. Glob. Pestic. Resist. Arthropods 2008, 5, 31. [Google Scholar]
- Bishop, B.A.; Grafius, E.J. Insecticide resistance in the Colorado potato beetle. Chrysomelidae Biol. 1996, 1, 355–377. [Google Scholar]
- Venette, R.C.; Hutchison, W.D. Invasive insect species: Global challenges, strategies & opportunities. Front. Insect Sci. 2021, 1, 650520. [Google Scholar]
- Bradshaw, C.J.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; Barbet-Massin, M.; Salles, J.M.; Simard, F.; Courchamp, F. Massive yet grossly underestimated global costs of invasive insects. Nat. Commun. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; et al. Complex responses of global insect pests to climate warming. Front. Ecol. Environ. 2020, 18, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [Green Version]
- Alyokhin, A.; Baker, M.; Mota-Sanchez, D.; Dively, G.; Grafius, E. Colorado Potato Beetle Resistance to Insecticides. Am. J. Potato Res. 2008, 85, 395–413. [Google Scholar] [CrossRef]
- Agosta, S.J. On ecological fitting, plant–insect associations, herbivore host shifts, and host plant selection. Oikos 2006, 114, 556–565. [Google Scholar] [CrossRef]
- Grapputo, A.; Boman, S.; Lindstroem, L.; Lyytinen, A.; Mappes, J. The voyage of an invasive species across continents: Genetic diversity of North American and European Colorado potato beetle populations. Mol. Ecol. 2005, 14, 4207–4219. [Google Scholar] [CrossRef]
- Weber, D. Colorado beetle: Pest on the move. Pestic. Outlook 2003, 14, 256–259. [Google Scholar] [CrossRef]
- Cingel, A.; Savić, J.; Vinterhalter, B.; Vinterhalter, D.; Kostić, M.; Šešlija Jovanović, D.; Smigocki, A.; Ninković, S. Growth and development of Colorado potato beetle larvae, Leptinotarsa decemlineata, on potato plants expressing the oryzacystatin II proteinase inhibitor. Transgenic Res. 2015, 24, 729–740. [Google Scholar] [CrossRef]
- Insecticides Market by Type (Pyrethroids, Organophosphorus, Carbamates, Organochlorine, Botanicals), Crop Type, Mode of Application, Formulation (WP, EC, SC, EW, ME, GR), and Region-Global Forecast to 2022. Available online: https://www.marketsandmarkets.com/Market-Reports/insecticide-market-142427569.html (accessed on 26 April 2020).
- Market Value of Bioinsecticides Worldwide in 2016 and 2020 with a Forecast for 2025. Available online: https://www.statista.com/statistics/1024754/market-value-bioinsecticides-globally/ (accessed on 26 April 2022).
- Johansen, C.A. Pesticides and pollinators. Annu. Rev. Entomol. 1977, 22, 177–192. [Google Scholar] [CrossRef] [Green Version]
- Copping, L.G.; Menn, J.J. Biopesticides: A review of their action, applications and efficacy. Pest Manag. Sci. 2000, 56, 651–676. [Google Scholar] [CrossRef]
- Pino, O.; Sánchez, Y.; Rojas, M.M. Plant secondary metabolites as an alternative in pest management. I: Background, research approaches and trends. Rev. Prot. Veg. 2013, 28, 81–89. [Google Scholar]
- Lima, P.S.A.; Santana, D.R.E.; Santos, C.C.A.; Silva, E.J.; Ribeiro, T.G.; Pinheiro, A.M.Í.; Santos, T.B.F.; Blank, F.A.; Araújo, A.P.A.; Bacci, L. Insecticide activity of botanical compounds against Spodoptera frugiperda and selectivity to the predatory bug Podisus nigrispinus. Crop Prot. 2020, 136, 105230. [Google Scholar] [CrossRef]
- Ma, S.; Jia, R.; Guo, M.; Qin, K.; Zhang, L. Insecticidal activity of essential oil from Cephalotaxus sinensis and its main components against various agricultural pests. Ind. Crops Prod. 2020, 150, 112403. [Google Scholar] [CrossRef]
- Ríos, N.; Stashenko, E.E.; Duque, J.E. Evaluation of the insecticidal activity of essential oils and their mixtures against Aedes aegypti (Diptera: Culicidae). Rev. Bras. Entomol. 2017, 61, 307–311. [Google Scholar] [CrossRef]
- Farag, M.A.; Ezzat, S.M.; Salama, M.M.; Tadros, M.G.; Serya, R.A. Anti-acetylcholinesterase activity of essential oils and their major constituents from four Ocimum species. Z Nat. C J. Biosci. 2016, 71, 393–402. [Google Scholar] [CrossRef]
- Mishra, B.B.; Tripathi, S.P.; Tripathi, C.P.M. Repellent effect of leaves essential oils from Eucalyptus globulus (Mirtaceae) and Ocimum basilicum (Lamiaceae) against two major stored grain insect pests of Coleopterons. Nat. Sci. 2012, 10, 50–54. [Google Scholar]
- Kiran, S.; Anupam, K.; Laluram, P.; Ramalakshmi, K.; Bhanu, P. Assessment of toxicity and biochemical mechanisms underlying the insecticidal activity of chemically characterized Boswellia carterii essential oil against insect pest of legume seeds. Pestic. Biochem. Phys. 2017, 139, 17–23. [Google Scholar]
- Ahmed, N.; Alam, M.; Saeed, M.; Ullah, H.; Iqbal, T.; Al-Mutairi, K.A.; Shahjeer, K.; Ullah, R.; Ahmed, S.; Ahmed, N.A.A.H.; et al. Botanical Insecticides Are a Non-Toxic Alternative to Conventional Pesticides in the Control of Insects and Pests. In Global Decline of Insects; El-Shafie, H., Ed.; IntechOpen: London, UK, 2021. [Google Scholar]
- Hummelbrunner, L.; Isman, M.B. Acute, Sublethal, Antifeedant, and Synergistic Effects of Monoterpenoid Essential Oil Compounds on the Tobacco Cutworm, Spodopteralitura (Lep., Noctuidae). J. Agric. Food Chem. 2001, 49, 715–720. [Google Scholar] [CrossRef]
- Hierro, I.; Valero, A.; Perez, P.; Gonzalez, P.; Cabo, M.; Montilla, P.; Navarro, C. Action of different monoterpenic compounds against Anisakis simplex L3 larvae. Phytomedicine 2004, 11, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Ajayi, E.; Appel, G.; Fadamiro, Y. Fumigation toxicity of essential oil monoterpenes to Callosobruchus maculatus (Coleoptera: Chrysomelidae: Bruchinae). J. Insects 2014, 2014, 917212. [Google Scholar] [CrossRef] [Green Version]
- Sadgrove, N.J.; Padilla-González, G.F.; Phumthum, M. Fundamental Chemistry of Essential Oils and Volatile Organic Compounds, Methods of Analysis and Authentication. Plants 2022, 11, 789. [Google Scholar] [CrossRef] [PubMed]
- Zhi-Jing, N.; Xin, W.; Yi, S.; Kiran, T.; Jinzhi, H.; Jian-Guo, Z.; Fei, H.; Zhao-Jun, W. Recent updates on the chemistry, bioactivities, mode of action, and industrial applications of plant essential oils. Trends Food Sci. Technol. 2021, 110, 78–89. [Google Scholar]
- Paes, J.L.; LRD’A, F.; Dhingra, O.D.; Cecon, P.R.; Silva, T.A. Insecticidal fumigant action of mustard essential oil against Sitophilus zeamais in maize grains. Crop Prot. 2012, 34, 56–58. [Google Scholar] [CrossRef]
- Koul, O.; Singh, R.; Kaur, B.; Kanda, D. Comparative study on the behavioral response and acute toxicity of some essential oil compounds and their binary mixtures to larvae of Helicoverpa armigera, Spodoptera litura and Chilo partellus. Ind. Crops Prod. 2013, 49, 428–436. [Google Scholar] [CrossRef]
- Shahriari, M.; Zibaee, A.; Sahebzadeh, N.; Shamakhi, L. Effects of α-pinene, trans-anethole, and thymol as the essential oil constituents on antioxidant system and acetylcholine esterase of Ephestia kuehniella Zeller (Lepidoptera: Pyralidae). Pestic. Biochem. Phys. 2018, 150, 40–47. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Santos, A.A.; Santana, A.S.; Lima, A.P.S.; Melo, C.R.; Santana, E.D.; Sampaio, T.S.; Blank, A.F.; Araújo, A.P.A.; Cristaldo, P.F.; et al. Essential oil of Lippia sidoides and its major compound thymol: Toxicity and walking response of populations of Sitophilus zeamais (Coleoptera: Curculionidae). Crop Prot. 2018, 112, 33–38. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G.; Canale, A.; Maggi, F.; Mártonfi, P. Exploring essential oils of Slovak medicinal plants for insecticidal activity: The case of Thymus alternans and Teucrium montanum subsp. jailae. Food Chem.Toxicol. 2020, 138, 111203. [Google Scholar] [CrossRef]
- Demeter, S.; Lebbe, O.; Hecq, F.; Nicolis, S.C.; Kenne Kemene, T.; Martin, H.; Fauconnier, M.L.; Hance, T. Insecticidal Activity of 25 Essential Oils on the Stored Product Pest, Sitophilus granarius. Foods 2021, 10, 200. [Google Scholar] [CrossRef]
- Pavela, R.; Maggi, F.; Petrelli, R.; Cappellacci, L.; Buccioni, M.; Palmieri, A.; Canale, A.; Benelli, G. Outstanding insecticidal activity and sublethal effects of Carlina acaulis root essential oil on the housefly, Musca domestica, with insights on its toxicity on human cells. Food Chem. Toxicol. 2020, 136, 111037. [Google Scholar] [CrossRef]
- Butterworth, J.; Morgan, E. Isolation of a Substance that suppresses Feeding in Locusts. Chem. Comm. 1968, 1, 23. [Google Scholar] [CrossRef]
- Abdel-Shafy, S.; Zayed, A.A. In vitro acaricidal effect of plant extract of neem seed oil (Azadirachta indica) on egg, immature, and adult stages of Hyalomma anatolicum excavatum (Ixodoidea: Ixodidae). Vet. Parasitol. 2002, 106, 89–96. [Google Scholar] [CrossRef]
- Tavares, W.S.; Costa, M.A.; Cruz, I.; Silveira, R.D.; Serrão, J.E.; Zanuncio, J.C. Selective effects of natural and synthetic insecticides on mortality of Spodoptera frugiperda (Lepidoptera: Noctuidae) and its predator Eriopis connexa (Coleoptera: Coccinellidae). J. Environ. Sci. Health B. 2010, 45, 557–561. [Google Scholar] [CrossRef]
- Scudeler, E.L.; dos Santos, D.C. Effects of neem oil (Azadirachta indica A. Juss) on midgut cells of predatory larvae Ceraeochrysa claveri (Navás, 1911) (Neuroptera: Chrysopidae). Micron 2013, 44, 125–132. [Google Scholar] [CrossRef]
- Dubey, N.K.; Shukla, R.A.V.I.N.D.R.A.; Kumar, A.S.H.O.K.; Singh, P.; Prakash, B. Global scenario on the application of natural products in integrated pest management programmes. Nat. Prod. Plant Pest Manag. 2011, 1, 1–20. [Google Scholar]
- Wei-Hong, T.; Zhan-qian, S. Research on effect of four natural ultraviolet light absorbers on photostabilization of azadirachtin-A. Agric. Sci. China 2006, 5, 855–858. [Google Scholar]
- Chaudhary, S.; Kanwar, R.K.; Sehgal, A.; Cahill, D.M.; Barrow, C.J.; Sehgal, R.; Kanwar, J.R. Progress on Azadirachta indica based biopesticides in replacing synthetic toxic pesticides. Front. Plant Sci. 2017, 8, 610. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Pesticide Evaluation Scheme, Vector Ecology and Management; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Bradberry, S.M.; Cage, S.A.; Proudfoot, A.T.; Vale, J.A. Poisoning due to pyrethroids. Toxicol. Rev. 2005, 24, 93–106. [Google Scholar] [CrossRef]
- Ramchandra, A.M.; Chacko, B.; Victor, P.J. Pyrethroid Poisoning. Indian J. Crit. Care Med. 2019, 23, 267–271. [Google Scholar] [CrossRef]
- Ray, D.E.; Forshaw, P.J. Pyrethroid insecticides: Poisoning syndromes, synergies, and therapy. J. Toxicol. Clin. Toxicol. 2000, 38, 95–101. [Google Scholar] [CrossRef]
- Isman, M.B. Commercial development of plant essential oils and their constituents as active ingredients in bioinsecticides. Phytochem. Rev. 2020, 19, 235–241. [Google Scholar] [CrossRef]
- Sparks, T.C.; Nauen, R. IRAC: Mode of action classification and insecticide resistance management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.R.; Scott, I.M.; Sims, S.R.; Trudeau, V.L.; Arnason, J.T. The effect of a synergistic concentration of a Piper nigrum extract used in conjunction with pyrethrum upon gene expression in Drosophila melanogaster. Insect. Mol. Biol. 2006, 15, 329–339. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Smagghe, G.; Stark, J.D.; Desneux, N. Pesticide-induced stress in arthropod pests for optimized integrated pest management programs. Annu. Rev. Entomol. 2016, 61, 43–62. [Google Scholar] [CrossRef] [Green Version]
- Van Toor, R.F.; Foster, S.P.; Anstead, J.A.; Mitchinson, S.; Fenton, B.; Kasprowicz, L. Insecticide resistance and genetic composition of Myzus persicae (Hemiptera: Aphididae) on field potatoes in New Zealand. Crop Prot. 2008, 27, 236–247. [Google Scholar] [CrossRef]
- Casida, J.E.; Durkin, K.A. Neuroactive insecticides: Targets, selectivity, resistance, and secondary effects. Annu. Rev. Entomol. 2013, 58, 99–117. [Google Scholar] [CrossRef]
- Owokotomo, I.A.; Ekundayo, O.; Abayomi, T.G.; Chukwuka, A.V. In-vitro anti-cholinesterase activity of essential oil from four tropical medicinal plants. Toxicol. Rep. 2015, 2, 850–857. [Google Scholar] [CrossRef] [Green Version]
- Park, C.G.; Jang, M.; Yoon, K.A.; Kim, J. Insecticidal and acetylcholinesterase inhibitory activities of Lamiaceae plant essential oils and their major components against Drosophila suzukii (Diptera: Drosophilidae). Ind. Crop. Prod. 2016, 89, 507–513. [Google Scholar] [CrossRef]
- López, V.; Cascella, M.; Benelli, G.; Maggi, F.; Gómez-Rincón, C. Green drugs in the fight against Anisakis simplex—larvicidal activity and acetylcholinesterase inhibition of Origanum compactum essential oil. Parasitol. Res. 2018, 117, 861–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva Barbosa, D.C.; Holanda, V.N.; de Assis, C.R.D.; de Oliveira Farias, J.C.R.; Henrique doNascimento, P.; da Silva, W.V.; Navarro, D.M.D.A.F.; da Silva, M.V.; de Menezes Lima, V.L.; dos Santos Correia, M.T. Chemical composition and acetylcholinesterase inhibitory potential, in silico, of Myrciaria floribunda (H. West ex Willd.) O. Berg fruit peel essential oil. Ind. Crops Prod. 2020, 151, 112372. [Google Scholar] [CrossRef]
- Bhavya, M.L.; Obulaxmi, S.; Devi, S.S. Efficacy of Ocimum tenuiflorum essential oil as grain protectant against coleopteran beetle, infesting stored pulses. J. Food Sci. Technol. 2021, 58, 1611–1616. [Google Scholar] [CrossRef]
- Isman, M.B. Plant essential oils for pest and disease management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Miyazawa, M.; Yamafuji, C. Inhibition of acetylcholinesterase activity by bicyclic monoterpenoids. J. Agric. Food Chem. 2005, 53, 1765–1768. [Google Scholar] [CrossRef]
- Tong, F.; Gross, A.D.; Dolan, M.C.; Coats, J.R. The phenolic monoterpenoid carvacrol inhibits the binding of nicotine to the housefly nicotinic acetylcholine receptor. Pest Manag. Sci. 2013, 69, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Bhavya, M.L.; Chandu, A.G.S.; Devi, S.S. Ocimum tenuiflorum oil, a potential insecticide against rice weevil with anti-acetylcholinesterase activity. Ind. Crops Prod. 2018, 126, 434–439. [Google Scholar] [CrossRef]
- Yeom, H.J.; Kang, J.S.; Kim, G.H.; Park, I.K. Insecticidal and acetylcholine esterase inhibition activity of Apiaceae plant essential oils and their constituents against adults of German cockroach (Blattella germanica). J. Agric. Food Chem. 2012, 60, 7194–7203. [Google Scholar] [CrossRef]
- Mills, C.; Cleary, B.V.; Walsh, J.J.; Gilmer, J.F. Inhibition of acetylcholinesterase by tea tree oil. J. Pharm. Pharmacol. 2004, 56, 375–379. [Google Scholar] [CrossRef]
- Patel, M.B.; Amin, D. Sphaeranthus indicus flower derived constituents exhibits synergistic effect against acetylcholinesterase and possess potential antiamnestic activity. J. Complement. Integr. Med. 2012, 9, 1. [Google Scholar] [CrossRef]
- Taylor, P.; Radić, Z. The cholinesterases: From genes to proteins. Annu. Rev. Pharmacol. Toxicol. 1994, 34, 281–320. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, B.; Yang, F.; Sun, Q.; Yang, Z.; Zhu, L. Chemical compositionand anti-acetyl cholinesterase activity of flower essential oils of artemisiaannuaat different flowering stage. Iran. J. Pharm. Res. 2011, 10, 265. [Google Scholar]
- Savelev, S.; Okello, E.; Perry, N.S.L.; Wilkins, R.M.; Perry, E.K. Synergistic and antagonistic interactions of anticholinesterase terpenoids in Salvia lavandulaefolia essential oil. Pharmacol. Biochem. Behav. 2003, 75, 661–668. [Google Scholar] [CrossRef]
- Anderson, J.A.; Coats, J.R. Acetylcholinesterase inhibition by nootkatone and carvacrol in arthropods. Pestic. Biochem. Physiol. 2012, 102, 124–128. [Google Scholar] [CrossRef] [Green Version]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular targets for components of essential oils in the insect nervous system—A review. Molecules 2017, 23, 34. [Google Scholar] [CrossRef] [Green Version]
- Raymond-Delpech, V.; Matsuda, K.; Sattelle, B.M.; Rauh, J.J.; Sattelle, D.B. Ion channels: Molecular targets of neuroactive insecticides. Invertebr. Neurosci. 2005, 5, 119–133. [Google Scholar] [CrossRef]
- Ashby, J.A.; McGonigle, I.V.; Price, K.L.; Cohen, N.; Comitani, F.; Dougherty, D.A.; Molteni, C.; Lummis, S.C. GABA binding to an insect GABA receptor: A molecular dynamics and mutagenesis study. Biophys. J. 2012, 103, 2071–2081. [Google Scholar] [CrossRef] [Green Version]
- Henderson, J.E.; Soderlund, D.M.; Knipple, D.C. Characterization of a putative γ-aminobutyric-acid (GABA) receptor β-subunit gene from Drosophila melanogaster. Biochem. Biophys. Res. Commun. 1993, 193, 474–482. [Google Scholar] [CrossRef]
- Tong, F.; Coats, J.R. Effects of monoterpenoid insecticides on [3H]-TBOB binding in house fly GABA receptor and 36Cl − uptakein American cockroach ventral nerve cord. Pestic. Biochem. Phys. 2010, 98, 317–324. [Google Scholar] [CrossRef]
- Höld, K.M.; Sirisoma, N.S.; Ikeda, T.; Narahashi, T.; Casida, J.E. α-Thujone (the active component of absinthe): γ-aminobutyric acid type A receptor modulation and metabolic detoxification. Proc. Natl. Acad. Sci. USA 2000, 97, 3826–3831. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Rao, Y.; Rao, Y. A subset of octopaminergic neurons are important for Drosophila aggression. Nat. Neurosci. 2008, 11, 1059–1067. [Google Scholar] [CrossRef]
- Evans, P.D. Multiple receptor types for octopamine in the locust. J. Physiol. Lond. 1981, 318, 99–122. [Google Scholar] [CrossRef] [Green Version]
- Roeder, T. A new octopamine receptor class in locust nervous tissue, the octopamine 3 (OA3) receptor. Life Sci. 1992, 50, 21–28. [Google Scholar] [CrossRef]
- Evans, P.D. A modulatory octopaminergic neurone increases cyclic nucleotide levels in locust skeletal muscle. J. Physiol. 1984, 348, 307–324. [Google Scholar] [CrossRef] [Green Version]
- Nathanson, J.A. Octopamine receptors, adenosine 3′, 5′-monophosphate, and neural control of firefly flashing. Science 1979, 203, 65–68. [Google Scholar] [CrossRef]
- Bodnarky, R.P. Cyclic nucleotides. In Endocrinology of Insects; Downer, R.G., Laufer, H., Eds.; Alan R. Liss. Inc.: New York, NY, USA, 1983; pp. 567–614. [Google Scholar]
- Enan, E. Insecticidal activity of essential oils: Octopaminergic sites of action. Comp. Biochem. Physiol. Part-C Toxicol. Pharmacol. 2001, 130, 325–337. [Google Scholar] [CrossRef]
- Enan, E.E. Molecular and pharmacological analysis of an octopamine receptor from American cockroach and fruit fly in response to plant essential oils. Arch. Insect Biochem. Physiol. 2005, 59, 161–171. [Google Scholar] [CrossRef]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef]
- Price, D.N.; Berry, M.S. Comparison of effects of octopamine and insecticidal essential oils on activity in the nerve cord, foregut, and dorsal unpaired median neurons of cockroaches. J. Insect Physiol. 2006, 52, 309–319. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Upadhyay, S. Repellent and insecticidal activities of Hyptis suaveolens (Lamiaceae) leaf essential oil against four stored-grain coleopteran pests. Int. J. Trop. Insect. Sci. 2009, 29, 219–228. [Google Scholar] [CrossRef]
- Turek, C.; Stintzing, F.C. Stability of essential oils: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 40–53. [Google Scholar] [CrossRef]
- Seng, K.K.; Loong, W.V. Introductory Chapter: From Microemulsions to Nanoemulsions. In Nanoemulsions-Properties, Fabrications and Applications; Koh, K.S., Wong, V.L., Eds.; IntechOpen: London, UK, 2019. [Google Scholar]
- Anton, N.; Vandamme, T.F. Nano-emulsions and micro-emulsions: Clarifications of the critical differences. Pharm. Res. 2011, 28, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Cimino, C.; Maurel, O.M.; Musumeci, T.; Bonaccorso, A.; Drago, F.; Souto, E.M.B.; Pignatello, R.; Carbone, C. Essential Oils: Pharmaceutical Applications and Encapsulation Strategies into Lipid-Based Delivery Systems. Pharmaceutics 2021, 13, 327. [Google Scholar] [CrossRef]
- Rodrigues, C.R.; Ferreira, A.M.; Vilhena, J.C.E.; Almeida, F.B.; Cruz, R.A.S.; Florentino, A.C.; Souto, R.N.P. Development of a larvicidal nanoemulsion with Copaiba (Copaifera duckei) oleoresin. Rev. Bras. Farmacogn. 2014, 24, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Duarte, J.L.; Amado, J.R.; Oliveira, A.E.; Cruz, R.A.; Ferreira, A.M.; Souto, R.N.; Falcão, D.Q.; Carvalho, J.C.; Fernandes, C.P. Evaluation of larvicidal activity of a nanoemulsion of Rosmarinus officinalis essential oil. Rev. Bras. Farmacogn. 2015, 25, 189–192. [Google Scholar] [CrossRef] [Green Version]
- Pant, M.; Dubey, S.; Patanjali, P.K.; Naik, S.N.; Sharma, S. Insecticidal activity of eucalyptus oil nanoemulsion with karanja and jatropha aqueous filtrates. Int. Biodeterior. Biodegrad. 2014, 91, 119–127. [Google Scholar] [CrossRef]
- Adak, T.; Barik, N.; Patil, N.B.; Govindharaj, G.P.P.; Gadratagi, B.G.; Annamalai, M.; Mukherjee, A.K.; Rath, P.C. Nanoemulsion of eucalyptus oil: An alternative to synthetic pesticides against two major storage insects Sitophilus oryzae (L.) and Tribolium castaneum (Herbst)) of rice. Ind. Crops Prod. 2020, 143, 111849. [Google Scholar] [CrossRef]
- Yasur, J.; Rani, P.U. Lepidopteran insect susceptibility to silver nanoparticles and measurement of changes in their growth, development and physiology. Chemosphere 2015, 124, 92–102. [Google Scholar] [CrossRef]
- Giunti, G.; Palermo, D.; Laudani, F.; Algeri, G.M.; Campolo, O.; Palmeri, V. Repellence and acute toxicity of a nano-emulsion of sweet orange essential oil toward two major stored grain insect pests. Ind. Crops Prod. 2019, 142, 111869. [Google Scholar] [CrossRef]
- Alizadeh-Sani, M.; Mohammadian, E.; McClements, D.J. Eco-friendly active packaging consisting of nanostructured biopolymer matrix reinforced with TiO2 and essential oil: Application for preservation of refrigerated meat. Food Chem. 2020, 322, 126782. [Google Scholar] [CrossRef]
- Mizielinska, M.; Nawrotek, P.; Stachurska, X.; Ordon, M.; Bartkowiak, A. Packaging covered with antiviral and antibacterial coatings based on ZnO nanoparticles supplemented with geraniol and carvacrol. Int. J. Mol. Sci. 2021, 22, 1717. [Google Scholar] [CrossRef]
- Simonin, M.; Richaume, A.; Guyonnet, J.P.; Dubost, A.; Martins, J.M.F.; Pommier, T. Titanium dioxide nanoparticles strongly impact soil microbial function by affecting archaeal nitrifiers. Sci. Rep. 2016, 6, 33643. [Google Scholar] [CrossRef]
- Jabir, M.S.; Taha, A.A.; Sahib, U.I. Linalool loaded on glutathione-modified gold nanoparticles: A drug delivery system for a successful antimicrobial therapy. Artif. Cells Nanomed. Biotechnol. 2018, 46, 345–355. [Google Scholar] [CrossRef]
- Aldosary, S.K.; El-Rahman, S.N.A.; Al-Jameel, S.S.; Alromihi, N.M. Antioxidant and antimicrobial activities of Thymus vulgaris essential oil contained and synthesis thymus (Vulgaris) silver nanoparticles. Braz. J. Biol. 2021, 83, e244675. [Google Scholar] [CrossRef]
- Suresh, U.; Murugan, K.; Panneerselvam, C.; Cianfaglione, K.; Wang, L.; Maggi, F. Encapsulation of sea fennel (Crithmum maritimum) essential oil in nanoemulsion and SiO2 nanoparticles for treatment of the crop pest Spodoptera litura and the dengue vector Aedes aegypti. Ind. Crops Prod. 2020, 158, 113033. [Google Scholar] [CrossRef]
- Attia, R.G.; Rizk, S.A.; Hussein, M.A.; Fattah, H.M.A.; Khalil, M.; Ma’Moun, S.A. Effect of cinnamon oil encapsulated with silica nanoparticles on some biological and biochemical aspects of the rice moth, Corcyra cephalonica (Staint.) (Lepidoptera: Pyralidae). Ann. Agric. Sci. 2020, 65, 1–5. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Hwang, I.-C.; Park, J.-W.; Park, H.-J. Photoprotection for deltamethrin using chitosan-coated beeswax solid lipid nanoparticles. Pest. Manag. Sci. 2012, 68, 1062–1068. [Google Scholar] [CrossRef]
- Lin, L.; Cui, H.; Zhou, H.; Zhang, X.; Bortolini, C.; Chen, M.; Liu, L.; Dong, M. Nanoliposomes containing Eucalyptus citriodora as antibiotic with specific antimicrobial activity. Chem. Comm. 2015, 51, 2653–2655. [Google Scholar] [CrossRef]
- Pan, Y.; Tikekar, R.V.; Nitin, N. Distribution of a model bioactive within solid lipid nanoparticles and nanostructured lipid carriers influences its loading efficiency and oxidative stability. Int. J. Pharm. 2016, 511, 322–330. [Google Scholar] [CrossRef] [Green Version]
- Nair, R.; Varghese, S.H.; Nair, B.G.; Maekawa, T.; Yoshida, Y.; Kumar, D.S. Nanoparticulate material delivery to plants. Plant Sci. 2010, 179, 154–163. [Google Scholar] [CrossRef]
- Lopez, M.D.; Maudhuit, A.; Pascual-Villalobos, M.J.; Poncelet, D. Development of formulations to improve the controlled-release of linalool to be applied as an insecticide. J. Agric. Food Chem. 2012, 60, 1187–1192. [Google Scholar] [CrossRef]
- Kumar, S.; Bhanjana, G.; Sharma, A.; Dilbaghi, N.; Sidhu, M.C.; Kim, K.H. Development of nanoformulation approaches for the control of weeds. Sci. Total Environ. 2017, 586, 1272–1278. [Google Scholar] [CrossRef]
- Ramasamy, T.; Ruttala, H.B.; Gupta, B.; Poudel, B.K.; Choi, H.G.; Yong, C.S.; Kim, J.O. Smart chemistry-based nanosized drug delivery systems for systemic applications: A comprehensive review. J. Control. Release 2017, 258, 226–253. [Google Scholar] [CrossRef]
- Perlatti, B.; Bergo, P.L.d.S.; Silva, M.F.d.G.F.d.; Fernandes, J.B.; Forim, M.R. Polymeric nanoparticle-based insecticides: A controlled release purpose for agrochemicals. In Insecticides−Development of Safer and More Effective Technologies; Trdan, S., Ed.; Intech Open Access Publisher: London, UK, 2013; pp. 523–550. [Google Scholar]
- Kulkarni, A.R.; Soppimath, K.S.; Aminabhavi, T.M.; Dave, A.M.; Mehta, M.H. Glutaraldehyde crosslinked sodium alginate beads containing liquid pesticide for soil application. J. Control. Release 2000, 63, 97–105. [Google Scholar] [CrossRef]
- Ye, Z.; Guo, J.; Wu, D.; Tan, M.; Xiong, X.; Yin, Y.; He, G. Photo-responsive shell cross-linked micelles based on carboxymethyl chitosan and their application in controlled release of pesticide. Carbohydr. Polym. 2015, 132, 520–528. [Google Scholar] [CrossRef]
- Vishwakarma, G.S.; Gautam, N.; Babu, J.N.; Mittal, S.; Jaitak, V. Polymeric encapsulates of essential oils and their constituents: A review of preparation techniques, characterization, and sustainable release mechanisms. Polym. Rev. 2016, 56, 668–701. [Google Scholar] [CrossRef]
- Campos, E.V.R.; Proença, P.L.F.; Oliveira, J.L.; Melville, C.C.; Vechia, J.F.D.; de Andrade, D.J.; Fraceto, L.F. Chitosan nanoparticles functionalized with β-cyclodextrin: A promising carrier for botanical pesticides. Sci. Rep. 2018, 8, 2067. [Google Scholar] [CrossRef]
- Ahmadi, Z.; Saber, M.; Bagheri, M.; Mahdavinia, G.R. Achillea millefolium essential oil and chitosan nanocapsules with enhanced activity against Tetranychus urticae. J. Pest Sci. 2018, 91, 837–848. [Google Scholar] [CrossRef]
- Rajkumar, V.; Gunasekaran, C.; Dharmaraj, J.; Chinnaraj, P.; Paul, C.A.; Kanithachristy, I. Structural characterization of chitosan nanoparticle loaded with Piper nigrum essential oil for biological efficacy against the stored grain pest control. Pestic. Biochem. Physiol. 2020, 166, 104566. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Plant Species | Insects | Major EO Constituents | Lethal (LC50) and Inhibition * (IC50 *) Concentrations | Ref. No |
|---|---|---|---|---|
| Cephalotaxus sinensis | Megoura japonica | α-Pinene | 8.82 mg/L 6.74 mg/L 7.35 mg/L 3.57 mg/L | [31] |
| Plutella xylostella | β-Caryophyllene | |||
| Sitophilus zeamais | Germacrene D | |||
| Brassica nigra | Sitophilus zeamais | Allyl isothiocyanate | 6.19 µL/L (larvae) 7.01 µL/L (adults) | [42] |
| / | Thymol | 189.7; 28.5; 290.8 µg/larva 462.4; 85.5; 431.5 µg/larva 412.1; 126.6; 406.8 µg/larva 409.7; 64.3; 378.6 µg/larva 550.3; 55.7; 414.7 µg/larva | [43] | |
| Chilo partellus | Linalool | |||
| Spodoptera litura | 1,8-Cineole | |||
| Helicoverpa armigera | trans-Anethole | |||
| Carvacrol | ||||
| / | Ephestia kuehniella | α-Pinene | 0.864 μL/mL * 0.490 μL/mL * 0.137 μL/mL * | [44] |
| trans-Anethole | ||||
| Thymol | ||||
| Lippia sidoides | Sitophilus zeamais | EO mix | 35.48 to 118.29 μL/L air | [45] |
| Thymol | 65.00 to 91.23 μL/L air | |||
| p-Cymene | 801.24 to 2188.83 μL/L air | |||
| Thymus vulgaris | Aedes aegypti | Thymol, p-Cymene | 45.73 mg/mL | [32] |
| Salvia officinalis | 1,8-Cineol, α-Thujone | 76.43 mg/mL | ||
| Lippia origanoides | Limonene, p-Cymene, α-Phellandrene | 53.79 mg/mL | ||
| Eucalyptus globulus | Thymol, p-Cymene | 92.55 mg/mL | ||
| Cymbopogon nardus | 1,8-Cineol | 75.85 mg/mL | ||
| Cymbopogon martinii | Citronellal, Citronellol, Geraniol | 114.65 mg/mL | ||
| Lippia alba | Geraniol | 72.34 mg/mL | ||
| Pelargonium graveolens | Carrvone, Limonene, Citronellol | 108.96 mg/mL | ||
| Thymus alternans | (E)-Nerolidol, Linalool, Germacrene D | 156.3 mg/L 103.7 mg/L 221.1 mg/L | [46] | |
| Spodoptera littoralis, Musca domestica, Culex quinquefasciatus | ||||
| Teucrium montanum subsp. jailae | Germacrene D, (E)-Caryophyllene | 56.7 mg/L 154.9 mg/L 180.5 mg/L | ||
| Allium sativum | Diallyl disulfide, Diallyl trisulfide | 0.64% | ||
| Cumimum cyminum | Cuminaldehyde γ-Terpinene, p-Cymene | 3.05% | ||
| Eucalyptus citriodora | β-Pinene | 2.98% | ||
| Eucalyptus dives | Citronellal | 2.03% | ||
| Gaultheria procumbens | Piperitone, α-Phellandrene Methyl salicylate | 1.59% | ||
| Illicum verum | trans-Anethole | 3.02% | ||
| Lavandulla hybrida super | Linalool, Linalyl acetate | 3.41% | ||
| Melaleuca alternifolia | Terpinene-4-ol, γ-Terpinene | 2.86% | ||
| Mentha arvensis | Menthol | 2.27% | ||
| Myristica fragrans | Sitophilus granaries | α-Pinene, α-Thujene, Sabinene, β-Pinene | 3.40% | [47] |
| Ocimum bassilicum spp. basilicum | Estragol, Linalool | 3.14% | ||
| Ocimum sanctum | Eugenol, β-Caryophyllene, Methyl eugenol | 1.77% | ||
| Origanum majorana | α-Pinene, 1,8-Cineole, Camphor, Camphene | 3.04% | ||
| Rosmarinus officinalis CT camphor | α-Pinene, 1,8-Cineole, Camphor, Camphene | 3.72% | ||
| Thymus vulgaris CT geraniol | Geraniol, Geranyl acetate | 2.90% | ||
| Carlina acaulis root | Musca domestica | Carlina oxide | 2.74 µL (male) 5.96 µL (female) | [48] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Devrnja, N.; Milutinović, M.; Savić, J. When Scent Becomes a Weapon—Plant Essential Oils as Potent Bioinsecticides. Sustainability 2022, 14, 6847. https://doi.org/10.3390/su14116847
Devrnja N, Milutinović M, Savić J. When Scent Becomes a Weapon—Plant Essential Oils as Potent Bioinsecticides. Sustainability. 2022; 14(11):6847. https://doi.org/10.3390/su14116847
Chicago/Turabian StyleDevrnja, Nina, Milica Milutinović, and Jelena Savić. 2022. "When Scent Becomes a Weapon—Plant Essential Oils as Potent Bioinsecticides" Sustainability 14, no. 11: 6847. https://doi.org/10.3390/su14116847
APA StyleDevrnja, N., Milutinović, M., & Savić, J. (2022). When Scent Becomes a Weapon—Plant Essential Oils as Potent Bioinsecticides. Sustainability, 14(11), 6847. https://doi.org/10.3390/su14116847