Author Contributions
Conceptualization, M.M., L.O., E.C. and C.C.; methodology, I.D.M.; software, L.O.; validation, M.M., I.D.M. and C.C.; formal analysis, L.O.; investigation, E.C.; resources, M.M.; data curation, E.C.; writing—original draft preparation, L.O., I.D.M. and C.C.; writing—review and editing, I.D.M. and C.C.; visualization, E.C.; supervision, M.M.; project administration, M.M. and C.C.; funding acquisition, M.M. All authors have read and agreed to the published version of the manuscript.
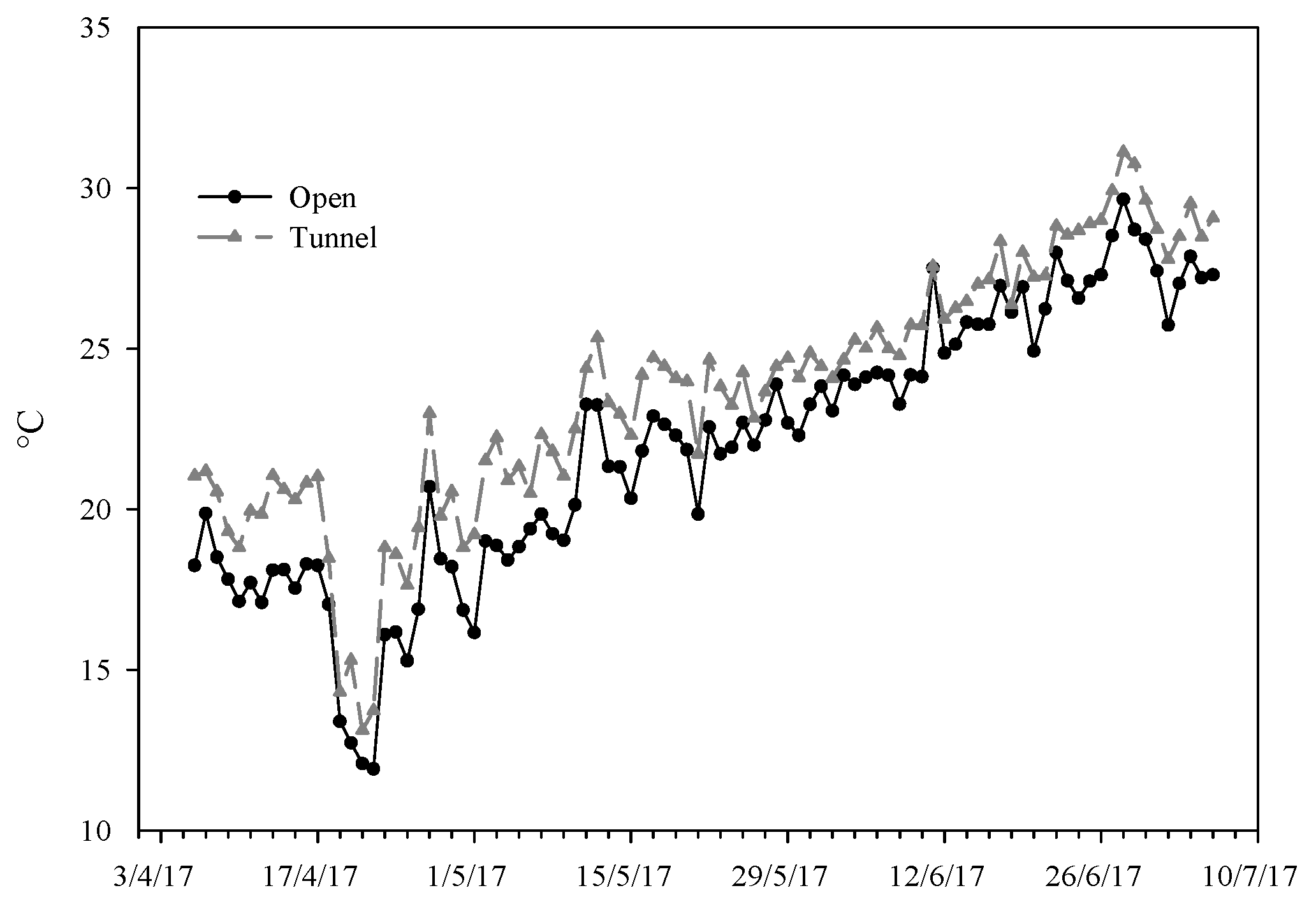
Figure 1.
Average temperature during the test period in the two environments: open (OF) and tunnel (PE).
Figure 1.
Average temperature during the test period in the two environments: open (OF) and tunnel (PE).
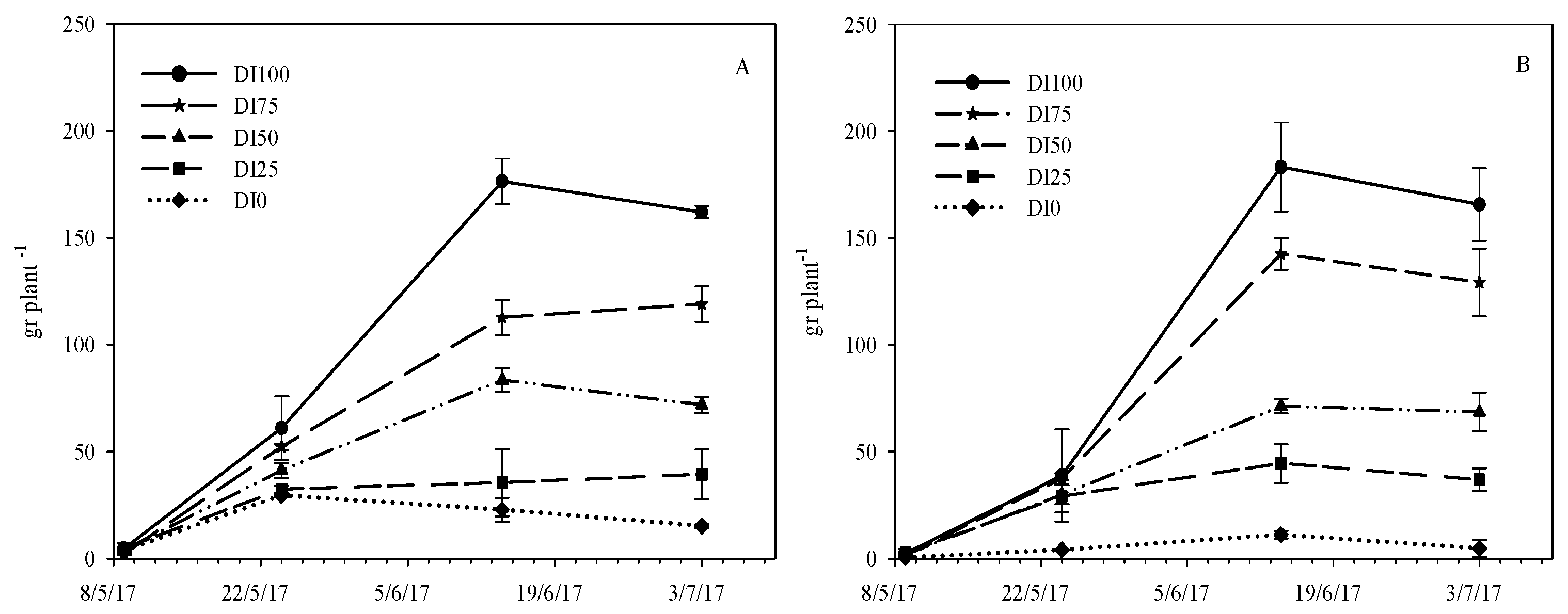
Figure 2.
Accumulation of total dry matter during the crop cycle as affected by five irrigation levels (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) in the two environments (PE = high temperatures (A) and OF = ordinary temperatures (B)).
Figure 2.
Accumulation of total dry matter during the crop cycle as affected by five irrigation levels (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) in the two environments (PE = high temperatures (A) and OF = ordinary temperatures (B)).
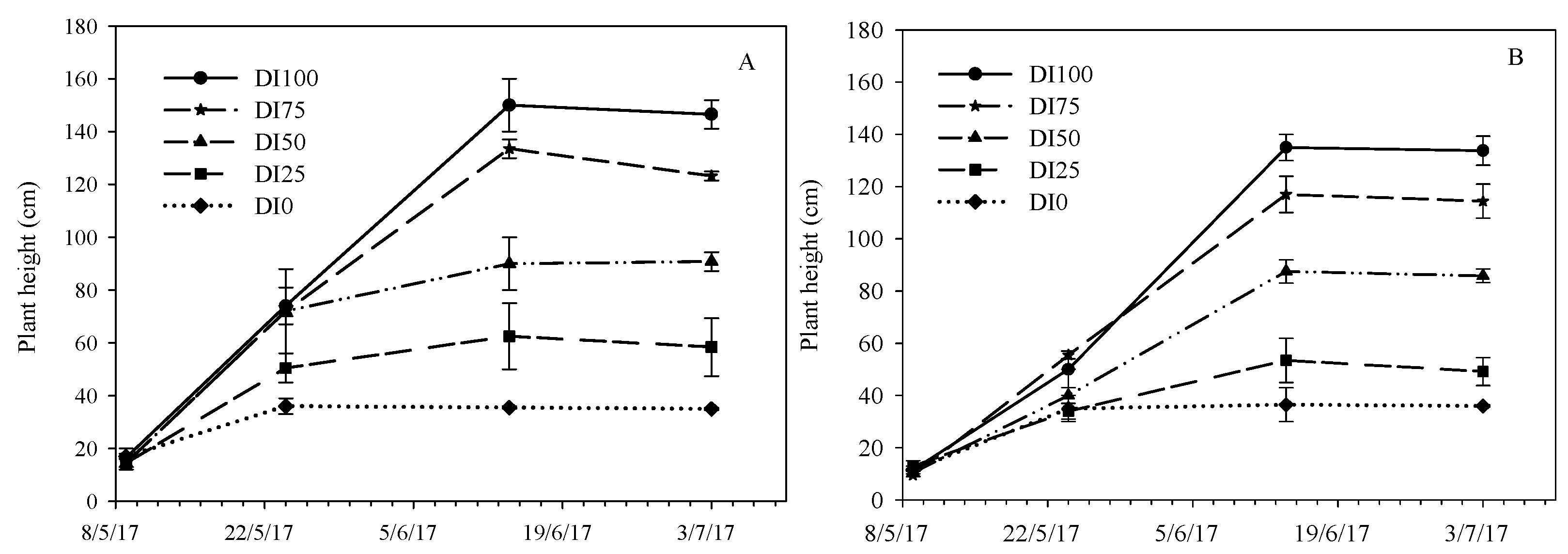
Figure 3.
Plant height during the crop cycle as affected by five irrigation levels (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) in the two environments (PE = high temperatures (A) and OF = ordinary temperatures (B)).
Figure 3.
Plant height during the crop cycle as affected by five irrigation levels (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) in the two environments (PE = high temperatures (A) and OF = ordinary temperatures (B)).
Figure 4.
Daily course of leaf water potential of maize potted plants grown under four irrigation levels (DI100, DI75, DI50, and DI25) in high-temperature (A) or ordinary temperature (B) conditions. Different letters within each column indicate significant differences according to Duncan’s multiple range test (p ≤ 0.05).
Figure 4.
Daily course of leaf water potential of maize potted plants grown under four irrigation levels (DI100, DI75, DI50, and DI25) in high-temperature (A) or ordinary temperature (B) conditions. Different letters within each column indicate significant differences according to Duncan’s multiple range test (p ≤ 0.05).
Table 1.
Water use of the irrigation treatments (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) during the whole cycle under the two environments (PE = high temperatures and OF = ordinary temperatures).
Table 1.
Water use of the irrigation treatments (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) during the whole cycle under the two environments (PE = high temperatures and OF = ordinary temperatures).
| Irrigation | Water Use (m3 ha−1) |
|---|
| PE | OF |
|---|
| DI100 | 3964.4 | 3314.9 |
| DI75 | 2978.5 | 2497.5 |
| DI50 | 1997.3 | 1680.1 |
| DI25 | 1011.4 | 862.7 |
| DI0 | 45.3 | 45.3 |
Table 2.
Physical and chemical properties of the test soil.
Table 2.
Physical and chemical properties of the test soil.
| Soil Properties | Units | Mean Values |
|---|
| Coarse sand | % | 38.6 |
| Fine sand | % | 40.5 |
| Silt | | 14.3 |
| Clay | | 6.6 |
| N—total (Kjeldahl method) | % | 0.16 |
| P2O5 (Olsen method) | ppm | 312.8 |
| K2O (Tetraphenylborate method) | ppm | 620.7 |
| Organic matter (Bichromate method) | % | 3.57 |
| NO3-N | ppm | 32.73 |
| NH4-N | ppm | 6.05 |
| pH | | 7.14 |
| Electrical conductivity | dS m−1 | 0.32 |
Table 3.
Interaction between environmental conditions (PE = high temperatures and OF = ordinary temperatures) and deficit irrigation application (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) on green leaf number, LAI (leaf area index), LA (leaf area), and ALA (average leaf area) at flowering.
Table 3.
Interaction between environmental conditions (PE = high temperatures and OF = ordinary temperatures) and deficit irrigation application (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) on green leaf number, LAI (leaf area index), LA (leaf area), and ALA (average leaf area) at flowering.
| Irrigation | Leaves (n pt−1) | LAI (m2 m−2) | LA (cm2 pt−1) | ALA (cm−2 leaf−1) |
|---|
| PE | OF | PE | OF | PE | OF | PE | OF |
|---|
| DI100 | 10.5 b | 11.0 b | 4.0 ab | 3.9 ab | 281.7 a | 263.2 b | 26.8 a | 23.9 b |
| DI75 | 11.0 b | 12.0 a | 3.9 ab | 4.1 a | 260.2 bc | 253.0 c | 23.7 b | 21.1 c |
| DI50 | 10.5 d | 12.0 a | 2.8 d | 3.8 b | 194.5 e | 235.5 d | 18.5 de | 19.6 cd |
| DI25 | 10.5 d | 12.0 a | 1.8 e | 3.1 c | 127.0 g | 191.6 e | 12.1 f | 16.0 e |
| DI0 | 7.5 c | 7.5 c | 1.3 f | 1.6 e | 121.3 g | 160.4 f | 16.2 e | 21.4 bc |
| Significance | | | | | |
| Irrigation (DI) | * | ** | ** | * |
| Environment (E) | * | * | * | * |
| DI × E | ** | * | ** | * |
Table 4.
Percentage incidence of different parts of plants in total dry matter for the five irrigation levels (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) in the PE = high temperatures.
Table 4.
Percentage incidence of different parts of plants in total dry matter for the five irrigation levels (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) in the PE = high temperatures.
| Irrigation | Stem | Green Leaves | Yellow Leaves | Ears |
|---|
| % Dry Matter |
|---|
| DI100 | 23.70 b | 10.45 a | 2.65 c | 63.20 a |
| DI75 | 27.00 b | 11.63 a | 3.31 c | 58.06 a |
| DI50 | 45.60 a | 8.28 a | 13.56 b | 32.57 b |
| DI25 | 45.20 a | 11.70 a | 18.60 b | 24.51 c |
| DI0 | 16.86 c | 0.00 b | 83.14 a | 0.00 d |
| Significance | | | | |
| Irrigation (DI) | ** | ** | ** | ** |
| Environment (E) | NS | NS | NS | NS |
| DI × E | NS | NS | NS | NS |
Table 5.
Percentage incidence of different parts of plants in total dry matter for the five irrigation levels (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) in the OF = ordinary temperatures.
Table 5.
Percentage incidence of different parts of plants in total dry matter for the five irrigation levels (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) in the OF = ordinary temperatures.
| Irrigation | Stem | Green Leaves | Yellow Leaves | Ears |
|---|
| % Dry Matter |
|---|
| DI100 | 26.35 c | 9.72 b | 5.75 c | 58.18 a |
| DI75 | 29.16 bc | 7.05 b | 8.92 c | 54.88 a |
| DI50 | 43.34 a | 10.19 b | 16.48 b | 30.00 b |
| DI25 | 47.73 a | 16.80 a | 20.46 b | 15.01 c |
| DI0 | 33.10 b | 13.88 a | 53.02 a | 0.00 d |
| Significance | | | | |
| Irrigation (DI) | ** | ** | ** | ** |
| Environment (E) | NS | NS | NS | NS |
| DI × E | NS | NS | NS | NS |
Table 6.
Interaction between environment (PE = high temperatures and OF = ordinary temperatures) and water treatment (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) on yield, plant height, and number of ears.
Table 6.
Interaction between environment (PE = high temperatures and OF = ordinary temperatures) and water treatment (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) on yield, plant height, and number of ears.
| Irrigation | Yield (t ha−1) | Plant Height (cm) | Ears (No. m−2) |
|---|
| PE | OF | PE | OF | PE | OF |
|---|
| DI100 | 9.9 a | 8.5 b | 146.6 a | 133.8 ab | 13.0 a | 11.0 b |
| DI75 | 5.8 c | 6.7 c | 123.2 bc | 114.4 c | 10.0 bc | 9.0 cd |
| DI50 | 1.7 d | 1.7 d | 90.8 d | 78.8 e | 8.0 de | 7.0 e |
| DI25 | 0.8 de | 0.4 e | 58.4 f | 49.2 f | 1.0 f | 1.0 f |
| DI0 | 0.0 e | 0.0 e | 35.7 g | 36.7 g | 0.0 f | 0.0 f |
| Significance | | | |
| Irrigation (DI) | ** | ** | ** |
| Environment (E) | NS | NS | NS |
| DI × E | ** | ** | ** |
Table 7.
Interaction between environment (PE = high temperatures and OF = ordinary temperatures) and water treatment (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) on basal diameter, length, and fertile part (F.P.—% on total length) of ears.
Table 7.
Interaction between environment (PE = high temperatures and OF = ordinary temperatures) and water treatment (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) on basal diameter, length, and fertile part (F.P.—% on total length) of ears.
| Irrigation | Basal Diameter (cm) | Length (cm) | F.P. (% on Total Length) |
|---|
| PE | OF | PE | OF | PE | OF |
|---|
| DI100 | 3.2 b | 3.7 a | 17.0 | 16.5 | 76.0 a | 77.4 a |
| DI75 | 3.1 b | 3.5 ab | 15.3 | 15.3 | 73.9 b | 75.7 ab |
| DI50 | 1.9 c | 2.2 c | 10.7 | 10.4 | 54.2 d | 67.5 c |
| DI25 | 1.3 d | 1.2 d | 5.0 | 4.2 | 27.8 e | 26.7 e |
| DI0 | nd | 0.6 e | nd | 2.4 | nd | 0.0 f |
| Significance | | | |
| Irrigation (DI) | ** | ** | ** |
| Environment (E) | NS | NS | NS |
| DI × E | * | NS | * |
Table 8.
Interaction between environment (PE = high temperatures and OF = ordinary temperatures) and water treatment (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) on water use efficiency (WUE).
Table 8.
Interaction between environment (PE = high temperatures and OF = ordinary temperatures) and water treatment (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) on water use efficiency (WUE).
| Irrigation | WUE (kg m−3) |
|---|
| PE | OF |
|---|
| DI100 | 2.49 a | 2.56 a |
| DI75 | 1.94 b | 2.70 a |
| DI50 | 0.86 c | 1.02 c |
| DI25 | 0.82 c | 0.90 c |
| DI0 | - | - |
| Significance | |
| Irrigation (DI) | ** |
| Environment (E) | * |
| DI × E | * |
Table 9.
Interaction between environment (PE = high temperatures and OF = ordinary temperatures) and water treatment (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) on total nitrogen, nitrogen use efficiency (NUE), and percentage protein into grain.
Table 9.
Interaction between environment (PE = high temperatures and OF = ordinary temperatures) and water treatment (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) on total nitrogen, nitrogen use efficiency (NUE), and percentage protein into grain.
| Irrigation | N (%) | NUE (t kg−1) | Protein (%) |
|---|
| PE | OF | PE | OF | PE | OF |
|---|
| DI100 | 4.75 f | 5.11 de | 0.062 a | 0.053 b | 10.75 e | 12.00 d |
| DI75 | 4.91 ef | 5.32 d | 0.036 d | 0.042 c | 13.20 c | 13.13 c |
| DI50 | 6.04 c | 6.20 bc | 0.011 e | 0.011 e | 13.13 c | 13.75 b |
| DI25 | 6.71 a | 6.80 a | 0.005 f | 0.003 f | 13.75 b | 14.94 a |
| DI0 | 6.48 ab | 6.34 bc | nd | nd | nd | nd |
| Significance | | | | | | |
| Irrigation (DI) | * | ** | * |
| Environment (E) | NS | NS | * |
| DI × E | ** | ** | * |
Table 10.
Main effects of deficit irrigation application treatment (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) and environmental conditions (PE = high temperatures and OF = ordinary temperatures) on leaf gas exchange and midday leaf water potential of maize potted plants (13 June 2017).
Table 10.
Main effects of deficit irrigation application treatment (DI100 = 100%, DI75 = 75%, DI50 = 50%, DI25 = 25%, and DI0 = 0% restoration of water lost by evapotranspiration) and environmental conditions (PE = high temperatures and OF = ordinary temperatures) on leaf gas exchange and midday leaf water potential of maize potted plants (13 June 2017).
| Treatments | Pn | Cì | gs | E | WUEi | ψlmd |
|---|
| µmol m−2 s−1 | µmol mol−1 | mmol m−2 s−1 | mol m−2 s−1 | µmolCO2 molH2O−1 | MPa |
|---|
| Irrigation (DI) | | | | | | |
| DI100 | 24.75 a | 25.00 c | 165.07 a | 4.54 a | 151.93 ab | −1.69 a |
| DI75 | 19.85 b | 24.63 c | 126.98 b | 3.75 b | 162.61 ab | −1.72 a |
| DI50 | 11.05 c | 58.84 b | 65.78 c | 2.34 c | 176.49 a | −1.93 ab |
| DI25 | 3.36 d | 170.18 a | 19.73 d | 0.88 d | 126.35 b | −2.22 b |
| DI0 | nd | nd | nd | nd | nd | −2.97 c |
| Environment (E) | | | | | | |
| PE | 16.24 a | 76.39 a | 105.74 a | 2.72 a | 145.99 a | −2.23 a |
| OF | 13.26 b | 62.93 a | 83.04 b | 3.04 a | 162.69 a | −1.99 b |
| Significance | | | | | | |
| DI | *** | *** | *** | *** | * | *** |
| E | * | NS | * | NS | NS | * |
| DI × E | NS | NS | NS | NS | NS | NS |











{kind=link}
{kind=link}
{kind=link}
{kind=link}