1. Introduction
The spatial distribution of wetland plants generally shows zonation along environmental gradients [
1], which could mainly be attributed to the influences of hydrological and biotic factors [
2,
3,
4,
5]. However, because multiple factors often function together, there is still no clear consensus on the mechanism of the formation of zonation patterns and plant community development in natural wetlands. On one hand, flooding causes excessive depletion of soil oxygen, which changed soil redox conditions [
6]. As a consequence, leaf stomatal conductance, photosynthesis, and biomass were influenced by inundation directly or indirectly [
7,
8,
9]. On the other hand, the effects of the interactions between different wetland species are numerous. Ellenberg [
2] demonstrated that, when interactions with other plant species were prevented, all species had very similar hydrological optima, but in contrast, in the presence of interspecific interactions, optimal growth considerably shifted to different positions along the hydrological gradient. This finding was also supported by the concept of niche presented by Hutchinson [
10]. Further research by Silvertown [
11] suggested that the fundamental niche of a species is narrowed to the realized niche as a consequence of interspecific competition and natural enemies. In fact, the extent to what hydrological and biotic factors affect plant species distribution could be dependent on many factors. For example, Lenssen et al. [
12] found that species distribution and richness along a freshwater flooding gradient were mainly dependent on hydrology in the frequently flooded zone, whereas at higher elevations, they were mainly dependent on plant interactions.
During the process of growth and development, plants compete with adjacent individuals for water, light, and nutrients. Competition, a ubiquitous phenomenon in plant communities, has long been considered as a major force shaping plant communities [
13]. However, the role of competition in community composition remains controversial, and this was discussed in the debate between Grime and Tilman on the role of competition in the formation of plant community structure [
14]. The conclusions of this debate have been included in a new conceptual model of stress-gradient hypothesis which predicts a shift of plant interaction from competition to facilitation with increasing physical stress [
15,
16]. To date, there are still many different opinions on the competition or facilitation relationships of plants along environmental gradients [
17].
The Sanjiang Plain lies in northeast China and it was once the largest national distribution area of freshwater marsh wetlands. Although the area has experienced dramatic reduction in wetland areas because of agricultural conversions, it is still an important distribution area of natural freshwater wetlands and it plays an important role in maintaining regional biodiversity. The distribution of plant communities in the wetlands of the Sanjiang Plain generally exhibits distinct plant zonation patterns along the water level gradients [
6,
18]. Typical plant species in these wetlands include
Carex lasiocarpa Ehrh.,
Carex pseudocuraica F. Schmidt, and
Glyceria spiculosa (Schmidt.) Roshev., and they generally occupy niches with similar hydrological conditions [
19]. As a result, individuals of these species are influenced by hydrological fluctuations as well as species interactions [
8]. In recent years, research was conducted on the distribution of wetland plants of the Sanjiang Plain under different water levels [
5,
20,
21], but studies focusing on the competition between
Carex species and
G. spiculosa at different water levels are still quite limited. Considering the fact that the water level of the typical inundated freshwater wetlands in the Sanjiang Plain generally varies from 0 to 40 cm above the soil surface [
6,
20,
21], we designed a controlled experiment in order to investigate the effect of different water levels (0–30 cm) on species interactions between
C. lasiocarpa and
G. spiculosa, and between
C. pseudocuraica and
G. spiculosa. The aims of this study were to: (1) investigate the response strategies of wetland plants to flooding, (2) investigate the effect of water level on interspecific competition, and (3) compare the interspecific relationships under different competition treatments. We aimed to discuss the mechanism of wetlands species distribution driven by interspecific competition and hydrological factors.
4. Discussion
C. lasiocarpa, and
C. pseudocuraica are typical plant species in Sanjiang Plain wetlands, and they generally occupy niches with similar hydrological conditions [
19]. In the present research, a controlled experiment was set up to explore the influence of water level and interspecific competition with
G. spiculosa on these two
Carex species.
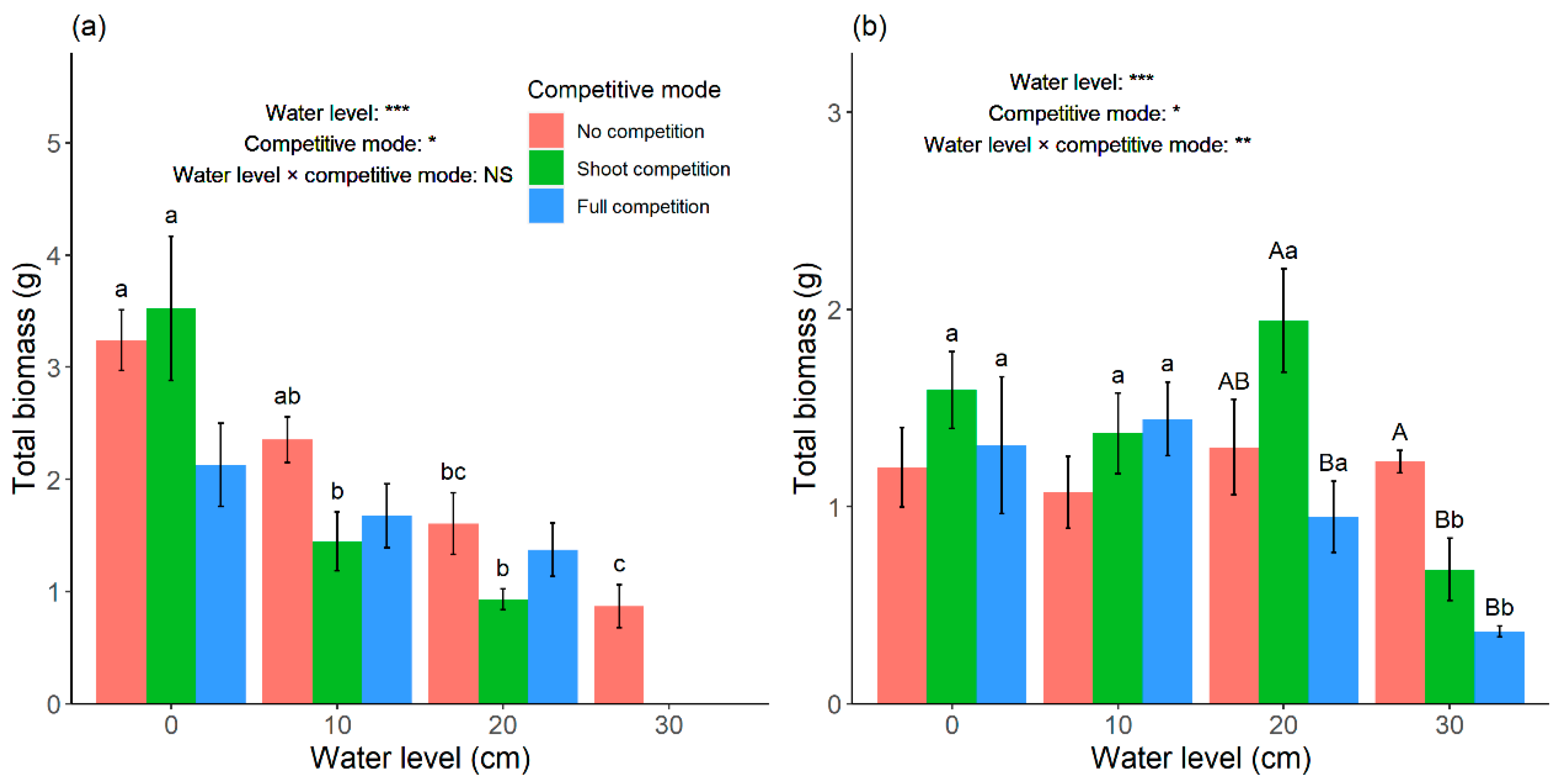
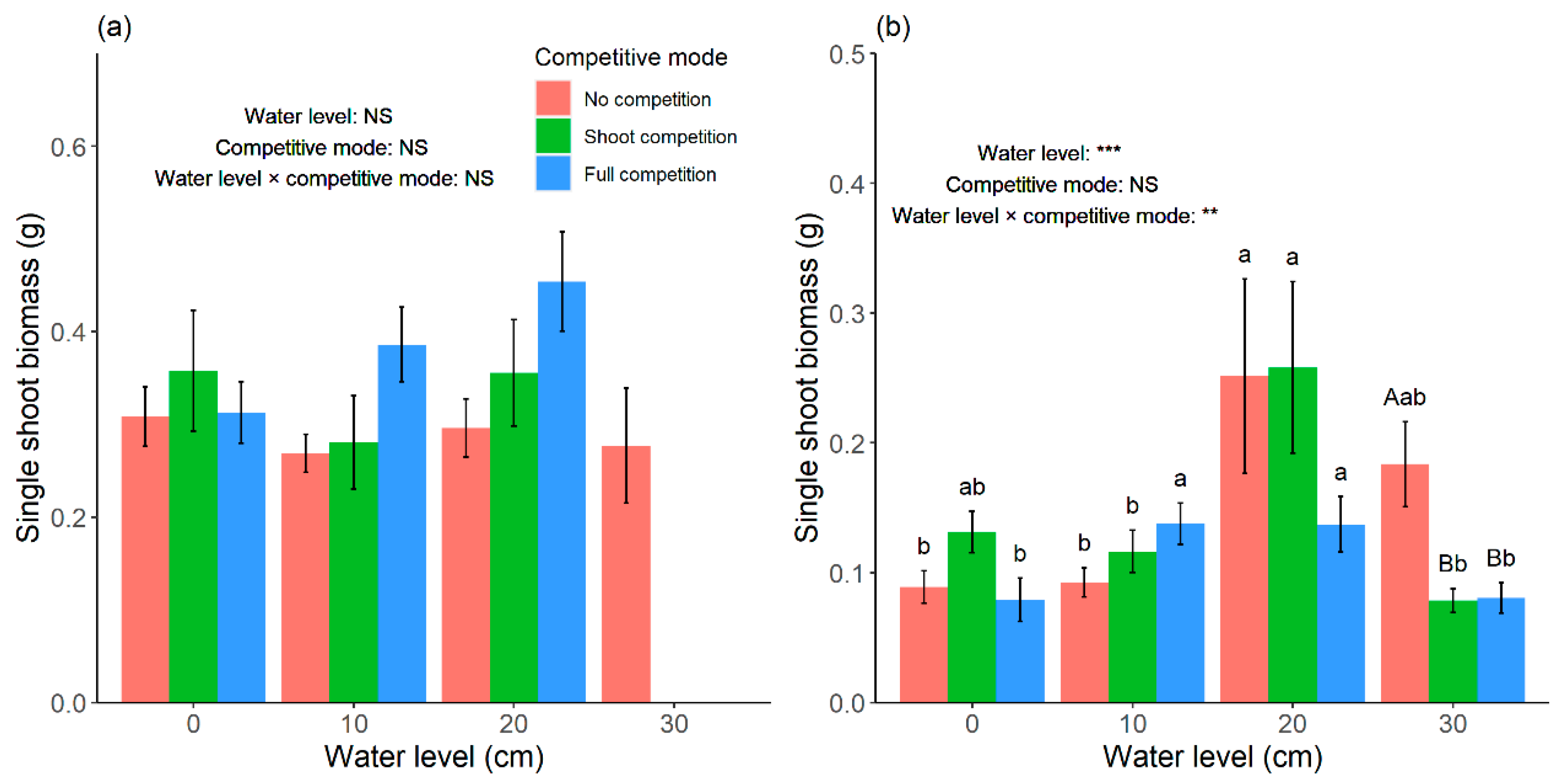
According to our results, total biomass of
C. lasiocarpa decreased as the water level increased in no competition conditions, which was consistent with the results of Luo et al. [
8] and Zhang et al. [
9]. The effects of water level on total biomass and propagule amount of
C. lasiocarpa were similar (
Figure 4a and
Figure 6a). This indicated the fundamental water level niche of
C. lasiocarpa was no more than 0 cm when evaluated by the total biomass and propagule amount indices. However, no significant differences in single shoot biomass of
C. lasiocarpa were observed among the four water levels (
Figure 5a). As
C. lasiocarpa is clonal plant, total biomass and propagule amount could represent the characteristics of a genetic individual, while single shoot biomass could represent the characteristics of a physiological individual [
29]. Our results suggested that the physiological individual and the whole genetic individual of
C. lasiocarpa use different trade-off strategies for surviving flooding stress. Genetic individuals were inhibited by deep water, whereas physiological individuals were not.
Unlike
C. lasiocarpa, there was no significant difference in total biomass and propagule amount of
C. pseudocuraica in no competition among the four water levels (
p > 0.05). When evaluated by single shoot biomass,
C. pseudocuraica preferred a deeper water level (
Figure 5b), which indicated that it had better resistance ability than
C. lasiocarpa under flooding stress. As the stems and leaves of
C. pseudocuraica are functionally plastic, e.g., they can have an increased number and thickness of vascular bundles in stems and leaves, elongated internode length, and more air cavities under flooding stress [
30,
31,
32]. Thus,
C. pseudocuraica is capable of adapting to large water level amplitude [
33]. Furthermore, because of the opposite trends of single shoot biomass and propagule amount of
C. pseudocuraica in this study, it could be inferred that in conditions of increased water levels, more resources were used for the preservation of physiological individuals than for clonal reproduction. This could also help to explain why
C. pseudocuraica was better adapted to the deep water conditions than
C. lasiocarpa during the experimental period.
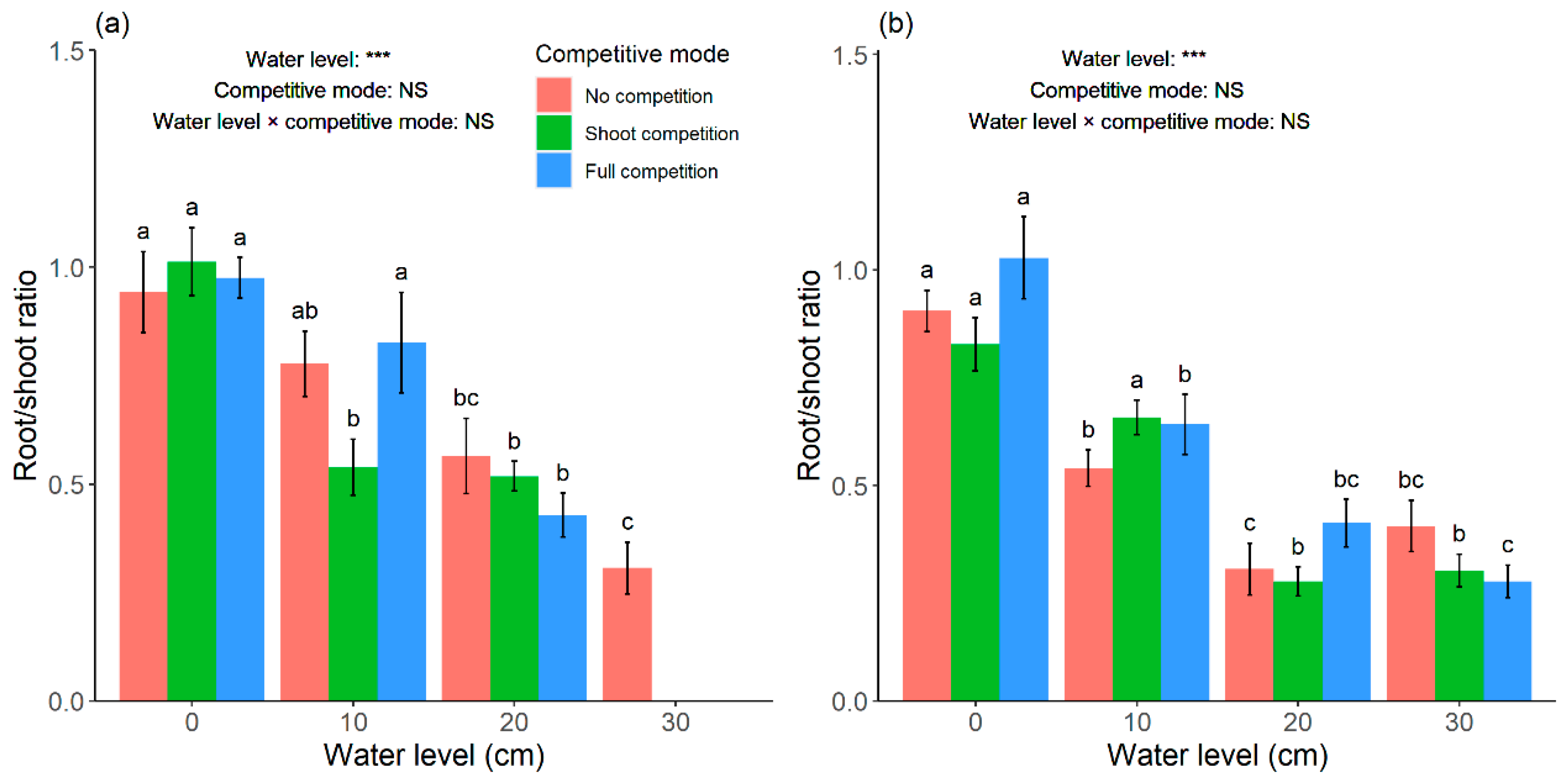
Root/shoot ratio showed a downward trend in the two
Carex species with the increase in water level in no competition (
Figure 7), which was similar to the results of Shi et al. [
22] and Zhang et al. [
9]. Interspecific competition did not significantly affect root/shoot ratio (
Figure 7), indicating that plants allocated more resources or energy to the shoots than to the roots in order to acquire more light and air, which aids photosynthesis [
34]. According to Cahill [
35], plants can alter root–shoot allocation in response to environmental conditions under certain conditions, which is also supported by the growth–balance hypothesis (optimal foraging theory), which states that plants prioritize resource allocation to organs responsible for acquiring the most limited resources [
36]. These resource allocation characteristics could explain, to some extent, the variations of root/shoot ratio under flooding stress in the present study.
In the natural inundated wetlands of the Sanjiang Plain, the optimum water level niche of
C. lasiocarpa generally varied from 13 to 29 cm with an optimal growing point of 21 cm [
20]. Similar to
C. lasiocarpa,
C. pseudocuraica was mostly distributed in the water level niche of 13–30 cm [
19]. Therefore, the realistic water level niches of
C. lasiocarpa and
C. pseudocuraica largely overlapped. This could not be the actual situation when competitions exist. In the current research, although total biomass of
C. lasiocarpa was not significantly influenced by competition modes, there existed the trend that competitive suppression by
G. spiculosa in full competition was decreasing along increasing water level (
Figure 4a). By contrast, total biomass of
C. pseudocuraica was relatively constant along water levels under no competition, but the competitive suppression by
G. spiculosa increased when water level was higher than 10 cm (
Figure 4b). Therefore, we could infer that the physiological niche of
C. lasiocarpa is emerged habitats, where the water level is 0 cm or lower; however, in a field environment, the coexistence of competitors and the relatively better adaptation of
C. lasiocarpa drive it away from emerged habitats. On the contrary, the actual water level niche of
C. pseudocuraica could tend to be far away from deep water. The interspecific competition could thus play an important role in the spatial differentiation of the actual water level niche of the two
Carex species.
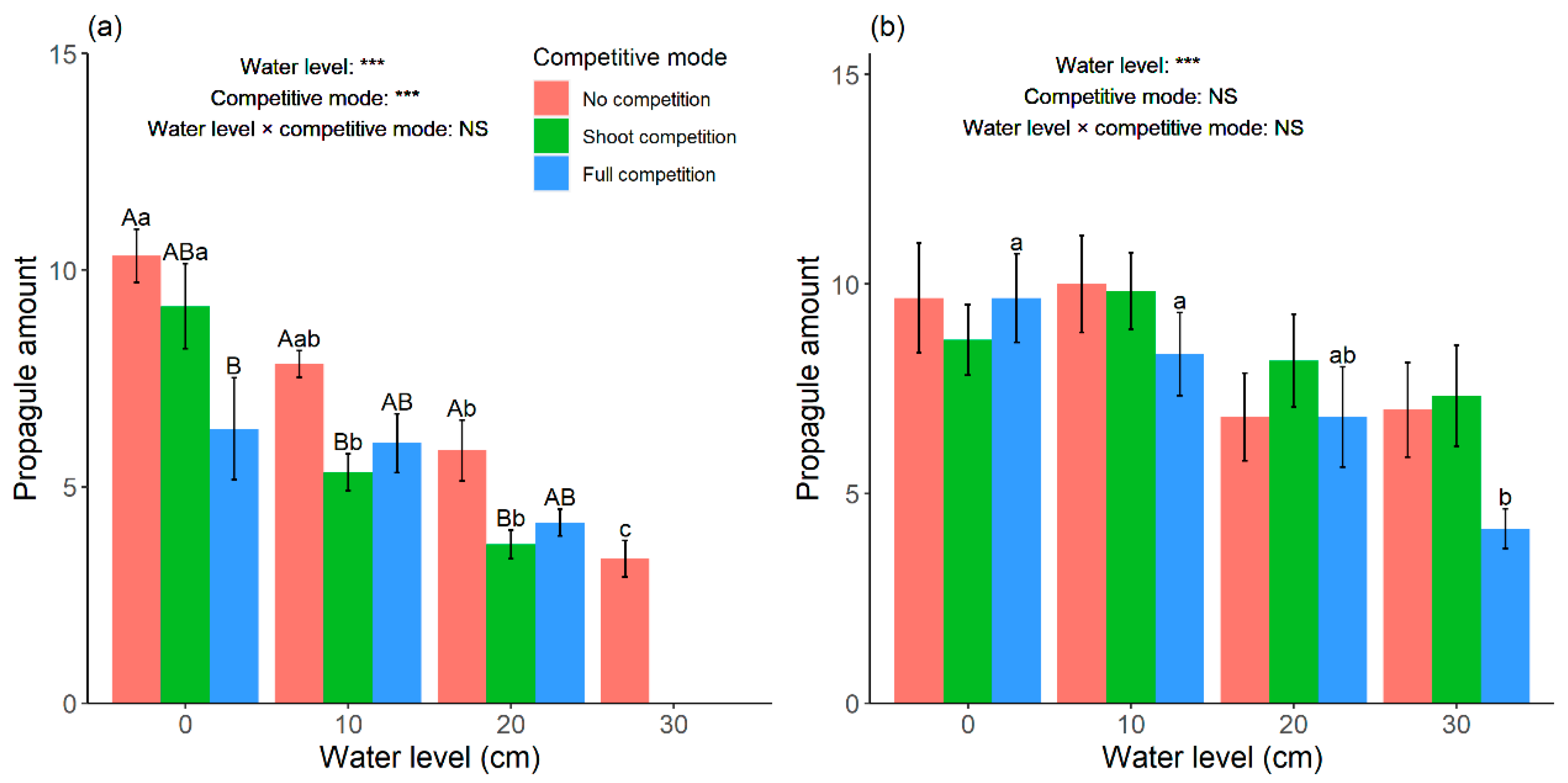
According to the variations of propagule amount under different competition modes along the increase in water level (
Figure 6a),
C. lasiocarpa tended to modulate resources allocation by reducing clonal reproduction to survive in the competition with
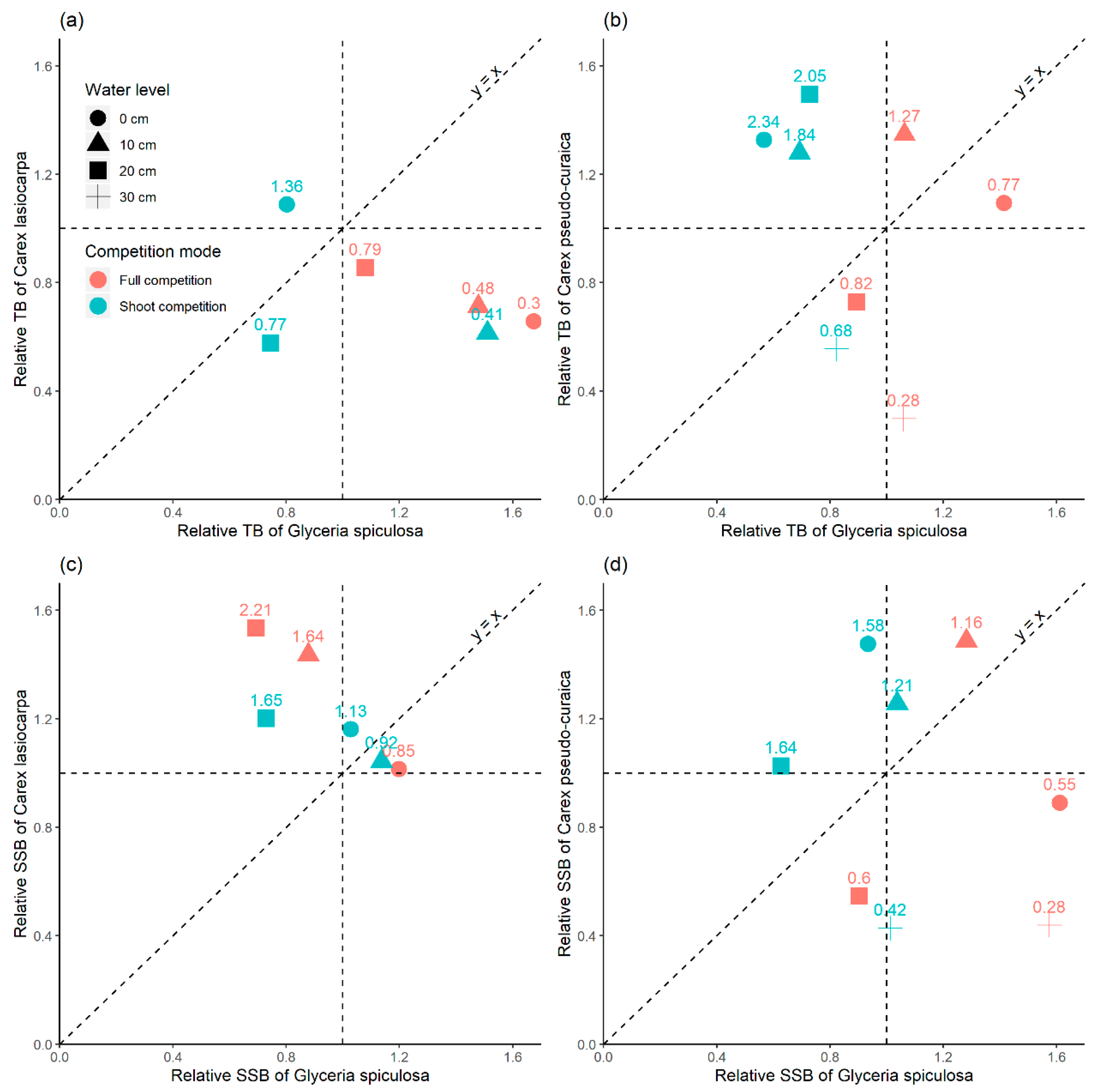
G. spiculosa. The slope change of relative single shoot biomass and relative total biomass between
C. lasiocarpa and
G. spiculosa also indicated that the stress imposed by
G. spiculosa on
C. lasiocarpa showed a decreasing trend with increasing water level (
Figure 8a,c), and this is consistent with the actual distribution of these species in natural wetland ecosystems [
19,
20]. The performances of
C. lasiocarpa and
G. spiculosa are in accordance with Ellenberg’s rule which states that competition can drive the distribution of a species away from its physiological optimum [
37]. Our results showed that based on its competitive ability and individual physiological performance,
C. lasiocarpa tended to choose a deeper water level niche in competition with
G. spiculosa compared to that in no competition. The hydrological niche segregation could mediate the coexistence of plants with similar hydrological niches and finally determine their actual distribution in wetlands.
At full competition (
C. pseudocuraica vs.
G. spiculosa), the slopes of relative total biomass and relative single shoot biomass at 0, 20 and 30 cm were smaller than 1 (
Figure 8b,d), indicating that shallow (0 cm) or relatively deep (>10 cm) water level are not optimal for the survival of
C. pseudocuraica. However,
C. pseudocuraica has greater adaptability than
G. spiculosa at 10 cm water level, which has been suggested by the high slopes (>1) of relative total biomass and relative single shoot biomass (
Figure 8b,d). According to our field observations,
C. pseudocuraica communities tend to be the first plants that colonize the waterlogged area after trampling disturbances, and their tangled roots (dead and alive) could gradually form floating organic layers, which often perform the similar function of soil. Considering the clonal reproduction mode of
C. pseudocuraica and the fact that it could be suppressed by
G. spiculosa in deep water level, we infer that
C. pseudocuraica often acts as the pioneer plant in natural wetlands and could further benefit the growth of other plants.
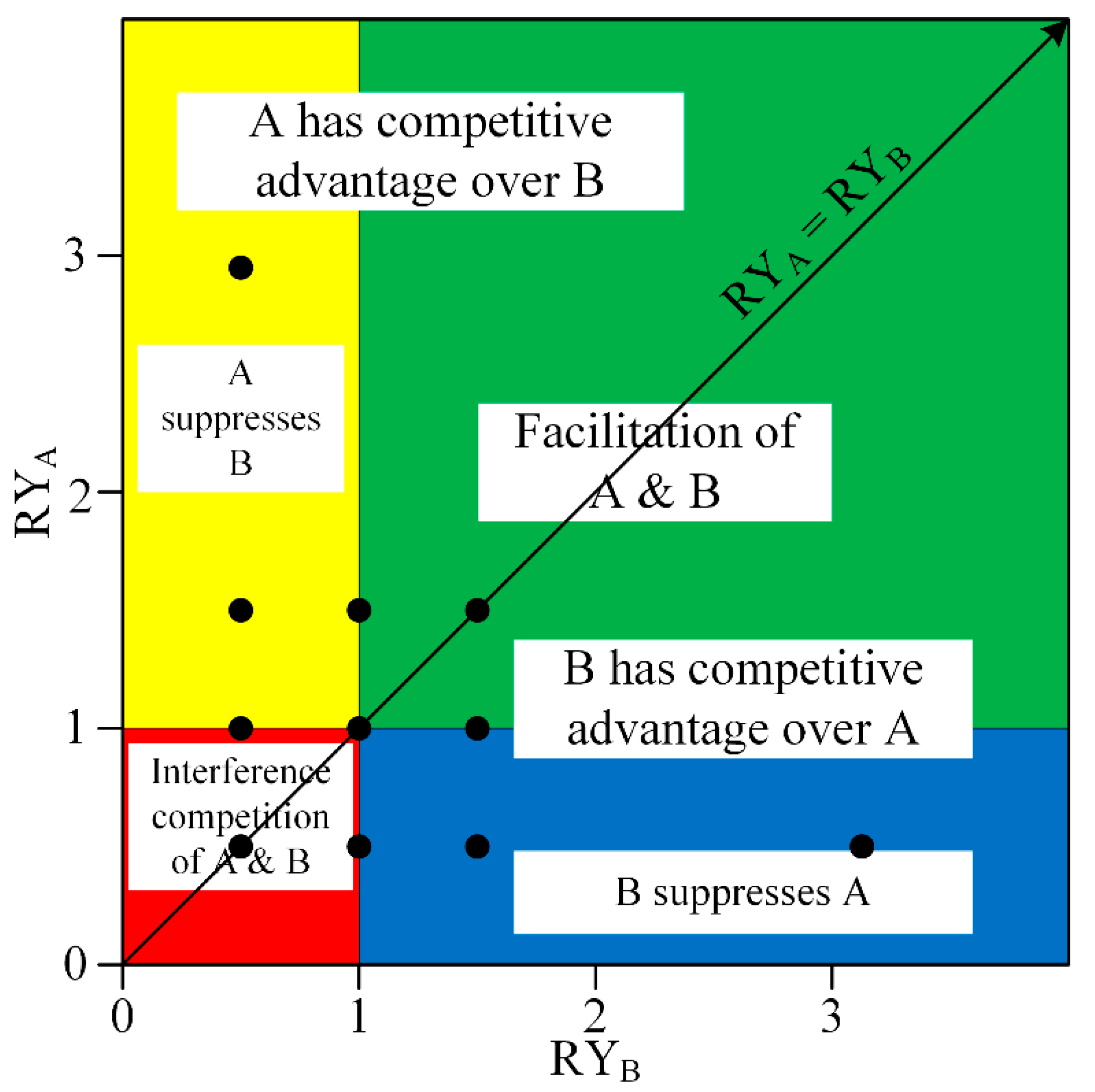
The stress-gradient hypothesis predicts a shift from net negative interactions in benign environments to net positive interactions in harsh environments [
15], which means that in
Figure 3, in cases when the flooding stress was strong enough, the relative indices should be located in top green area (Facilitation of A and B). As evaluated by relative total biomass, in the relationship between
C. lasiocarpa and
G. spiculosa, only
G. spiculosa was facilitated. Generally, in this research, facilitation changed to competition with the increase in flooding stress (
Figure 8). This may be because flooding is a non-resource stress in permanently inundated wetlands of the Sanjiang Plain. Although in permanently flooded wetlands, water availability is not a limited resource, other resources, such as rhizosphere air, could be limited by flooding. Our results were consistent with an alternative of stress-gradient hypothesis of that facilitation generally switches to competition in highly flooding stressed conditions [
17].
Competitive response is associated with traits related to resource acquisition [
38]. As root competition is more size-symmetric than shoot competition and involves multiple resources [
39], root competition and shoot competition will show different characteristics. Here, we inferred root competition ability from the difference between the slope of full competition. At 0 cm water level, shoot competitive ability of
C. lasiocarpa was greater than that of
G. spiculosa, and root competition ability weaker than that of
G. spiculosa (
Figure 8a). However, it was the opposite at 10 and 20 cm water level. The relative changes in root competition abilities between
C. lasiocarpa and
G. spiculosa along water levels could be the main reason why the realistic niche of
C. lasiocarpa was quite different from its physiological one. When comparing the relative competitive abilities between
C. pseudocuraica and
G. spiculosa, we found the shoot competitive ability of
C. pseudocuraica was greater than that of
G. spiculosa, and root competitive ability was weaker than that of
G. spiculosa at 0–20 cm water levels. At 30 cm water level, both shoot and root competitive abilities of
C. pseudocuraica were weaker than those of
G. spiculosa (
Figure 8b). These results suggested that the relatively weak position of
C. pseudocuraica when competing with
G. spiculosa could mainly be ascribed to its low root competition ability.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







