
Effect of Methyl Jasmonate, Cytokinin, and Lavender Oil on Antioxidant Enzyme System of Apricot Fruit (Prunus armeniaca L.)

 , , ,
, , ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Material Preparation for Antioxidative Enzyme Assays
2.3. Ascorbate Peroxidase (APX) Activity
2.4. Catalase (CAT) Activity
2.5. Superoxide Dismutase (SOD) Activity
2.6. Lipid Peroxidation
2.7. Statistical Analysis
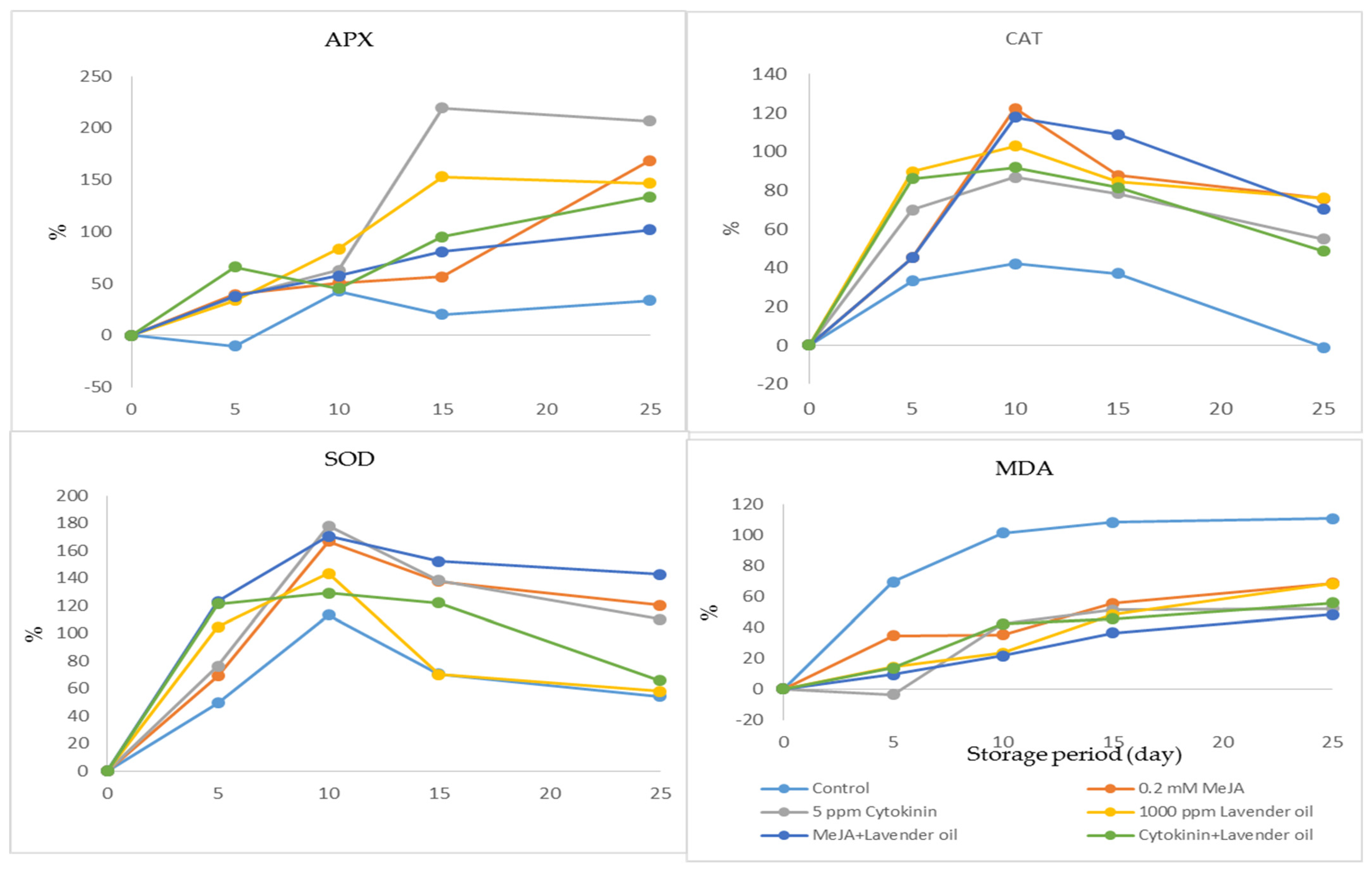
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ercisli, S.; Esitken, A.; Cangi, R.; Sahin, F. Adventitious root formation of kiwifruit in relation to sampling date, IBA and Agrobacterium rubi inoculation. Plant Growth Regul. 2003, 41, 133–137. [Google Scholar] [CrossRef]
- Campbell, O.E.; Merwin, I.A.; Padilla-Zakour, O.I. Characterization and the effect of maturity at harvest on the phenolic and carotenoid content of northeast USA apricot (Prunus armeniaca) varieties. J. Agric. Food Chem. 2013, 61, 12700–12710. [Google Scholar] [CrossRef] [PubMed]
- Dogan, H.; Ercisli, S.; Jurikova, T.; Temim, E.; Leto, A.; Hadziabulic, A.; Tosun, M.; Narmanlioglu, H.K.; Zia-Ul-Haq, M. Physicochemical and antioxidant characteristics of fruits of cape gooseberry (Physalis peruviana L.) from Turkey. Oxid. Commun. 2014, 37, 1005–1014. [Google Scholar]
- Hu, W.; Yang, H.; Tie, W.; Yan, Y.; Ding, Z.; Liu, Y.; Wu, C.; Wang, J.; Reiter, R.J.; Tan, D.X.; et al. Natural variation in banana varieties highlights the role of melatonin in postharvest ripening and quality. J. Agric. Food Chem. 2017, 65, 9987–9994. [Google Scholar] [CrossRef]
- Abd El-Naby, S.K.M.A.; Mohamed, A.A.A.; El-Naggar, Y.I.M. Effect of melatonin, GA3 and NAA on vegetative growth, yield and quality of ‘Canino’ apricot fruits. Acta Sci. Pol. Hortorum Cultus 2019, 18, 167–174. [Google Scholar] [CrossRef]
- Engin, S.P.; Mert, C. The effects of harvesting time on the physicochemical components of aronia berry. Turk. J. Agric. For. 2020, 44, 361–370. [Google Scholar] [CrossRef]
- Karatas, N.; Sengul, M. Some important physicochemical and bioactive characteristics of the main apricot cultivars from Turkey. Turk. J. Agric. For. 2020, 44, 651–661. [Google Scholar] [CrossRef]
- Medina-Santamarina, J.; Zapata, P.J.; Valverde, J.M.; Valero, D.; Serrano, M.; Guillén, F. Melatonin treatment of apricot trees leads to maintenance of fruit quality attributes during storage at chilling and non-chilling temperatures. Agronomy 2021, 11, 917. [Google Scholar] [CrossRef]
- Ercisli, S. Apricot culture in Turkey. Sci. Res. Essays 2009, 4, 715–719. [Google Scholar]
- Gecer, M.K.; Kan, T.; Gundogdu, M.; Ercisli, S.; Ilhan, G.; Sagbas, H.I. Physicochemical characteristics of wild and cultivated apricots (Prunus armeniaca L.) from Aras valley in Turkey. Genet. Resour. Crop Evol. 2020, 67, 935–945. [Google Scholar] [CrossRef]
- FAOSTAT. 2019. Available online: http://www.fao.org/faostat/en/#compare (accessed on 28 January 2021).
- Egea, M.I.; Sánchez-Bel, P.; Martínez-Madrid, M.C.; Flores, F.B.; Romojaro, F. The effect of beta ionization on the antioxidant potential of “Búlida” apricot and its relationship with quality. Postharvest Biol. Technol. 2007, 46, 63–70. [Google Scholar] [CrossRef]
- Cavusoglu, S.; Islek, F.; Yilmaz, N.; Tekin, O. Kayısıda (Prunus armeniaca L.) metil jasmonate, sitokinin ve lavanta yağı uygulamalarının hasat sonrası fizyolojisi üzerine etkileri. Yüzüncü Yıl Univ. Tarm Bilimleri Derg. 2020, 30, 136–146. [Google Scholar]
- Rampáčková, E.; Göttingerová, M.; Gála, P.; Kiss, T.; Ercişli, S.; Nečas, T. Evaluation of Protein and Antioxidant Content in Apricot Kernels as a Sustainable Additional Source of Nutrition. Sustainability 2021, 13, 4742. [Google Scholar] [CrossRef]
- Göttingerová, M.; Kumšta, M.; Nečas, T. Health-benefitting Biologically Active Substances in Edible Apricot Flowers. HortScience 2020, 55, 1372–1377. [Google Scholar] [CrossRef]
- Hossein, M.; Mahmood, G.; Davood, B. Effect of different coatings on post-harvest quality and bioactive compounds of pomegranate (Punica granatum L.) fruits. J. Food Sci. Technol. 2015, 52, 4507–4514. [Google Scholar]
- Nourozi, F.; Sayyari, M. Enrichment of Aloe vera gel with basil seed mucilage preserve bioactive compounds and postharvest quality of apricot fruits. Sci. Hortic. 2020, 262, 109041. [Google Scholar] [CrossRef]
- Sortino, G.; Saletta, F.; Puccio, S.; Scuderi, D.; Allegra, A.; Inglese, P.; Farina, V. Extending the shelf life of white peach fruit with 1-Methylcyclopropene and Aloe arborescens edible coating. Agriculture 2020, 10, 151. [Google Scholar] [CrossRef]
- Vick, B.A.; Zimmerman, D.C. Biosynthesis of jasmonic acid by several plant species. Plant Physiol. 1984, 75, 458–461. [Google Scholar] [CrossRef]
- Gundlach, H.; Muller, M.J.; Kutchan, T.M.; Zenk, M.H. Jasmonic acid is a signal transducer in elicitor-induced plant cell cultures. Proc. Natl. Acad. Sci. USA 1992, 89, 2389–2393. [Google Scholar] [CrossRef]
- Mueller, M.J.; Brodschelm, W.; Spannagl, E.; Zenk, M.H. Signaling in the elicitation process is mediated through the octadecanoid pathway leading to jasmonic acid. Proc. Natl. Acad. Sci. USA 1993, 90, 7490–7494. [Google Scholar] [CrossRef] [PubMed]
- Meir, S.; Philosoph-Hadas, S.; Lurie, S.; Droby, S.; Akerman, M.; Zauberman, G.; Shapiro, B.; Cohen, E.; Fuchs, Y. Reduction of chilling injury in stored avocado, grapefruit, and bell pepper by methyl jasmonate. Can. J. Bot. 1996, 74, 870–874. [Google Scholar] [CrossRef]
- Pérez, A.G.; Sanz, C.; Olías, R.; Olías, J.M. Effect of methyl jasmonate on in vitro strawberry ripening. J. Agric. Food Chem. 1997, 45, 3733–3737. [Google Scholar] [CrossRef]
- Zhu, Z.; Tian, S.P. Resistant responses of tomato fruit treated with exogenous methyl jasmonate to Botrytis cinerea infection. Sci. Hortic. 2012, 142, 38–43. [Google Scholar] [CrossRef]
- Kays, S.J. Postharvest Physiology of Perishable Plant Products; Van Nostrand Reinhold: New York, NY, USA, 1991; p. 507. [Google Scholar]
- Clarke, S.F.; Jameson, P.E.; Downs, C. The influence of 6-benzylaminopurine on post-harvest senescence of floral tissues of broccoli (Brassica oleracea var Italica). Plant Growth Regul. 1994, 14, 21–27. [Google Scholar] [CrossRef]
- Fuller, G.; Kuhnle, J.A.; Corse, J.W.; Mackey, B.E. Use of natural cytokinin’s to extend the storage life of broccoli (Brassica oleracea, Itaica group). J. Am. Soc. Hort. Sci. 1977, 102, 480–484. [Google Scholar]
- Cavusoglu, S.; Halloran, N. The effect of pre-harvest cytokinin application total phenolic content and polyphenoloxidase on post-harvest physiology of cauliflower (Brassica oleracea L. Botrytis). In Proceedings of the XXVIII International Horticultural Congress, Lisboa, Portugal, 22–26 August 2010; p. 114. [Google Scholar]
- Mastromatteo, M.; Conte, A.; Del Nobile, M.A. Combined used of modified atmosphere packaging and natural compounds for food preservation. Food Eng. Rev. 2010, 2, 28–38. [Google Scholar] [CrossRef]
- Deans, S.G.; Ritchie, G. Antimicrobial properties of plant essential oils. Int. J. Food Microbiol. 1987, 5, 165–180. [Google Scholar] [CrossRef]
- Kim, H.Y.; Lee, Y.J.; Hong, K.H.; Kwon, Y.K.; Sim, K.C.; Lee, J.Y.; Cho, H.Y.; Kim, I.S.; Han, S.B.; Lee, C.W.; et al. Isolation of antimicrobial substances from natural products and their preservative effects. Food Sci. Biotechnol. 2001, 10, 59–71. [Google Scholar]
- Alzoreky, N.S.; Nakahara, K. Antimicrobial activity of extracts from some edible plants commonly consumed in Asia. Int. Food Microbial. 2002, 80, 223–230. [Google Scholar] [CrossRef]
- Packiyasothy, E.V.; Kyle, S. Antimicrobial properties of some herb essential oil. Food Aust. 2002, 54, 384–387. [Google Scholar]
- Zaika, L.L. Spice and herbs: Their antimicrobial activity and its determinaton. J. Food Saf. 1988, 9, 97–118. [Google Scholar] [CrossRef]
- Meng, D.; Zhang, Y.; Yang, R.; Wang, J.; Zhang, X.; Sheng, J.; Wang, J.; Fan, Z. Arginase participates in the methyl jasmonate-regulated quality maintenance of postharvest Agaricus bisporus fruit bodies. Postharvest Biol. Technol. 2017, 132, 7–14. [Google Scholar] [CrossRef]
- Jebara, S.; Jebara, M.; Limam, F.; Aouani, M.E. Changes in ascorbate peroxidase, catalase, guaiacol peroxidase and superoxide dismutase activities in common bean (Phaseolus vulgaris) nodules under salt stress. J. Plant Physiol. 2005, 162, 929–936. [Google Scholar] [CrossRef]
- Guneri Bagci, E. Determination of Physiological and Biochemical Parameters Symptomatic for Oxidative Stress in Chickpea (Cicer arietinum L.) Cultivars under Droughtvars under Drought. Ph.D. Thesis, Ankara University, Ankara, Turkey, 2010. [Google Scholar]
- Sun, J.; You, X.; Li, L.; Peng, H.; Su, W.; Li, C.; He, Q.; Liao, F. Effects of a phospholipase D inhibitor on postharvest enzymatic browning and oxidative stress of litchi fruit. Postharvest Biol. Technol. 2011, 62, 288–294. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Zhou, Q.; Ma, C.; Cheng, S.; Wei, B.; Liu, X.; Ji, S. Changes in antioxidative metabolism accompanying pitting development in stored blueberry fruit. Postharvest Biol. Technol. 2014, 88, 88–95. [Google Scholar] [CrossRef]
- Liu, Y.; Yanga, X.; Zhua, S.; Wang, Y. Postharvest application of MeJA and NO reduced chilling injury in cucumber (Cucumis sativus) through inhibition of H2O2 accumulation. Postharvest Biol. Technol. 2016, 119, 77–83. [Google Scholar] [CrossRef]
- Cao, S.; Zheng, Y.; Wang, K.; Rui, H.; Tang, S. Effect of methyl jasmonate on cell wall modification of loquat fruit in relation to chilling injury after harvest. Food Chem. 2009, 118, 641–647. [Google Scholar] [CrossRef]
- Esim, N.; Atici, O. Nitric oxide improves chilling tolerance of maize by affecting apoplastic antioxidative enzymes in leaves. Plant Growth Regul. 2014, 72, 29–38. [Google Scholar] [CrossRef]
- Wu, B.; Guo, Q.; Li, Q.; Ha, Y.; Li, X.; Chen, W. Impact of postharvest nitric oxide treatment on antioxidant enzymes and related genes in banana fruit in response to chilling tolerance. Postharvest Biol. Technol. 2014, 92, 157–163. [Google Scholar] [CrossRef]
- Venkatachalam, K.; Meenune, M. Effect of methyl jasmonate on physiological and biochemical quality changes of longkong fruit under low temperature storage. Fruits 2015, 70, 69–75. [Google Scholar] [CrossRef]
- Mangena, T.; Muyima, N.Y.O. Comparative evaluation of the antimicrobial activities of essential oils of Artemisia afra, Pteronia incana and Rosemarinus officinalis on selected bacteria and yeast strains. Lett. Appl. Microbiol. 1999, 28, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.X.; Duan, L.S.; Tian, X.L.; Wang, B.M.; Eneji, A.E.; Li, Z.H. Coronatinealleviatessalinity stress in cotton by improving the antioxidative defense system and radical-scavenging activity. J. Plant Physiol. 2008, 165, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Wang, Q.; Lv, J.; Gao, L.; Zuo, J.; Shi, J. Amelioration of postharvest chilling injury in cowpea (Vigna sinensis) by methyl jasmonate (MeJA) treatments. Sci. Hortic. 2016, 203, 95–101. [Google Scholar] [CrossRef]
- Gan, S.; Amasino, M.R. Cytokinins in plant senescence: From spray and pray to clone and play. Bioassays 1996, 18, 557–565. [Google Scholar] [CrossRef]
- Hwang, L.; Sheen, J.; Muller, B. Cytokinin signaling networks. Ann. Rev. Plant Biol. 2012, 63, 353–380. [Google Scholar] [CrossRef]
- Markovich, O.; Steiner, E.; Kouril, S.; Tarkowski, P.; Aharoni, A.; Elbaum, R. Silicon promotes cytokinin biosynthesis and delays senescence in Arabidopsis and Sorghum. Plant. Cell Environ. 2017, 40, 1189–1196. [Google Scholar] [CrossRef]
- Yu, T.; Wang, L.P.; Yin, Y.; Feng, F.Q.; Zheng, X.D. Suppression of postharvest blue mould of apple fruit by Cryptococcus laurentii and N-6-benzyladenine. J. Sci. Food Agric. 2008, 88, 1266–1271. [Google Scholar] [CrossRef]
- Zheng, X.D.; Yu, T.; Chen, R.L.; Huang, B.; Wu, V.C.H. Inhibiting Penicillium expansum infection on pear fruit by Cryptococcus laurentii and cytokinin. Postharvest Biol. Technol. 2007, 45, 221–227. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, X.; Zhang, Z.; Jiang, G.; Feng, L.; Duan, X.; Jiang, Y. 6-Benzylaminopurine improves the quality of harvested litchi fruit. Postharvest Biol. Technol. 2018, 143, 137–142. [Google Scholar] [CrossRef]
- Massolo, J.F.; Lemoine, M.L.; Chaves, A.R.; Concellon, A.; Vicente, A.R. Benzylaminopurine (BAP) treatments delay cell wall degradation and softening, improving quality maintenance of refrigerated summer squash. Postharvest Biol. Technol. 2014, 93, 122–129. [Google Scholar] [CrossRef]
- Chanjirakul, K.; Wang, S.Y.; Wang, C.Y.; Siriphanich, J. Effect of natural volatile compounds on antioxidant capacity and antioxidant enzymes in raspberries. Postharvest Biol. Technol. 2006, 40, 106–115. [Google Scholar] [CrossRef]
- Ghasemnezhad, M.; Javaherdashti, M. Effect of methyl jasmonate treatment on antioxidant capacity, internal quality and postharvest life of raspberry fruit. Casp. J. Environ. Sci. 2008, 1, 73–78. [Google Scholar]
- Giménez, M.J.; Valverde, J.M.; Valero, D.; Zapata, P.J.; Castillo, S.; Serrano, M. Postharvest methyl salicylate treatments delay ripening and maintain quality attributes and antioxidant compounds of ‘Early Lory’ sweet cherry. Postharvest Biol. Technol. 2016, 117, 102–109. [Google Scholar] [CrossRef]
- Muengkaew, R.; Chaiprasart, P.; Warrington, I. Changing of physiochemical properties and color development of mango fruit sprayed methyl Jasmonate. Sci. Hortic. 2016, 198, 70–77. [Google Scholar] [CrossRef]
- Awad, A.H.R.; Parmar, A.; Ali, M.R.; El-Mogy, M.M.; Abdelgawad, K.F. Extending the Shelf-Life of Fresh-Cut Green Bean Pods by Ethanol, Ascorbic Acid, and Essential Oils. Foods 2021, 10, 1103. [Google Scholar] [CrossRef]
- Javed, S.; Shoaib, A.; Mahmood, Z.; Mushtaq, S.; Iftikhar, S. Analysis of phytochemical constituents of Eucalyptus citriodora L. responsible for antifungal activity against post-harvest fungi. Nat. Prod. Res. 2012, 26, 1732–1736. [Google Scholar] [CrossRef]
- Petretto, G.L.; Chessa, M.; Piana, A.; Masia, M.D.; Foddai, M.; Mangano, G.; Culeddu, N.; Afifi Fatma, U.; Pintore, G. Chemical and biological study on the essential oil of Artemisia caerulescens L. ssp. Densiflora (Viv.). Nat. Prod. Res. 2013, 27, 1709–1715. [Google Scholar] [CrossRef]
- Bouabidi, W.; Hanana, M.; Gargouri, S.; Amri, I.; Fezzani, T.; Ksontini, M.; Jamoussi, B.; Hamrouni, L. Chemical composition, phytotoxic and antifungal properties of Ruta chalepensis L. essential oils. Nat. Prod. Res. 2015, 29, 864–868. [Google Scholar] [CrossRef]
- Moghaddam, M.; Taheri, P.; Pirbalouti, A.G.; Mehdizadeh, L. Chemical composition and antifungal activity of essential oil from the seed of Echinophora platyloba DC. against phytopathogens fungi by two different screening methods. LWT-Food Sci. Technol. 2015, 61, 536–542. [Google Scholar] [CrossRef]
- Dupuy, N.; Gaydou, V.; Kister, J. Quantitative analysis of lavender (Lavandula angustifolia) essential oil using multiblock data from infrared spectroscopy. Am. J. Analyt. Chem. 2014, 5, 633–645. [Google Scholar] [CrossRef]


{kind=link}
| Treatments | Storage Periods (Day) | |||||
|---|---|---|---|---|---|---|
| 0 | 5 | 10 | 15 | 25 | Average | |
| APX | ||||||
| Control | 0.204 ± 0.009 | 0.183 ± 0.004 | 0.289 ± 0.027 | 0.250 ± 0.124 | 0.262 ± 0.216 | 0.237 ± 0.040 B |
| 0.2 mM MeJA | 0.204 ± 0.009 | 0.275 ± 0.199 | 0.308 ± 0.042 | 0.318 ± 0.012 | 0.533 ± 0.283 | 0.327 ± 0.064 AB |
| 5 ppm cytokinin | 0.204 ± 0.009 | 0.277 ± 0.043 | 0.332 ± 0.020 | 0.655 ± 0.127 | 0.624 ± 0.008 | 0.418 ± 0.065 A |
| 1000 ppm lavender | 0.204 ± 0.009 | 0.275 ± 0.074 | 0.360 ± 0.276 | 0.510 ± 0.098 | 0.503 ± 0.029 | 0.370 ± 0.061 AB |
| MeJA + lavender | 0.204 ± 0.009 | 0.280 ± 0.001 | 0.321 ± 0.013 | 0.370 ± 0.058 | 0.410 ± 0.004 | 0.317 ± 0.026 AB |
| Cytokinin + lavender | 0.204 ± 0.009 | 0.347 ± 0.213 | 0.293 ± 0.054 | 0.398 ± 0.018 | 0.464 ± 0.258 | 0.341 ± 0.059 AB |
| Average | 0.204 ± 0.003 c | 0.273 ± 0.040 c | 0.317 ± 0.036 bc | 0.417 ± 0.048 ab | 0.466 ± 0.064 a | |
| p values; p treatments = 0.285; p storage periods = 0.004; p treatments × p storage periods = 0.984 | ||||||
| CAT | ||||||
| Control | 0.065 ± 0.002 | 0.087 ± 0.004 | 0.093 ± 0.002 | 0.089 ± 0.012 | 0.064 ± 0.020 | 0.079 ± 0.005 B |
| 0.2 mM MeJA | 0.065 ± 0.002 | 0.095 ± 0.004 | 0.144 ± 0.002 | 0.122 ± 0.004 | 0.115 ± 0.004 | 0.108 ± 0.009 A |
| 5 ppm cytokinin | 0.065 ± 0.002 | 0.111 ± 0.008 | 0.122 ± 0.004 | 0.116 ± 0.006 | 0.101 ± 0.005 | 0.103 ± 0.007 AB |
| 1000 ppm lavender | 0.065 ± 0.002 | 0.123 ± 0.001 | 0.132 ± 0.012 | 0.121 ± 0.043 | 0.115 ± 0.003 | 0.111 ± 0.010 A |
| MeJA + lavender | 0.065 ± 0.002 | 0.095 ± 0.002 | 0.142 ± 0.002 | 0.136 ± 0.005 | 0.111 ± 0.016 | 0.109 ± 0.010 A |
| Cytokinin + lavender | 0.065 ± 0.002 | 0.121 ± 0.003 | 0.125 ± 0.002 | 0.118 ± 0.000 | 0.097 ± 0.007 | 0.105 ± 0.008 A |
| Averages | 0.065 ± 0.000 d | 0.105 ± 0.004 bc | 0.126 ± 0.005 a | 0.117 ± 0.007 ab | 0.100 ± 0.006 b | |
| p values; p treatments = 0.001; p storage periods = 0.001; p treatments × p storage periods = 0.278 | ||||||
| SOD | ||||||
| Control | 163.96 ± 9.42 | 243.87 ± 8.81 | 348.60 ± 3.36 | 279.10 ± 6.29 | 252.50 ± 4.12 | 257.61 ± 19.98 B |
| 0.2 mM MeJA | 163.96 ± 9.42 | 276.70 ± 1.93 | 436.20 ± 5.52 | 388.76 ± 3.29 | 360.31 ± 5.18 | 325.19 ± 32.01 AB |
| 5 ppm cytokinin | 163.96 ± 9.42 | 288.41 ± 2.11 | 453.24 ± 18.40 | 389.78 ± 4.01 | 379.64 ± 2.65 | 335.01 ± 33.62 AB |
| 1000 ppm lavender | 163.96 ± 9.42 | 334.62 ± 3.70 | 397.45 ± 3.14 | 278.28 ± 6.03 | 258.24 ± 3.01 | 286.51 ± 26.10 AB |
| MeJA + lavender | 163.96 ± 9.42 | 364.30 ± 2.77 | 442.46 ± 1.90 | 412.68 ± 10.24 | 397.38 ± 6.64 | 356.16 ± 33.20 A |
| Cytokinin + lavender | 163.96 ± 9.42 | 362.04 ± 4.07 | 374.82 ± 4.06 | 362.33 ± 6.84 | 271.76 ± 16.75 | 306.98 ± 27.01 AB |
| Average | 163.964 ± 2.84 c | 311.656 ± 13.66 b | 408.79 ± 11.79 a | 351.823 ± 16.32 b | 319.97 ± 18.36 b | |
| p values; p treatments = 0.001; p storage periods = 0.001; p treatments × p storage periods = 0.001 | ||||||
| Treatments | Storage Periods (Day) | |||||
|---|---|---|---|---|---|---|
| 0 | 5 | 10 | 15 | 25 | Average | |
| MDA | ||||||
| Control | 3.020 ± 0.203 | 5.153 ± 0.877 | 5.996 ± 0.897 | 6.278 ± 0.204 | 6.304 ± 0.408 | 5.350 ± 0.459 A |
| 0.2 mM MeJA | 3.020 ± 0.203 | 4.039 ± 0.188 | 4.114 ± 0.733 | 4.687 ± 0.081 | 5.108 ± 0.610 | 4.193 ± 0.278 B |
| 5 ppm cytokinin | 3.020 ± 0.203 | 3.001 ± 1.538 | 4.267 ± 0.089 | 4.572 ± 0.325 | 4.584 ± 0.150 | 3.889 ± 0.339 B |
| 1000 ppm lavender | 3.020 ± 0.203 | 3.429 ± 0.173 | 3.774 ± 0.985 | 4.483 ± 0.418 | 4.950 ± 1.569 | 3.931 ± 0.369 B |
| MeJA + lavender | 3.020 ± 0.203 | 3.303 ± 0.127 | 3.618 ± 0.547 | 4.098 ± 0.034 | 4.462 ± 0.085 | 3.700 ± 0.196 B |
| Cytokinin + lavender | 3.020 ± 0.203 | 3.428 ± 0.090 | 4.320 ± 0.695 | 4.401 ± 0.338 | 4.682 ± 0.051 | 3.970 ± 0.243 B |
| Average | 3.020 ± 0.061 c | 3.725 ± 0.307 bc | 4.348 ± 0.320 ab | 4.753 ± 0.228 a | 5.015 ± 0.284 a | |
| p values; p treatments = 0.001; p storage periods 0.001; p treatments × p storage periods 0.969 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavusoglu, S.; Yilmaz, N.; Islek, F.; Tekin, O.; Sagbas, H.I.; Ercisli, S.; Rampáčková, E.; Nečas, T. Effect of Methyl Jasmonate, Cytokinin, and Lavender Oil on Antioxidant Enzyme System of Apricot Fruit (Prunus armeniaca L.). Sustainability 2021, 13, 8565. https://doi.org/10.3390/su13158565
Cavusoglu S, Yilmaz N, Islek F, Tekin O, Sagbas HI, Ercisli S, Rampáčková E, Nečas T. Effect of Methyl Jasmonate, Cytokinin, and Lavender Oil on Antioxidant Enzyme System of Apricot Fruit (Prunus armeniaca L.). Sustainability. 2021; 13(15):8565. https://doi.org/10.3390/su13158565
Chicago/Turabian StyleCavusoglu, Seyda, Nurettin Yilmaz, Firat Islek, Onur Tekin, Halil Ibrahim Sagbas, Sezai Ercisli, Eliška Rampáčková, and Tomáš Nečas. 2021. "Effect of Methyl Jasmonate, Cytokinin, and Lavender Oil on Antioxidant Enzyme System of Apricot Fruit (Prunus armeniaca L.)" Sustainability 13, no. 15: 8565. https://doi.org/10.3390/su13158565
APA StyleCavusoglu, S., Yilmaz, N., Islek, F., Tekin, O., Sagbas, H. I., Ercisli, S., Rampáčková, E., & Nečas, T. (2021). Effect of Methyl Jasmonate, Cytokinin, and Lavender Oil on Antioxidant Enzyme System of Apricot Fruit (Prunus armeniaca L.). Sustainability, 13(15), 8565. https://doi.org/10.3390/su13158565