
Sustainable Agriculture: Nutritional Benefits of Wheat–Soybean and Maize–Sunflower Associations for Hibernation and Reproduction of Endangered Common Hamsters


Abstract
1. Introduction
2. Materials and Methods
2.1. Selection of Supplemental Crops Based on Consultation with Farmers
2.2. Study Species and Housing Conditions
2.3. Diet Composition
2.4. Macronutrient Content of the Diets
2.5. Hamster Body Mass, Food Intake and Food Preferences
2.6. Geometrical Representation of Nutrient Intake
2.7. Activity Index
2.8. Reproductive Success and Litter Size
2.9. Data Analyses
2.10. Ethics
3. Results
3.1. Macronutrient Content of the Diets
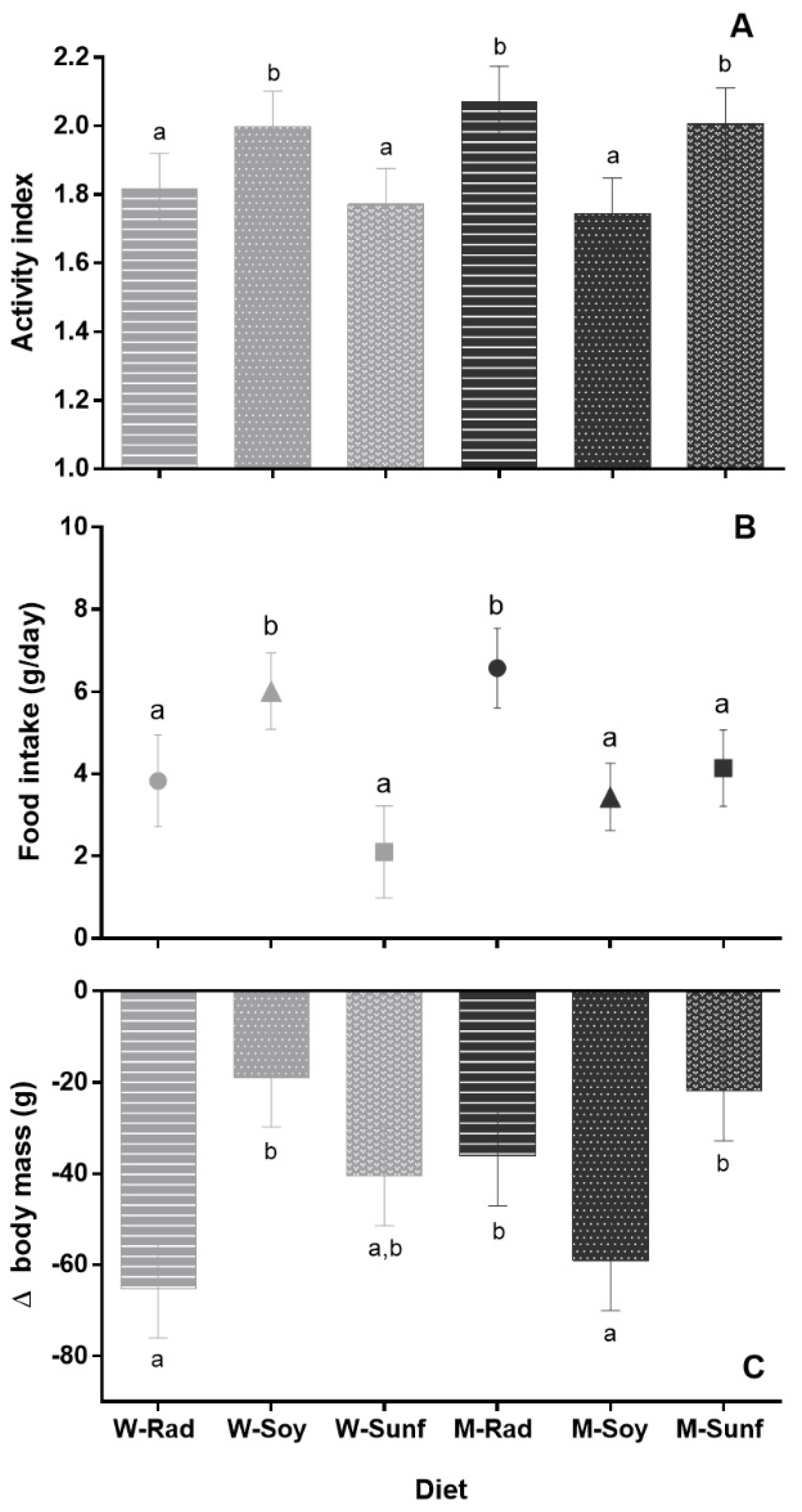
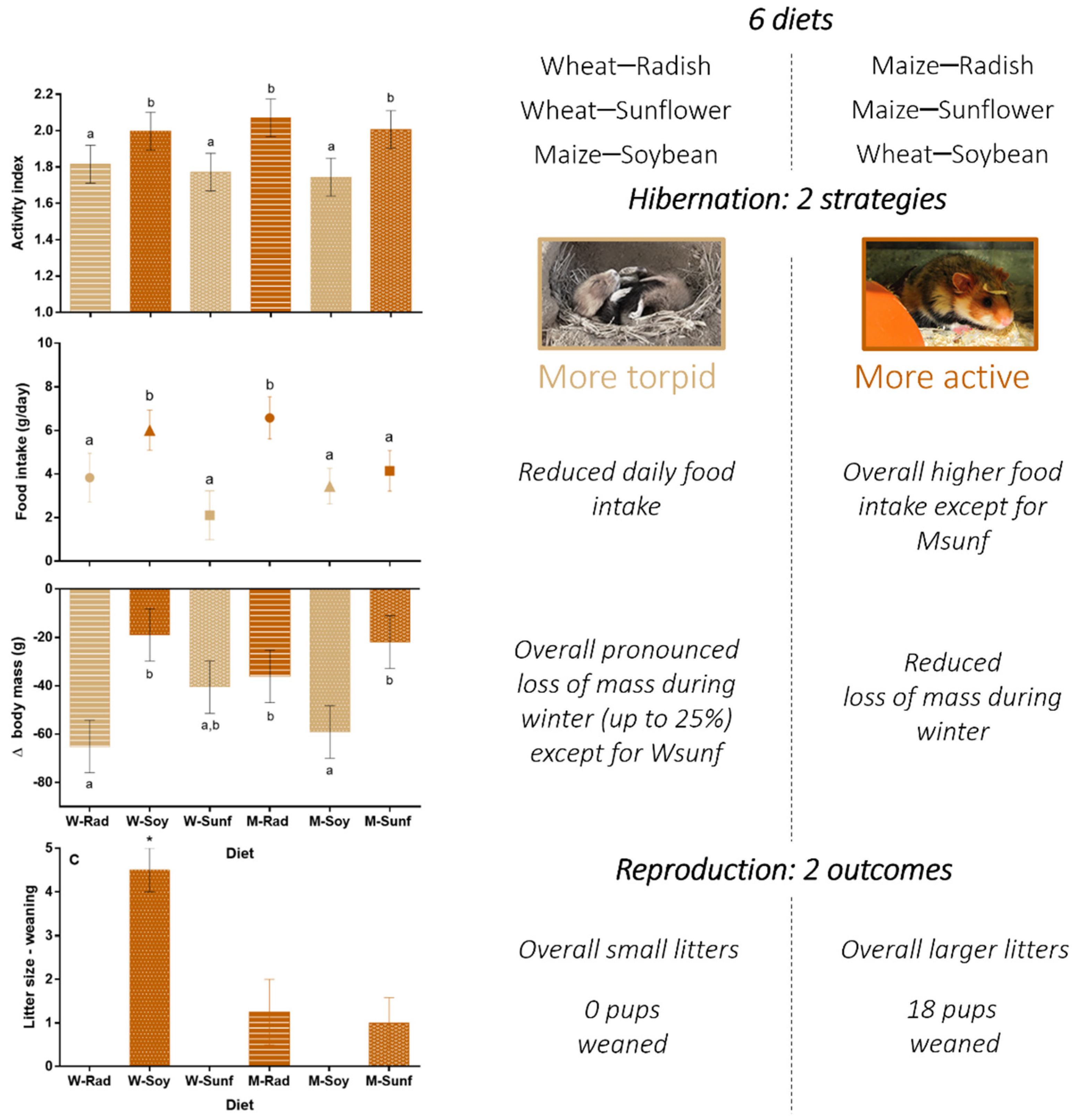
3.2. Activity Index during Hibernation
3.3. Food Intake and Body Mass Loss during Hibernation
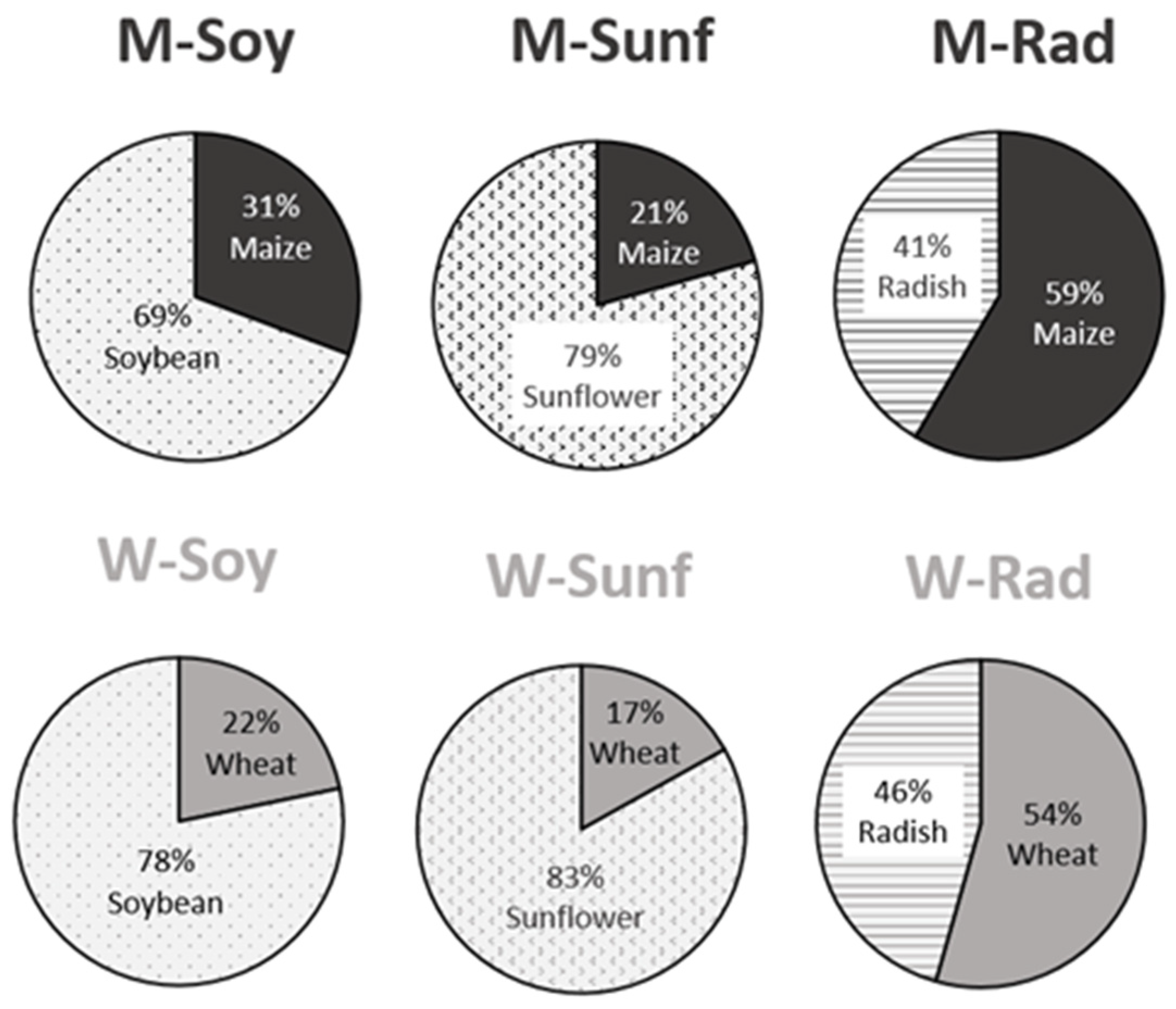
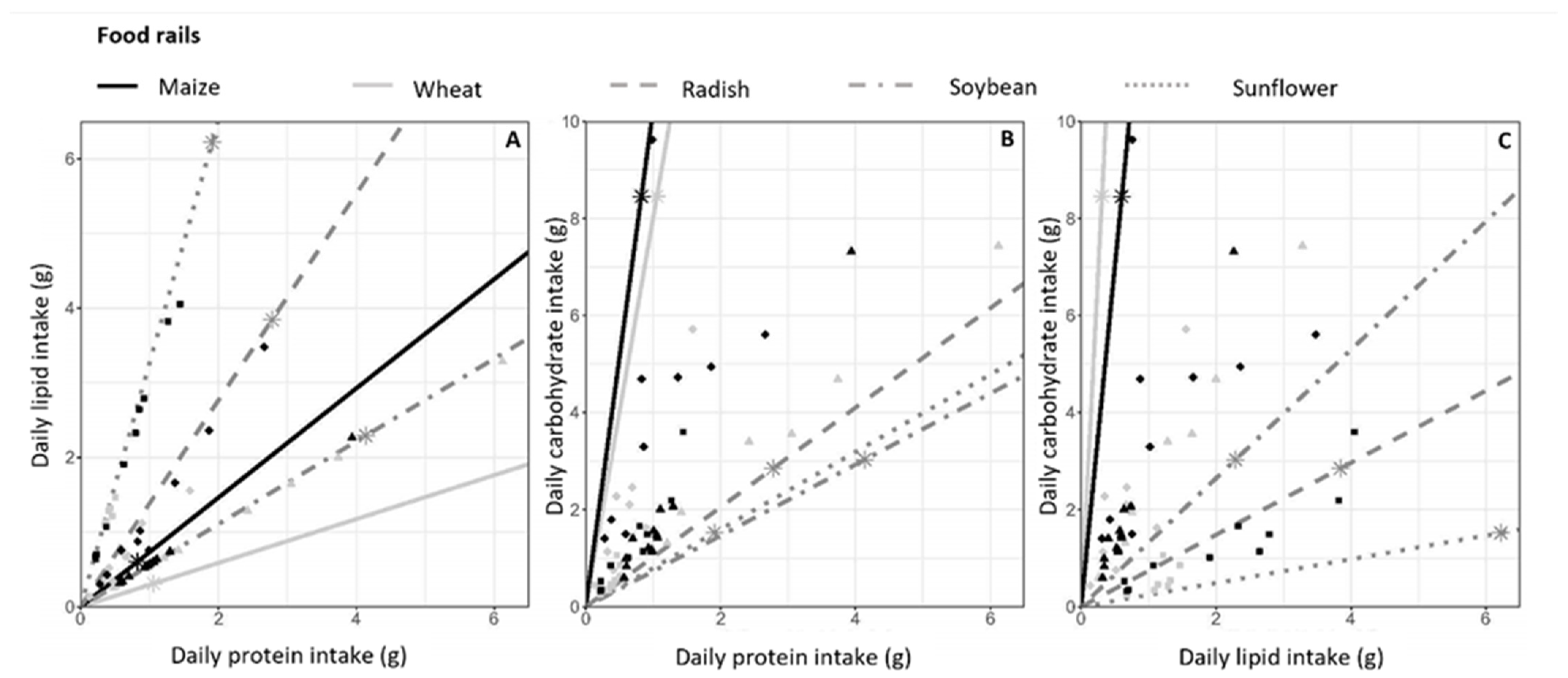
3.4. Food Selection and Macronutrient Intake
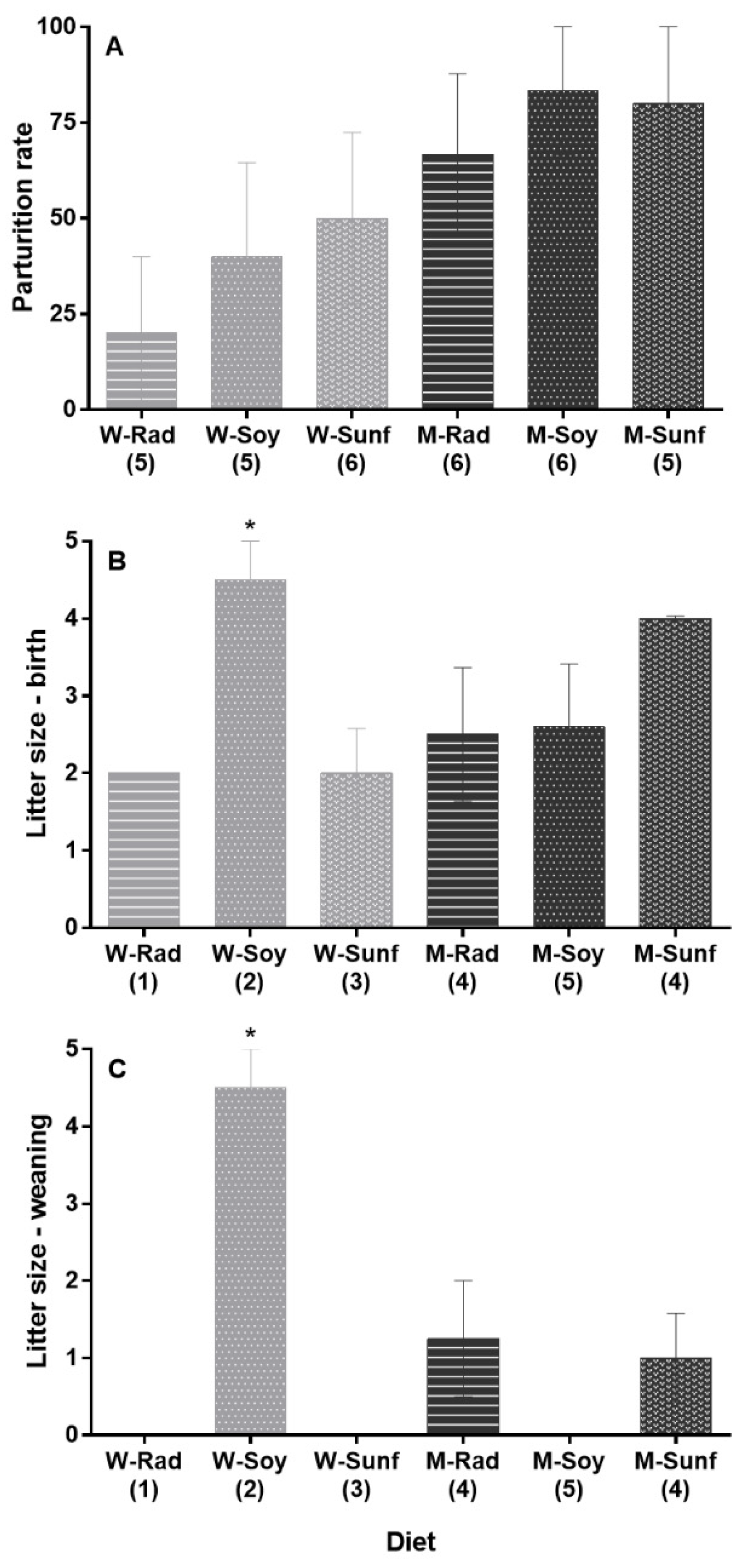
3.5. Female Reproductive Success
4. Discussion
4.1. Activity Index and ΔBody Mass: Sex and Diet Effects
4.2. Diet Effect on Reproduction
4.3. Favorable Crop Associations and Recommendations
4.4. From Knowledge to Action
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.U.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the Earth’s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed]
- Stoate, C.; Boatman, N.; Borralho, R.; Carvalhob, C.R.; Snoo, G.; Eden, P. Ecological impacts of arable intensification in Europe. J. Environ. Manag. 2001, 63, 337–365. [Google Scholar] [CrossRef]
- Benton, T.G.; Bryant, D.M.; Cole, L.; Crick, H.Q.P. Linking agricultural practice to insect and bird populations: A historical study over three decades. J. Appl. Ecol. 2002, 39, 673–687. [Google Scholar] [CrossRef]
- Geiger, F.; Bengtsson, J.; Berendse, F.; Weisser, W.W.; Emmerson, M.; Morales, M.B.; Ceryngier, P.; Liira, J.; Tscharntke, T.; Winqvist, C.; et al. Persistent negative effects of pesticides on biodiversity and biological control potential on European farmland. Basic Appl. Ecol. 2010, 11, 97–105. [Google Scholar] [CrossRef]
- Wickramasinghe, L.P.; Harris, S.; Jones, G.; Jennings, N.V. Abundance and Species Richness of Nocturnal Insects on Organic and Conventional Farms: Effects of Agricultural Intensification on Bat Foraging. Conserv. Biol. 2004, 18, 1283–1292. [Google Scholar] [CrossRef]
- Didham, R.K.; Basset, Y.; Collins, C.M.; Leather, S.R.; Littlewood, N.A.; Menz, M.H.M.; Müller, J.; Packer, L.; Saunders, M.E.; Schönrogge, K.; et al. Interpreting insect declines: Seven challenges and a way forward. Insect Conserv. Divers. 2020, 13, 103–114. [Google Scholar] [CrossRef]
- Guzman, L.M.; Johnson, S.A.; Mooers, A.O.; M’Gonigle, L.K. Using historical data to estimate bumble bee occurrence: Variable trends across species provide little support for community-level declines. Biol. Conserv. 2021, 257, 109141. [Google Scholar] [CrossRef]
- Hędrzak, M.; Badach, E.; Kornaś, S. Preliminary Assumptions for Identification of the Common Hamster (Cricetus cricetus) as a Service Provider in the Agricultural Ecosystem. Sustainability 2021, 13, 6793. [Google Scholar] [CrossRef]
- Banaszek, A.; Bogomolov, P.; Feoktistova, N.; La Haye, M.; Monecke, S.; Reiners, T.E.; Rusin, M.; Surov, A.; Weinhold, U.; Ziomek, J. Cricetus cricetus. IUCN Red List of Threaten Species. Available online: https://dx.doi.org/10.2305/IUCN.UK.2020-2.RLTS.T5529A111875852.en (accessed on 14 September 2021).
- Surov, A.; Banaszek, A.; Bogomolov, P.; Feoktistova, N.; Monecke, S. Dramatic global decrease in the range and reproduction rate of the European hamster Cricetus cricetus. Endanger. Species Res. 2016, 31, 119–145. [Google Scholar] [CrossRef]
- Chaigne, A.; Tissier, M.L.; Habold, C.; Eidenschenck, J.; Uhlrich, B. Le Grand hamster (Cricetus cricetus) en Alsace, quel devenir? In Bourgogne Nature—Les Mammifères Sauvages, Recolonisation et Réémergence; Actes du 37ème colloque de la SFEPM: St-Brisson, France, 2014; pp. 312–322. [Google Scholar]
- Weinhold, U. Draft European action plan for the conservation of the common hamster (Cricetus cricetus L., 1758). In Proceedings of the Convention on the Conservation of European Wildlife and Natural Habitats (28th Meeting Standing Committee), Strasbourg, France, 24–27 November 2008. [Google Scholar]
- La Haye, M.; Swinnen, K.; Kuiters, A.; Leirs, H.; Siepel, H. Modelling population dynamics of the Common hamster (Cricetus cricetus): Timing of harvest as a critical aspect in the conservation of a highly endangered rodent. Biol. Conserv. 2014, 180, 53–61. [Google Scholar] [CrossRef]
- Tissier, M.L.; Handrich, Y.; Robin, J.-P.; Weitten, M.; Pevet, P.; Kourkgy, C.; Habold, C. How maize monoculture and increasing winter rainfall have brought the hibernating European hamster to the verge of extinction. Sci. Rep. 2016, 6, 25531. [Google Scholar] [CrossRef] [PubMed]
- Tissier, M.L.; Marchandeau, S.; Habold, C.; Handrich, Y.; Eidenschenck, J.; Kourkgy, C. Weeds as a predominant food source: A review of the diet of common hamsters Cricetus cricetus in farmlands and urban habitats. Mammal Rev. 2018, 49, 152–170. [Google Scholar] [CrossRef]
- Agreste Statistique Agricole 2014, 2017, 2018. Available online: https://agreste.agriculture.gouv.fr/agreste-web/ (accessed on 18 May 2021).
- Tissier, M.L.; Handrich, Y.; Dallongeville, O.; Robin, J.-P.; Habold, C. Diets derived from maize monoculture cause maternal infanticides in the endangered European hamster due to a vitamin B3 deficiency. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162168. [Google Scholar] [CrossRef]
- Tissier, M.L.; Kletty, F.; Handrich, Y.; Habold, C. Monocultural sowing in mesocosms decreases the species richness of weeds and invertebrates and critically reduces the fitness of the endangered European hamster. Oecologia 2018, 186, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Humphries, M.M.; Thomas, D.W.; Kramer, D.L. The Role of Energy Availability in Mammalian Hibernation: A Cost-Benefit Approach. Physiol. Biochem. Zool. 2003, 76, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Franceschini-Zink, C.; Millesi, E. Population Development and Life Expectancy in Common Hamsters. 2008. Available online: https://ucris.univie.ac.at/portal/en/publications/population-development-and-life-expectancy-in-common-hamsters(2302eda9-2118-47ce-869d-e2ced0930227).html (accessed on 12 March 2016).
- Millesi, E.; Strijkstra, A.M.; Hoffmann, I.E.; Dittami, J.P.; Daan, S. Sex and Age Differences in Mass, Morphology, and Annual Cycle in European Ground Squirrels, Spermophilus citellus. J. Mammal. 1999, 80, 218–231. [Google Scholar] [CrossRef][Green Version]
- Hoogland, J.L. The Black-Tailed Prairie Dog: Social Life of a Burrowing Mammal; University of Chicago Press: Chicago, IL, USA, 1995. [Google Scholar]
- Dark, J. Annual lipid cycles in hibernators: Integration of physiology and behavior. Annu. Rev. Nutr. 2005, 25, 469–497. [Google Scholar]
- Humphries, M.M.; Thomas, D.W.; Kramer, D.L. Torpor and Digestion in Food-Storing Hibernators. Physiol. Biochem. Zool. 2001, 74, 283–292. [Google Scholar] [CrossRef]
- Weitten, M.; Tissier, M.L.; Robin, J.-P.; Habold, C. Dietary proteins improve hibernation and subsequent reproduction in the European hamster, Cricetus cricetus. Am. J. Physiol. Integr. Comp. Physiol. 2018, 315, R848–R855. [Google Scholar] [CrossRef]
- Siutz, C.; Nemeth, M.; Wagner, K.-H.; Quint, R.; Ruf, T.; Millesi, E. Effects of food store quality on hibernation performance in common hamsters. PLoS ONE 2017, 12, e0185913. [Google Scholar] [CrossRef]
- Arnold, W.; Giroud, S.; Valencak, T.G.; Ruf, T. Ecophysiology of Omega Fatty Acids: A Lid for Every Jar. Physiology 2015, 30, 232–240. [Google Scholar] [CrossRef]
- Munro, D.; Thomas, D.W. The role of polyunsaturated fatty acids in the expression of torpor by mammals: A review. Zoology 2004, 107, 29–48. [Google Scholar] [CrossRef]
- Jastroch, M.; Giroud, S.; Barrett, P.; Geiser, F.; Heldmaier, G.; Herwig, A. Seasonal Control of Mammalian Energy Balance: Recent Advances in the Understanding of Daily Torpor and Hibernation. J. Neuroendocr. 2016, 28. [Google Scholar] [CrossRef] [PubMed]
- Munro, D.; Thomas, D.W.; Humphries, M.M. Torpor patterns of hibernating eastern chipmunks Tamias striatus vary in response to the size and fatty acid composition of food hoards. J. Anim. Ecol. 2005, 74, 692–700. [Google Scholar] [CrossRef]
- Humphries, M.M.; Thomas, D.W.; Hall, C.L.; Speakman, J.R.; Kramer, D.L. The energetics of autumn mast hoarding in eastern chipmunks. Oecologia 2002, 133, 30–37. [Google Scholar] [CrossRef]
- Siutz, C.; Millesi, E. Torpor patterns in common hamsters with and without access to food stores. J. Comp. Physiol. B 2017, 187, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Tarnoczi, T.J.; Berkes, F. Sources of information for farmers’ adaptation practices in Canada’s Prairie agro-ecosystem. Clim. Chang. 2009, 98, 299. [Google Scholar] [CrossRef]
- Marr, E.J.; Howley, P. The accidental environmentalists: Factors affecting farmers’ adoption of pro-environmental activities in England and Ontario. J. Rural. Stud. 2019, 68, 100–111. [Google Scholar] [CrossRef]
- AFZ, I. CIRAD, FAO Feedipedia. Anim Feed Ressources Inf Syst. Available online: http://www.feedipedia.org/ (accessed on 24 July 2020).
- USDA, Nutrition Data. In USDA SR-21, Nutrition Data. Available online: http://nutritiondata.self.com/ (accessed on 1 September 2016).
- HeckenBenner, B.; de PontBriand, S. CIPAN: Quand L’outil Réglementaire Devient un Atout Agronomique et Faunistique. Available online: https://docplayer.fr/21149047-Cipan-quand-l-outil-reglementaire-devient-un-atout-agronomique-et-faunistique.html (accessed on 24 July 2020).
- Chen, G.; Weil, R.R. Penetration of cover crop roots through compacted soils. Plant Soil 2010, 331, 31–43. [Google Scholar] [CrossRef]
- Mandal, S.; Yadav, S.; Singh, R.; Begum, G.; Suneja, P.; Singh, M. Correlation studies on oil content and fatty acid profile of some Cruciferous species. Genet. Resour. Crop. Evol. 2002, 49, 551–556. [Google Scholar] [CrossRef]
- Kjeldahl, C. A new method for the determination of nitrogen in organic matter. Z. Anal. Chem. 1883, 22, 366. [Google Scholar] [CrossRef]
- Campbell, R.R.; Leatherland, J.F. Estimating Body Protein and Fat from Water Content in Lesser Snow Geese. J. Wildl. Manag. 1980, 44, 438. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Raubenheimer, D.; Simpson, S.J. Integrative models of nutrient balancing: Application to insects and vertebrates. Nutr. Res. Rev. 1997, 10, 151–179. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.J.; Raubenheimer, D. A multi-level analysis of feeding behaviour: The geometry of nutritional decisions. Philos. Trans. R. Soc. B Biol. Sci. 1993, 342, 381–402. [Google Scholar] [CrossRef]
- Concannon, P.W.; Fullam, L.A.; Baldwin, B.H.; Tennant, B.C. Effects of Induction Versus Prevention of Hibernation on Reproduction in Captive Male and Female Woodchucks (Marmota Monax)1. Biol. Reprod. 1989, 41, 255–261. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; ISBN 978-0-387-95364-9. [Google Scholar]
- Zar, J.H. Biostatistical Analysis; Pearson Prentice Hall./Pearson Education. Inc.: Upper Saddle River, NJ, USA, 1999; 255p. [Google Scholar]
- Michener, G.R. Sexual differences in over-winter torpor patterns of Richardson’s ground squirrels in natural hibernacula. Oecologia 1992, 89, 397–406. [Google Scholar] [CrossRef]
- Siutz, C.; Franceschini, C.; Millesi, E. Sex and age differences in hibernation patterns of common hamsters: Adult females hibernate for shorter periods than males. J. Comp. Physiol. B 2016, 186, 801–811. [Google Scholar] [CrossRef]
- Siutz, C.; Valent, M.; Ammann, V.; Niebauer, A.; Millesi, E. Sex-specific effects of food supplementation on hibernation performance and reproductive timing in free-ranging common hamsters. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Nechay, G.; Hamar, M.; Grulich, I. The Common Hamster (Cricetus cricetus [L.]); A Review. EPPO Bull. 1977, 7, 255–276. [Google Scholar] [CrossRef]
- Al-Badry, K.S.; Taha, H.M. Hibernation hypothermia and metabolism in hedgehogs—Changes in free amino acids and related compounds. Comp. Biochem. Physiol. Part A Physiol. 1982, 72, 541–547. [Google Scholar] [CrossRef]
- Giroud, S.; Stalder, G.; Gerritsmann, H.; Kübber-Heiss, A.; Kwak, J.; Arnold, W.; Ruf, T. Dietary Lipids Affect the Onset of Hibernation in the Garden Dormouse (Eliomys quercinus): Implications for Cardiac Function. Front. Physiol. 2018, 9, 1235. [Google Scholar] [CrossRef] [PubMed]
- Geiser, F. Influence of polyunsaturated and saturated dietary lipids on adipose tissue, brain and mitochondrial membrane fatty acid composition of a mammalian hibernator. Biochim. Et Biophys. Acta (BBA) Lipids Lipid Metab. 1990, 1046, 159–166. [Google Scholar] [CrossRef]
- Carey, H.V.; Andrews, M.T.; Martin, S.L. Mammalian Hibernation: Cellular and Molecular Responses to Depressed Metabolism and Low Temperature. Physiol. Rev. 2003, 83, 1153–1181. [Google Scholar] [CrossRef]
- Villemey, A.; Besnard, A.; Grandadam, J.; Eidenschenck, J. Testing restocking methods for an endangered species: Effects of predator exclusion and vegetation cover on common hamster (Cricetus cricetus) survival and reproduction. Biol. Conserv. 2013, 158, 147–154. [Google Scholar] [CrossRef]
- Wencel, M.-C.; Migot, P. Le grand hamster en France en 1998. In Proceedings of the International Hamster Workgroup Meeting, Muttersholtz, France, 31 October–1 November 1998; pp. 32–37. [Google Scholar]
- Tissier, M.L.; Habold, C.; Kletty, F.; Eidenschenck, J.; Marchandeau, S.; Handrich, Y.; Oswald, P.; Revel-Mouroz, A.; Kourkgy, C. Concilier agriculture et préservation de la faune de plaine: Le cas du grand hamster en Alsace. Faune Sauvage 2019, 322, 31–39. [Google Scholar]
- Amand, B.; Duponteil, A.; Strosser, P.; Boos, M. National Actions Plan for the Common Hamster Cricetus Cricetus; MEDDE: Paris, France, 2012; p. 175. [Google Scholar]
- Virion, M.-C. Plan National D’actions en Faveur du Hamster Commun Cricetus cricetus et de la Biodiversité de la Plaine d’Alsace 2019–2028. Direction Régionale de l’Environnement, de l’Aménagement et du Logement Grand Est. 130p. Ministère de la Transition Écologique et Solidaire. Available online: https://www.ecologie.gouv.fr/sites/default/files/pna-Hamster-final_compressed.pdf (accessed on 1 December 2021).
- Glavan, M.; Železnikar, Š.; Velthof, G.; Boekhold, S.; Langaas, S.; Pintar, M. How to Enhance the Role of Science in European Union Policy Making and Implementation: The Case of Agricultural Impacts on Drinking Water Quality. Water 2019, 11, 492. [Google Scholar] [CrossRef]
- Arlettaz, R.; Schaub, M.; Fournier, J.; Reichlin, T.S.; Sierro, A.; Watson, J.; Braunisch, V. From Publications to Public Actions: When Conservation Biologists Bridge the Gap between Research and Implementation. BioScience 2010, 60, 835–842. [Google Scholar] [CrossRef]
- Gregory, R.; Arvai, J.; Gerber, L.R. Structuring Decisions for Managing Threatened and Endangered Species in a Changing Climate. Conserv. Biol. 2013, 27, 1212–1221. [Google Scholar] [CrossRef]
- Naujokaitis-Lewis, I.; Pomara, L.Y.; Zuckerberg, B. Delaying conservation actions matters for species vulnerable to climate change. J. Appl. Ecol. 2018, 55, 2843–2853. [Google Scholar] [CrossRef]
- Fischer, C.; Wagner, C. Can agri-environmental schemes enhance non-target species? Effects of sown wildflower fields on the common hamster (Cricetus cricetus) at local and landscape scales. Biol. Conserv. 2016, 194, 168–175. [Google Scholar] [CrossRef]
- Bald, V.; Boetzl, F.A.; Krauss, J. Where do hamsters go after cereal harvest? A case study. Basic Appl. Ecol. 2021, 54, 98–107. [Google Scholar] [CrossRef]
- Hagy, H.M.; Linz, G.M.; Bleier, W.J. Wildlife Conservation Sunflower Plots and Croplands as Fall Habitat for Migratory Birds. Am. Midl. Nat. 2010, 164, 119–135. [Google Scholar] [CrossRef]
- Jones, G.A.; Sieving, K. Intercropping sunflower in organic vegetables to augment bird predators of arthropods. Agric. Ecosyst. Environ. 2006, 117, 171–177. [Google Scholar] [CrossRef]
- LoCascio, G.M.; Aguirre, L.; Irwin, R.E.; Adler, L.S. Pollen from multiple sunflower cultivars and species reduces a common bumblebee gut pathogen. R. Soc. Open Sci. 2019, 6, 190279. [Google Scholar] [CrossRef]
- Giacomini, J.J.; Leslie, J.; Tarpy, D.; Palmer-Young, E.C.; Irwin, R.E.; Adler, L.S. Medicinal value of sunflower pollen against bee pathogens. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Ramesh, K.; Ashok, K.P.; Biswas, A.K. Best Management Practices for Soybean under Soybean-Wheat System to Minimize the Impact of Climate Change. Indian J. Fertil. 2017, 13, 42–55. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Period | Photoperiod | Ta | Humidity |
|---|---|---|---|---|
| 10–16 December | Food habituation | Natural (fall, winter) | 10 ± 1 °C | 55–65% |
| 16 December–8 March | Hibernation | Natural (winter) | 10 ± 1 °C | 55–65% |
| 8 March–10 June | Reproduction | Natural (spring, summer) | 20 ± 1 °C | 50–60% |
| Food Item | DM (% of FM) | Water (% of FM) | Minerals (% of DM) | Lipids (% of DM) | Proteins (% of DM) | Carbohydrates (% of DM) | Energy (kJ/g of DM) |
|---|---|---|---|---|---|---|---|
| Maize | 92.87 ± 0.43 | 7.13 ±0.43 | 1.17 ± 0.13 | 6.03 ± 0.27 | 8.25 ± 0.10 | 84.54 ± 0.24 | 18.21 ± 0.10 |
| Wheat | 91.17 ± 0.47 | 8.83 ± 0.47 | 1.77 ± 0.05 | 3.10 ± 0.09 | 10.56 ± 0.12 | 84.57 ± 0.17 | 17.55 ± 0.13 |
| Radish | 94.37 ± 0.27 | 5.63 ± 0.27 | 5.32 ± 0.03 | 38.42 ± 0.46 | 27.78 ± 0.15 | 28.48 ± 0.37 | 26.40 ± 0.23 |
| Soybean | 93.88 ± 0.49 | 6.12 ± 0.49 | 5.47 ± 0.17 | 22.89 ± 0.18 | 41.36 ± 0.33 | 30.28 ± 0.36 | 23.53 ± 0.11 |
| Sunflower | 95.87 ± 0.15 | 4.13 ± 0.15 | 3.49 ± 0.08 | 62.21 ± 0.32 | 19.08 ± 0.14 | 15.21 ± 0.32 | 31.38 ± 0.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tissier, M.L.; Kletty, F.; Robin, J.-P.; Habold, C. Sustainable Agriculture: Nutritional Benefits of Wheat–Soybean and Maize–Sunflower Associations for Hibernation and Reproduction of Endangered Common Hamsters. Sustainability 2021, 13, 13521. https://doi.org/10.3390/su132413521
Tissier ML, Kletty F, Robin J-P, Habold C. Sustainable Agriculture: Nutritional Benefits of Wheat–Soybean and Maize–Sunflower Associations for Hibernation and Reproduction of Endangered Common Hamsters. Sustainability. 2021; 13(24):13521. https://doi.org/10.3390/su132413521
Chicago/Turabian StyleTissier, Mathilde Louise, Florian Kletty, Jean-Patrice Robin, and Caroline Habold. 2021. "Sustainable Agriculture: Nutritional Benefits of Wheat–Soybean and Maize–Sunflower Associations for Hibernation and Reproduction of Endangered Common Hamsters" Sustainability 13, no. 24: 13521. https://doi.org/10.3390/su132413521
APA StyleTissier, M. L., Kletty, F., Robin, J.-P., & Habold, C. (2021). Sustainable Agriculture: Nutritional Benefits of Wheat–Soybean and Maize–Sunflower Associations for Hibernation and Reproduction of Endangered Common Hamsters. Sustainability, 13(24), 13521. https://doi.org/10.3390/su132413521