
Bio-Mitigation of Carbon Dioxide Using Desmodesmus sp. in the Custom-Designed Pilot-Scale Loop Photobioreactor

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media Preparation, Microalgae Strain, Culture Conditions, and Inoculum Preparation
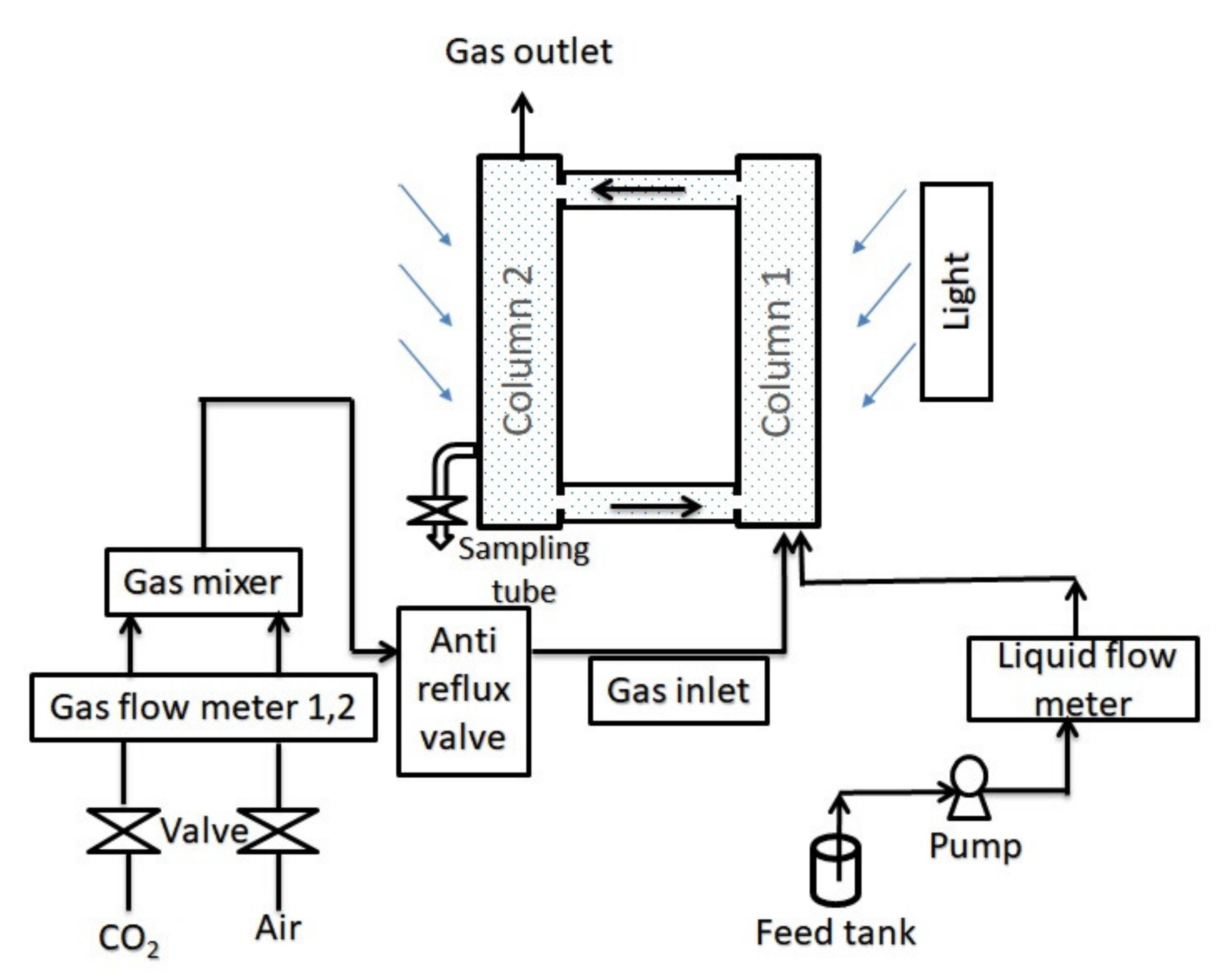
2.2. Experimental Setup
2.3. Experimental Procedure
2.4. Measurement of Biomass Growth Rate
2.5. Determination of Growth Kinetic Parameters
2.6. Determination of Chlorophyll Content
2.7. Biochemical Compositional Analysis
2.7.1. Analysis of Total Carbohydrate (CHO) Content
2.7.2. Analysis of Total Protein Content
2.7.3. Analysis of Total Lipid Content
3. Results and Discussion
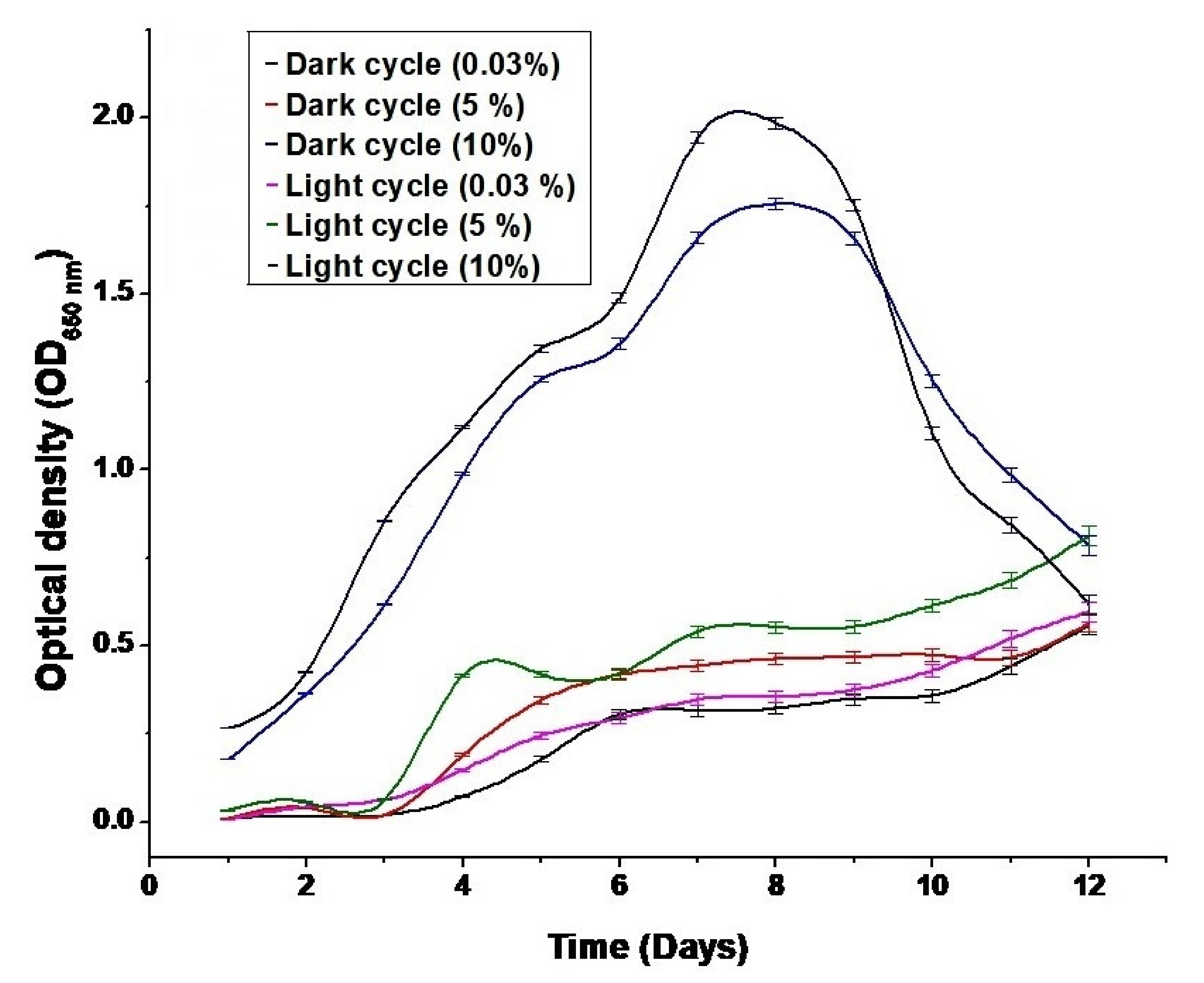
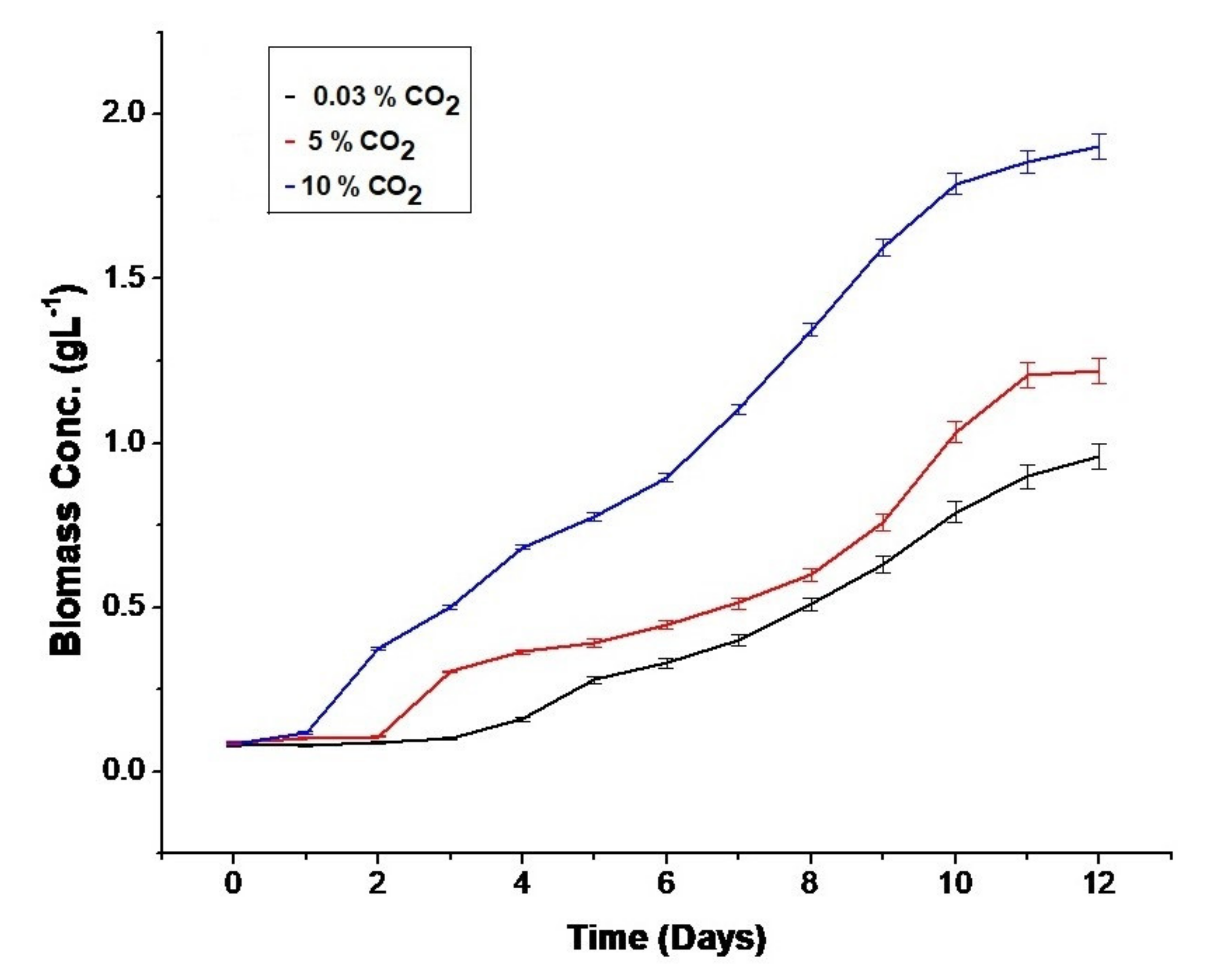
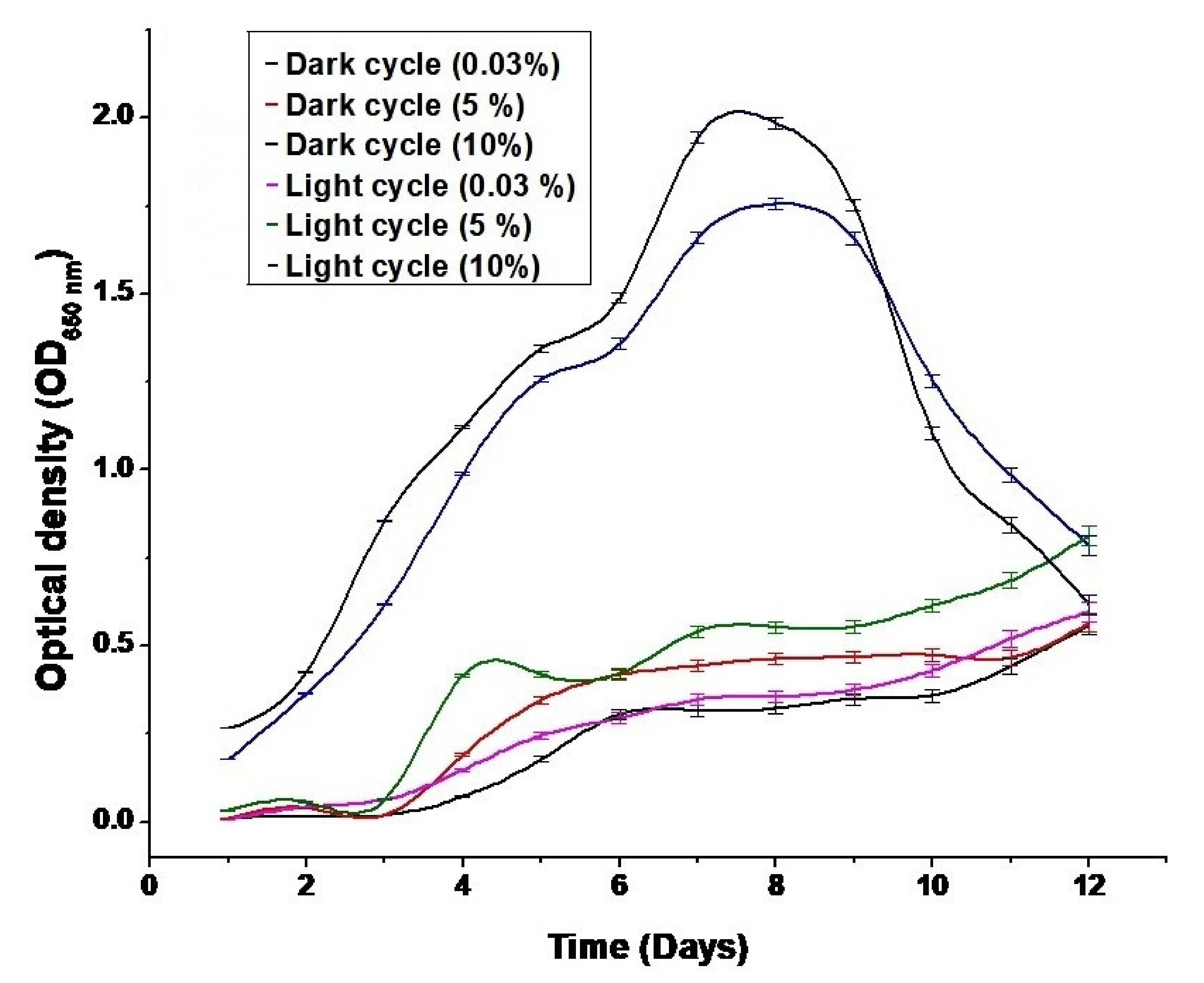
3.1. Effect of CO2 Concentration on Biomass Growth Rate and Optical Density Values
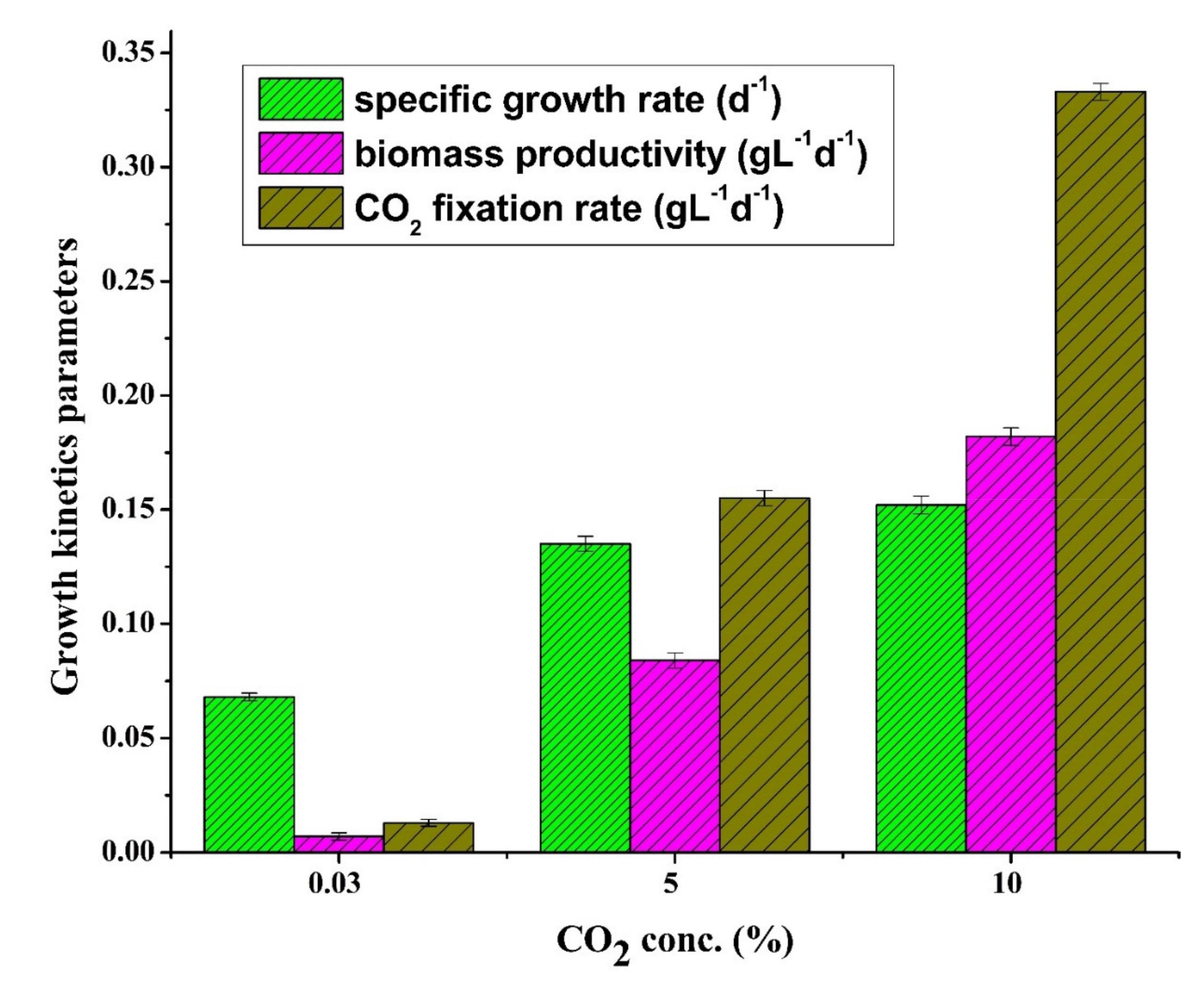
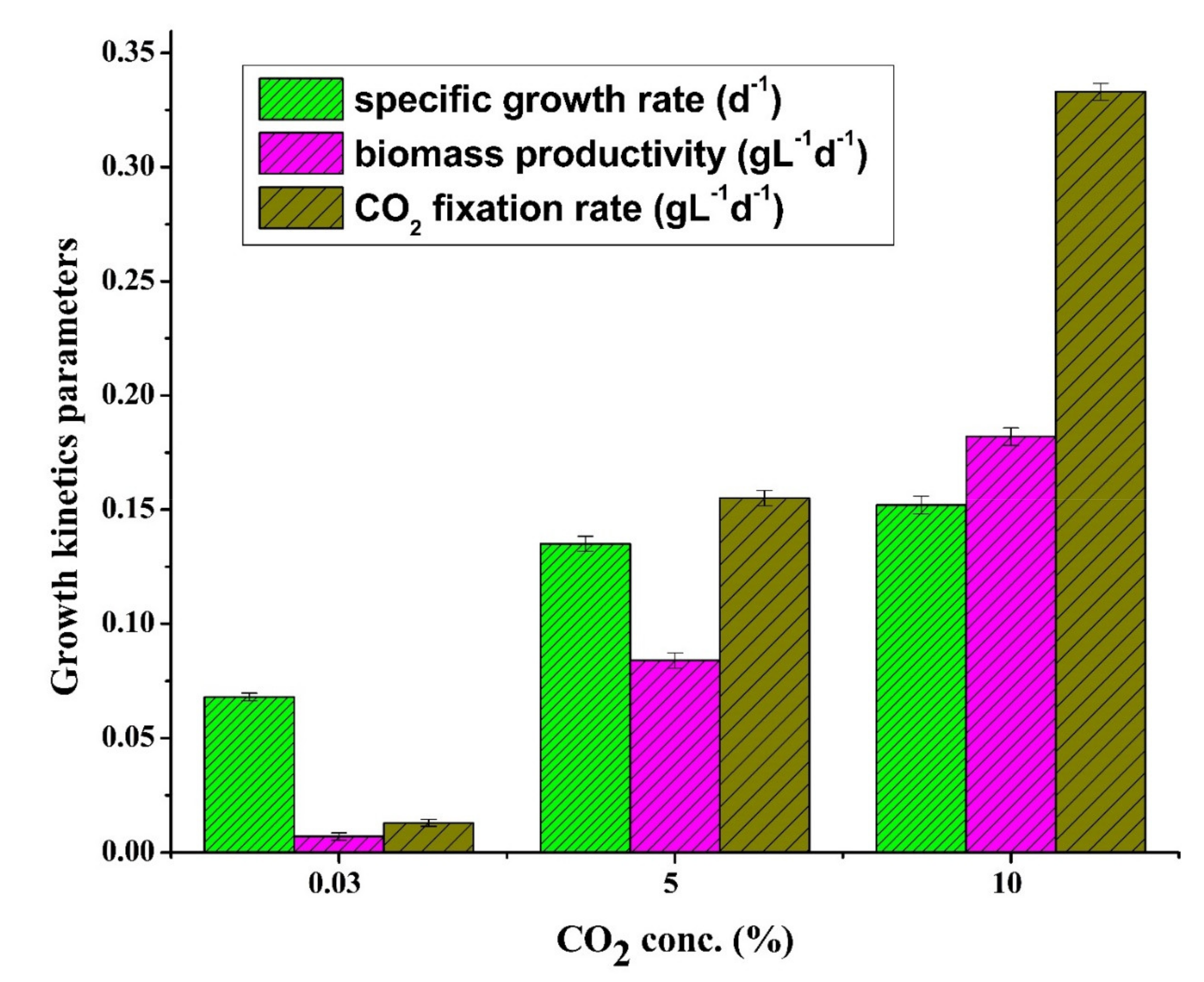
3.2. Effects of CO2 Concentration on Growth Kinetic Parameters
3.2.1. Specific Growth Rate
3.2.2. Biomass Productivity
3.2.3. CO2 Fixation Rate
3.3. Effect of CO2 Concentration on Biochemical Composition of Desmodesmus sp.
4. Performance Comparison of Loop Bioreactor
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramanathan, V. The greenhouse theory of climate change: A test by an inadvertent global experiment. Science 1988, 240, 293–300. [Google Scholar] [CrossRef]
- Mendiara, T.; García-Labiano, F.; Abad, A.; Gayán, P.; de Diego, L.F.; Izquierdo, M.; Adánez, J. Negative CO2 emissions through the use of biofuels in chemical looping technology: A review. Appl. Energy 2018, 232, 657–684. [Google Scholar] [CrossRef]
- Sharma, J.; Kumar, S.; Kumar, V.; Malyan, S.K.; Mathimani, T.; Bishnoi, N.R.; Pugazhendhi, A. Upgrading of microalgal consortia with CO2 from fermentation of wheat straw for the phycoremediation of domestic wastewater. Bioresour. Technol. 2020, 305, 123063. [Google Scholar] [CrossRef]
- Agriculture, Agro-biodiversity and Climate Change. Available online: https://www.un.org/en/ecosoc/docs/pdfs/agriculture.pdf (accessed on 27 August 2021).
- Calabrò, P.S. Greenhouse gases emission from municipal waste management: The role of separate collection. Waste Manag. 2009, 29, 2178–2187. [Google Scholar] [CrossRef]
- Indian Network for Climate Change Assessment (INCCA). India: Green House Gas Emissions 2007; Ministry of Environment and Forests (MoEF), Government of India: New Delhi, India, 2010. [Google Scholar]
- Sources of Greenhouse Gas Emissions. Available online: https://www.epa.gov/ghgemissions/sources-greenhouse-gas-emissions (accessed on 27 August 2021).
- Chen, Y.C.; Lo, S.L. Evaluation of greenhouse gas emissions for several municipal solid waste management strategies. J. Clean Prod. 2016, 113, 606–612. [Google Scholar] [CrossRef]
- Vetter, S.H.; Sapkota, T.; Hillier, J.; Stirling, C.M.; Macdiarmid, J.I.; Aleksandrowicz, L.; Green, R.; Joy, E.J.; Dangour, A.D.; Smith, P. Greenhouse gas emissions from agricultural food production to supply Indian diets: Implications for climate change mitigation. Agric. Ecosyst. Environ. 2017, 237, 234–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahid, A.; Malik, S.; Zhu, H.; Xu, J.; Nawaz, M.Z.; Nawaz, S.; Alam, A.; Mehmood, M.A. Cultivating microalgae in wastewater for biomass production, pollutant removal, and atmospheric carbon mitigation; a review. Sci. Total Environ. 2020, 704, 135303. [Google Scholar] [CrossRef]
- Earth System Research Laboratory, Global Monitoring, DivisionNational Oceanic and Atmospheric Administration, U.S. Department of Commerce. The NOAA Earth System Research Laboratory, Gl. 2020. Available online: http://www.esrl.noaa.gov/gmd/ccgg/trends (accessed on 5 January 2020).
- Solomon, S.; Plattner, G.K.; Knutti, R.; Friedlingstein, P. Irreversible climate change due to carbon dioxide emissions. Proc. Natl. Acad. Sci. USA 2009, 106, 1704–1709. [Google Scholar] [CrossRef] [Green Version]
- Anand, A.; Raghuvanshi, S.; Gupta, S. Trends in carbon dioxide (CO2) fixation by microbial cultivations. Curr. Sustain. Energy Rep. 2020, 7, 40–47. [Google Scholar] [CrossRef]
- De Silva, G.P.D.; Ranjith, P.G.; Perera, M.S.A. Geochemical aspects of CO2 sequestration in deep saline aquifers: A review. Fuel 2015, 155, 128–143. [Google Scholar] [CrossRef]
- IPCC (Intergovernmental Panel on Climate Change). Climate Change 2013: The Physical Science Basis; Working Group I Contribution to the IPCC Fifth Assessment Report; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Seyed Hosseini, N.; Shang, H.; Scott, J.A. Biosequestration of industrial off-gas CO2 for enhanced lipid productivity in open microalgae cultivation systems. Renew Sustain. Energy Rev. 2018, 92, 458–469. [Google Scholar] [CrossRef]
- Barahoei, M.; Hatamipour, M.S.; Afsharzadeh, S. CO2 capturing by chlorella vulgaris in a bubble column photo-bioreactor; Effect of bubble size on CO2 removal and growth rate. J. CO2 Util. 2020, 37, 9–19. [Google Scholar] [CrossRef]
- Khandelwal, A.; Anand, A.; Raghuvanshi, S.; Gupta, S. Integrated approach for microbial carbon dioxide (CO2) fixation process and wastewater treatment for the production of hydrocarbons: Experimental studies. J Environ. Chem. Eng. 2021, 9, 105116. [Google Scholar] [CrossRef]
- Ho, S.H.; Chen, C.Y.; Lee, D.J.; Chang, J.S. Perspectives on microalgal CO2 emission mitigation systems—A review. Biotechnol. Adv. 2011, 29, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Nocito, F.; Dibenedetto, A. Atmospheric CO2 mitigation technologies: Carbon capture utilization and storage. Curr. Opin. Green Sustain. Chem. 2020, 21, 34–43. [Google Scholar] [CrossRef]
- Könst, P.; Mireles, I.H.; Van Der Stel, R.; Van Os, P.; Goetheer, E. Integrated system for capturing CO2 as feedstock for algae production. Energy Procedia 2017, 114, 7126–7132. [Google Scholar] [CrossRef]
- Farrelly, D.J.; Everard, C.D.; Fagan, C.C.; McDonnell, K.P. Carbon sequestration and the role of biological carbon mitigation: A review. Renew Sustain. Energy Rev 2013, 21, 712–727. [Google Scholar] [CrossRef]
- Salbitani, G.; Barone, C.M.A.; Carfagna, S. Effect of bicarbonate on growth of the oleaginous microalga Botryococcus braunii. Int. J. Plant Biol. 2019, 10, 35–37. [Google Scholar] [CrossRef] [Green Version]
- Mountourakis, F.; Papazi, A.; Kotzabasis, K. The microalga chlorella vulgaris as a natural bioenergetic system for effective CO2 mitigation—New perspectives against global warming. Symmetry 2021, 13, 997. [Google Scholar] [CrossRef]
- Anguselvi, V.; Ebhin Masto, R.; Mukherjee, A.; Kumar Singh, P. CO2 Capture for industries by algae. Algae 2019, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Alami, A.H.; Tawalbeh, M.; Alasad, S.; Ali, M.; Alshamsi, M.; Aljaghoub, H. Cultivation of Nannochloropsis algae for simultaneous biomass applications and carbon dioxide capture. Energy Sources Part A Recover. Util Environ. Eff. 2021, 1–12. [Google Scholar] [CrossRef]
- Ullmann, J.; Grimm, D. Algae and their potential for a future bioeconomy, landless food production, and the socio-economic impact of an algae industry. Org. Agric. 2021, 11, 261–267. [Google Scholar] [CrossRef]
- Wang, B.; Li, Y.; Wu, N.; Lan, C.Q. CO2 bio-mitigation using microalgae. Appl. Microbiol. Biotechnol. 2008, 79, 707–718. [Google Scholar] [CrossRef]
- Leong, Y.K.; Chew, K.W.; Chen, W.H.; Chang, J.S.; Show, P.L. Reuniting the biogeochemistry of algae for a low-carbon circular bioeconomy. Trends Plant Sci. 2021, 26, 729–740. [Google Scholar] [CrossRef]
- Ho, S.H.; Chen, C.Y.; Yeh, K.L.; Chen, W.M.; Lin, C.Y.; Chang, J.S. Characterization of photosynthetic carbon dioxide fixation ability of indigenous Scenedesmus obliquus isolates. Biochem. Eng. J. 2010, 53, 57–62. [Google Scholar] [CrossRef]
- Nayak, M.; Karemore, A.; Sen, R. Sustainable valorization of flue gas CO2 and wastewater for the production of microalgal biomass as a biofuel feedstock in closed and open reactor systems. RSC Adv. 2016, 6, 9111–91120. [Google Scholar] [CrossRef]
- Lam, M.K.; Lee, K.T. Renewable and sustainable bioenergies production from palm oil mill effluent (POME): Win-win strategies toward better environmental protection. Biotechnol. Adv. 2011, 29, 124–141. [Google Scholar] [CrossRef] [PubMed]
- Ugwu, C.U.; Aoyagi, H.; Uchiyama, H. Photobioreactors for mass cultivation of algae. Bioresour. Technol. 2008, 99, 4021–4028. [Google Scholar] [CrossRef]
- Achinas, S.; Achinas, V.; Euverink, G.J.W. Microbiology and biochemistry of anaerobic digesters: A brief overview. In Bioreactors: Sustainable Design and Industrial Applications in Mitigation of GHG Emissions, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 17–26. [Google Scholar] [CrossRef]
- Kumar, K.; Dasgupta, C.N.; Nayak, B.; Lindblad, P.; Das, D. Development of suitable photobioreactors for CO2 sequestration addressing global warming using green algae and cyanobacteria. Bioresour. Technol. 2011, 102, 4945–4953. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Jiang, F.; Wang, L.; Yang, C. Design of photobioreactors for mass cultivation of photosynthetic organisms. Engineering 2017, 3, 318–329. [Google Scholar] [CrossRef]
- Sharma, A.K.; Tripathi, K. Loop reactors—Advancing reactors realm: A review. Reactors 2016. [Google Scholar]
- Coker, A.K. Industrial and laboratory reactors—Chemical reaction hazards and process integration of reactors. Ludwig’s Appl. Process. Des. Chem. Petrochem. Plants 2015, 1095–1208. [Google Scholar] [CrossRef]
- Nagappan, S.; Verma, S.K. Growth model for raceway pond cultivation of Desmodesmus sp. MCC34 isolated from a local water body. Eng. Life Sci. 2016, 16, 45–52. [Google Scholar] [CrossRef]
- Abou-Shanab, R.A.I.; Ji, M.K.; Kim, H.C.; Paeng, K.J.; Jeon, B.H. Microalgal species growing on piggery wastewater as a valuable candidate for nutrient removal and biodiesel production. J. Environ. Manag. 2013, 115, 257–264. [Google Scholar] [CrossRef]
- Sundaram, S.; Thakur, I.S. Biosurfactant production by a CO2 sequestering Bacillus sp. strain ISTS2. Bioresour. Technol. 2015, 188, 247–250. [Google Scholar] [CrossRef]
- Abreu, A.P.; Fernandes, B.; Vicente, A.A.; Teixeira, J.; Dragone, G. Mixotrophic cultivation of Chlorella vulgaris using industrial dairy waste as organic carbon source. Bioresour. Technol. 2012, 118, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Anjos, M.; Fernandes, B.D.; Vicente, A.A.; Teixeira, J.A.; Dragone, G. Optimization of CO2 bio-mitigation by Chlorella vulgaris. Bioresour. Technol. 2013, 139, 149–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Roy, A.S.; Mohanty, K.; Ghoshal, A.K. Enhanced CO2 sequestration by a novel microalga: Scenedesmus obliquus SA1 isolated from bio-diversity hotspot region of Assam, India. Bioresour. Technol. 2013, 143, 369–377. [Google Scholar] [CrossRef]
- Tang, D.; Han, W.; Li, P.; Miao, X.; Zhong, J. CO2 biofixation and fatty acid composition of Scenedesmus obliquus and Chlorella pyrenoidosa in response to different CO2 levels. Bioresour. Technol. 2011, 102, 3071–3076. [Google Scholar] [CrossRef] [PubMed]
- Mohsenpour, S.F.; Willoughby, N. Effect of CO2 aeration on cultivation of microalgae in luminescent photobioreactors. Biomass Bioenergy 2016, 85, 168–177. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Mecozzi, M. Estimation of total carbohydrate amount in environmental samples by the phenol-sulphuric acid method assisted by multivariate calibration. Chemom. Intell. Lab. Syst. 2005, 79, 84–90. [Google Scholar] [CrossRef]
- Wang, H.; Nche-Fambo, F.A.; Yu, Z.; Chen, F. Using microalgal communities for high CO2-tolerant strain selection. Algal Res. 2018, 35, 253–261. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Varshney, P.; Beardall, J.; Bhattacharya, S.; Wangikar, P.P. Isolation and biochemical characterisation of two thermophilic green algal species- Asterarcys quadricellulare and Chlorella sorokiniana, which are tolerant to high levels of carbon dioxide and nitric oxide. Algal Res. 2018, 30, 28–37. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Varshney, P.; Sohoni, S.; Wangikar, P.P.; Beardall, J. Effect of high CO2 concentrations on the growth and macromolecular composition of a heat- and high-light-tolerant microalga. J. Appl. Phycol. 2016, 28, 2631–2640. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, G.; Karemore, A.; Dash, S.K.; Sen, R. Performance evaluation of a green process for microalgal CO2 sequestration in closed photobioreactor using flue gas generated in-situ. Bioresour. Technol. 2015, 191, 399–406. [Google Scholar] [CrossRef]
- Kaiwan-Arporn, P.; Hai, P.D.; Thu, N.T.; Annachhatre, A.P. Cultivation of cyanobacteria for extraction of lipids. Biomass Bioenergy 2012, 44, 142–149. [Google Scholar] [CrossRef]
- Wang, X.W.; Liang, J.R.; Luo, C.S.; Chen, C.P.; Gao, Y.H. Biomass, total lipid production, and fatty acid composition of the marine diatom Chaetoceros muelleri in response to different CO2 levels. Bioresour. Technol. 2014, 161, 124–130. [Google Scholar] [CrossRef]
- Widdel, F. Theory and measurement of bacterial growth. MethodsX 2019, 6, 2248–2257. [Google Scholar] [CrossRef]
- Chae, S.R.; Hwang, E.J.; Shin, H.S. Single cell protein production of Euglena gracilis and carbon dioxide fixation in an innovative photo-bioreactor. Bioresour. Technol. 2006, 97, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Chiarini, A.; Quadrio, M. The light/dark cycle of microalgae in a thin-layer photobioreactor. J. Appl. Phycol. 2021, 33, 183–195. [Google Scholar] [CrossRef]
- Xue, S.; Su, Z.; Cong, W. Growth of Spirulina platensis enhanced under intermittent illumination. J. Biotechnol. 2011, 151, 271–277. [Google Scholar] [CrossRef]
- Toledo-Cervantes, A.; Morales, M.; Novelo, E.; Revah, S. Carbon dioxide fixation and lipid storage by Scenedesmus obtusiusculus. Bioresour. Technol. 2013, 130, 652–658. [Google Scholar] [CrossRef]
- Chiang, C.L.; Lee, C.M.; Chen, P.C. Utilization of the cyanobacteria Anabaena sp. CH1 in biological carbon dioxide mitigation processes. Bioresour. Technol. 2011, 102, 5400–5405. [Google Scholar] [CrossRef]
- Yang, Q.; Li, H.; Wang, D.; Zhang, X.; Guo, X.; Pu, S.; Guo, R.; Chen, J. Utilization of chemical wastewater for CO2 emission reduction: Purified terephthalic acid (PTA) wastewater-mediated culture of microalgae for CO2 bio-capture. Appl. Energy 2020, 276, 115502. [Google Scholar] [CrossRef]
- Chinnasamy, S.; Ramakrishnan, B.; Bhatnagar, A.; Das, K.C. Biomass production potential of a wastewater alga chlorella vulgaris ARC 1 under elevated levels of CO2 and temperature. Int. J. Mol. Sci. 2009, 10, 518–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, M.E.; Sánchez, S.; Jiménez, J.M.; El Yousfi, F.; Muñoz, L. Nitrogen and phosphorus removal from urban wastewater by the microalga Scenedesmus obliquus. Bioresour. Technol. 2000, 73, 263–272. [Google Scholar] [CrossRef]
- Chiu, S.Y.; Kao, C.Y.; Chen, C.H.; Kuan, T.C.; Ong, S.C.; Lin, C.S. Reduction of CO2 by a high-density culture of Chlorella sp. in a semicontinuous photobioreactor. Bioresour. Technol. 2008, 99, 3389–3396. [Google Scholar] [CrossRef]
- Yen, H.W.; Ho, S.H.; Chen, C.Y.; Chang, J.S. CO2, NOx and SOx removal from flue gas via microalgae cultivation: A critical review. Biotechnol. J. 2015, 10, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Ugwu, C.U.; Ogbonna, J.C.; Tanaka, H. Improvement of mass transfer characteristics and productivities of inclined tubular photobioreactors by installation of internal static mixers. Appl. Microbiol. Biotechnol. 2002, 58, 600–607. [Google Scholar] [CrossRef]
- Sydney, E.; Sturm, W.; de Carvalho, J.; Soccol, V.T.; Larroche, C.; Pandey, A.; Soccol, C.R. Potential carbon dioxide fixation by industrially important microalgae. Bioresour. Technol. 2010, 101, 5892–5896. [Google Scholar] [CrossRef]
- Kumar, K.; Banerjee, D.; Das, D. Carbon dioxide sequestration from industrial flue gas by Chlorella sorokiniana. Bioresour. Technol. 2014, 152, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Nayak, M.; Karemore, A.; Sen, R. Performance evaluation of microalgae for concomitant wastewater bioremediation, CO2 biofixation and lipid biosynthesis for biodiesel application. Algal Res. 2016, 16, 216–223. [Google Scholar] [CrossRef]
- Carlozzi, P. Dilution of solar radiation through “culture” lamination in photobioreactor rows facing south-north: A way to improve the efficiency of light utilization by cyanobacteria (Arthrospira platensis). Biotechnol. Bioeng. 2003, 81, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.O.; Acién Fernández, F.G.; Guerrero, E.C.; Rao, K.K.; Grima, E.M. Outdoor helical tubular photobioreactors for microalgal production: Modeling of fluid-dynamics and mass transfer and assessment of biomass productivity. Biotechnol. Bioeng. 2003, 82, 62–73. [Google Scholar] [CrossRef] [PubMed]
- López, M.G.-M.; Sanchez, E.D.R.; López, J.C.; Fernandez, F.G.A.; Sevilla, J.M.F.; Rivas, J.; Guerrero, M.; Grima, E.M. Comparative analysis of the outdoor culture of Haematococcus pluvialis in tubular and bubble column photobioreactors. J. Biotechnol. 2006, 123, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Kargupta, W.; Ganesh, A.; Mukherji, S. Estimation of carbon dioxide sequestration potential of microalgae grown in a batch photobioreactor. Bioresour. Technol. 2015, 180, 370–375. [Google Scholar] [CrossRef]
- Ye, Q.; Cheng, J.; Guo, W.; Xu, J.; Li, K.; Zhou, J. Serial lantern-shaped draft tube enhanced flashing light effect for improving CO2 fixation with microalgae in a gas-lift circumflux column photobioreactor. Bioresour. Technol. 2018, 255, 156–162. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biochemical Composition | Inlet Concentration of CO2 (v/v) | ||
|---|---|---|---|
| 0.03% | 5% | 10% | |
| Total carbohydrates (% DCW) | 14.6 ± 1.5 | 17.2 ± 2.0 | 20.7 ± 2.4 |
| Proteins (% DCW) | 14.4 ± 1.2 | 25 ± 1.1 | 32.3 ± 2.5 |
| Lipids (% DCW) | 15.5 ± 0.5 | 40 ± 2.0 | 42 ± 1.0 |
| Chlorophyll a, b (mg L−1) | 0.12 0.15 | 0.13 0.17 | 0.14 0.19 |
| Species | Cultivation Time (Day) | Mode/ (Volume, L/Working Volume, L) | CO2 conc. (% v/v) | Max. Biomass Produced (XMax) (g L−1) | Biomass Productivity (P) (g L−1 d−1) | CO2 Fixation Rate (RCO2) (g L−1 d−1) | Carbohydrate (% DCW) | Protein (% DCW) | Lipids (% DCW) | References |
|---|---|---|---|---|---|---|---|---|---|---|
| Chlorella sp. | 8 | Column Photobioreactors, (0.8) | 2 | 1.21 | 0.15 | 0.28 | - | - | - | Chiu et al., 2008 [67] |
| Chlorella vulgaris | 15 | Bio Flow fermenter, (11/8) | 10 | 1.94 | 0.13 | 0.25 | 16.74 | 40.95 | 9.95 | Sydney et al., 2010 [70] |
| Scenedesmus obliquus | 6 | Erlenmeyer flask, (0.650) | 10 | 1.84 | 0.15 | 0.29 | - | - | 22 | Tang et al., 2011 [45] |
| Chlorella sorokiniana | 8 | Airlift photobioreactor, (1.4) | 4 | 1.1 | 0.15 | - | - | - | 20.93 | Kumar et al., 2014 [71] |
| Scenedesmus sp. | 7 | Airlift photobioreactor, (0.5) | 2.5 | 1.3 | 0.19 | 0.35 | 10.4 | - | 35.6 | Nayak et al., 2016 [72] |
| Scenedesmus sp. | 7 | Bubble-column photobioreactor, (0.5) | 2.5 | 1.37 | 0.196 | 0.37 | - | - | 33.3 | Nayak et al., 2016 [72] |
| Acutodesmus sp. | 5 | Erlenmeyer flasks, (0.5/0.2) | 20 | 1.65 | - | - | 34.52 | 38.78 | 11.67 | Yadav et al., 2015 [55] |
| A. quadricellulare | 6 | Laboratory scale photobioreactor, (0.8/0.680) | 5 | 1.29 | - | - | 33.4 | 30.3 | 44 | Varshney et al., 2018 [53] |
| Desmodesmus sp. MCC34 | 18 | Raceway pond, (1000) | - | 1.9 | - | - | - | - | 0.103 | Nagappan et al., 2016 [39] |
| Porphyridium cruentum | - | Airlift tubular, (200) | - | 3.0 | 1.50 | - | - | - | - | Yen et al., 2015 [71] |
| Chlorella sorokiniana | - | Inclined tubular, (6.0) | 5 | 1.50 | 1.47 | - | - | - | - | Ugwu et al., 2002 [69] |
| Arthrospira platensis | - | Undular row tubular, (11) | - | - | 2.70 | - | - | - | - | Carlozzi P., 2003 [73] |
| Phaeodactylum tricornutum | 9 | Outdoor helical tubular, (75) | - | 2.95 | 1.40 | - | - | - | - | Hall et al., 2003 [74] |
| Haematococcus pluvialis | 16 | Bubble-column, (55) | - | 1.4 | 0.06 | - | - | - | - | Lopez et al., 2006 [75] |
| Chlorella pyrenoidosa | 1.25 | Tubular batch reactors, (0.660) | 10 | 0.11 | 0.096 | - | - | - | Kargupta et al., 2015 [76] | |
| Chlorella PY-ZU1 | 4.5 | Cylindrical PBR (6) | 15 | - | 0.47 | 0.87 | - | - | - | Ye at al., 2018 [77] |
| Desmodesmus sp. | 12 | Loop photobioreactor, (34/26) | 0.03 | 0.96 ± 0.04 | 0.018 ± 0.002 | 0.013 ± 0.001 | 14.6 ± 1.5 | 14.4 ± 1.2 | 15.5 ± 0.5 | Present study |
| Desmodesmus sp. | 12 | Loop photobioreactor, (34/26) | 5 | 1.219 ± 0.04 | 0.084 ± 0.003 | 0.155 ± 0.003 | 17.2 ± 2.0 | 25 ± 1.1 | 40 ± 2.0 | Present study |
| Desmodesmus sp. | 12 | Loop photobioreactor, (34/26) | 10 | 1.903 ± 0.04 | 0.185 ± 0.004 | 0.333 ± 0.004 | 20.7 ± 2.4 | 32.3 ± 2.5 | 42 ± 1.0 | Present study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anand, A.; Tripathi, K.; Kumar, A.; Gupta, S.; Raghuvanshi, S.; Verma, S.K. Bio-Mitigation of Carbon Dioxide Using Desmodesmus sp. in the Custom-Designed Pilot-Scale Loop Photobioreactor. Sustainability 2021, 13, 9882. https://doi.org/10.3390/su13179882
Anand A, Tripathi K, Kumar A, Gupta S, Raghuvanshi S, Verma SK. Bio-Mitigation of Carbon Dioxide Using Desmodesmus sp. in the Custom-Designed Pilot-Scale Loop Photobioreactor. Sustainability. 2021; 13(17):9882. https://doi.org/10.3390/su13179882
Chicago/Turabian StyleAnand, Abhishek, Kaustubh Tripathi, Amit Kumar, Suresh Gupta, Smita Raghuvanshi, and Sanjay Kumar Verma. 2021. "Bio-Mitigation of Carbon Dioxide Using Desmodesmus sp. in the Custom-Designed Pilot-Scale Loop Photobioreactor" Sustainability 13, no. 17: 9882. https://doi.org/10.3390/su13179882
APA StyleAnand, A., Tripathi, K., Kumar, A., Gupta, S., Raghuvanshi, S., & Verma, S. K. (2021). Bio-Mitigation of Carbon Dioxide Using Desmodesmus sp. in the Custom-Designed Pilot-Scale Loop Photobioreactor. Sustainability, 13(17), 9882. https://doi.org/10.3390/su13179882