
Impact of Secondary Succession in the Xerothermic Grassland on the Population of the Eastern Pasque Flower (Pulsatilla patens)—Preliminary Studies


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Sampling
2.3. Data Analysis
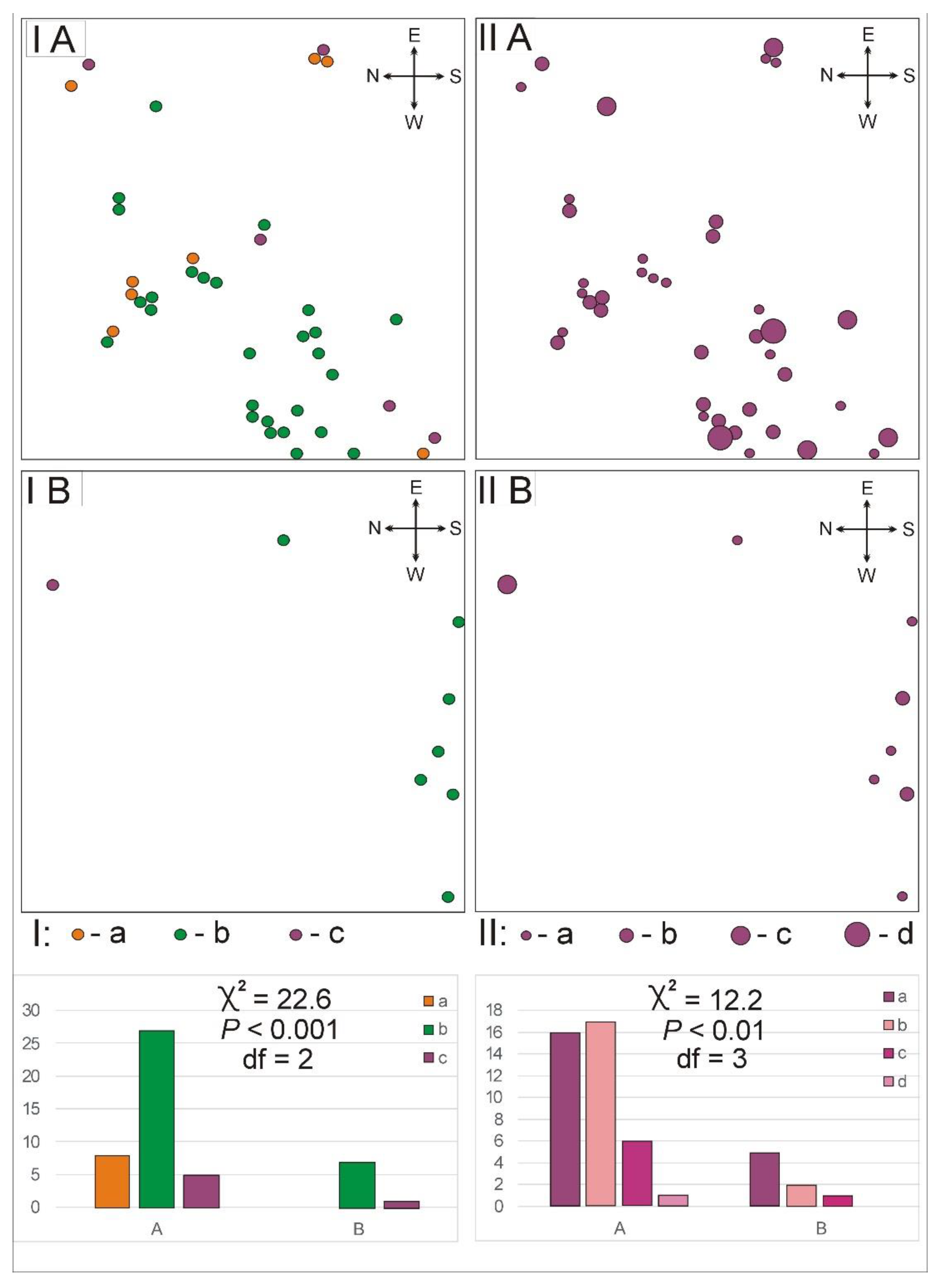
3. Results
3.1. The Habitat Conditions
3.2. The Population Structure
3.3. The Traits of Individuals
4. Discussion
5. Conclusions
- The total abundance of Pulsatilla patens in the open xerothermic grassland is five times higher than in the overgrowing xerothermic grassland, (40:8 leaf rosettes, respectively).
- A randomly clustering distribution of P. patens was noted only in the open xerothermic grassland (in the overgrowing xerothermic grassland a random type of distribution occurred).
- The density structure of the rosettes of P. patens differed significantly between the study plots. Leaf rosettes showed from 6 to 10 leaves in the open xerothermic grassland, whereas only from 1 to 5 leaves in the overgrowing xerothermic grassland.
- The difference on the basis of age structure of the P. patens shows that only in the open xerothermic grassland juvenile individuals were noted.
- The mean number of leaves in the rosettes was higher in the open xerothermic grassland, but the leaf dimensions were higher in the overgrowing xerothermic grassland (a shadowing caused by a process of secondary succession increased the height and width of the leaves).
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bartemucci, P.; Messier, C.H.; Canham, C.H.D. Overstory influences on light attenuation patterns and understory plant community diversity and composition in southern boreal forests of Quebec. Can. J. For. Res. 2006, 36, 2065–2079. [Google Scholar] [CrossRef]
- Domke, G.M.; Caspersen, J.P.; Jones, T.A. Light attenuation following selection harvesting in northern hardwood forests. For. Ecol. Manag. 2007, 239, 182–190. [Google Scholar] [CrossRef]
- Kostrakiewicz, K. The influence of shadow created by adjacent plants on phenotypic plasticity of endangered species Trollius europaeus L. (Ranunculacae). Pol. J. Ecol. 2009, 57, 625. [Google Scholar]
- Zervoudakis, G.; Salahas, G.; Kaspiris, G.; Konstantopoulou, E. Influence of light intensity on growth and physiological characteristics of common sage (Salvia officinalis L.). Braz. Arch. Biol. Technol. 2012, 55, 89–95. [Google Scholar] [CrossRef]
- Navvab, M. Daylighting aspects for plant growth in interior environments. Light Eng. 2009, 17, 46–54. [Google Scholar]
- Rezazadeh, A.; Harkess, R.L.; Telmadarrehei, T. The Effect of light intensity and temperature on flowering and morphology of potted red firespike. Horticulturae 2018, 4, 36. [Google Scholar] [CrossRef]
- Li, J.; Zheng, Z.; Xie, H.; Zhao, N.; Gao, Y. Increased soil nutrition and decreased light intensity drive species loss after eight years grassland enclosures. Sci. Rep. 2017, 7, 44525. [Google Scholar] [CrossRef] [PubMed]
- Poptcheva, K.; Schwartze, P.; Vogel, A.; Kleinebecker, T.; Holzel, N. Changes in wet meadow vegetation after 20 years of different management in a field experiment (North-West Germany). Agric. Ecosyst. Environ. 2009, 134, 108–114. [Google Scholar] [CrossRef]
- Michalska-Hejduk, D.; Kopeć, D. Dynamics of semi-natural vegetation with a focus on Molinion meadows after 50 years of strict protection. Pol. J. Environ. Stud. 2012, 21, 1731–1741. [Google Scholar]
- Marriott, C.A.; Hood, K.; Fisher, J.M.; Pakeman, R.J. Long-term impacts of extensive grazing and abandonment on the species composition, richness, diversity and productivity of agricultural grassland. Agric. Ecosyst. Environ. 2009, 134, 190–200. [Google Scholar] [CrossRef]
- Ignatavičius, G.; Sinkevičius, S.; Ložytė, A. Effects of grassland management on plant communities. Ekologija 2013, 59, 99–110. [Google Scholar] [CrossRef]
- European Communities (Ed.) Council Directive 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora. 2009. Available online: http://ec.europa.eu/environment/nature/legislation/habitatsdirective/index_en.htm (accessed on 15 August 2020).
- Liess, A.; Lange, K.; Schulz, F.; Piggott, J.J.; Matthaei, C.D.; Townsend, C.R. Light, nutrients and grazing interact to determine diatom species richness via changes to productivity, nutrient state and grazer activity. J. Ecol. 2009, 97, 326–336. [Google Scholar] [CrossRef]
- Poschlod, P.; Bakker, J.P.; Kahmen, S. Changing land use and its impact on biodiversity. Basic Appl. Ecol. 2005, 6, 93–98. [Google Scholar] [CrossRef]
- Łazarski, G. Protected, rare and endangered vascular plant species in the Chęciny Hills and Dyminy Range (Małopolska Upland)—part I. Grassland and thermophilous fringe species. Fragm. Florist. Geobot. Pol. 2019, 26, 49–73. [Google Scholar]
- Łazarski, G.; Podgórska, M. New and highly threatened locality of Pulsatilla patens subsp. patens (Ranunculaceae) in the Świętokrzyskie Mountains (Małopolska Upland). Fragm. Florist. Geobot. Pol. 2020, 27, 527–535. [Google Scholar]
- Meusel, H.; Jäger, E.; Weinert, E. Vergleichende Chorologie der Zentraleuropäischen Flora; Karten. Fischer: Jena, Germany, 1965. [Google Scholar]
- Hulten, E.; Fries, M. Atlas of North European Vascular Plants North of the Tropic of Cancer; Koeltz: Konigstein, Germany, 1986. [Google Scholar]
- Tutin, T.G.; Akeroyd, J.R. Pulsatilla Miller. In Flora Europaea, Vol. 1: Psilotaceae to Platanaceae, 2nd ed.; Tutin, T.G., Burges, N.A., Chater, A.O., Edmondson, J.R., Heywood, V.H., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1993; pp. 264–266. [Google Scholar]
- Zając, A.; Zając, M. Distribution Atlas of Vascular Plants in Poland; Laboratory of Computer Chorology, Institute of Botany, Jagiellonian University: Kraków, Poland, 2001. [Google Scholar]
- Council of Europe. Convention on the Conservation of European Wildlife and Natural Habitats. 1979. Available online: http://conventions.coe.int/Treaty/en/Treaties/Word/104.doc (accessed on 15 August 2020).
- Bilz, M. Pulsatilla patens. The IUCN Red List of Threatened Species 2011: E.T165908A6162193. 2011. Available online: https://www.iucnredlist.org/species/165908/6162193 (accessed on 15 August 2020).
- Kalliovirta, M.; Ryttäri, T.; Heikkinen, R.K. Population structure of a threatened plant, Pulsatilla patens, in boreal forests: Modelling relationships to overgrowth and site closure. Biodivers. Conserv. 2006, 15, 3095–3108. [Google Scholar] [CrossRef]
- Podgórska, M.; Bróż, E. The extinction of stations of Pulsatilla patens (L.) Mill. subsp. patens (American pasqueflower) on the Małopolska Upland. Acta Soc. Bot. Pol. Suppl. 2010, 79, 53. [Google Scholar]
- Łazarski, G.; Podgórska, M.; Bróż, E. Pasque flowers at the Natura 2000 Site Wzgórza Chęcińsko- Kieleckie and its vicinity—distribution and conservation status. Chrońmy Przyr. Ojczysta 2018, 74, 37–51. [Google Scholar]
- Uotila, P. Decline of Anemone patens (Ranunculaceae) in Finland. Acta Univ. Ups. Symb. Bot. Ups. 1996, 31, 205–210. [Google Scholar]
- Pilt, I.; Kukk, Ü. Pulsatilla patens and Pulsatilla pratensis (Ranunculaceae) in Estonia: Distribution and ecology. Proc. Nat. Acad. Sci. USA 2002, 51, 242–256. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie, Grundzüge der Vegetationskunde [Plant Sociology, Basics of Vegetation Science], 3rd ed.; Springer: Berlin/Heidelberg, Germany, 1964. [Google Scholar] [CrossRef]
- Kostrakiewicz-Gierałt, K.; Podgórska, M. Regeneration of the rare meadow species Iris sibirica in a postcultural land. Bot. Lett. 2020, 167, 331–339. [Google Scholar] [CrossRef]
- Jarzyna, K.; Podgórska, M.; Szwed, M.; Jóźwiak, M. A simple light meter as a device for studying the influence of seasonal changes of light conditions on the phenology of herbaceous undergrowth species in a fertile beach forest. Balt. For. 2018, 249, 148–157. [Google Scholar]
- Comeau, P.G. Measuring Light in a Forest; Extension Note 42; British Columbia Ministry of Forest, Research Branch: Vancouver, BC, Canada, 2000. [Google Scholar]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Vascular Plants of Poland. An Annotated Checklist; W. Szafer Institute of Botany PAS: Kraków, Poland, 2020; p. 526. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- StatSoft. STATISTICA (Data Analysis Software System), Version 6.1.; StatSoft, Inc.: Tulsa, OK, USA, 2003. [Google Scholar]
- Valkó, O.; Żmihorski, M.; Biurrun, I.; Loos, J.; Labadessa, R.; Venn, S. Ecology and conservation of steppes and semi-natural grasslands. Hacquetia 2016, 15, 5–14. [Google Scholar] [CrossRef]
- Valkó, O.; Venn, S.; Żmihorski, M.; Biurrun, I.; Labadessa, R.; Loos, J. The challenge of abandonment for the sustainable management of Palaearctic natural and semi-natural grasslands. Hacquetia 2018, 17, 5–16. [Google Scholar] [CrossRef]
- Dzwonko, Z. Effect of changes in land use during the 20th century on woodland and calcareous grassland vegetation in southern Poland. Acta Univ. Lodz Folia Biol. Oecologica 2011, 7, 27–48. [Google Scholar] [CrossRef]
- Bąba, W. Changes in the structure and floristic composition of the limestone grasslands after cutting trees and shrubs and mowing. Acta Soc. Bot. Pol. 2003, 72, 61–69. [Google Scholar] [CrossRef]
- Bąba, W. The species composition and dynamics in well-preserved and restored calcareous xerothermic grasslands (South Poland). Biol. Bratisl. 2004, 59, 447–456. [Google Scholar]
- Bąba, W.; Kompała-Bąba, A. Do small-scale gaps in calcareous grassland swards facilitate seedling establishment? Acta Soc. Bot. Pol. 2005, 74, 125–131. [Google Scholar] [CrossRef]
- Dzwonko, Z.; Loster, S. Changes in plant species composition in abandoned and restored limestone grasslands—the effects of tree and shrub cutting. Acta Soc. Bot. Pol. 2008, 77, 67–75. [Google Scholar] [CrossRef][Green Version]
- Kostrakiewicz-Gierałt, K. The size structure of ramets in Dianthus superbus L. in mosaic meadow vegetation. Acta Agrobot. 2013, 66, 23–30. [Google Scholar] [CrossRef][Green Version]
- Kubíková, P.; Zeidler, M. Habitat demands and population characteristics of the rare plant species Gladiolus imbricatus L. in the Frenštát region (NE Moravia, the Czech Republic). Čas. Slez. Zemského Muz. Sér. A 2011, 60, 154–164. [Google Scholar] [CrossRef][Green Version]
- Juśkiewicz-Swaczyna, B. Population structure of Pulsatilla patens in relation to the habitat quality. Tuexenia 2010, 30, 457–466. [Google Scholar]
- Ciosek, M.T.; Piórek, K.; Sikorski, R.; Trębicka, A. Population dynamics of Pulsatilla patens (L.) Mill. in a new locality in Poland. Biodivers. Res. Conserv. 2016, 41, 61–68. [Google Scholar] [CrossRef]
- Kricsfalusy, V. Variations in the life cycle of Anemone patens L. (Ranunculaceae) in wild populations of Canada. Plants 2016, 5, 29. [Google Scholar] [CrossRef]
- Podgórska, M. The long-term changes of forest communities as an effect of former iron-ore mining activities and current forest management: Importance for local biodiversity. Pol. J. Ecol. 2016, 64, 35–44. [Google Scholar] [CrossRef]
- Sienkiewicz, A.; Łaska, G. Application of Bayesian networks in evaluation of current status and protection of Pulsatilla patens (L.) Mill. Ecosphere 2021, 12, e03337. [Google Scholar] [CrossRef]
- Sultangazina, G.J.; Kuprijanov, O.A.; Kuprijanov, A.N.; Beyshov, R.S. Coenoflora Pulsatilla patens (L.) Mill. s. l. in northern Kazakhstan. Bull. Natl. Acad. Sci. Repub. Kazakhstan 2019, 380, 83–92. [Google Scholar] [CrossRef]
- Juśkiewicz-Swaczyna, B.; Choszcz, D. Effect of habitat quality on the structure of populations of Pulsatilla patens (L.) Mill. (Ranunculaceae)—Rare and endangered species in European flora. Pol. J. Ecol. 2012, 60, 567–576. [Google Scholar]


{kind=link}
| Conditions | A | B | The Mann–Whitney U Test Value | p Value | |
|---|---|---|---|---|---|
| plants | The maximum height of the neighbouring plants (cm) | 53.4 (34–90) | 98.9 (78–117) | U = −3.55 | p ≤ 0.001 |
| The minimum height of the neighbouring plants (cm) | 8.0 (4–15) | 47.6 (26–60) | U = −3.77 | p ≤ 0.001 | |
| light | The light (klx) | 16.97 (15.12–17.45) | 0.44 (0.32–0.55) | U = 2.88 | p ≤ 0.001 |
| soil | pH KCl | 7.6 (7.3–7.7) | 7.4 (7.2–7.5) | U = 0.48 | p = 0.63 |
| Ca (mg/l) | 4135 (4125–4142) | 4125 (4121–4126) | U = 0.53 | p = 0.71 | |
| Mg (mg/100 g) | 1337 (1332–1338) | 1338 (1331–1340) | U = −1.28 | p = 0.21 | |
| K (mg/100 g) | 57.1 (55.1–57.4) | 54.7 (54.1–54.8) | U = 0.16 | p = 0.87 | |
| P (mg/10 0g) | 17.9 (16.2–18.0) | 16.1 (15.9–17.0) | U = −0.16 | p = 0.87 |
| No. of Relevé | 1 | 2 | |
|---|---|---|---|
| Date (d/m/y) | - | 5 June 2020 | 5 June 2020 |
| Area of relevé [m2] | 25 | 25 | |
| Coverage of tree layer a [%] | - | 25 | |
| Coverage of shrub layer b1 [%] | - | 25 | |
| Coverage of shrub layer b2 [%] | 2 | 40 | |
| Coverage of herb layer c [%] | 85 | 100 | |
| No. of species in relevé | 38 | 41 | |
| Trees and shrubs: | |||
| ChCl. Vaccinio-Piceetea Br.-Bl. in Br.-Bl. et al. 1939: | |||
| Pinus sylvestris a | . | 2 | |
| Pinus sylvestris b1 | . | 2 | |
| Pinus sylvestris b2 | + | + | |
| Pinus sylvestris c | + | . | |
| ChCl. Crataego-Prunetea Tx. 1962: | |||
| Rhamnus cathartica b2 | . | 2 | |
| Cornus sanguinea b2 | . | 1 | |
| Cornus sanguinea c | . | + | |
| Prunus spinosa b2 | . | + | |
| Crataegus monogyna b2 | . | + | |
| Rosa rubiginosa c | + | . | |
| Others: | |||
| Frangula alnus b2 | . | 2 | |
| Frangula alnus c | . | + | |
| Betula pendula b1 | . | + | |
| Juniperus communis b2 | . | + | |
| Juniperus communis c | + | . | |
| Quercus robur c | + | . | |
| Padus avium b2 | . | + | |
| Herbs: | |||
| ChAll. Cirsio-Brachypodion pinnati Hadač et Klika in Klika et Hadač 1944: | |||
| Asperula tinctoria c | + | 1 | |
| Seseli annuum c | + | + | |
| Viola rupestris c | + | . | |
| ChO. Festucetalia valesiacae Soó 1947: | |||
| Thymus marschallianus c | + | + | |
| ChCl. Festuco-Brometea Br.-Bl. et Tx. ex Soó 1947: | |||
| Brachypodium pinnatum c | 3 | 3 | |
| Filipendula vulgaris c | + | 2 | |
| Helianthemum nummularium subsp. obscurum c | 1 | + | |
| Euphorbia cyparissias c | + | + | |
| Veronica spicata c | + | + | |
| Carlina vulgaris c | . | + | |
| Plantago media c | + | + | |
| Carex caryophyllea c | . | + | |
| ChCl. Trifolio medii-Geranietea sanguinei T. Müller 1962: | |||
| Anemone sylvestris c | 2 | 1 | |
| Peucedanum oreoselinum c | 1 | 2 | |
| Viola hirta c | + | 1 | |
| Anthericum ramosum c | 1 | 2 | |
| Polygonatum odoratum c | . | 1 | |
| Coronilla varia c | + | + | |
| Galium verum c | + | . | |
| ChCl. Molinio-Arrhenatheretea Tx. 1937: | |||
| Galium boreale c | 2 | 2 | |
| Knautia arvensis c | . | 1 | |
| Taraxacum officinale c | + | . | |
| Galium mollugo | . | + | |
| Anthyllis vulneraria c | + | . | |
| Trifolim montanum c | + | . | |
| Plantago lanceolata c | + | . | |
| Centaurea jacea c | . | + | |
| Others: | |||
| Pulsatilla patens c | 2 | 1 | |
| Chamaecytisus ruthenicus c | 2 | 2 | |
| Festuca trachyphylla c | 1 | 1 | |
| Sanguisorba minor c | 1 | + | |
| Pimpinella saxifraga c | + | + | |
| Vicia angustifolia c | + | + | |
| Briza media c | + | + | |
| Prunella grandiflora c | 1 | . | |
| Poa angustifolia c | + | 1 | |
| Carex montana c | + | . | |
| Polygala comosa c | + | . | |
| Carlina acaulis c | + | . | |
| Hieracium pilosella c | + | . | |
| Viola collina c | . | + | |
| Vincetoxicum hirundinaria c | . | + | |
| Primula veris c | . | + | |
| Traits | A | B | The Mann–Whitney U Test Value | p Value |
|---|---|---|---|---|
| The number of leaves in rosettes | 6.6 (1–18) | 5.7 (2–14) | U = 141.0 | p = 0.59 |
| The height of the greatest leaf in rosette | 11.2 (4.5–17) | 18.1 (12.5–23) | U = 22.0 | p = 0.0001 |
| The height of the lowest leaf in rosette | 6.9 (3–14) | 12.9 (7.5–18.5) | U = 28.5 | p= 0.0003 |
| The width of the largest leaf blade in rosette | 6.2 (2–9.8) | 8.1 (6.5–10) | U = 50.5 | p = 0.002 |
| The width of the least leaf blade in rosette | 3.9 (1.5–6.2) | 5.4 (3.5–8) | U = 89.5 | p = 0.051 |
| The length of the greatest lobe of the leaf in rosette | 3.4 (1–5.5) | 4.3 (3.5–5.5) | U = 53.5 | p = 0.003 |
| The length of the lowest lobe of the leaf in rosette | 2.3 (0.5–3.5) | 3.4 (2–5) | U = 72.5 | p= 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podgórska, M.; Łazarski, G. Impact of Secondary Succession in the Xerothermic Grassland on the Population of the Eastern Pasque Flower (Pulsatilla patens)—Preliminary Studies. Sustainability 2021, 13, 12575. https://doi.org/10.3390/su132212575
Podgórska M, Łazarski G. Impact of Secondary Succession in the Xerothermic Grassland on the Population of the Eastern Pasque Flower (Pulsatilla patens)—Preliminary Studies. Sustainability. 2021; 13(22):12575. https://doi.org/10.3390/su132212575
Chicago/Turabian StylePodgórska, Monika, and Grzegorz Łazarski. 2021. "Impact of Secondary Succession in the Xerothermic Grassland on the Population of the Eastern Pasque Flower (Pulsatilla patens)—Preliminary Studies" Sustainability 13, no. 22: 12575. https://doi.org/10.3390/su132212575
APA StylePodgórska, M., & Łazarski, G. (2021). Impact of Secondary Succession in the Xerothermic Grassland on the Population of the Eastern Pasque Flower (Pulsatilla patens)—Preliminary Studies. Sustainability, 13(22), 12575. https://doi.org/10.3390/su132212575