
Locally Available Organic Waste for Counteracting Strawberry Decline in a Mountain Specialized Cropping Area

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Preliminary Soil Health Evaluation by Greenhouse Bio-Assay
2.3. Field Trial
2.3.1. Pre-Plant Treatments and Field Trial
2.3.2. Evaluation of Strawberry Production
2.4. Soil Sampling and Molecular Analysis
2.4.1. Rhizosphere Soil Sampling
2.4.2. Soil DNA Extraction
2.4.3. Quantitative PCR (qPCR)
2.4.4. Bacterial Community DGGE Fingerprinting (PCR-DGGE)
2.5. Statistical Analysis
3. Results
3.1. Original Soil Features
3.2. Soil Healthevalution by Greenhouse Bio-Assay
3.3. Strewberry Yield in Response to Soil Treatment in Pre-Plantof Field Trial
3.3.1. Strawberry Yield Response
3.3.2. Correlation between Crop Yield and Quantitative Microbial Changes in Soil
3.3.3. Qualitative Changes of Soil Bacterial Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Broeckling, C.D.; Broz, A.K.; Bergelson, J.; Manter, D.K.; Vivanco, J.M. Root exudates regulate soil fungal community composition and diversity. Appl. Environ. Microbiol. 2008, 74, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Eisenhauer, N.; Lanoue, A.; Strecker, T.; Scheu, S.; Steinauer, K.; Thakur, M.P.; Mommer, L. Root biomass and exudates link plant diversity with soil bacterial and fungal biomass. Sci. Rep. 2017, 7, 44641. [Google Scholar] [CrossRef] [PubMed]
- Lemanceau, P.; Maron, P.A.; Mazurier, S.; Mougel, C.; Pivato, B.; Plassart, P.; Ranjard, L.; Revellin, C.; Tardy, V.; Wipf, D. Understanding and managing soil biodiversity: A major challenge in agroecology. Agron. Sustain. Dev. 2014, 35, 67–81. [Google Scholar] [CrossRef]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for Management of Soilborne Diseases in Crop Production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef]
- Koike, S.T.; Gordon, T.R.; Daugovish, O.; Ajwa, H.; Bolda, M.; Subbarao, K. Recent Developments on Strawberry Plant Collapse Problems in California Caused by Fusarium and Macrophomina. Int. J. Fruit Sci. 2013, 13, 76–83. [Google Scholar] [CrossRef]
- López-Medina, J.; López-Aranda, J.M.; Medina, J.J.; Miranda, L.; Flores, F. Chemical and non-chemical alternatives to methyl bromide fumigation of soil for strawberry production. J. Hortic. Sci. Biotechnol. 2003, 78, 597–604. [Google Scholar] [CrossRef]
- Pinkerton, J.N.; Ivors, K.L.; Reeser, P.W.; Bristow, P.R.; Windom, G.E. The use of soil solarization for the management of soilborne plant pathogens in strawberry and red raspberry production. Plant Dis. 2002, 86, 645–651. [Google Scholar] [CrossRef]
- Shennan, C.; Muramoto, J.; Koike, S.; Baird, G.; Fennimore, S.; Samtani, J.; Bolda, M.; Dara, S.; Daugovish, O.; Lazarovits, G.; et al. Anaerobic soil disinfestation is an alternative to soil fumigation for control of some soilborne pathogens in strawberry production. Plant Pathol. 2018, 67, 51–66. [Google Scholar] [CrossRef]
- Camprubí, A.; Estaún, V.; Bakali, M.A.; Garcia-Figueres, F.; Calvet, C. Alternative strawberry production using solarization, metham sodium and beneficial soil microbes as plant protection methods. Agron. Sustain. Dev. 2007, 27, 179–184. [Google Scholar] [CrossRef]
- You, M.P.; Sivasithamparam, K. Changes in microbial populations of an avocado plantation mulch suppressive of Phytophthora cinnamomi. Appl. Soil Ecol. 1995, 2, 33–43. [Google Scholar] [CrossRef]
- Mazzola, M. Mechanisms of natural soil suppressiveness to soilborne diseases. Antonie Van Leeuwenhoek 2002, 81, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Bongiorno, G.; Postma, J.; Bünemann, E.K.; Brussaard, L.; de Goede, R.G.M.; Mäder, P.; Tamm, L.; Thuerig, B. Soil suppressiveness to Pythium ultimum in ten European long-term field experiments and its relation with soil parameters. Soil Biol. Biochem. 2019, 133, 174–187. [Google Scholar] [CrossRef]
- Bonilla, N.; Cazorla, F.M.; Martínez-Alonso, M.; Hermoso, J.M.; González-Fernández, J.J.; Gaju, N.; Landa, B.B.; de Vicente, A. Organic amendments and land management affect bacterial community composition, diversity and biomass in avocado crop soils. Plant Soil 2012, 357, 215–226. [Google Scholar] [CrossRef]
- Franke-Whittle, I.H.; Juárez, M.F.D.; Insam, H.; Schweizer, S.; Naef, A.; Topp, A.R.; Kelderer, M.; Rühmer, T.; Baab, G.; Henfrey, J.; et al. Performance evaluation of locally available composts to reduce replant disease in apple orchards of central Europe. Renew. Agric. Food Syst. 2020, 34, 543–557. [Google Scholar] [CrossRef]
- Wang, S.Y.; Lin, S.S. Composts as soil supplement enhanced plant growth and fruit quality of strawberry. J. Plant Nutr. 2002, 25, 2243–2259. [Google Scholar] [CrossRef]
- Lloyd, M.; Kluepfel, D.; Gordon, T. Evaluation of Four Commercial Composts on Strawberry Plant Productivity and Soil Characteristics in California. Int. J. Fruit Sci. 2016, 16, 84–107. [Google Scholar] [CrossRef]
- Millner, P.D.; Ringer, C.E.; Maas, J.L. Suppression of strawberry root disease with animal manure composts. Compost Sci. Util. 2004, 12, 298–307. [Google Scholar] [CrossRef]
- Fang, X.; You, M.P.; Barbetti, M.J. Reduced severity and impact of Fusarium wilt on strawberry by manipulation of soil pH, soil organic amendments and crop rotation. Artic. Eur. J. Plant Pathol. 2012, 134, 619–629. [Google Scholar] [CrossRef]
- Fagerström, A.; Seadi, A.; Rasi, T.; Briseid, S. The Role of Anaerobic Digestion and Biogas in the Circular Economy; IEA Bioenergy: Cork, Ireland, 2018; Volume 37, ISBN 978-1-910154-46-5. [Google Scholar]
- European Union. Directive 2009/28/EC of the European Parliament and of the Council of 23 April 2009 on the Promotion of the Use of Energy from Renewable Sources and Amending and Subsequently Repealing Directives 2001/77/EC and 2003/30/EC; European Commission: Bruxelles, Belgium, 2009.
- Barzee, T.J.; Edalati, A.; El-Mashad, H.; Wang, D.; Scow, K.; Zhang, R. Digestate biofertilizers support similar or higher tomato yields and quality than mineral fertilizer in a subsurface drip fertigation system. Front. Sustain. Food Syst. 2019, 3, 58. [Google Scholar] [CrossRef]
- Tambone, F.; Scaglia, B.; D’Imporzano, G.; Schievano, A.; Orzi, V.; Salati, S.; Adani, F. Assessing amendment and fertilizing properties of digestates from anaerobic digestion through a comparative study with digested sludge and compost. Chemosphere 2010, 81, 577–583. [Google Scholar] [CrossRef]
- Podmirseg, S.M.; Waldhuber, S.; Knapp, B.A.; Insam, H.; Goberna, M. Robustness of the autochthonous microbial soil community after amendment of cattle manure or its digestate. Biol. Fertil. Soils 2019, 55, 565–576. [Google Scholar] [CrossRef]
- Rubel, F.; Brugger, K.; Haslinger, K.; Auer, I. The climate of the European Alps: Shift of very high resolution Köppen-Geiger climate zones 1800–2100. Meteorol. Z. 2017, 26, 115–125. [Google Scholar] [CrossRef]
- Manici, L.M.; Caputo, F.; Rossi, A.; Topp, A.R.; Zago, M.; Kelderer, M. Thermal disturbance of fertile soils to search for new biological control options in strawberry crops affected by yield decline. Biol. Control 2018, 126, 65–73. [Google Scholar] [CrossRef]
- Hoffmann, G. Methodenbuch, Band 1, Die Untersuchung von Böden, vierte Auflage,1. Teillieferung, 2. Teillieferung 1997; VDLUFA-Verlag: Darmstadt, Germany, 1997. [Google Scholar]
- Tessmer, M.A.; Appezzato-da-Glória, B.; Antoniolli, L.R. Influence of growing sites and physicochemical features on the incidence of lenticel breakdown in “Gala” and “Galaxy” apples. Sci. Hortic. 2016, 205, 119–126. [Google Scholar] [CrossRef]
- Caputo, F.; Nicoletti, F.; De Luca Picione, F.; Manici, L.M. Rhizospheric changes of fungal and bacterial communities in relation to soil health of multi-generation apple orchards. Biol. Control 2015, 88, 8–17. [Google Scholar] [CrossRef]
- Alaniz, S.; León, M.; Vicent, A.; García-Jiménez, J.; Abad-Campos, P.; Armengol, J. Characterization of Cylindrocarpon species associated with black foot disease of grapevine in Spain. Plant Dis. 2007, 91, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Widmer, F.; Seidler, R.J.; Gillevet, P.M.; Watrud, L.S.; Di Giovanni, G.D. A highly selective PCR protocol for detecting 16S rRNA genes of the genus Pseudomonas (sensu stricto) in environmental samples. Appl. Environ. Microbiol. 1998, 64, 2545–2553. [Google Scholar] [CrossRef]
- El Fantroussi, S.; Verschuere, L.; Verstraete, W.; Top, E.M. Effect of phenylurea herbicides on soil microbial communities estimated by analysis of 16S rRNA gene fingerprints and community-level physiological profiles. Appl. Environ. Microbiol. 1999, 65, 982–988. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar] [CrossRef]
- Zdruli, P.; Jones, R.J.A.; Montanarella, L. Organic Matter in the Soils of Southern Europe; Office for Official Publications of the European Communities: Luxembourg, 2004; Volumw EUR 21083.
- Saxton, K.E.; Rawls, W.J.; Romberger, J.S.; Papendick, R.I. Estimating Generalized Soil-water Characteristics from Texture1. Soil Sci. Soc. Am. J. 1986, 50, 1031. [Google Scholar] [CrossRef]
- Alvarenga, P.; Palma, P.; Mourinha, C.; Farto, M.; Dôres, J.; Patanita, M.; Cunha-Queda, C.; Natal-da-Luz, T.; Renaud, M.; Sousa, J.P. Recycling organic wastes to agricultural land as a way to improve its quality: A field study to evaluate benefits and risks. Waste Manag. 2017, 61, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Manici, L.M.; Caputo, F.; Baruzzi, G. Additional experiences to elucidate the microbial component of soil suppressiveness towards strawberry black root rot complex. Ann. Appl. Biol. 2005, 146, 421–431. [Google Scholar] [CrossRef]
- Sung, J.-M.Y.-N.S.-S. Identification of Cylindrocarpon destructans Associated with Root Rot Disease of Strawberry. Korean J. Mycol. 1985, 13, 179–183. [Google Scholar]
- Weber, R.W.S.; Entrop, A.P. Dactylonectria torresensis als wichtigste Komponente der Schwarzen Wurzelfäule an Erdbeeren und Himbeeren in Norddeutschland. Erwerbs-Obstbau 2017, 59, 157–169. [Google Scholar] [CrossRef]
- Van der Voort, M.; Kempenaar, M.; van Driel, M.; Raaijmakers, J.M.; Mendes, R. Impact of soil heat on reassembly of bacterial communities in the rhizosphere microbiome and plant disease suppression. Ecol. Lett. 2016, 19, 375–382. [Google Scholar] [CrossRef] [PubMed]
- The Council of European Communities. Council Directive of 12 December 1991 Concerning the Protection of Waters against Pollution Caused by Nitrates from Agricultural Sources (91/676/EEC); European Commission: Bruxelles, Belgium, 1991.
- De Corato, U. Soil microbiome manipulation gives new insights in plant disease-suppressive soils from the perspective of a circular economy: A critical review. Sustainability 2021, 13, 1688. [Google Scholar] [CrossRef]
- Pérez-Piqueres, A.; Edel-Hermann, V.; Alabouvette, C.; Steinberg, C. Response of soil microbial communities to compost amendments. Soil Biol. Biochem. 2006, 38, 460–470. [Google Scholar] [CrossRef]
- Craft, C.M.; Nelson, E.B. Microbial properties of composts that suppress damping-off and root rot of creeping bentgrass caused by Pythium graminicola. Appl. Environ. Microbiol. 1996, 62, 1550–1557. [Google Scholar] [CrossRef] [PubMed]
- Expósito, R.G.; de Bruijn, I.; Postma, J.; Raaijmakers, J.M. Current insights into the role of Rhizosphere bacteria in disease suppressive soils. Front. Microbiol. 2017, 8, 2529. [Google Scholar] [CrossRef]
- De Souza, J.T.; Weller, D.M.; Raaijmakers, J.M. Frequency, diversity, and activity of 2,4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. in Dutch take-all decline soils. Phytopathology 2003, 93, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Chin-A-Woeng, T.F.C.; Bloemberg, G.V.; Lugtenberg, B.J.J. Phenazines and their role in biocontrol by Pseudomonas bacteria. New Phytol. 2003, 157, 503–523. [Google Scholar] [CrossRef]
- Manici, L.M.; Caputo, F.; Cappelli, G.A.; Ceotto, E. Can repeated soil amendment with biogas digestates increase soil suppressiveness toward non-specific soil-borne pathogens in agricultural lands? Renew. Agric. Food Syst. 2020, 1–12. [Google Scholar] [CrossRef]
- Barłóg, P.; Hlisnikovský, L.; Kunzová, E. Effect of Digestate on Soil Organic Carbon and Plant-Available Nutrient Content Compared to Cattle Slurry and Mineral Fertilization. Agronomy 2020, 10, 379. [Google Scholar] [CrossRef]
- UEE Directive 2008/98/EC on Waste and Repealing Certain Directives. Available online: https://www.bmu.de/en/law/directive-200898ec-on-waste-and-repealing-certain-directives/ (accessed on 9 February 2021).
- Nkoa, R. Agricultural benefits and environmental risks of soil fertilization with anaerobic digestates: A review. Agron. Sustain. Dev. 2014, 34, 473–492. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Texture 1 | SOM (%) | pH | C/N | Mineral N 2 (mg kg−1) | Olsen-P (mg kg−1) |
|---|---|---|---|---|---|
| silt-loam | 3.3 | 6.9 | 7 | 8 | 28 |
| Fungal Species | r | Plant Relationship |
|---|---|---|
| Rhizoctonia sp. AG-A & AG-G | 0.24 | Neutral 1 |
| F. oxysporum | −0.06 | neutral |
| F. solani | 0.64 | beneficial |
| F. acuminatum | 0.71 | beneficial |
| Pestalotia longisetula | −0.72 | pathogenic |
| Cadophora sp. | 0.51 | beneficial |
| Mucor hiemalis | 0.34 | neutral |
| Cylindrocarpon-like fungi | −0.78 | pathogenic |
| Acremonium sp. | 0.03 | neutral |
| Paecilomyces lilacinus | −0.34 | neutral |
| Penicillum spp | 0.34 | neutral |
| Pythium sp. | −0.62 | pathogenic |
| Factors | DF | p-Value |
|---|---|---|
| Year (2019–2020) | 1 | Ns 1 |
| Treatment | 3 | 0.0007 |
| Y × T | 3 | 0.0002 |
| Year | count | Average (g) |
| 2019 | 12 | 134.6 2 A |
| 2020 | 12 | 152.4 A |
| Citric Acid (%) | Sugar (Brix) | Firmness (Durofel Index) | Color Index | |
|---|---|---|---|---|
| 2019–2020 | * 1 | *** | *** | *** |
| 2019 | ||||
| Untreated | 70 | 6.77 | 37.23 | 74.51 |
| Dazomet | 82 | 6.93 | 35.50 | 70.74 |
| Compost | 73 | 6.33 | 34.53 | 79.97 |
| Digestate | 81 | 7.24 | 35.23 | 85.45 |
| Total mean | 77 | 6.82 | 35.63 | 77.67 |
| ns | ns | ns | ns | |
| 2020 | ||||
| Untreated | 62 | 9.03 | 43.97 | 49.63 |
| Dazomet | 66 | 7.96 | 42.37 | 48.21 |
| Compost | 70 | 8.72 | 41.40 | 49.88 |
| Digestate | 77 | 9.63 | 48.77 | 48.14 |
| Total mean | 69 | 8.83 | 44.13 | 48.97 |
| ns | ns | ns | ns |
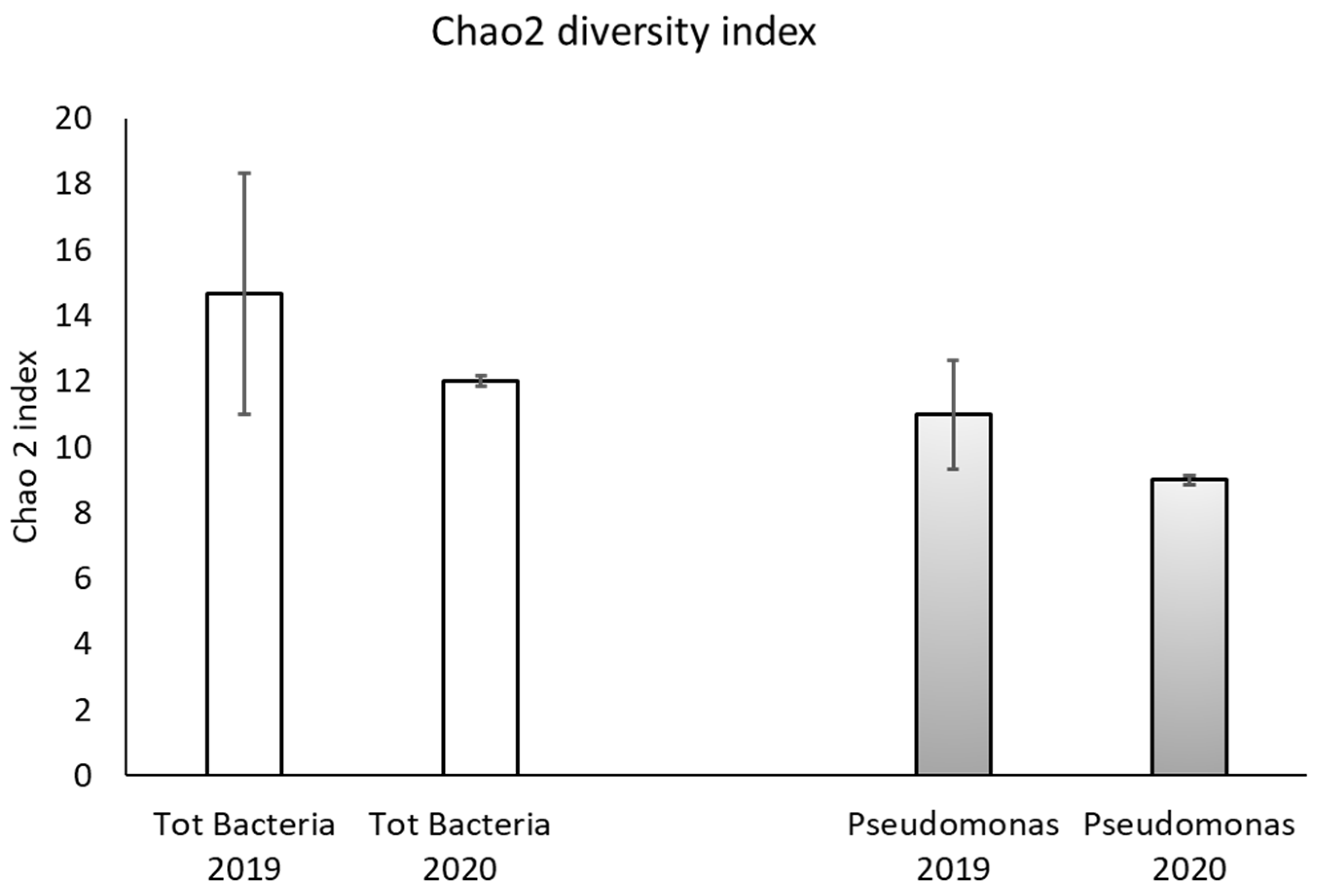
| 2019 | 2020 | |
|---|---|---|
| Total bacteria | ** | ns |
| Pseudmonas spp. | * | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soppelsa, S.; Manici, L.M.; Caputo, F.; Zago, M.; Kelderer, M. Locally Available Organic Waste for Counteracting Strawberry Decline in a Mountain Specialized Cropping Area. Sustainability 2021, 13, 3964. https://doi.org/10.3390/su13073964
Soppelsa S, Manici LM, Caputo F, Zago M, Kelderer M. Locally Available Organic Waste for Counteracting Strawberry Decline in a Mountain Specialized Cropping Area. Sustainability. 2021; 13(7):3964. https://doi.org/10.3390/su13073964
Chicago/Turabian StyleSoppelsa, Sebastian, Luisa Maria Manici, Francesco Caputo, Massimo Zago, and Markus Kelderer. 2021. "Locally Available Organic Waste for Counteracting Strawberry Decline in a Mountain Specialized Cropping Area" Sustainability 13, no. 7: 3964. https://doi.org/10.3390/su13073964
APA StyleSoppelsa, S., Manici, L. M., Caputo, F., Zago, M., & Kelderer, M. (2021). Locally Available Organic Waste for Counteracting Strawberry Decline in a Mountain Specialized Cropping Area. Sustainability, 13(7), 3964. https://doi.org/10.3390/su13073964