Pool-Type Fishway Design for a Potamodromous Cyprinid in the Iberian Peninsula: The Iberian Barbel—Synthesis and Future Directions

 ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Swimming Performance of the Iberian Barbel and Hydrodynamics
3. Hydraulics of Pool-Type Fishways
3.1. Flow Regime and Discharge
3.2. Water Surface Profile
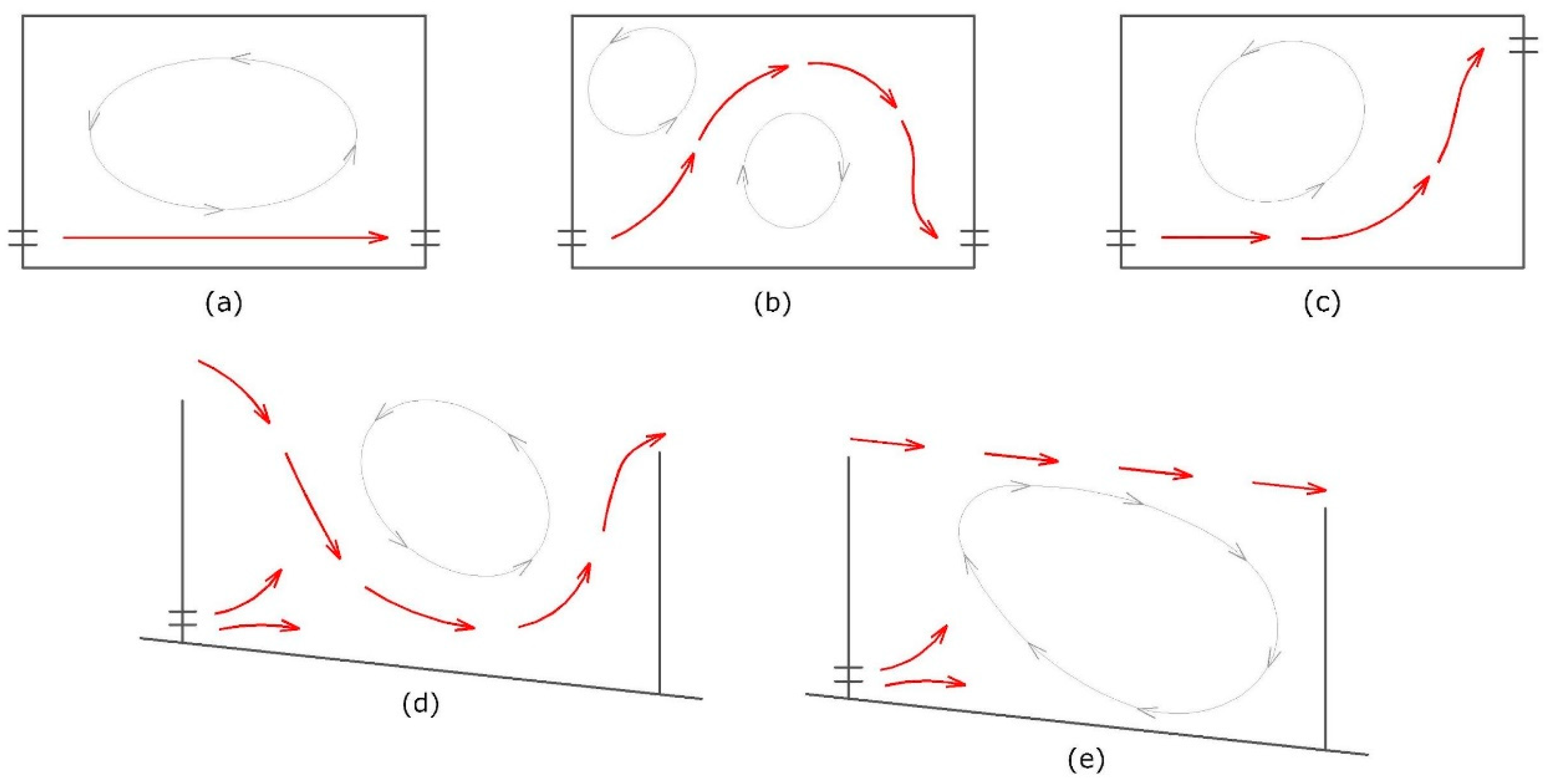
3.3. Flow Pattern, Velocity Distribution and Turbulence Levels in the Pools
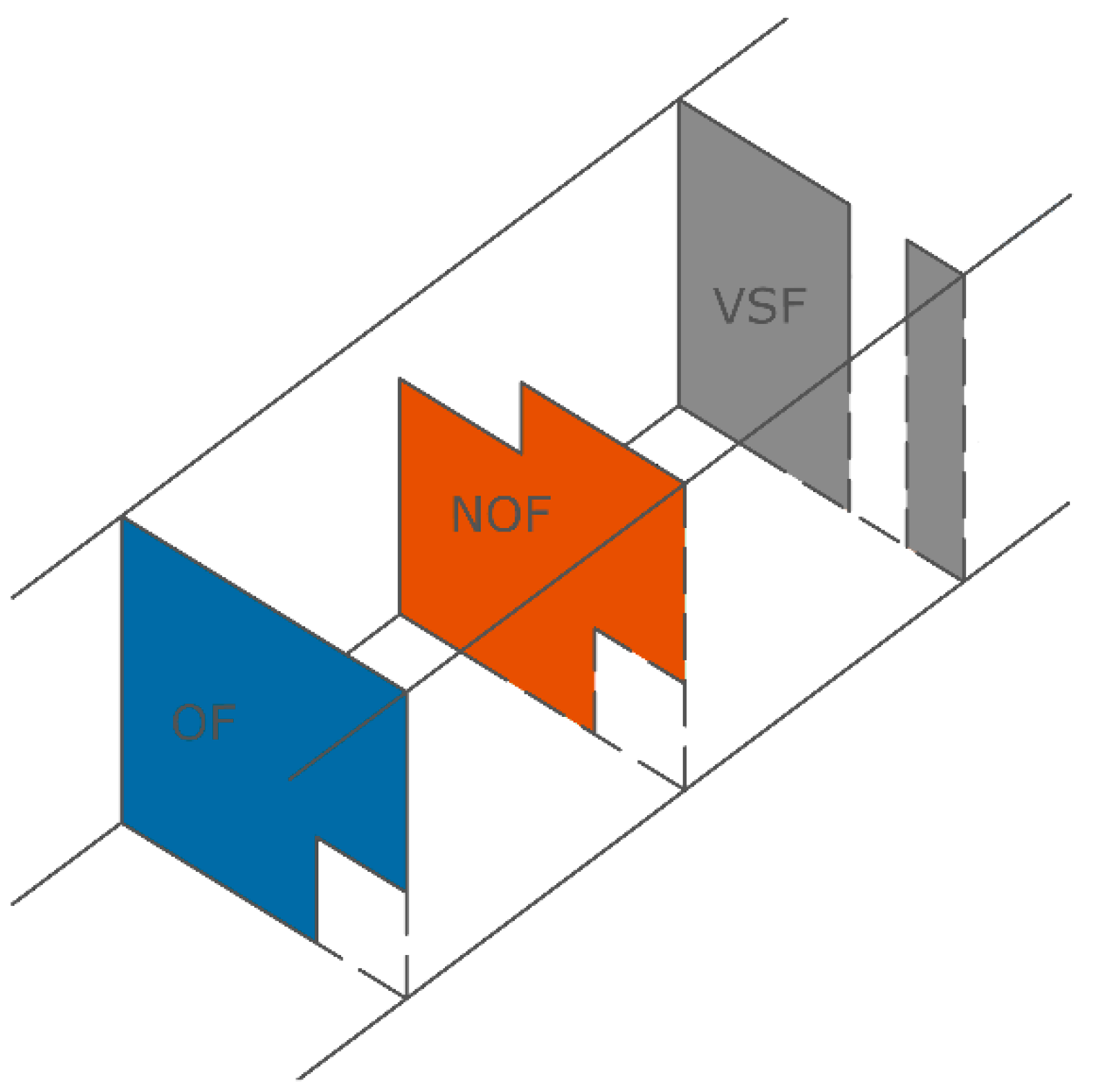
4. Comparative Passage Performance of Fishway Types for the Iberian Barbel
4.1. Overview of Conditions Tested
4.2. Overview of Main Findings
4.2.1. NOF
4.2.2. OF with Smooth Bottom
4.2.3. OF with Bottom Substrates
4.2.4. Vertical Slot Fishways (VSF)
5. Guidelines for Design of Fishways
- (1)
- Conduct fishway studies and evaluations preferably during spring and early-summer when fish are motivated to migrate upstream due to adequate environmental conditions, such as river discharge and water temperature. This should also include the dimensioning of the fishway itself, considering the range of river flows and water levels experienced during this season and the associated topographic works.
- (2)
- Combine surface notches with submerged orifices for barbel passage. Submerged orifices were clearly the preferential route choice of barbel to upstream movements in pool-and-weir fishways (>75% of total movements) so their use should always be encouraged, if possible with surface notches, to provide alternative routes of passage, when the former, which are more easily prone to clogging, become blocked by woody debris. This may be of particular importance in remote locations, where regular checking and maintenance of the fishway is seldom performed.
- (3)
- Provide streaming flow conditions within pool-type fishways whenever surface notches are present. Streaming flow proved to be a more efficient flow regime by facilitating upstream passage. Such conditions should be favored in detriment of plunging ones. Provided that sufficient water is available, this could be achieved by increasing the fishway discharge and allowing submersion of the crest of the notches as a consequence of increasing the water level of the downstream pools.
- (4)
- Encourage the implementation of offset orifice arrangements. Offset orifices were shown to provide a higher passage success and to be more efficient in passing barbel, particularly smaller individuals, with minimal delay. Contrarily, straight orifices arrangements should be avoided as they may cause higher shear stress in the vicinity of the orifices, larger vortices, and more pronounced changes in velocity, which seemed to have hampered upstream movements.
- (5)
- Consider the placement of submerged structures (e.g., artificial or natural boulders) in the pool’s bottom. This will allow attenuating the high turbulence levels (particularly, the horizontal RSS) detected in the vicinity of the submerged orifices, which seemed to hamper and delay the movement of small fish. This placement of structural elements should also be favored in the pools itself to reduce the size of potential recirculation regions that might trap fish.
- (6)
- Favor relative depths of flow (d/h)—as the ratio between the mean water depth in the pool (d) and the height of artificial bottom structures (h)—higher than 4. This ratio was found to be more beneficial to fish passage since it creates small reverse-flow fields that might aid upstream movements. In addition, it favors the presence of negative vertical (i.e., descendent) velocities that best orientate the fish within the pathways created by such structures.
- (7)
- Use submerged structures embedded on pools bottom at high density, combined with higher fishway discharge. This should be encouraged as it significantly reduced fish transit time; thus, it is particularly important to be considered in case of longer fishways, where the energy expenditure of migrating fish may be higher and hence fish may be more prone to fatigue. If, however, fishway discharge is a constraint, a lower density design should be employed instead, because lower density yielded higher passage success with low discharge. Whichever the case, regular inspections and maintenance of the pools is necessary due to potential clogging by floating and dragged debris.
- (8)
- Provide an adequate fishway discharge, which should be sufficiently attractive for the fish, but not excessive. Fishway discharge is a critical variable as barbel’s passage was found to be flow-related, being higher at higher discharges, but not excessively high (flow in the present study encompassed a high variability, i.e., from 38.5 L·s−1 to 402 L·s−1). The optimal value will depend on the size of the fishway considered (larger/wider fishways will require higher flows to adequately operate) and on the range of flow variations in the river. In some cases, it might be important to consider the provision of an extra flow to increase attraction or to provide an additional fish entrance.
- (9)
- Provide water velocities within the pools in the order of 0.40 m·s−1–0.60 m·s−1. Water velocity within the pools was found to be a key-feature for effective fish passage in pool-type fishways. Water velocities should be sufficiently attractive for fish but should not exceed fish swimming capacity. Excessive velocities and the consequent excessive turbulence will make it difficult for fish to keep oriented while trying to negotiate the fishway. For the barbel, mean water velocities within the pools ranging between 0.40 m·s−1 and 0.60 m·s−1 seemed to provide adequate conditions for fishway negotiation.
- (10)
- Provide head drops between pools of 16 cm–20 cm. This will provide water velocities in the slots (in case of VSF) of up to 1.7 m·s−1, therefore below the desirable threshold of 2 m·s−1 (limited established for cyprinids [105]). Although being dependent on head drop between the pools, such velocities are relatively unaffected by discharge.
- (11)
- Use vertical slots featuring slots with a single lateral baffle. This should be preferred in detriment to those presenting an additional central baffle. Although significant differences between both slot configurations were not found on the number of successful upstream movements, configurations with a single lateral baffle may be a better option, because it requires a lower discharge to operate, making it a more cost-effective design.
- (12)
- Consider the use of multi-slot fishways (MSF) in detriment of standard vertical-slot fishways (VSF). Despite their similar performance in terms of successful negotiations, MSF were shown to be more discharge-efficient than VSF, because they required 31% less water to operate for the same water depth in the pools. Therefore, velocity and turbulence are expected to present lower magnitudes, which will favor the negotiation by smaller individuals. Nonetheless, managers should bear in mind that due to their lower operational discharge and design characteristics, MSF could lack appropriate attraction flow while being more prone to clogging. To overcome the problem of adequate attraction flow, an additional discharge at the fishway entrance could be provided to increase attraction and make these fishways more competitive with the dominant flow.
- (13)
- Take into consideration the non-uniformity of flow caused by variations in river water levels upon designing and evaluating the performance of fishways. The non-uniform scenarios, namely the “backwater” profile and the “drawdown” profile, were shown to affect fish distribution due to a change in local hydrodynamics of the pool, and should be considered as they are representative for the changes in up- and downstream water levels that naturally occur with changes in river flow over the year. In such cases, fishways may need to be adapted, for example with adaptable sills in the most downstream cross-walls or submerged pre-barrages downstream of the fishway as suggested by Fuentes-Pérez et al. [74].
6. Missing Pieces and Future Directions
6.1. Science: Ecology and Technology
6.2. Society
6.3. Policy
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benitez, J.-P.; Dierckx, A.; Matondo, B.N.; Rollin, X.; Ovidio, M. Movement behaviours of potamodromous fish within a large anthropised river after the reestablishment of the longitudinal connectivity. Fish. Res. 2018, 207, 140–149. [Google Scholar] [CrossRef]
- Ordeix, M.; González, G.; Sanz-Ronda, F.J.; Santos, J.M. Restoring fish migration in the rivers of the Iberian Peninsula. In From Sea to Source 2.0. Protection and Restoration of Fish Migration in Rivers Worldwide; Brink, K., Gough, P., Royte, J., Schollema, P.P., Wanningen, H., Eds.; World Fish Migration Foundation: Groningen, The Netherlands, 2018; pp. 174–179. [Google Scholar]
- Santos, R.E.; Pinto-Coelho, R.M.; Fonseca, R.; Simões, N.R.; Zanchi, F.B. The decline of fisheries on the Madeira River, Brazil: The high cost of the hydroelectric dams in the Amazon Basin. Fish. Manag. Ecol. 2018, 25, 380–391. [Google Scholar] [CrossRef]
- Mazumder, D.; Williams, R.J.; Wen, L.; Saintilan, N.; Walsh, C.T. Impoundment constraint of fish niche diversity in a temperate Australian river. Hydrobiologia 2016, 771, 195–206. [Google Scholar] [CrossRef]
- Olden, J.D.; Kennard, M.J.; Leprieur, F.; Tedesco, P.A.; Winemiller, K.O.; García-Berthou, E. Conservation biogeography of freshwater fishes: Recent progress and future challenges. Divers. Distrib. 2010, 16, 496–513. [Google Scholar] [CrossRef]
- Agostinho, A.A.; Pelicice, F.M.; Gomes, L.C. Dams and the fish fauna of the Neotropical region: Impacts and management related to diversity and fisheries. Braz. J. Boil. 2008, 68, 1119–1132. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Boil. Rev. 2005, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- European Environment Agency. Reservoirs and Dams. 2018. Available online: https://www.eea.europa.eu/archived/archived-content-water-topic/reservoirs-and-dams (accessed on 14 February 2020).
- Centro Ibérico de Restauración Fluvial (CIREF). An analysis of river fragmentation in the Spanish river basins. Technical report developed by Ecohidráulica, S.L., Madrid, Spain, 2016. Available online: www.cirefluvial.com (accessed on 19 April 2020).
- Silva, A.T.; Lucas, M.C.; Castro-Santos, T.; Katopodis, C.; Baumgartner, L.J.; Thiem, J.D.; Aarestrup, K.; Pompeu, P.S.; O’Brien, G.C.; Braun, D.C.; et al. The future of fish passage science, engineering, and practice. Fish Fish. 2017, 19, 340–362. [Google Scholar] [CrossRef]
- Larinier, M. Pool fishways, pre-barrages and natural bypass channels. Bull. Français de la Pêche et de la Pisciculture 2002, 54–82. [Google Scholar] [CrossRef]
- Clay, C.H.; Eng, P. Design of Fishways and Other Fish Facilities, 2nd ed.; Lewis Publishers, CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Larinier, M.; Marmulla, G. Fish passes: Types, principles and geographical distribution an overview. In Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries, Phnom Penh, Cambodia, 11–14 February 2003; Volume II, pp. 183–206. [Google Scholar]
- Gough, P.; Philipsen, P.; Schollema, P.P.; Wanningn, H. Chapter 5: Solutions for hazards and obstacles. In From Sea to Source: International Guidance for the Restoration of Fish Migration Highways; Regional Water Authority Hunze en Aa’s: AD Veendam, The Netherlands, 2012. [Google Scholar]
- Santos, J.M.; Silva, A.T.; Katopodis, C.; Pinheiro, P.; Pinheiro, A.N.; Bochechas, J.; Ferreira, M.T. Ecohydraulics of pool-type fishways: Getting past the barriers. Ecol. Eng. 2012, 48, 38–50. [Google Scholar] [CrossRef]
- Pena, L.; Puertas, J.; Bermúdez, M.; Cea, L.; Peña, E. Conversion of Vertical Slot Fishways to Deep Slot Fishways to Maintain Operation during Low Flows: Implications for Hydrodynamics. Sustainability 2018, 10, 2406. [Google Scholar] [CrossRef]
- Quaresma, A.; Romão, F.; Branco, P.; Ferreira, M.T.; Pinheiro, A.N. Multi slot versus single slot pool-type fishways: A modelling approach to compare hydrodynamics. Ecol. Eng. 2018, 122, 197–206. [Google Scholar] [CrossRef]
- Bombač, M.; ČETINA, M.; Novak, G. Study on flow characteristics in vertical slot fishways regarding slot layout optimization. Ecol. Eng. 2017, 107, 126–136. [Google Scholar] [CrossRef]
- Bermúdez, M.; Puertas, J.; Cea, L.; Pena, L.; Balairón, L. Influence of pool geometry on the biological efficiency of vertical slot fishways. Ecol. Eng. 2010, 36, 1355–1364. [Google Scholar] [CrossRef]
- Rajaratnam, N.; Katopodis, C.; Solanki, S. New designs for vertical slot fishways. Can. J. Civ. Eng. 1992, 19, 402–414. [Google Scholar] [CrossRef]
- Baumgartner, L.J.; Marsden, T.; Singhanouvong, D.; Phonekhampheng, O.; Stuart, I.G.; Thorncraft, G. Using an experimentalin situfishway to provide key design criteria for lateral fish passage in tropical rivers: A case study from the mekong river, central Lao Pdr. River Res. Appl. 2011, 28, 1217–1229. [Google Scholar] [CrossRef]
- Barrett, J.; Mallen-Cooper, M. The Murray River’s ’Sea to Hume Dam’ fish passage program: Progress to date and lessons learned. Ecol. Manag. Restor. 2006, 7, 173–183. [Google Scholar] [CrossRef]
- Williams, J.G.; Armstrong, G.; Katopodis, C.; Larinier, M.; Travade, F. Thinking Like a Fish: A Key Ingredient for Development of Effective Fish Passage Facilities at River Obstructions. River Res. Appl. 2011, 28, 407–417. [Google Scholar] [CrossRef]
- Mallen-Cooper, M.; Brand, D.A. Non-salmonids in a salmonid fishway: What do 50 years of data tell us about past and future fish passage? Fish. Manag. Ecol. 2007, 14, 319–332. [Google Scholar] [CrossRef]
- Noonan, M.J.; Grant, J.W.A.; Jackson, C.D. A quantitative assessment of fish passage efficiency. Fish Fish. 2011, 13, 450–464. [Google Scholar] [CrossRef]
- Foulds, W.L.; Lucas, M.C. Extreme inefficiency of two conventional, technical fishways used by European river lamprey (Lampetra fluviatilis). Ecol. Eng. 2013, 58, 423–433. [Google Scholar] [CrossRef]
- Marriner, B.A.; Baki, A.B.; Zhu, D.Z.; Cooke, S.J.; Katopodis, C. The hydraulics of a vertical slot fishway: A case study on the multi-species Vianney-Legendre fishway in Quebec, Canada. Ecol. Eng. 2016, 90, 190–202. [Google Scholar] [CrossRef]
- Thiem, J.D.; Binder, T.R.; Dumont, P.; Hatin, D.; Hatry, C.; Katopodis, C.; Stamplecoskie, K.M.; Cooke, S.J. Multispecies fish passage behaviour in a vertical slot fishway on the Richelieu River, Quebec, Canada. River Res. Appl. 2012, 29, 582–592. [Google Scholar] [CrossRef]
- Lucas, M.C.; Mercer, T.; McGinty, S.; Armstrong, J.D. Development and evaluation of a flat-bed pass. In Advances in Fish Telemetry; Moore, A., Russell, I., Eds.; CEFAS: Suffolk, UK, 2000; pp. 117–127. [Google Scholar]
- Ferreira, M.T.; Sousa, L.V.D.C.; Santos, J.M.; Reino, L.; Oliveira, J.; Almeida, P.; Cortes, R.V. Regional and local environmental correlates of native Iberian fish fauna. Ecol. Freshw. Fish 2007, 16, 504–514. [Google Scholar] [CrossRef]
- Sociedad Ibérica de Ictiología (SIBIC) Iberian Barbel-Species Info. Available online: http://www.cartapiscicola.es/#/species/bboc (accessed on 11 September 2018).
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Publications Kottelat: Cornol, Switzerland, 2007; ISBN 9782839902984. [Google Scholar]
- Doadrio, I.; Perea, S.; Garzón-Heydt, P.; González, J.L. Ictiofauna Continental Española: Bases Para su Seguimiento; Ministerio de Medio Ambiente y Medio Rural y Marino, Centro de Publicaciones: Madrid, Spain, 2011; ISBN 9788449111587. [Google Scholar]
- Silva, A.T.; Santos, J.M.; Ferreira, M.T.; Pinheiro, A.N.; Katopodis, C. Effects of water velocity and turbulence on the behaviour of Iberian barbel (Luciobarbus bocagei, Steindachner 1864) in an experimental pool-type fishway. River Res. Appl. 2011, 27, 360–373. [Google Scholar] [CrossRef]
- Mateus, C.; Quintella, B.; Almeida, P. The critical swimming speed of Iberian barbelBarbus bocageiin relation to size and sex. J. Fish Boil. 2008, 73, 1783–1789. [Google Scholar] [CrossRef]
- Roskov, Y.; Ower, G.; Orrell, T.; Nicolson, D.; Bailly, N.; Kirk, P.M.; Bourgoin, T.; DeWalt, R.E.; Decock, W.; Nieukerken, E.; et al. FishBase (version Feb 2018). In Species 2000 & ITIS Catalogue of Life, 2020-02-24; Froese, R., Pauly, D., Eds.; Species 2000; Naturalis: Leiden, The Netherlands; ISSN 2405-8858. Available online: www.catalogueoflife.org/col (accessed on 4 April 2020).
- Alexandre, C.; Romão, F.; Almeida, P.; Quintella, B.; Ferreira, A.F. Swimming performance and ecomorphology of the Iberian barbel Luciobarbus bocagei (Steindachner, 1864) on permanent and temporary rivers. Ecol. Freshw. Fish 2013, 23, 244–258. [Google Scholar] [CrossRef]
- Silva, A.T.; Katopodis, C.; Santos, J.M.; Ferreira, M.T.; Pinheiro, A.N. Cyprinid swimming behaviour in response to turbulent flow. Ecol. Eng. 2012, 44, 314–328. [Google Scholar] [CrossRef]
- Silva, A.T.; Santos, J.M.; Ferreira, M.T.; Pinheiro, A.N.; Katopodis, C. Passage efficiency of offset and straight orifices for upstream movements of Iberian barbel in a pool-type fishway. River Res. Appl. 2010, 28, 529–542. [Google Scholar] [CrossRef]
- Santos, J.M.; Branco, P.; Katopodis, C.; Ferreira, M.T.; Pinheiro, A.N. Retrofitting pool-and-weir fishways to improve passage performance of benthic fishes: Effect of boulder density and fishway discharge. Ecol. Eng. 2014, 73, 335–344. [Google Scholar] [CrossRef]
- Alexandre, C.; Quintella, B.; Silva, A.T.; Mateus, C.; Romão, F.; Branco, P.; Ferreira, M.T.; Almeida, P. Use of electromyogram telemetry to assess the behavior of the Iberian barbel (Luciobarbus bocagei Steindachner, 1864) in a pool-type fishway. Ecol. Eng. 2013, 51, 191–202. [Google Scholar] [CrossRef]
- Bravo-Córdoba, F.J.; Sanz-Ronda, F.J.; Ruiz-Legazpi, J.; Valbuena-Castro, J.; Makrakis, S. Vertical slot versus submerged notch with bottom orifice: Looking for the best technical fishway type for Mediterranean barbels. Ecol. Eng. 2018, 122, 120–125. [Google Scholar] [CrossRef]
- Plaut, I. Critical swimming speed: Its ecological relevance. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 131, 41–50. [Google Scholar] [CrossRef]
- Lighthill, M.J. Hydromechanics of Aquatic Animal Propulsion. Annu. Rev. Fluid Mech. 1969, 1, 413–446. [Google Scholar] [CrossRef]
- Franssen, N.R. Anthropogenic habitat alteration induces rapid morphological divergence in a native stream fish. Evol. Appl. 2011, 4, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.S. Fishes of the World, 4th ed.; Wiley: New York, NY, USA, 2006. [Google Scholar]
- Wheeler, A.; Lelek, A. The Freshwater Fishes of Europe. Vol. 9. Threatened Fishes of Europe. Copeia 1988, 1988, 1099. [Google Scholar] [CrossRef]
- Doadrio, I.; Casado, P. Nota sobre la ictiofauna continental de los yacimientos de la cuenca de Guadix-Baza (Granada). In Geologıa y Paleontologıa de la Cuenca de Guadix-Baza; Alberdi, M.T., Bonadonna, F.P., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 1989; pp. 139–150. [Google Scholar]
- Sanz-Ronda, F.J.; Bravo-Córdoba, F.; Fuentes-Perez, J.F.; Castro-Santos, T. Ascent ability of brown trout, Salmo trutta, and two Iberian cyprinids−Iberian barbel, Luciobarbus bocagei, and northern straight-mouth nase, Pseudochondrostoma duriense− in a vertical slot fishway. Knowl. Manag. Aquat. Ecosyst. 2016, 417, 10. [Google Scholar] [CrossRef]
- Castro-Santos, T. Optimal swim speeds for traversing velocity barriers: An analysis of volitional high-speed swimming behavior of migratory fishes. J. Exp. Boil. 2005, 208, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, D.S.; Skorobagatov, M.A.; Shtaf, L.G. The critical current velocity of fish and the degree of flow turbulence. Rep. USSR Acad. Sci. 1982, 267, 1019–1021. [Google Scholar]
- Ruiz-Legazpi, J.; Sanz-Ronda, F.J.; Fuentes-Pérez, J.F. Influence of environmental and biometric factors on the swimming capacity of the Iberian barbel (Luciobarbus bocagei Steindachner, 1864), an endemic potamodromous cyprinid of the Iberian Peninsula. Limnetica 2018, 37, 251–265. [Google Scholar] [CrossRef]
- Sanz-Ronda, F.J.; Ruiz-Legazpi, J.; Córdoba, F.J.B.; Makrakis, S.; Castro-Santos, T.; Sanz-Ronda, F.J. Sprinting performance of two Iberian fish: Luciobarbus bocagei and Pseudochondrostoma duriense in an open channel flume. Ecol. Eng. 2015, 83, 61–70. [Google Scholar] [CrossRef]
- Lupandin, A.I. Effect of Flow Turbulence on Swimming Speed of Fish. Boil. Bull. 2005, 32, 461–466. [Google Scholar] [CrossRef]
- Cada, G.; Carlson, T.; Ferguson, J.; Richmond, M.; Sale, M. Exploring the Role of Shear Stress and Severe Turbulence in Downstream Fish Passage. In Proceedings of the Waterpower Conference 1999; Brookshier, P.A., Ed.; American Society of Civil Engineers (ASCE): Las Vegas, NV, USA, 1999; p. 10. [Google Scholar]
- Tritico, H.M.; Cotel, A.J. The effects of turbulent eddies on the stability and critical swimming speed of creek chub (Semotilus atromaculatus). J. Exp. Boil. 2010, 213, 2284–2293. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.C. A review of fish swimming mechanics and behaviour in altered flows. Philos. Trans. R. Soc. B Boil. Sci. 2007, 362, 1973–1993. [Google Scholar] [CrossRef] [PubMed]
- FAO. DVWK Fish Passes: Design, Dimensions and Monitoring; Food and Agriculture Organization of the United Nations: Rome, Italy, 2002; ISBN 3895540277. [Google Scholar]
- Katopodis, C.; Williams, J.G. Not all fishways are created equal. In Advances in Fish Passage Research. In Proceedings of the 11th International Symposium on Ecohydraulics, Melbourne, Australia, 7–12 February 2016. [Google Scholar]
- Katopodis, C. Introduction to Fishway Design; Freshwater Institute, Department of Fisheries and Oceans: Manitoba, ON, Canada, 1992. [Google Scholar]
- Bombač, M.; Novak, G.; Mlacnik, J.; ČETINA, M. Extensive field measurements of flow in vertical slot fishway as data for validation of numerical simulations. Ecol. Eng. 2015, 84, 476–484. [Google Scholar] [CrossRef]
- Katopodis, C.; Williams, J.G. The development of fish passage research in a historical context. Ecol. Eng. 2012, 48, 8–18. [Google Scholar] [CrossRef]
- Rajaratnam, N.; Katopodis, C.; Mainali, A. Pool-orifice and pool-orifice-weir fishways. Can. J. Civ. Eng. 1989, 16, 774–777. [Google Scholar] [CrossRef]
- Boiten, W.; Dommerholt, A. Standard design of the Dutch pool and orifice fishway. Int. J. River Basin Manag. 2006, 4, 219–227. [Google Scholar] [CrossRef]
- Yagci, O. Hydraulic aspects of pool-weir fishways as ecologically friendly water structure. Ecol. Eng. 2010, 36, 36–46. [Google Scholar] [CrossRef]
- Wu, S.; Rajaratnam, N.; Katopodis, C. Structure of Flow in Vertical Slot Fishway. J. Hydraul. Eng. 1999, 125, 351–360. [Google Scholar] [CrossRef]
- Tauber, M.; Mader, D. Hydraulic comparison of standard vertical slot and multi structure slot fish bypass. In Proceedings of the first European IAHR Congress, Edinburgh, UK, 4–6 May 2010. [Google Scholar]
- Laine, A.; Jokivirta, T.; Katopodis, C. Atlantic salmon, Salmo salar L., and sea trout, Salmo trutta L., passage in a regulated northern river—Fishway efficiency, fish entrance and environmental factors. Fish. Manag. Ecol. 2002, 9, 65–77. [Google Scholar] [CrossRef]
- Romão, F.; Quaresma, A.; Branco, P.; Santos, J.M.; Amaral, S.; Ferreira, M.T.; Katopodis, C.; Pinheiro, A.N. Passage performance of two cyprinids with different ecological traits in a fishway with distinct vertical slot configurations. Ecol. Eng. 2017, 105, 180–188. [Google Scholar] [CrossRef]
- USFWS (U.S. Fish and Wildlife Service). Fish Passage Engineering Design Criteria; USFWS, Northeast Region R5: Hadley, MA, USA, 2019. [Google Scholar]
- O’Connor, J.; Mallen-Cooper, M.; Stuart, I. Performance, operation and maintenance guidelines for fishways and fish passage works. In Arthur Rylah Institute for Environmental Research Technical Report No. 262 for the Water and Catchments Group, Department of Environment, Land, Water and Planning; Arthur Rylah Institute for Environmental Research, Department of Environment, Land, Water and Planning: Heidelberg, Germany, 2015. [Google Scholar]
- Rajaratnam, N.; Katopodis, C.; Mainali, A. Plunging and Streaming Flows in Pool and Weir Fishways. J. Hydraul. Eng. 1988, 114, 939–944. [Google Scholar] [CrossRef]
- Ead, S.; Katopodis, C.; Sikora, G.J.; Rajaratnam, N. Flow regimes and structure in pool and weir fishways. J. Environ. Eng. Sci. 2004, 3, 379–390. [Google Scholar] [CrossRef]
- Fuentes-Perez, J.F.; Sanz-Ronda, F.J.; De Azagra, A.M.; García-Vega, A. Non-uniform hydraulic behavior of pool-weir fishways: A tool to optimize its design and performance. Ecol. Eng. 2016, 86, 5–12. [Google Scholar] [CrossRef]
- Fuentes-Perez, J.F.; García-Vega, A.; Sanz-Ronda, F.J.; Paredes, A.M.D.A. Villemonte’s approach: A general method for modeling uniform and non-uniform performance in stepped fishways. Knowl. Manag. Aquat. Ecosyst. 2017, 418, 23. [Google Scholar] [CrossRef]
- Fuentes-Perez, J.F.; Branco, P.; Tuhtan, J.A.; Ferreira, M.T.; Kruusmaa, M.; Branco, P. Spatial preferences of Iberian barbel in a vertical slot fishway under variable hydrodynamic scenarios. Ecol. Eng. 2018, 125, 131–142. [Google Scholar] [CrossRef]
- Fuentes-Perez, J.F.; Sanz-Ronda, F.J.; Paredes, A.M.D.A.; García-Vega, A. Modeling Water-Depth Distribution in Vertical-Slot Fishways under Uniform and Nonuniform Scenarios. J. Hydraul. Eng. 2014, 140, 06014016. [Google Scholar] [CrossRef]
- Puertas, J.; Cea, L.; Bermúdez, M.; Pena, L.; Rodriguez, A.; Rabuñal, J.R.; Balairón, L.; Lara, Á.; Aramburu, E. Computer application for the analysis and design of vertical slot fishways in accordance with the requirements of the target species. Ecol. Eng. 2012, 48, 51–60. [Google Scholar] [CrossRef]
- Rodriguez, A.; Bermúdez, M.; Rabuñal, J.R.; Puertas, J.; Dorado, J.; Peña, L.; Balairón, L. Optical Fish Trajectory Measurement in Fishways through Computer Vision and Artificial Neural Networks. J. Comput. Civ. Eng. 2011, 25, 291–301. [Google Scholar] [CrossRef]
- Rodriguez, A.; Bermúdez, M.; Rabuñal, J.R.; Puertas, J. Fish tracking in vertical slot fishways using computer vision techniques. J. Hydroinform. 2014, 17, 275–292. [Google Scholar] [CrossRef]
- Calluaud, D.; Pineau, G.; Texier, A.; David, L. Modification of vertical slot fishway flow with a supplementary cylinder. J. Hydraul. Res. 2014, 52, 614–629. [Google Scholar] [CrossRef]
- Wang, R.; David, L.; Larinier, M. Contribution of experimental fluid mechanics to the design of vertical slot fish passes. Knowl. Manag. Aquat. Ecosyst. 2010, 2. [Google Scholar] [CrossRef]
- Puertas, J.; Pena, L.; Teijeiro, T. Experimental Approach to the Hydraulics of Vertical Slot Fishways. J. Hydraul. Eng. 2004, 130, 10–23. [Google Scholar] [CrossRef]
- Kim, J. Hydraulic characteristics by weir type in a pool-weir fishway. Ecol. Eng. 2001, 16, 425–433. [Google Scholar] [CrossRef]
- Branco, P.; Santos, J.M.; Katopodis, C.; Pinheiro, A.N.; Ferreira, M.T. Pool-Type Fishways: Two Different Morpho-Ecological Cyprinid Species Facing Plunging and Streaming Flows. PLoS ONE 2013, 8, e65089. [Google Scholar] [CrossRef]
- Santos, J.M.; Branco, P.; Silva, A.T.; Katopodis, C.; Pinheiro, A.N.; Viseu, T.; Ferreira, M.T. Effect of two flow regimes on the upstream movements of the Iberian barbel (Luciobarbus bocagei) in an experimental pool-type fishway. J. Appl. Ichthyol. 2012, 29, 425–430. [Google Scholar] [CrossRef]
- Katopodis, C. Developing a toolkit for fish passage, ecological flow management and fish habitat works. J. Hydraul. Res. 2005, 43, 451–467. [Google Scholar] [CrossRef]
- Silva, A.T.; Santos, J.M.; Franco, A.C.; Ferreira, M.T.; Pinheiro, A.N. Selection of Iberian barbel Barbus bocagei (Steindachner, 1864) for orifices and notches upon different hydraulic configurations in an experimental pool-type fishway. J. Appl. Ichthyol. 2009, 25, 173–177. [Google Scholar] [CrossRef]
- Quintella, B.R.; Mateus, C.S.; Silva, A.T.; Ferreira, M.T.; Almeida, P.R. Assessing the Behavioural Response of the Iberian Barbel (Barbus bocagei) in a Pool-And-Weir Type Fishway Prototype Using EMG Telemetry. In Proceedings of the 33rd IAHR Congress, Vancouver, BC, Canada, 9–14 August 2009. [Google Scholar]
- Aramburu, E.; Morcillo, F.; Castillo, M.; Lara, A. Research on fish behavior in a fishway in the Hydraulic Laboratory of the Centre for Hydrographic Studies. Conclusions of the experiment period from 2009 to 2012. Ing. Civ. 2014, 173, 33–42. [Google Scholar]
- Branco, P.; Santos, J.M.; Katopodis, C.; Viseu, T.; Ferreira, M.T.; Pinheiro, A.N. The influence of bottom rugosity on the performance of upstream fish movements through a pool-type fishway under different discharges. proceedings of the 36th IAHR Congress, The Hague, Netherlands, 28 June–3 July 2015. [Google Scholar]
- Bravo-Córdoba, F.J.; Sanz-Ronda, F.J.; Ruiz-Legazpi, J.; Celestino, L.; Makrakis, S. Fishway with two entrance branches: Understanding its performance for potamodromous Mediterranean barbels. Fish. Manag. Ecol. 2017, 25, 12–21. [Google Scholar] [CrossRef]
- Romão, F.; Branco, P.; Quaresma, A.; Amaral, S.; Pinheiro, A.N. Effectiveness of a multi-slot vertical slot fishway versus a standard vertical slot fishway for potamodromous cyprinids. Hydrobiologia 2018, 816, 153–163. [Google Scholar] [CrossRef]
- Romão, F.; Santos, J.M.; Katopodis, C.; Pinheiro, A.N.; Branco, P. How Does Season Affect Passage Performance and Fatigue of Potamodromous Cyprinids? An Experimental Approach in a Vertical Slot Fishway. Water 2018, 10, 395. [Google Scholar] [CrossRef]
- Romão, F.; Quaresma, A.; Santos, J.M.; Branco, P.; Pinheiro, A.N. Cyprinid passage performance in an experimental multislot fishway across distinct seasons. Mar. Freshw. Res. 2019, 70, 881. [Google Scholar] [CrossRef]
- Pedescoll, A.; Aguado, R.; Marcos, C.; González, G. Performance of a Pool and Weir Fishway for Iberian Cyprinids Migration: A Case Study. Fishes 2019, 4, 45. [Google Scholar] [CrossRef]
- Mader, H.; Brandl, A.; Käfer, S. Design and Function Monitoring of an Enature® Vertical Slot Fish Pass in a Large Potamal River in Carinthia/Austria. Water 2020, 12, 551. [Google Scholar] [CrossRef]
- Guiny, E.; Armstrong, J.; Ervine, D.A. Preferences of mature male brown trout and Atlantic salmon parr for orifice and weir fish pass entrances matched for peak velocities and turbulence. Ecol. Freshw. Fish 2003, 12, 190–195. [Google Scholar] [CrossRef]
- Amaral, S.; Branco, P.; Katopodis, C.; Ferreira, M.T.; Pinheiro, A.N.; Santos, J.M. To swim or to jump? Passage behaviour of a potamodromous cyprinid over an experimental broad-crested weir. River Res. Appl. 2017, 34, 174–182. [Google Scholar] [CrossRef]
- Santos, J.M.; Ferreira, M.T.; Godinho, F.; Bochechas, J.; Ferreira, M.T. Efficacy of a nature-like bypass channel in a Portuguese lowland river. J. Appl. Ichthyol. 2005, 21, 381–388. [Google Scholar] [CrossRef]
- Acharya, M.; Kells, J.A.; Katopodis, C. Hydraulics of micro-channels in nature-like fishways. In Proceedings of the 15th Hydrotechnical Conference of the Canadian Society for Civil Engineering, Victoria, BC, Canada, 30 May–2 June 2001. [Google Scholar]
- Hinch, S.G.; Rand, P.S. Swim speeds and energy use of upriver-migrating sockeye salmon (Oncorhynchus nerka): Role of local environment and fish characteristics. Can. J. Fish. Aquat. Sci. 1998, 55, 1821–1831. [Google Scholar] [CrossRef]
- Tarrade, L.; Texier, A.; David, L.; Larinier, M. Topologies and measurements of turbulent flow in vertical slot fishways. Hydrobiologia 2008, 609, 177–188. [Google Scholar] [CrossRef]
- Pavlov, D.S.; Lupandin, A.I.; Skorobogatov, M.A. The effects of flow turbulence on the behavior and distribution of fish. J. Ichthyol. 2000, 40, S232–S261. [Google Scholar]
- Larinier, M. Fish passage experience at small-scale hydro-electric power plants in France. Hydrobiology 2008, 609, 97–108. [Google Scholar] [CrossRef]
- Castro-Santos, T.; Cotel, A.; Webb, P.W. Fishway evaluations for better bioengineering–an integrative approach. In Challenges for Diadromous Fishes in a Dynamic Global Environment; Haro, A.J., Smith, K.L., Rulifson, R.A., Moffit, C.M., Klauda, R.J., Dadswell, M.J., Cunjak, R.A., Cooper, J.E., Beal, K.L., Avery, T.S., Eds.; American Fisheries Society: Bethesda, MD, USA, 2009; pp. 557–575. [Google Scholar]
- Lucas, M.C.; Frear, P.A. Effects of a flow-gauging weir on the migratory behaviour of adult barbel, a riverine cyprinid. J. Fish Boil. 1997, 50, 382–396. [Google Scholar] [CrossRef]
- Roscoe, D.W.; Hinch, S.G. Effectiveness monitoring of fish passage facilities: Historical trends, geographic patterns and future directions. Fish Fish. 2010, 11, 12–33. [Google Scholar] [CrossRef]
- Cabral, M.J.; Almeida, J.; Almeida, P.R.; Dellinger, T.; Ferrand de Almeida, N.; Oliveira, M.E.; Palmeirim, J.M.; Queiroz, A.I.; Rogado, L.; Santos-Reis, M. Livro Vermelho dos Vertebrados de Portugal; Instituto de Conservação da Natureza: Lisboa, Portugal, 2005; p. 660. [Google Scholar]
- Doadrio, I. Atlas y Libro Rojo de los Peces Continentales de España; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2002; p. 374. [Google Scholar]
- Skelton, P.H.; Tweddle, D.; Jackson, P.B.N. Cyprinids of Africa. In Cyprinid Fishes; Springer Science and Business Media LLC: Berlin, Germany, 1991; Volume 3, pp. 211–239. [Google Scholar]
- Banarescu, P.; Coad, B.W. Cyprinids of Eurasia. In Cyprinid Fishes: Systematics, Biology, and Exploitation; Springer Science and Business Media LLC: London, UK; New York, NY, USA, 1991; Volume 3, pp. 127–155. [Google Scholar]
- Migration of Freshwater Fishes; Wiley: New York, NY, USA, 2001.
- Santo, M. Dispositivos de Passagem Para Peixes em Portugal; Direcção-Geral dos Recursos Florestais: Lisboa, Portugal, 2005; p. 137.
- Silvertown, J. A new dawn for citizen science. Trends Ecol. Evol. 2009, 24, 467–471. [Google Scholar] [CrossRef]
- Mameri, D.; Rivaes, R.; Oliveira, J.; Pádua, J.; Ferreira, M.T.; Santos, J.M. Passability of Potamodromous Species through a Fish Lift at a Large Hydropower Plant (Touvedo, Portugal). Sustainability 2020, 12, 172. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Study | Ref. | Type | Main Findings |
|---|---|---|---|
| Silva et al., 2009 | [88] | NOF | Orifices clear choice to negotiate the fishway Orifices and notches equally used during streaming flow Orifices used much more than notches during plunging flow |
| Quintella et al., 2009 | [89] | OF | EMG telemetry useful to monitor barbel in pool-type fishways |
| Silva et al., 2011 | [34] | OF | Larger adults with higher passage success than smaller ones Time to negotiate the fishway lower for larger adults Horizontal RSS as the most important hydraulic variable |
| Silva et al., 2012 | [39] | OF | Offset orifice configuration better than the straight one RSS the one that most strongly influenced fish movements |
| Silva et al., 2012 | [38] | OF | Higher passage rates/lower time with offset orifices Size-related behavioral responses to turbulence RSS most important turbulence descriptor Behavior of larger fish strongly affected by eddies |
| Alexandre et al., 2013 | [41] | OF | Burst swimming was required to move through the orifices Horizontal RSS as the most important hydraulic variable |
| Branco et al., 2013 | [85] | NOF | Species used the notches more readily during streaming flow Species more successful during streaming flow Streaming flow most suitable with different species |
| Santos et al., 2013 | [86] | OF | Lower relative depth of flow more beneficial to fish passage |
| Santos et al., 2014 | [40] | OF | Fish passage success discharge-related Fish passage success independent of boulder density Higher boulder density and discharge lowered time of passage |
| Aramburu et al., 2014 | [90] | VSF | Iberian barbel had higher passage success than Mediterranean barbel and Iberian nase Larger fish were more successful in negotiating the fishway Lower fish passage at higher discharge |
| Branco et al., 2015 | [91] | OF | Boulders: greater fish passage success at higher discharge Grass: greater fish passage success at lower discharge |
| Rodríguez et al., 2015 | [80] | VSF | Fish avoided high velocity and turbulent areas Fish rested frequently in the upstream low-velocity areas of the pools |
| Sanz-Ronda et al., 2016 | [49] | VSF | Barbel and nase ascended easily the VSF Barbel and nase performed better than trout Fishway discharge affected fish motivation Fishway discharge did not influence passage success |
| Romão et al., 2017 | [69] | VSF | No difference in the number of upstream movements between the slot configurations C1 and C2. C2 is more cost-effective because it requires less water. |
| Bravo-Cordoba et al., 2018 | [92] | NOF | Environmental variables were important for fish entrance. No preference between path routes Dual-entrance fishway could be important for conservation |
| Bravo-Cordoba et al., 2018 | [42] | VSF + NOF | Ascent ability in VSF and NOF were similar Larger fish displayed lower transit time in ascension Motivation greater in VSF, though not relevant for success |
| Fuentes-Pérez et al., 2018 | [76] | VSF | Broader use of the fishway pools with lower head drop Protection by the walls was key in the use of space VSF designed with uniform flow can be unsuitable when non-uniform conditions are present |
| Romão et al., 2018 | [93] | VSF | No difference in passage performance between VSF and MSF MSF is preferable as it requires less water (31%) to operate |
| Romão et al., 2018 | [94] | VSF | No differences in fish success between spring and autumn Fish passage evaluation can be extended to early-autumn |
| Romão et al., 2019 | [95] | VSF | No differences in fish success between spring and autumn MSF can be considered a cost-effective design for the barbel |
| Pedescoll et al., 2019 | [96] | NOF | Upstream passage efficiency was the greatest for barbel (60%) Entry efficiency was low for the barbel (3.8%) Fishway has limited efficiency for small fish |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.T.; Bermúdez, M.; Santos, J.M.; Rabuñal, J.R.; Puertas, J. Pool-Type Fishway Design for a Potamodromous Cyprinid in the Iberian Peninsula: The Iberian Barbel—Synthesis and Future Directions. Sustainability 2020, 12, 3387. https://doi.org/10.3390/su12083387
Silva AT, Bermúdez M, Santos JM, Rabuñal JR, Puertas J. Pool-Type Fishway Design for a Potamodromous Cyprinid in the Iberian Peninsula: The Iberian Barbel—Synthesis and Future Directions. Sustainability. 2020; 12(8):3387. https://doi.org/10.3390/su12083387
Chicago/Turabian StyleSilva, Ana T., María Bermúdez, José M. Santos, Juan R. Rabuñal, and Jerónimo Puertas. 2020. "Pool-Type Fishway Design for a Potamodromous Cyprinid in the Iberian Peninsula: The Iberian Barbel—Synthesis and Future Directions" Sustainability 12, no. 8: 3387. https://doi.org/10.3390/su12083387
APA StyleSilva, A. T., Bermúdez, M., Santos, J. M., Rabuñal, J. R., & Puertas, J. (2020). Pool-Type Fishway Design for a Potamodromous Cyprinid in the Iberian Peninsula: The Iberian Barbel—Synthesis and Future Directions. Sustainability, 12(8), 3387. https://doi.org/10.3390/su12083387