
Effect of Micronutrient and Hormone on Microalgae Growth Assessment for Biofuel Feedstock

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Specific Strains of Microalgae
2.2. Experimental Setup and Growth Strategy
2.2.1. Separate Effects of Micronutrients and Hormone (First Set of Experiments)
2.2.2. Combined Effects of Micronutrients and Hormone (Second Set of Experiments)
2.3. Photobioreactor Setup and Microalgal Growth Analyses
2.4. Dry Weight (for ZAA1)
2.5. DNA Extraction, Polymerase Chain Reaction (PCR) Amplification, and Sequencing
2.6. Determination of Chlorophyll and Carotenoid Contents
2.7. Phylogenetic Tree Construction
Statistical Analysis
3. Results and Discussions
3.1. Isolation and Identification of Microalgae
3.2. PCR Amplification and Sequencing
3.3. Growth Rate of Microalgal Strains under Separate Hormone and Micronutrient Treatments (First Set of Experiments)
3.3.1. Growth Analysis of ZAA1 and ZAA2 Strains
3.3.2. Growth Analysis of ZAA3 and AZH Strains
3.3.3. AZS Strain
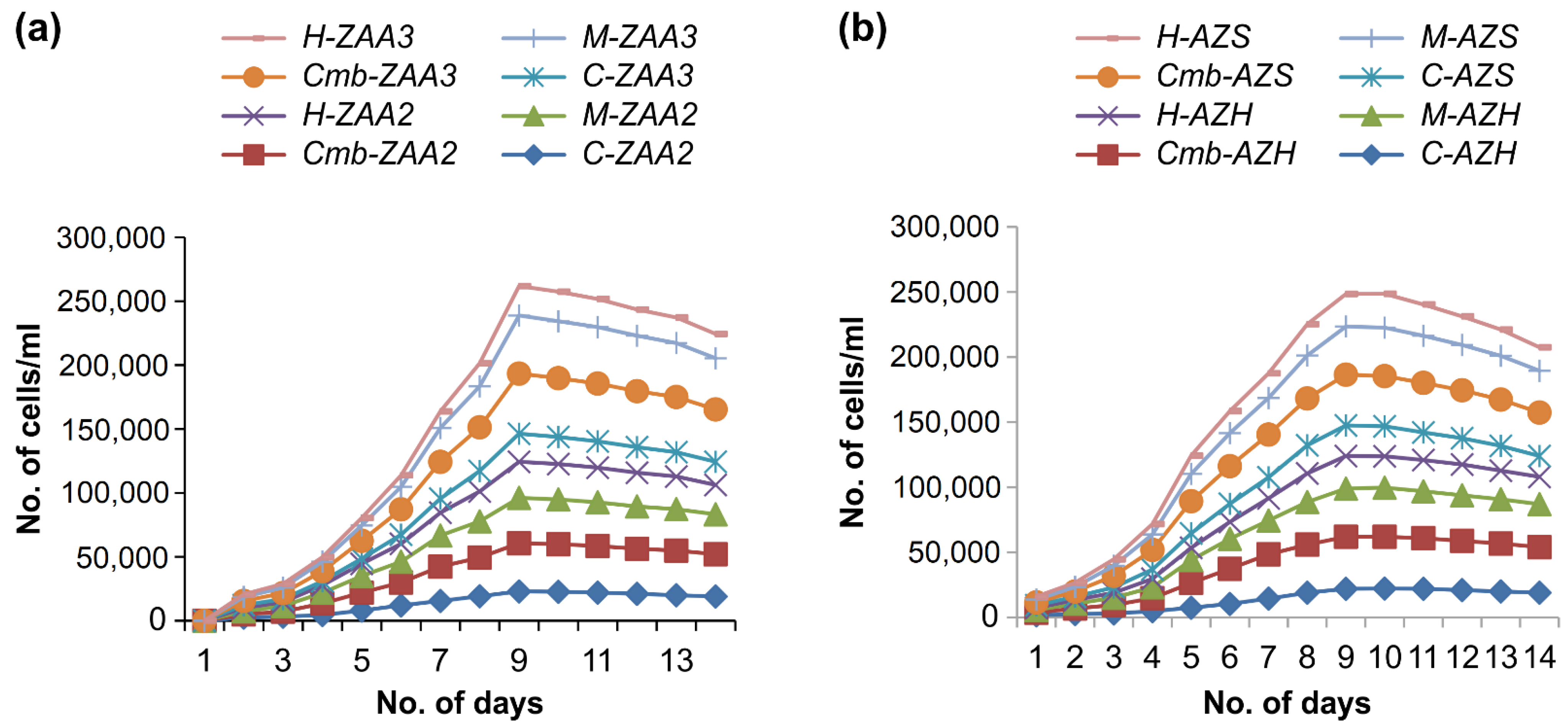
3.4. The Combined Effect of Hormone and Micronutrients (Second Set of Experiments)
3.5. Effects of Individual Treatments with Micronutrients and Hormone on Photosynthetic Pigments
3.5.1. Chlorophyll A
3.5.2. Chlorophyll B
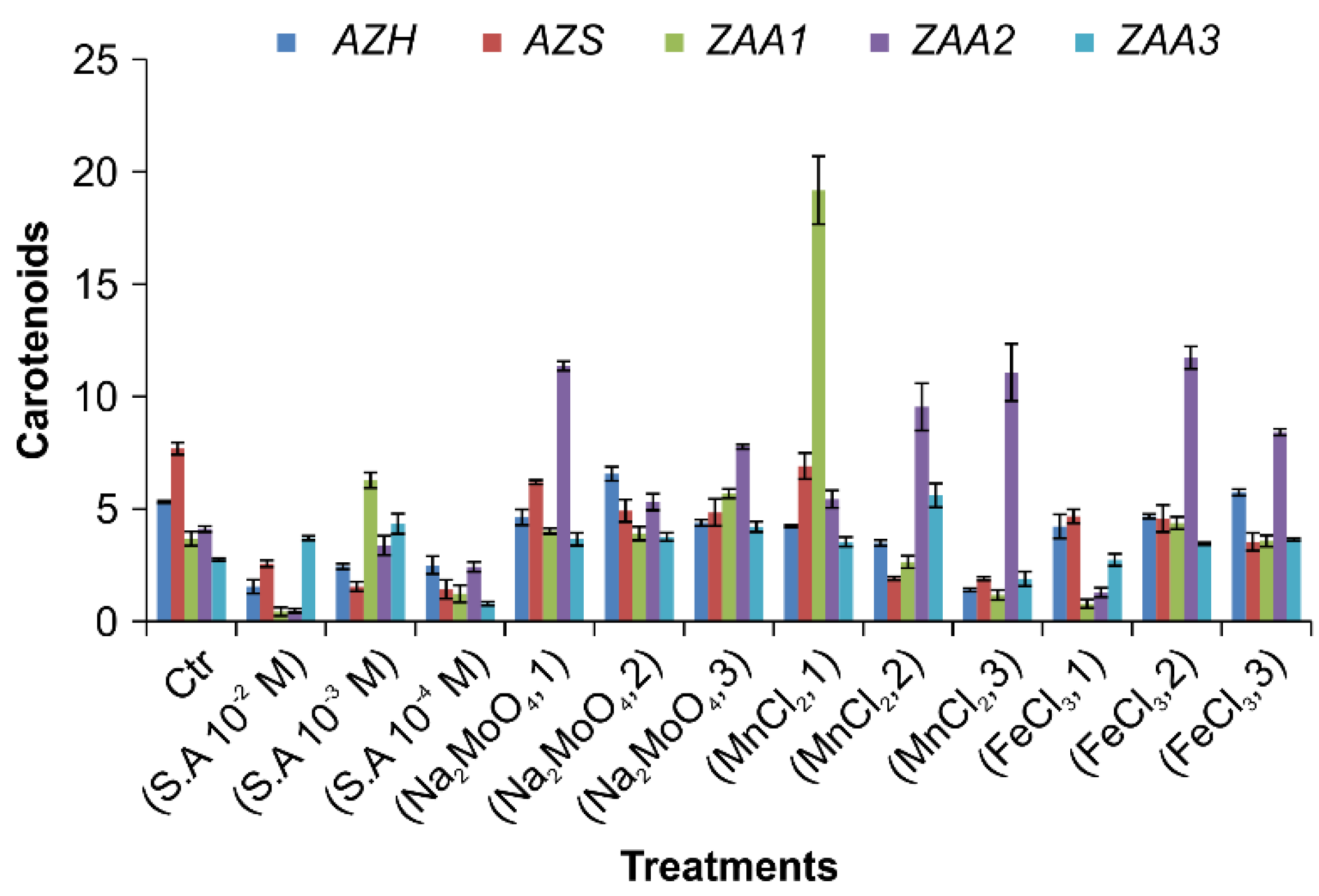
3.5.3. Carotenoids
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Williams, P.J.; Laurens, L.M.L. Microalgae as biodiesel & biomass feedstocks: Review & analysis of the biochemistry, energetics & economics. Energy Environ. Sci. 2010, 3, 554–590. [Google Scholar]
- Fargione, J.; Hill, J.; Tilman, D.; Polasky, S.; Hawthorne, P. Land Clearing and the Biofuel Carbon Debt. Science 2008, 319, 1235–1238. [Google Scholar] [CrossRef] [PubMed]
- Melillo, J.M.; Reilly, J.M.; Kicklighter, D.W.; Gurgel, A.C.; Cronin, T.W.; Paltsev, S.; Felzer, B.S.; Wang, X.; Sokolov, A.P.; Schlosser, C.A. Indirect emissions from biofuels: How important? Science 2009, 326, 1397–1399. [Google Scholar] [CrossRef] [PubMed]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Richmond, A.; Hu, Q. Handbook of Microalgal Culture; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013. [Google Scholar]
- Graham, L.; Graham, J.; Wilcox, L. Algae, 2nd ed.; Benjamin Cummings: San Francisco, CA, USA, 2009. [Google Scholar]
- Lee, R.A.; Lavoie, J.-M. From first- to third-generation biofuels: Challenges of producing a commodity from a biomass of increasing complexity. Anim. Front. 2013, 3, 6–11. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Song, M.; Pei, H.; Hu, W.; Ma, G. Evaluation of the potential of 10 microalgal strains for biodiesel production. Bioresour. Technol. 2013, 141, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Michelon, W.; Da Silva, M.L.B.; Mezzari, M.P.; Pirolli, M.; Prandini, J.M.; Soares, H.M. Effects of Nitrogen and Phosphorus on Biochemical Composition of Microalgae Polyculture Harvested from Phycoremediation of Piggery Wastewater Digestate. Appl. Biochem. Biotechnol. 2016, 178, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Jazzar, S.; Berrejeb, N.; Messaoud, C.; Marzouki, M.N.; Smaali, I. Growth Parameters, Photosynthetic Performance, and Biochemical Characterization of Newly Isolated Green Microalgae in Response to Culture Condition Variations. Appl. Biochem. Biotechnol. 2016, 179, 1290–1308. [Google Scholar] [CrossRef]
- Huesemann, M.; Crowe, B.; Waller, P.; Chavis, A.; Hobbs, S.; Edmundson, S.; Wigmosta, M. A validated model to predict microalgae growth in outdoor pond cultures subjected to fluctuating light intensities and water temperatures. Algal Res. 2016, 13, 195–206. [Google Scholar] [CrossRef]
- Juneja, A.; Ceballos, R.M.; Murthy, G.S. Effects of Environmental Factors and Nutrient Availability on the Biochemical Composition of Algae for Biofuels Production: A Review. Energies 2013, 6, 4607–4638. [Google Scholar] [CrossRef]
- Chen, M.; Tang, H.; Ma, H.; Holland, T.C.; Ng, K.S.; Salley, S.O. Effect of nutrients on growth and lipid accumulation in the green algae Dunaliella tertiolecta. Bioresour. Technol. 2011, 102, 1649–1655. [Google Scholar] [CrossRef] [PubMed]
- Mojaat, M.; Pruvost, J.; Foucault, A.; Legrand, J. Effect of organic carbon sources and Fe2+ ions on growth and β-carotene accumulation by Dunaliella salina. Biochem. Eng. J. 2008, 39, 177–184. [Google Scholar] [CrossRef]
- Chernikova, A.A.; Tsoglin, L.N.; Markelova, A.G.; Zorin, S.N.; Mazo, V.K.; Pronina, N.A. Capacity of Spirulina platensis to accumulate manganese and its distribution in cell. Russ. J. Plant Physiol. 2006, 53, 800–806. [Google Scholar] [CrossRef]
- Araújo, P.H.M.; Santana, J.K.S.; Sassi, R.; da Costa, D.C.; Filho, N.R.A.; Cordeiro, A.M.T.M.; Gondim, A.D.; Santos, N.A. Renewable source hydrocarbons obtaining from microalgae by catalytic deoxygenation. Biomass Convers. Biorefinery 2021, 1–8. [Google Scholar] [CrossRef]
- Manoj, B.S.; Ahlawat, S.; Chavan, M.; Karosiya, A. Successive production of biodiesel and bioethanol feedstock from the Cosmarium sp. Int. J. Chem. Stud. 2018, 6, 550–554. [Google Scholar] [CrossRef]
- Chi, N.T.L.; Duc, P.A.; Mathimani, T.; Pugazhendhi, A. Evaluating the potential of green alga Chlorella sp. for high biomass and lipid production in biodiesel viewpoint. Biocatal. Agric. Biotechnol. 2019, 17, 184–188. [Google Scholar] [CrossRef]
- Kim, H.S.; Park, W.-K.; Lee, B.; Seon, G.; Suh, W.I.; Moon, M.; Chang, Y.K. Optimization of heterotrophic cultivation of Chlorella sp. HS2 using screening, statistical assessment, and validation. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Vo, H.-N.-P.; Bui, X.-T.; Nguyen, T.-T.; Nguyen, D.D.; Dao, T.-S.; Cao, N.-D.-T.; Vo, T.-K.-Q. Effects of nutrient ratios and carbon dioxide bio-sequestration on biomass growth of Chlorella sp. in bubble column photobioreactor. J. Environ. Manag. 2018, 219, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Alishah Aratboni, H.; Rafiei, N.; Garcia-Granados, R.; Alemzadeh, A.; Morones-Ramírez, J.R. Biomass and lipid induction strategies in microalgae for biofuel production and other applications. Microb. Cell Factories 2019, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Yadav, G.; Sekar, M.; Kim, S.-H.; Geo, V.E.; Bhatia, S.K.; Sabir, J.S.; Chi, N.T.L.; Brindhadevi, K.; Pugazhendhi, A. Lipid content, biomass density, fatty acid as selection markers for evaluating the suitability of four fast growing cyanobacterial strains for biodiesel production. Bioresour. Technol. 2021, 325, 124654. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.K.; Sundaram, S.; Patel, A.K.; Kalra, A. Characterization of Seven Species of Cyanobacteria for High-Quality Biomass Production. Arab. J. Sci. Eng. 2017, 43, 109–121. [Google Scholar] [CrossRef]
- Tsavatopoulou, V.D.; Aravantinou, A.F.; Manariotis, I.D. Biofuel conversion of Chlorococcum sp. and Scenedesmus sp. biomass by one-and two-step transesterification. Biomass Convers. Biorefinery 2019, 1–9. [Google Scholar] [CrossRef]
- Rehman, Z.U.; Anal, A.K. Enhanced lipid and starch productivity of microalga (Chlorococcum sp. TISTR 8583) with nitrogen limitation following effective pretreatments for biofuel production. Biotechnol. Rep. 2019, 21, e00298. [Google Scholar] [CrossRef] [PubMed]
- Ruangsomboon, S. Effect of light, nutrient, cultivation time and salinity on lipid production of newly isolated strain of the green microalga, Botryococcus braunii KMITL 2. Bioresour. Technol. 2012, 109, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Andersen, R.A. Algal Culturing Techniques; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Chaichalerm, S.; Pokethitiyook, P.; Yuan, W.; Meetam, M.; Sritong, K.; PugKaew, W.; Kungvansaichol, K.; Kruatrachue, M.; Damrongphol, P. Culture of microalgal strains isolated from natural habitats in Thailand in various enriched media. Appl. Energy 2012, 89, 296–302. [Google Scholar] [CrossRef]
- Hai, T.; Ahlers, H.; Gorenflo, V.; Steinbüchel, A. Axenic cultivation of anoxygenic phototrophic bacteria, cyanobacteria, and microalgae in a new closed tubular glass photobioreactor. Appl. Microbiol. Biotechnol. 2000, 53, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Dupré, C.; Legrand, J.; Grizeau, D. Extracellular hydrocarbon and intracellular lipid accumulation are related to nutrient-sufficient conditions in pH-controlled chemostat cultures of the microalga Botryococcus braunii SAG 30.81. Algal Res. 2016, 17, 244–252. [Google Scholar] [CrossRef]
- Wu, L.F.; Chen, P.C.; Lee, C.M. The effects of nitrogen sources and temperature on cell growth and lipid accumulation of microalgae. Int. Biodeterior. Biodegrad. 2013, 85, 506–510. [Google Scholar] [CrossRef]
- González-Fernández, C.; Mahdy, A.; Ballesteros, I.; Ballesteros, M. Impact of temperature and photoperiod on anaerobic biodegradability of microalgae grown in urban wastewater. Int. Biodeterior. Biodegrad. 2016, 106, 16–23. [Google Scholar] [CrossRef]
- Halim, R.; Harun, R.; Danquah, M.K.; Webley, P.A. Microalgal cell disruption for biofuel development. Appl. Energy 2012, 91, 116–121. [Google Scholar] [CrossRef]
- Agostoni, M.; Lucker, B.F.; Smith, M.A.; Kanazawa, A.; Blanchard, G.J.; Kramer, D.M.; Montgomery, B.L. Competition-based phenotyping reveals a fitness cost for maintaining phycobilisomes under fluctuating light in the cyanobacterium Fremyella diplosiphon. Algal Res. 2016, 15, 110–119. [Google Scholar] [CrossRef]
- Bergmann, P.; Trösch, W. Repeated fed-batch cultivation of Thermosynechococcus elongatus BP-1 in flat-panel airlift photobioreactors with static mixers for improved light utilization: Influence of nitrate, carbon supply and photobioreactor design. Algal Res. 2016, 17, 79–86. [Google Scholar] [CrossRef]
- Blank, C.E.; Hinman, N.W. Cyanobacterial and algal growth on chitin as a source of nitrogen; ecological, evolutionary, and biotechnological implications. Algal Res. 2016, 15, 152–163. [Google Scholar] [CrossRef]
- Coyer, J.A.; Robertson, D.L.; Alberte, R.S. Genetic variability within a population and between diploid/haploid tissue macrocystis pyrifera. J. Phycol. 1994, 30, 545–552. [Google Scholar] [CrossRef]
- Sapp, M.; Schwaderer, A.S.; Wiltshire, K.H.; Hoppe, H.-G.; Gerdts, G.; Wichels, A. Species-Specific Bacterial Communities in the Phycosphere of Microalgae? Microb. Ecol. 2007, 53, 683–699. [Google Scholar] [CrossRef] [PubMed]
- Burja, A.M.; Tamagnini, P.; Bustard, M.T.; Wright, P.C. Identification of the green alga, Chlorella vulgaris (SDC1) using cyanobacteria derived 16S rDNA primers: Targeting the chloroplast. FEMS Microbiol. Lett. 2001, 202, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Li, L.; Cui, J.; Liu, Q.; Ding, Y.; Liu, J. Screening and phylogenetic analysis of lipid-rich microalgae. Algal Res. 2015, 11, 381–386. [Google Scholar] [CrossRef]
- Feng, D.-F.; Doolittle, R.F. Progressive alignment and phylogenetic tree construction of protein sequences. Methods Enzymol. 1990, 183, 375–387. [Google Scholar]
- Song, L.; Qin, J.G.; Su, S.; Xu, J.; Clarke, S.; Shan, Y. Micronutrient Requirements for Growth and Hydrocarbon Production in the Oil Producing Green Alga Botryococcus braunii (Chlorophyta). PLoS ONE 2012, 7, e41459. [Google Scholar] [CrossRef] [PubMed]
- Volland, S.; Bayer, E.; Baumgartner, V.; Andosch, A.; Lütz, C.; Sima, E.; Lütz-Meindl, U. Rescue of heavy metal effects on cell physiology of the algal model system Micrasterias by divalent ions. J. Plant Physiol. 2014, 171, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Fanning, K.; Netzel, M.; Schenk, P.M. Induced carotenoid accumulation in Dunaliella salina and Tetraselmis suecica by plant hormones and UV-C radiation. Appl. Microbiol. Biotechnol. 2015, 99, 9407–9416. [Google Scholar] [CrossRef] [PubMed]
- Czerpak, R.; Bajguz, A.; Gromek, M.; Kozłowska, G.; Nowak, I. Activity of salicylic acid on the growth and biochemism of Chlorella vulgaris Beijerinck. Acta Physiol. Plant. 2002, 24, 45–52. [Google Scholar] [CrossRef]
- Mc Gee, D.; Archer, L.; Fleming, G.T.; Gillespie, E.; Touzet, N. The effect of nutrient and phytohormone supplementation on the growth, pigment yields and biochemical composition of newly isolated microalgae. Process. Biochem. 2020, 92, 61–68. [Google Scholar] [CrossRef]
- Battah, M.; El-Ayoty, Y.; Abomohra, A.E.-F.; El-Ghany, S.A.; Esmael, A. Effect of Mn2+, Co2+ and H2O2 on biomass and lipids of the green microalga Chlorella vulgaris as a potential candidate for biodiesel production. Ann. Microbiol. 2014, 65, 155–162. [Google Scholar] [CrossRef]
- Pirson, A.; Bergmann, L. Manganese requirement and carbon source in Chlorella. Nature 1955, 176, 209–210. [Google Scholar] [CrossRef]
- Bruce, M. Organo-Transition Metal Chemistry-A Guide to the Literature 1950–1970. In Advances in Organometallic Chemistry; Elsevier: Amsterdam, The Netherlands, 1972; Volume 10, pp. 273–346. [Google Scholar]
- Clarkson, D.T.; Hanson, J.B. The Mineral Nutrition of Higher Plants. Annu. Rev. Plant Physiol. 1980, 31, 239–298. [Google Scholar] [CrossRef]
- Campbell, L.C.; Nable, R.O. Physiological Functions of Manganese in Plants; Springer: New York, NY, USA, 1988; pp. 139–154. [Google Scholar]
- Rains, D. Mineral metabolism. In Plant Biochemistry; Bonner, J., Varner, J.E., Eds.; Academic Press: New York, NY, USA, 1976. [Google Scholar]
- Pirson, A.; Tichy, C.; Wilhelmi, G. Stoffwechsel und mineralsalzernährung Einzelliger Grünalgen. Planta 1951, 40, 199–253. [Google Scholar] [CrossRef]
- Garnham, G.W.; Codd, G.A.; Gadd, G.M. Effect of nutritional regime on accumulation of cobalt, manganese and zinc by green microalgae. FEMS Microbiol. Lett. 1992, 98, 45–50. [Google Scholar] [CrossRef][Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Methods | Chemicals | Concentrations | |||
|---|---|---|---|---|---|
| Hormone | Salicylic acid (S.A) | 10−2 M | 10−4 M | 10−6 M | |
| Micronutrient | Sodium molybdenum oxide (Na2MoO4) | 0.40 µM | 0.60 µM | 0.80 µM | |
| Manganese chloride (MnCl2) | 0.50 µM | 0.70 µM | 0.90 µM | ||
| Iron chloride (FeCl3) | 0.20 µM | 0.40 µM | 0.60 µM | ||
| Sampling Site and Conditions | Name of Species | Morphological Characteristics | Accession Number | Close Relatives | Similarity Index | Microscopic Image |
|---|---|---|---|---|---|---|
| University lawn (Uni-L), pH 6.8, temperature 24 °C | (AZH) Chlorella sp. | Unicellular, bilateral oval shape, pyronide present | MF114595 | JN559374.1 | 88% |  |
| Bio-park (Bp), pH 6.3, temperature 24 °C | (AZS) Chlorococcum sp. | Unicellular, circular shape, cup-shaped chloroplast | MF114593 | JX221788.1 | 92% |  |
| Institute of Management Science (IMS), pH 6.9, temperature 24 °C | (ZAA1) Phormidium sp. | Fiber-like structure grows with a long spindle form, unicellular | MF140241 | MF140241.1 | 100% |  |
| Rice field (Rf), pH 8.1, temperature 28 °C | (ZAA3) Cosmarium sp. | Tetrachotomous branching, perpendicular to the surface of colony, unicellular | MF114594 | KJ877699.1 | 86% |  |
| Old Shujabad (OS), pH 7.4, temperature 30 °C | (ZAA2) Chlorella sp. | Unicellular, exists in colony, cup-shaped chloroplast | MF114592 | GU291326.1 | 84% |  |
| Measurements | Strains | SA 10−2 M | S.A 10−4 M | SA 10−6 M | NaMo4 0.4 µM | NaMoO4 0.6 µM | NaMoO4 0.8 µM | MnCl2 0.5 µM | MnCl2 0.7 µM | MnCl2 0.9 µM | FeCl3 0.2 µM | FeCl3 0.4 µM | FeCl3 0.6 µM |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chlorophyll A (mg/g) | ZAA1 | 0.1171 ± 0.0201 | 0.1272 ± 0.017 | 0.1757 ± 0.0186 | 0.2477 ± 0.0214 | 0.3236 ± 0.0281 | 0.3875 ± 0.0124 | 0.1412 ± 0.0228 | 0.1817 ± 0.0194 | 0.1817 ± 0.0194 | 0.249 ± 0.0257 | 0.4447 ± 0.0244 | 0.551 ± 0.0203 |
| ZAA2 | 0.1413 ± 0.0202 | 0.1914 ± 0.0153 | 0.211 ± 0.0217 | 0.3352 ± 0.0222 | 0.3993 ± 0.0239 | 0.323 ± 0.0115 | 0.2392 ± 0.0159 | 0.1796 ± 0.0153 | 0.2065 ± 0.0158 | 0.3737 ± 0.0103 | 0.4433 ± 0.0107 | 0.3389 ± 0.0281 | |
| ZAA3 | 0.1627 ± 0.0204 | 0.25 ± 0.0122 | 0.2091 ± 0.0197 | 0.3584 ± 0.0145 | 0.4067 ± 0.0167 | 0.2978 ± 0.0181 | 0.2624 ± 0.014 | 0.198 ± 0.0213 | 0.1835 ± 0.0113 | 0.2041 ± 0.0102 | 0.5394 ± 0.0199 | 0.4787 ± 0.016 | |
| AZH | 0.1851 ± 0.01 | 0.2088 ± 0.033 | 0.2361 ± 0.0183 | 0.3459 ± 0.0086 | 0.503 ± 0.0173 | 0.4655 ± 0.0168 | 0.2989 ± 0.0125 | 0.4025 ± 0.00918 | 0.334 ± 0.012 | 0.4577 ± 0.0219 | 0.5794 ± 0.0173 | 0.4707 ± 0.017 | |
| AZS | 0.2088 ± 0.0111 | 0.2429 ± 0.0139 | 0.298 ± 0.0102 | 0.4025 ± 0.0121 | 0.4859 ± 0.0133 | 0.4486 ± 0.0265 | 0.284 ± 0.022 | 0.3367 ± 0.0228 | 0.3865 ± 0.0201 | 0.4576 ± 0.0291 | 0.5074 ± 0.046 | 0.3252 ± 0.0207 | |
| Chlorophyll B (mg/g) | ZAA1 | 0.09375 ± 0.01037 | 0.09935 ± 0.024181 | 0.315719 ± 0.023256 | 0.73211 ± 0.05258 | 0.804814 ± 0.015297 | 0.3677 ± 0.03404 | 0.13488 ± 0.03303 | 0.20505 ± 0.04236 | 0.65798 ± 0.0438 | 0.57058 ± 0.02651 | 0.87368 ± 0.02367 | 0.61523 ± 0.0308 |
| ZAA2 | 0.08485 ± 0.02028 | 0.186017 ± 0.01902 | 0.091736 ± 0.025541 | 0.05354 ± 0.01917 | 0.463006 ± 0.026463 | 0.22385 ± 0.02105 | 0.51141 ± 0.03922 | 0.10287 ± 0.03134 | 0.12946 ± 0.01461 | 0.08683 ± 0.01698 | 0.13385 ± 0.03812 | 0.25741 ± 0.02391 | |
| ZAA3 | 0.79518 ± 0.03918 | 0.664446 ± 0.02389 | 0.842581 ± 0.048591 | 0.63124 ± 0.02654 | 0.717881 ± 0.021089 | 0.616131 ± 0.06543 | 0.20564 ± 0.04097 | 0.18543 ± 0.03821 | 0.83656 ± 0.02893 | 0.41679 ± 0.01686 | 0.78801 ± 0.03884 | 0.64316 ± 0.02728 | |
| AZH | 0.77487 ± 0.02113 | 0.471821 ± 0.021832 | 0.6489 ± 0.103712 | 0.47316 ± 0.04979 | 0.33621 ± 0.03459 | 0.35468 ± 0.02349 | 0.66925 ± 0.00989 | 0.27505 ± 0.0216 | 0.71086 ± 0.01428 | 0.57291 ± 0.03043 | 0.38467 ± 0.02769 | 0.59745 ± 0.01642 | |
| AZS | 0.48207 ± 0.01944 | 0.548749 ± 0.024679 | 0.427653 ± 0.022751 | 0.18158 ± 0.02913 | 0.328115 ± 0.019466 | 0.30226 ± 0.02619 | 0.30246 ± 0.03064 | 0.48315 ± 0.02577 | 0.471 ± 0.03469 | 0.47072 ± 0.06262 | 0.58966 ± 0.03006 | 0.77913 ± 0.07585 | |
| Carotenoids (mg/g) | ZAA1 | 0.4376 ± 0.18961 | 6.279645 ± 0.342229 | 1.231069 ± 0.382523 | 4.0227 ± 0.11885 | 3.910981 ± 0.304809 | 5.68487 ± 0.2053 | 19.1798 ± 1.50787 | 2.64593 ± 0.2769 | 1.1772 ± 0.22342 | 0.78878 ± 0.19189 | 4.37719 ± 0.27326 | 3.57572 ± 0.24983 |
| ZAA2 | 0.48769 ± 0.09245 | 3.380871 ± 0.428483 | 2.431123 ± 0.219307 | 11.3628 ± 0.36715 | 5.309117 ± 0.367158 | 7.76896 ± 0.09943 | 5.43946 ± 0.39154 | 9.54366 ± 1.05015 | 11.0765 ± 1.27241 | 1.29222 ± 0.20923 | 11.733 ± 0.50088 | 8.41567 ± 0.14368 | |
| ZAA3 | 3.70892 ± 0.0974 | 4.344771 ± 0.449426 | 0.795548 ± 0.085358 | 3.65585 ± 0.28242 | 3.761879 ± 0.180967 | 4.20928 ± 0.23091 | 3.54076 ± 0.21322 | 5.60778 ± 0.53143 | 1.8951 ± 0.32474 | 2.73708 ± 0.26823 | 3.45322 ± 0.0565 | 3.64747 ± 0.06035 | |
| AZH | 1.54641 ± 0.30667 | 2.447025 ± 0.121471 | 2.505814 ± 0.392535 | 4.63407 ± 0.34712 | 6.567664 ± 0.310251 | 4.38534 ± 0.14101 | 4.23357 ± 0.05658 | 3.4905 ± 0.1282 | 1.40034 ± 0.07346 | 4.22837 ± 0.53237 | 4.67849 ± 0.1003 | 5.73855 ± 0.14241 | |
| AZS | 2.57232 ± 0.14578 | 1.550937 ± 0.212485 | 1.430827 ± 0.416397 | 6.18919 ± 0.09059 | 4.925682 ± 0.497577 | 4.84973 ± 0.61001 | 6.91673 ± 0.57915 | 1.91282 ± 0.07618 | 1.90541 ± 0.07665 | 4.67638 ± 0.31029 | 4.57829 ± 0.31029 | 3.54275 ± 0.39245 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslam, A.; Rasul, S.; Bahadar, A.; Hossain, N.; Saleem, M.; Hussain, S.; Rasool, L.; Manzoor, H. Effect of Micronutrient and Hormone on Microalgae Growth Assessment for Biofuel Feedstock. Sustainability 2021, 13, 5035. https://doi.org/10.3390/su13095035
Aslam A, Rasul S, Bahadar A, Hossain N, Saleem M, Hussain S, Rasool L, Manzoor H. Effect of Micronutrient and Hormone on Microalgae Growth Assessment for Biofuel Feedstock. Sustainability. 2021; 13(9):5035. https://doi.org/10.3390/su13095035
Chicago/Turabian StyleAslam, Ayesha, Sumaira Rasul, Ali Bahadar, Nazia Hossain, Muhammad Saleem, Sabir Hussain, Lubna Rasool, and Hamid Manzoor. 2021. "Effect of Micronutrient and Hormone on Microalgae Growth Assessment for Biofuel Feedstock" Sustainability 13, no. 9: 5035. https://doi.org/10.3390/su13095035
APA StyleAslam, A., Rasul, S., Bahadar, A., Hossain, N., Saleem, M., Hussain, S., Rasool, L., & Manzoor, H. (2021). Effect of Micronutrient and Hormone on Microalgae Growth Assessment for Biofuel Feedstock. Sustainability, 13(9), 5035. https://doi.org/10.3390/su13095035