Novel Sources of Variation in Grain Yield, Components and Mineral Traits Identified in Wheat Amphidiploids Derived from Thinopyrum bessarabicum (Savul. & Rayss) Á. Löve (Poaceae) under Saline Soils in India


Abstract
1. Introduction
2. Materials and Methods
2.1. Field Locations
2.2. Planting Material
2.3. Experimental Managements
2.4. Supplementary Lighting
2.5. Trait Measurements
2.6. Grain Mineral Concentration
2.7. Statistical Analysis
3. Results
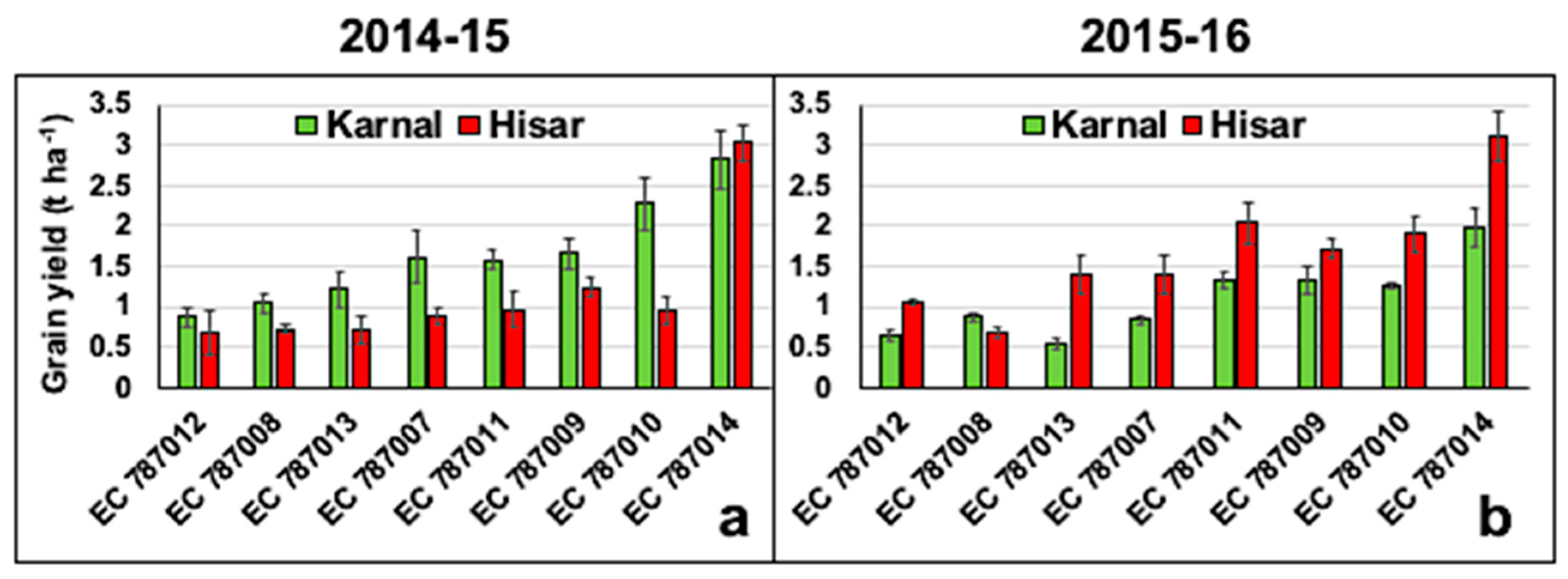
3.1. Differences in Grain Yield of Eight Amphidiploids at Karnal (Normal) and Hisar (Saline) Sites
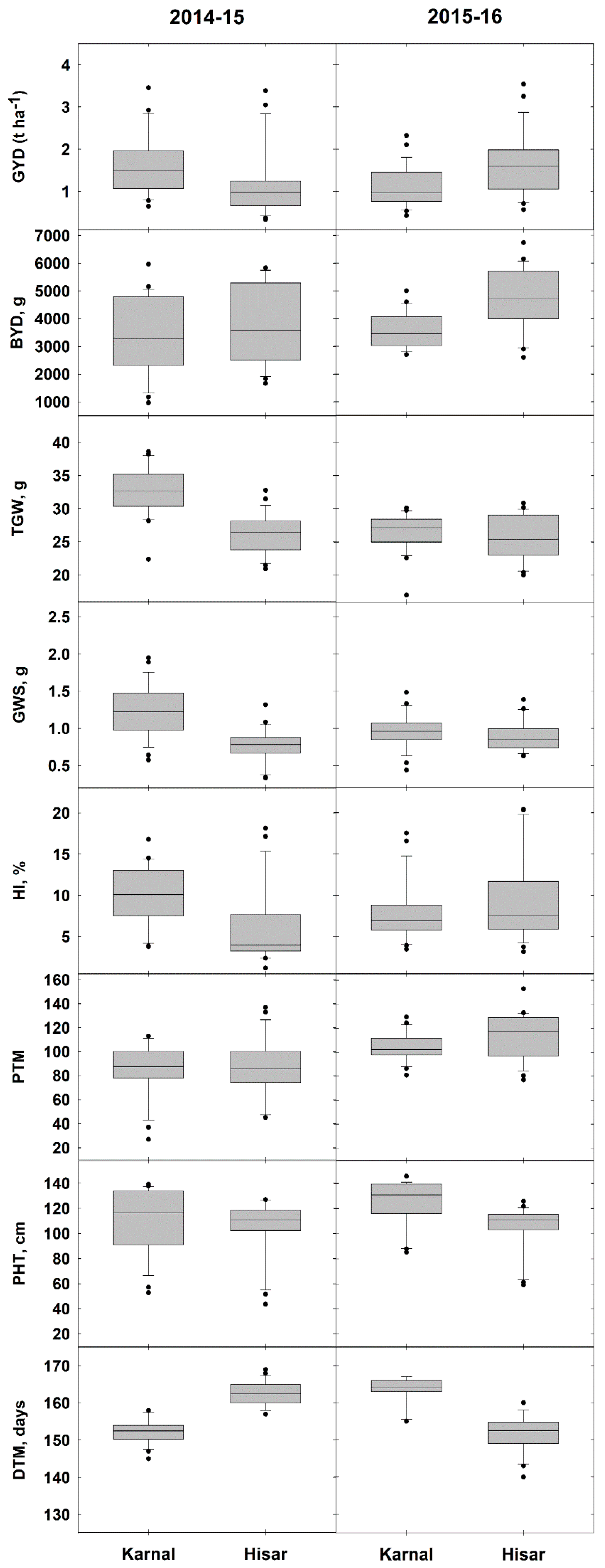
3.2. Differences in Grain Yield and Yield Component Traits between Karnal (Normal) and Hisar (Saline) Sites
3.3. Comparison of Grain Yield Reduction between Karnal (Normal) and Hisar (Saline) Sites, in 36 Indian Wheat Genotypes and Eight Amphidiploids
3.4. Variance Components Associated with Grain Yield and Component Traits
3.5. Correlation among Grain Yield and Yield Component Traits at Karnal (Normal) and Hisar (Saline) Sites
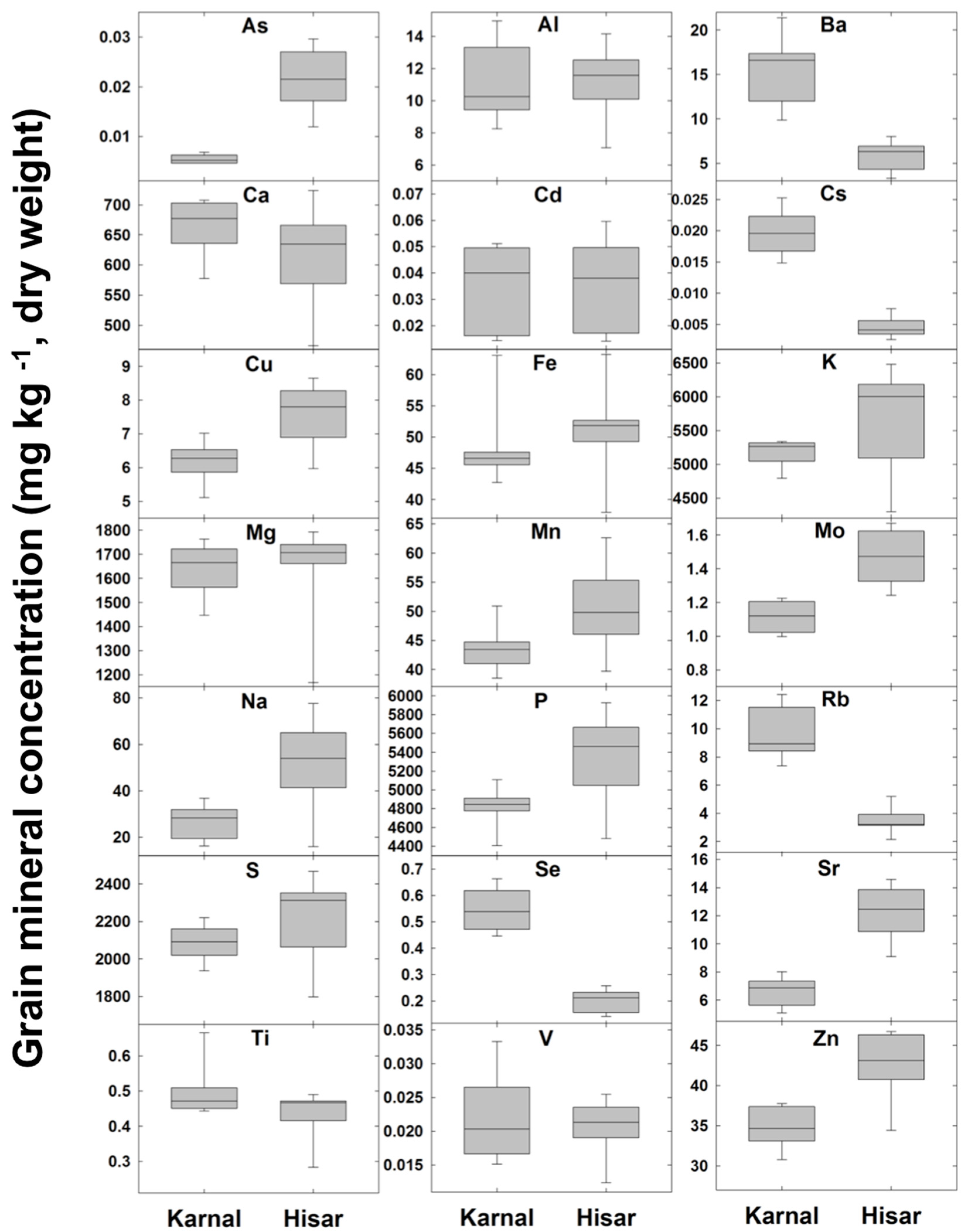
3.6. Variation in Wheat Grain Zn Concentration and the Grain Ionome
3.7. Differences in Grain Zn Concentration and the Grain Ionome between Karnal (Normal) and Hisar (Saline) Sites
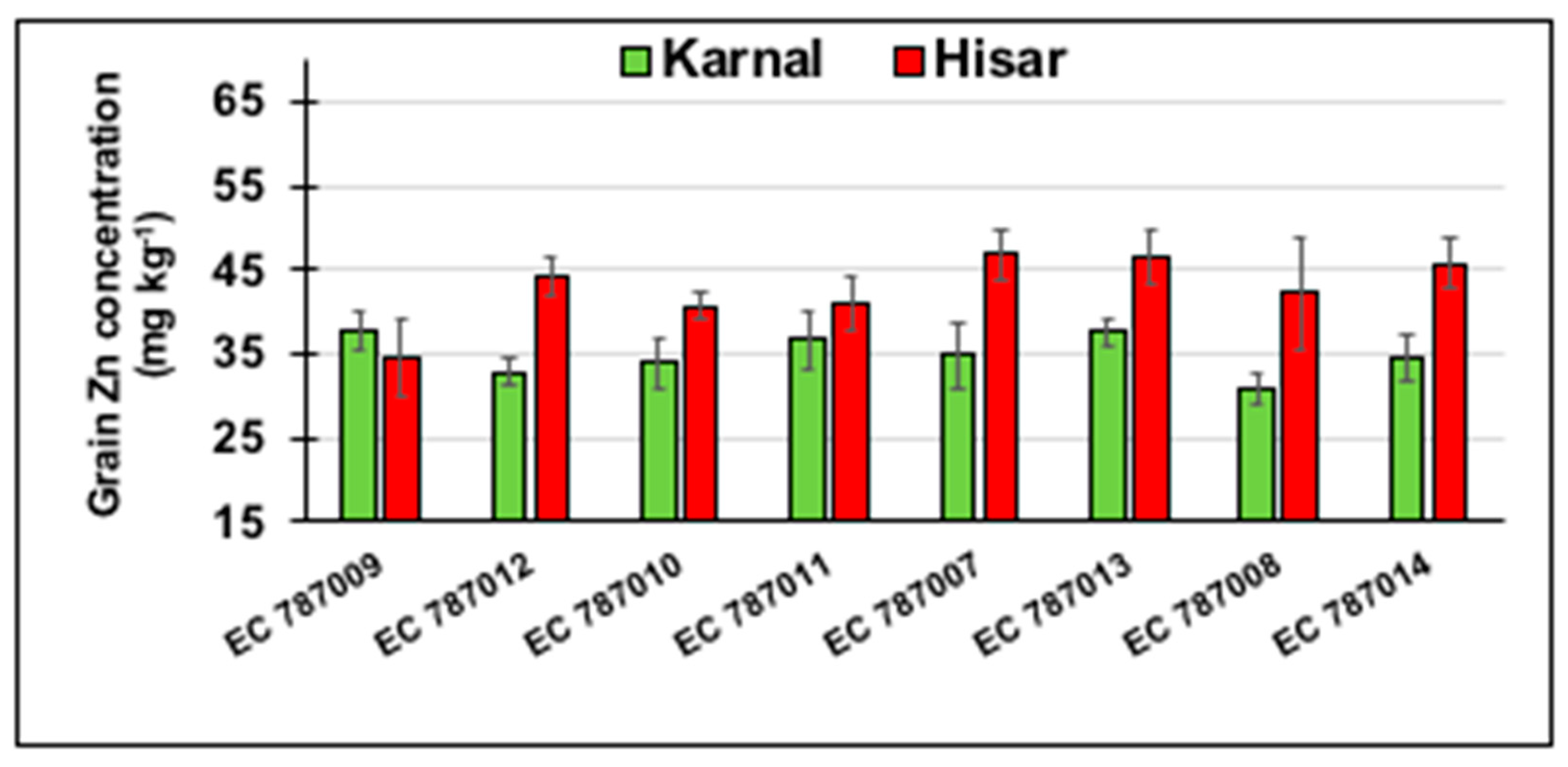
3.8. Differences in Grain Zn Concentration of Eight Amphidiploids between Karnal (Normal) and Hisar (Saline) Sites
3.9. Correlations between Grain Zn Concentration and the Grain Ionome and Grain Yield
3.10. Variance Components Associated with Grain Ionome
4. Discussion
4.1. Significant Performance of Amphidiploids for Grain Yield and Yield Components under Saline Soils
4.2. Characterisation of Amphidiploids for Grain Zn Concentration and the Grain Ionome
4.3. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shieferaw, B.; Smale, M.; Braun, H.J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef]
- United States Department of Agriculture. Available online: https:/www.fas.usda.gov/data/grain-world-markets-and-trade (accessed on 15 September 2020).
- United States Department of Agriculture. Available online: https:/www.fas.usda.gov/data/india-grain-voluntary-1 (accessed on 15 September 2020).
- King, J.C.; Brown, K.H.; Gibson, R.S.; Krebs, N.F.; Lowe, N.M.; Siekmann, J.H.; Raiten, D.J. Biomarkers of nutrition for development (BOND)-Zinc review. J. Nutr. 2016, 146, 858S–885S. [Google Scholar] [CrossRef]
- Kumssa, D.B.; Joy, E.J.M.; Ander, E.L.; Watts, M.J.; Young, S.D.; Walker, S.; Broadley, M.R. Dietary calcium and zinc deficiency risks are decreasing but remain prevalent. Sci. Rep. 2015, 5, 10974. [Google Scholar] [CrossRef]
- Gregory, P.J.; Wahbi, A.; Adu-Gyamfi, J.; Heiling, M.; Gruber, R.; Joy, E.J.M. Approaches to reduce zinc and iron deficits in food systems. Glob. Food Secur. 2017, 15, 1–10. [Google Scholar] [CrossRef]
- Zhao, F.J.; Su, Y.H.; Dunham, S.J.; Rakszrgi, M.; Bedo, Z.; McGrath, S.P.; Shewry, P.R. Variation in mineral micronutrient concentrations in grain of wheat lines of diverse origin. J. Cereal Sci. 2009, 49, 290–295. [Google Scholar] [CrossRef]
- Chen, X.P.; Zhang, Y.Q.; Tong, Y.P.; Xue, Y.F.; Liu, D.Y.; Zhang, W.; Deng, Y.; Meng, Q.F. Harvesting more grain zinc of wheat for human health. Sci. Rep. 2017, 7, 7016. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Torun, A.; Millet, E.; Feldman, M.; Fahima, T.; Korol, A.; Nevo, E.; Braun, H.J.; Ozkan, H. Triticum dicoccoides: An important genetic resource for increasing zinc and iron concentration in modern cultivated wheat. Soil Sci. 2004, 50, 1047–1054. [Google Scholar] [CrossRef]
- Rawat, N.; Tiwari, V.K.; Singh, N.; Randhawa, G.S.; Singh, K.; Chhuneja, P.; Dhaliwal, H.S. Evaluation and utilization of Aegilops and wild Triticum species for enhancing iron amnd zinc content n wheat. Genet. Resour. Crop Evol. 2008, 56, 53–64. [Google Scholar] [CrossRef]
- Zaharieva, M.; Ayana, N.G.; Al Hakimi, A.; Misra, S.C.; Monneveux, P. Cultivated emmer wheat (Triticum diccocum Schrank), an old crop with a promising future: A review. Genet. Resour. Crop Evol. 2010, 57, 937–962. [Google Scholar] [CrossRef]
- Fiore, M.C.; Mercati, F.; Spina, A.; Blangiforti, S.; Venora, G.; Dell’Acqua, M.; Lupini, A.; Preiti, G.; Monti, M.; Pe, M.E.; et al. High-throughput genotype, morphology, and quality traits evaluation for the assessment of genetic diversity of wheat Landraces from Sicily. Plant 2019, 8, 116. [Google Scholar] [CrossRef]
- Alfeo, V.; Jaskula-Goiris, B.; Venora, G.; Schimmenti, E.; Aerts, G.; Todaro, A. Screening of durum wheat landraces (Triticum turgidum subsp. durum) for the malting suitability. J. Cereal Sci. 2018, 83, 101–109. [Google Scholar] [CrossRef]
- Hasegawa, P.M. Sodium (Na+) homeostasis and salt tolerance of plants. Environ. Exp. Bot. 2013, 92, 19–31. [Google Scholar] [CrossRef]
- Pessarakli, M.; Szabolcs, I. Soil salinity and sodicity as particular plant/crop stress factors. In Handbook of Plant and Crop Stress, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 3–21. [Google Scholar]
- Mandal, A.K.; Sharma, R.C.; Singh, G.; Dagar., J.C. Computerized Database on Salt Affected Soil in India; Sehgal, V.K., Singh, R., Eds.; Central Soil Salinity Research Institute: Karnal, India, 2010; p. 28. [Google Scholar]
- Sharma, K.L.; Mandal, B.; Venkateswarlu, B. Soil Quality and Productivity Improvement under Rainfed Conditions–Indian Perspectives. In Resource Management for Sustainable Agriculture; Abrol, V., Sharma, P., Eds.; InTech: Rijeka, Croatia, 2012; pp. 203–238. [Google Scholar]
- Khokhar, J.S.; Sareen, S.; Tyagi, B.S.; Singh, G.; Chowdhury, A.K.; Dhar, T.; Singh, V.; King, I.P.; Young, S.D.; Broadley, M.R. Characterising variation in wheat traits under hostile soil conditions in India. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Rahnama, A.; Munns, R.; Poustini, K.; Watt, M. A screening method to identify genetic variation in root growth response to a salinity gradient. J. Exp. Bot. 2011, 62, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Frossard, E.; Bucher, M.; Machler, F.; Mozafar, A.; Hurrell, R. Potential for increasing the content and bioavailability of Fe, Zn and Ca in plants for human nutrition. J. Sci. Food Agr. 2000, 80, 861–879. [Google Scholar] [CrossRef]
- Gorham, J. Salt Tolerance in the Triticeae-K/Na Discrimination in Synthetic Hexaploid Wheats. J. Exp. Bot. 1990, 41, 623–627. [Google Scholar] [CrossRef]
- Gorham, J.; Bristol, A.; Young, E.M.; Wyn-Jones, R.G. The presence of the enhanced K/Na discrimination trait in diploid Triticum species. Theor. Appl. Genet. 1991, 82. [Google Scholar] [CrossRef]
- Mujeeb-Kazi, A.; De Leon, J.L.D. Conventional and alien genetic diversity for salt tolerant wheats: Focus on current status and new germplasm development. In Prospects for Saline Agriculture; Ahmad, R., Malik, K.A., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 69–82. [Google Scholar]
- Gorham, J.; Mcdonnell, E.; Budrewicz, E.; Jones, R.G.W. Salt tolerance in the Triticeae: Growth and solute accumulation in leaves of Thinopyrum bessarabicum. J. Exp. Bot. 1985, 36, 1021–1031. [Google Scholar] [CrossRef]
- King, I.P.; Forster, B.P.; Law, C.C.; Cant, K.A.; Orford, S.E.; Gorham, J.; Reader, S.; Miller, T.E. Introgression of salt-tolerance genes from Thinopyrum bessarabicum into wheat. New Phytol. 1997, 137, 75–81. [Google Scholar] [CrossRef]
- King, I.P.; Law, C.N.; Cant, K.A.; Orford, S.E.; Reader, S.M.; Miller, T.E. Tritipyrum, a potential new salt-tolerant cereal. Plant Breed. 1997, 116, 127–132. [Google Scholar] [CrossRef]
- Qi, Z.; Du, P.; Qian, B.; Zhuang, L.; Chen, H.; Chen, T.; Shen, J.; Guo, J.; Feng, Y.; Pei, Z. Characterization of a wheat-Thinopyrum bessarabicum (T2JS-2BS.2BL) translocation line. Theor. Appl. Genet. 2010, 121, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Colmer, T.D.; Flowers, T.J.; Munns, R. Use of wild relatives to improve salt tolerance in wheat. J. Exp. Bot. 2006, 57, 1059–1078. [Google Scholar] [CrossRef]
- Forster, B.P.; Gorham, J.; Miller, T.E. Salt Tolerance of an Amphiploid between Triticum aestivum and Agropyron junceum. Plant Breed. 1987, 98, 1–8. [Google Scholar] [CrossRef]
- King, I.P.; Purdie, K.A.; Orford, S.E.; Reader, S.M.; Miller, T.E. Detection of homoeologous chiasma formation in Triticum durum x Thinopyrum bessarabicum hybrids using genomic in situ hybridization. Heredity 1993, 71, 369–372. [Google Scholar] [CrossRef]
- Ardalani, S.; Mirzaghaderi, G.; Badakhshan, H. A Robertsonian translocation from Thinopyrum bessarabicum into bread wheat confers high iron and zinc contents. Plant Breed 2016, 135, 286–290. [Google Scholar] [CrossRef]
- Black, R.E.; Victoria, G.; Walker, P.; Bhutta, A.; Christian, P.; de Onis, M.; McGregor, S.G.; Katz, J.; Martorell, R.; Uauy, R. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 2013, 382, 427–451. [Google Scholar] [CrossRef]
- World Health Organisation. The Double Burden of Malnutrition. Available online: http://www.who.int/nutrition/publications/doubleburdenmalnutrition-policybrief/en (accessed on 13 September 2020).
- Khokhar, J.S.; Sareen, S.; Tyagi, B.S.; Singh, G.; Wilson, L.; King, I.P.; Young, S.D.; Broadley, M.R. Variation in grain Zn concentration, and the grain ionome, in field-grown Indian wheat. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Murphy, K.M.; Hoagland, L.A.; Reeves, P.G.; Baik, B.K.; Jones, S.S. Nutritional and quality characteristics expressed in 31 perennial wheat breeding lines. Renew. Agric. Food Syst. 2009, 24, 285–292. [Google Scholar] [CrossRef]
- Nemeth, C.; Yang, C.Y.; Kasprzak, P.; Hubbart, S.; Scholefield, D.; Mehra, S.; Skipper, E.; King, I.; King, J. Generation of amphidiploids from hybrids of wheat and related species from the genera Aegilops, Secale, Thinopyrum, and Triticum as a source of genetic variation for wheat improvement. Genome. 2015, 58, 71–79. [Google Scholar] [CrossRef]
- Grewal, S.; Yang, C.; Edward, S.H.; Scholefield, D.; Ashling, S.; Burridge, A.J.; King, I.P.; King, J. Characterisation of Thinopyrum bessarabicum chromosomes through genome-wide introgressions into wheat. Theor. Appl. Genet. 2017, 131, 389–406. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, D.; Schnurbusch, T. Variance components, heritability and correlation analysis of anther and overy size during the floral development of bread wheat. J. Exp. Bot. 2015, 66, 3099–3111. [Google Scholar] [CrossRef]
- Hassani, H.S.; King, I.P.; Reader, S.M.; Caligari, P.D.S.; Miller, T.E. Can tritipyrum, a new salt tolerant potential amphiploid, be a successful cereal like triticale? J. Agri. Sci. Tech. 2000, 2, 177–195. [Google Scholar]
- Javed, I.U.H.; Akhtar, S.; Akram, M.; Arfan, M.; Yasmin, S. Differential yield responses of barley genotypes to NaCl salinity. Int. J. Agric. Biol. 2003, 5, 233–235. [Google Scholar]
- Akbarpour, O.A.; Dehghani, H.; Rousta, M.J. Evaluation of salt stress of Iranian wheat germplasm under field conditions. Crop Pasture Sci. 2015, 66, 770–781. [Google Scholar] [CrossRef]
- Chamekh, Z.; Ayadi, S.; Karmous, C.; Trifa, Y.; Amara, H.; Boudabbous, K.; Yousfi, S.; Dolors, S.; Araus, J.L. Comparative effect of salinity on growth, grain yield, water use efficiency, delta (13)C and delta(15)N of landraces and improved durum wheat varieties. Plant Sci. 2016, 251, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Noori, S.A.S.; Khalaj, H.; Labbafi, M.R. Effect of different salinity levels on morpho-physiological characters of 8 wheat genotypes (Triticum aestivum L.). Iranian J. Plant Physio. 2010, 1, 108–117. [Google Scholar]
- Turki, N.; Harrabi, M.; Okuno, K. Effect of salinity on grain yield and quality of wheat and genetic relationships among durum and common wheat. J. Arid Land Stud. 2012, 22, 311–314. [Google Scholar]
- McDonald, G.K.; Taylor, J.D.; Verbyla, A.; Kuchel, H. Assessing the importance of subsoil constraints to yield of wheat and its implications for yield improvement. Crop Pasture Sci. 2013, 63, 1043–1065. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Islam, A.; Colmer, T.D. Hordeum marinum-wheat amphiploids maintain higher leaf K+:Na+ and suffer less leaf injury than wheat parents in saline conditions. Plant Soil. 2011, 348, 365–377. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Zhu, M.; Shabala, S.; Shabala, L.; Fan, Y.; Zhou, M.X. Evaluating predictive values of various physiological indices for salinity stress tolerance in wheat. J. Agron. Crop Sci. 2016, 202, 115–124. [Google Scholar] [CrossRef]
- Stoyanov, H. Characteristics of amphidiploids in the Triticum-Aegilops-Haynaldia-Agropyron group. Scientific Papers-Series A. Agronomy 2012, 55, 249–255. [Google Scholar]
- Ghazali, S.; Mirzaghaderi, G.; Majdi, M. Production of a novel Robertsonian translocation from Thinopyrum bessarabicum into bread wheat. Cytol. Genet. 2015, 49, 378–381. [Google Scholar] [CrossRef]
- Marais, F.; Simsek, S.; Poudel, R.S.; Cookman, D.; Somo, M. Development of hexaploid (AABBJJ) Tritipyrums with rearranged Thinopyrum distichum (Thunb.) A love-derived genomes. Crop Sci. 2014, 54, 2619–2630. [Google Scholar] [CrossRef]
- Singh, R.; Velu, G. Zinc-biofortified wheat: Harnessing genetic diversity for improved nutritional quality. In Science Brief: Biofortification; Andersson, M.S., Bouis, H., Eds.; CIMMYT, HarvestPlus and the Global Crop Diversity Trust: Bonn, Germany, 2017. [Google Scholar]
- Joshi, A.K.; Crossa, J.; Arun, B.; Chand, R.; Trethowan, R.; Vargas, M.; Ortiz-Monasterio, I. Genotype x environment interaction for zinc and iron concentration of wheat grain in eastern Gangetic plains of India. Field Crop Res. 2010, 116, 268–277. [Google Scholar] [CrossRef]
- Welch, R.M.; Graham, R.D. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef]
- Ajiboye, B.; Cakmak, I.; Paterson, D.; de Jonge, M.D.; Howard, D.L.; Stacey, S.P.; Torun, A.A.; Aydin, N.; McLaughlin, M.J. X-ray fluorescence microscopy of zinc localization in wheat grains biofortified through foliar zinc applications at different growth stages under field conditions. Plant Soil 2015, 392, 357–370. [Google Scholar] [CrossRef]
- Cakmak, I.; Kalayci, M.; Kaya, Y.; Torun, A.A.; Aydin, N.; Wang, Y.; Arisoy, Z.; Erdem, H.; Yazici, A.; Gokmen, O.; et al. Biofortification and localization of zinc in wheat grain. J. Agric. Food Chem. 2010, 58, 9092–9102. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Growing Zone | Grid | Sowing | Harvesting | Year | Soil Type | Cropping Pattern |
|---|---|---|---|---|---|---|---|
| IIWBR, Karnal | NWPZ | 29.70° N | 18.11.14 | 20.04.15 | 2014–2015 | Normal | Rice–Wheat |
| 76.99° E | 12.11.15 | 23.04.16 | 2015–2016 | ||||
| IIWBR, Hisar | NWPZ | 29.18° N | 27.11.14 | 24.04.15 | 2014–2015 | Saline | Cotton–Wheat |
| 75.70° E | 24.11.15 | 22.04.16 | 2015–2016 |
| Amphidiploids | Pedigree | Genomic Constitution |
|---|---|---|
| EC 787007 | Triticum. turgidum L. cv. Langdon x Thinopyrum bessarabicum (Savul. & Rayss) Á. Löve (Poaceae) | AABBJJ |
| EC 787008 | T. turgidum L. cv. Macoun x T. bessarabicum | AABBJJ |
| EC 787009 | T. turgidum L. cv. Karim x T. bessarabicum | AABBJJ |
| EC 787010 | T. turgidum L. cv. Neodur x T. bessarabicum | AABBJJ |
| EC 787011 | T. turgidum L. cv. Creso x T. bessarabicum | AABBJJ |
| EC 787012 | T. turgidum L. cv. Azaziah x T. bessarabicum | AABBJJ |
| EC 787013 | T. turgidum L. cv. Stewart x T. bessarabicum | AABBJJ |
| EC 787014 | Triticum aestivum L. cv. Chinese Spring x T. bessarabicum | AABBDDJJ |
| Traits | Variation (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| G | E | G*E | Residual | H2 | |||||
| 2015 | 2016 | 2015 | 2016 | 2015 | 2016 | 2015 | 2016 | 2015 + 2016 | |
| Grain yield per plot (GYD) | 67.7 ** | 63.7 ** | 9.8 ** | 16.7 ** | 7.2 | 7.0 | 15.3 | 12.6 | 39.1 |
| Biological yield (BYD) | 77.5 ** | 41.6 ** | 3.1 * | 27.3 ** | 6.2 | 13.3 * | 13.2 | 17.8 | 49.8 |
| Germination % (Germ. %) | 67.0 ** | 67.1 ** | 4.9 * | 14.5 ** | 13.9 * | 4.4 | 14.1 | 14.0 | 77.7 |
| 1000 grain weight (TGW) | 18.8 | 25.1 | 49.3 ** | 2.0 | 4.4 | 26.0 | 27.6 | 46.8 | 56.3 |
| Grain weight per spike (GWS) | 13.2 | 50.6 ** | 41.5 ** | 2.5 | 9.8 | 4.7 | 35.5 | 42.2 | 64.9 |
| Harvest Index (HI) | 58.0 ** | 84.8 ** | 16.2 ** | 3.1 ** | 15.5 ** | 5.5 * | 10.2 | 6.6 | 59.6 |
| Productive tillers per meter (PTM) | 45.9 ** | 23.1 | 0.1 | 8.1 | 15.4 | 22.9 | 38.6 | 46.0 | 40.8 |
| Plant height at maturity (PHT) | 88.3 ** | 65.6 ** | 0.8 | 23.9 ** | 7.2 ** | 5.7 ** | 3.7 | 4.7 | 82.4 |
| Days to maturity (DTM) | 15.0 ** | 26.8 ** | 71.9 ** | 67.5 ** | 6.9 | 2.9 * | 6.2 | 2.8 | 89.1 |
| Degrees of freedom (d.f.) | 7 | 1 | 7 | 32 | |||||
| Elements | Mean | Median | SD | Range | LOD | Heritability |
|---|---|---|---|---|---|---|
| Zn | 39.7 | 40.0 | 8.20 | 21.2–58.9 | 0.572 | 43.7 |
| Others | ||||||
| As | 0.0154 | 0.0106 | 0.0146 | 0.0016–0.0646 | 0.00319 | 45.7 |
| Al | 11.1 | 11.1 | 4.22 | 2.26–26.3 | 4.53 | 27.3 |
| Ba | 9.47 | 8.2 | 6.1 | 2–24.1 | 0.0291 | 78.0 |
| Ca | 636 | 631 | 94.8 | 273–819 | 7.65 | 77.3 |
| Cd | 0.0351 | 0.033 | 0.0196 | 0.00634–0.0999 | 0.00121 | 56.4 |
| Cs | 0.0102 | 0.00586 | 0.00853 | 0.000271–0.0302 | 0.000542 | 58.5 |
| Cu | 7.01 | 7 | 1.44 | 3.53–9.99 | 0.355 | 69.4 |
| Fe | 50.1 | 49.3 | 8.33 | 22.1–72.8 | 4.76 | 56.0 |
| K | 5520 | 5660 | 843 | 2920–7140 | 2.02 | 77.6 |
| Mg | 1650 | 1670 | 217 | 643–1950 | 0.942 | 62.4 |
| Mn | 47.8 | 47.8 | 9.24 | 18.9–69.8 | 0.238 | 65.3 |
| Mo | 1.34 | 1.3 | 0.33 | 0.609–2.36 | 0.077 | 67.5 |
| Na | 42.3 | 34.2 | 25.4 | 13–103 | 5.39 | 56.0 |
| P | 5160 | 5220 | 657 | 2530–6590 | 1.84 | 48.3 |
| Rb | 5.78 | 4.45 | 3.77 | 0.468–15.2 | 0.00599 | 73.6 |
| S | 2180 | 2170 | 257 | 947–2620 | 106 | 59.9 |
| Se | 0.331 | 0.289 | 0.206 | 0.0912–0.911 | 0.00342 | 01.2 |
| Sr | 10.1 | 9.66 | 3.92 | 4.18-18.4 | 0.0886 | 34.9 |
| Ti | 0.46 | 0.456 | 0.117 | 0.126–0.736 | 0.251 | 29.1 |
| V | 0.0212 | 0.0201 | 0.00833 | 0.00251–0.0468 | 0.00503 | 09.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khokhar, J.S.; Sareen, S.; Tyagi, B.S.; Wilson, L.; Young, S.; King, J.; King, I.; Broadley, M.R. Novel Sources of Variation in Grain Yield, Components and Mineral Traits Identified in Wheat Amphidiploids Derived from Thinopyrum bessarabicum (Savul. & Rayss) Á. Löve (Poaceae) under Saline Soils in India. Sustainability 2020, 12, 8975. https://doi.org/10.3390/su12218975
Khokhar JS, Sareen S, Tyagi BS, Wilson L, Young S, King J, King I, Broadley MR. Novel Sources of Variation in Grain Yield, Components and Mineral Traits Identified in Wheat Amphidiploids Derived from Thinopyrum bessarabicum (Savul. & Rayss) Á. Löve (Poaceae) under Saline Soils in India. Sustainability. 2020; 12(21):8975. https://doi.org/10.3390/su12218975
Chicago/Turabian StyleKhokhar, Jaswant Singh, Sindhu Sareen, Bhudeva Singh Tyagi, Lolita Wilson, Scott Young, Julie King, Ian King, and Martin R. Broadley. 2020. "Novel Sources of Variation in Grain Yield, Components and Mineral Traits Identified in Wheat Amphidiploids Derived from Thinopyrum bessarabicum (Savul. & Rayss) Á. Löve (Poaceae) under Saline Soils in India" Sustainability 12, no. 21: 8975. https://doi.org/10.3390/su12218975
APA StyleKhokhar, J. S., Sareen, S., Tyagi, B. S., Wilson, L., Young, S., King, J., King, I., & Broadley, M. R. (2020). Novel Sources of Variation in Grain Yield, Components and Mineral Traits Identified in Wheat Amphidiploids Derived from Thinopyrum bessarabicum (Savul. & Rayss) Á. Löve (Poaceae) under Saline Soils in India. Sustainability, 12(21), 8975. https://doi.org/10.3390/su12218975