Validation of Francis–Kaplan Turbine Blade Strike Models for Adult and Juvenile Atlantic Salmon (Salmo Salar, L.) and Anadromous Brown Trout (Salmo Trutta, L.) Passing High Head Turbines

Abstract
1. Introduction
2. Materials and Methods
2.1. The Turbines
2.2. Telemetry Fish Tracking Studies
2.3. Validation Approach
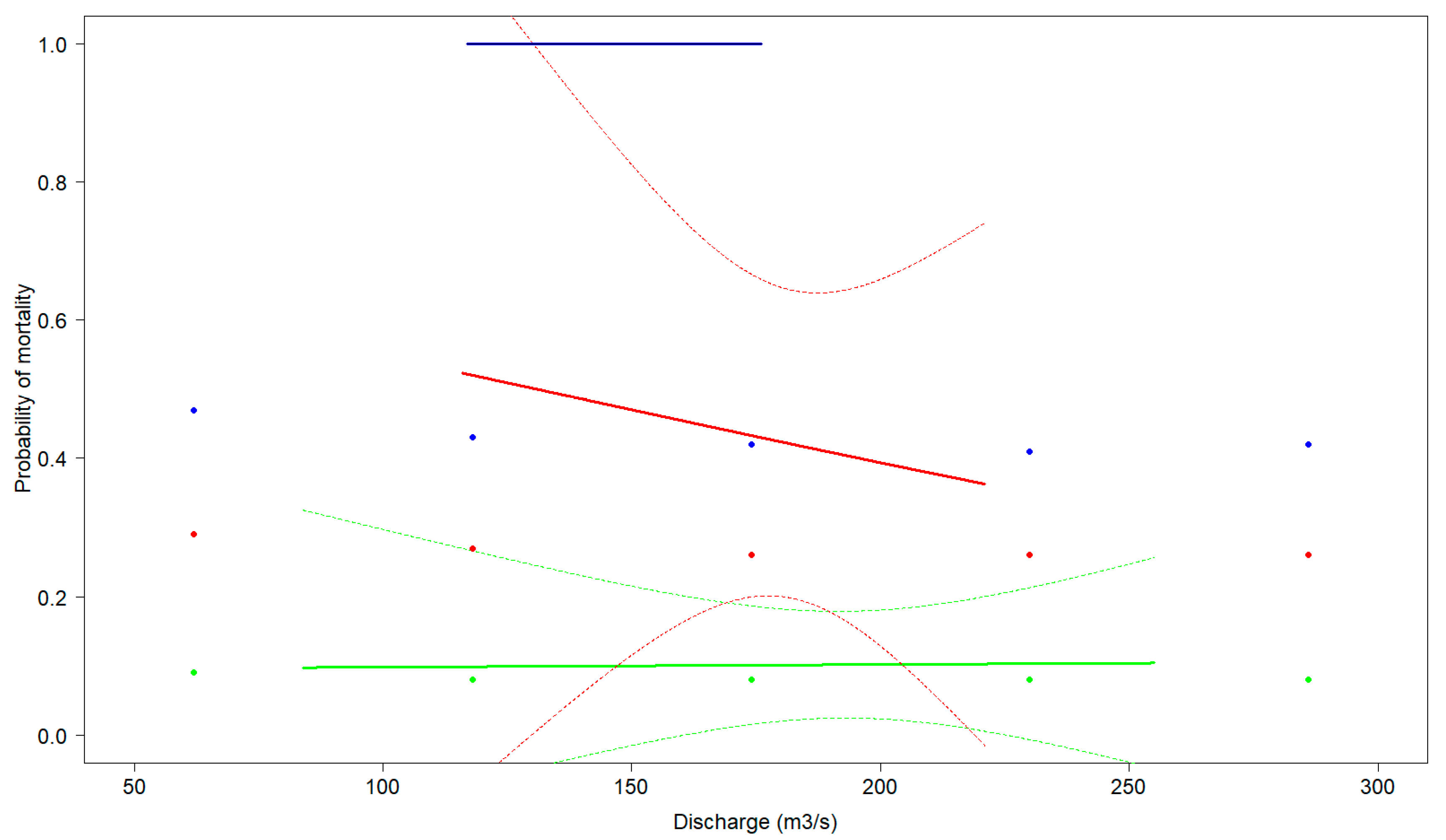
3. Results
3.1. Telemetry Turbine Passage Mortality
3.2. Validation of the Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bevelhimer, M.; Pracheil, B.M.; Fortner, A.M.; Saylor, R.; Deck, K.L. Mortality and injury assessment for three species of fish exposed to simulated turbine blade strike. Can. J. Fish. Aquat. Sci. 2019, 76, 2350–2363. [Google Scholar] [CrossRef]
- Silva, A.T.; Lucas, M.C.; Castro-Santos, T.; Katopodis, C.; Baumgartner, L.J.; Thiem, J.D.; Aarestrup, K.; Pompeu, P.S.; O’Brien, G.C.; Braun, D.C.; et al. The future of fish passage science, engineering, and practice. Fish Fish. 2017, 19, 340–362. [Google Scholar] [CrossRef]
- Pracheil, B.M.; DeRolph, C.R.; Schramm, M.P.; Bevelhimer, M.S. A fish-eye view of riverine hydropower systems: The current understanding of the biological response to turbine passage. Rev. Fish Boil. Fish. 2016, 26, 153–167. [Google Scholar] [CrossRef]
- Calles, O.; Olsson, I.C.; Comoglio, C.; Kemp, P.S.; Blunden, L.; Schmitz, M.; Greenberg, L.A. APPLIED ISSUES: Size-dependent mortality of migratory silver eels at a hydropower plant, and implications for escapement to the sea. Freshw. Boil. 2010, 55, 2167–2180. [Google Scholar] [CrossRef]
- Heisey, P.G.; Mathur, D.; Phipps, J.L.; Avalos, J.C.; Hoffman, C.E.; Adams, S.W.; De-Oliveira, E. Passage survival of European and American eels at Francis and propeller turbines. J. Fish Boil. 2019, 95, 1172–1183. [Google Scholar] [CrossRef]
- Deng, Z.; Carlson, T.J.; Duncan, J.P.; Richmond, M.C.; Dauble, D.D. Use of an autonomous sensor to evaluate the biological performance of the advanced turbine at Wanapum Dam. J. Renew. Sustain. Energy 2010, 2, 053104. [Google Scholar] [CrossRef]
- Colotelo, A.H.; Goldman, A.E.; Wagner, K.A.; Brown, R.S.; Deng, Z.; Richmond, M.C. A comparison of metrics to evaluate the effects of hydro-facility passage stressors on fish. Environ. Rev. 2017, 25, 1–11. [Google Scholar] [CrossRef]
- Montén, E. Fish and Turbines: Fish Injuries During Passage Through Power Station Turbines; Norstedts Tryckeri: Stockholm, Sweden, 1985; pp. 1–111. [Google Scholar]
- Von Raben, K. Regarding the Problem of Mutilations of Fishes by Hydraulic Turbines. Wasserwirtschaft 1957, 4, 97–100. [Google Scholar]
- Turnpenny, A.W. Mechanisms of fish damage in low head turbines: An experimental appraisal. In Fish Migration and Fish Bypasses; Fishing News Books: Farnham, UK, 1998. [Google Scholar]
- Van Esch, B.P.M.; Spierts, I. Validation of a model to predict fish passage mortality in pumping stations. Can. J. Fish. Aquat. Sci. 2014, 71, 1910–1923. [Google Scholar] [CrossRef]
- Skalski, J.R.; Mathur, D.; Heisey, P.G. Effects of Turbine Operating Efficiency on Smolt Passage Survival. N. Am. J. Fish. Manag. 2002, 22, 1193–1200. [Google Scholar] [CrossRef]
- Cada, G.F. The Development of Advanced Hydroelectric Turbines to Improve Fish Passage Survival. Fish 2001, 26, 14–23. [Google Scholar] [CrossRef]
- Ruggles, C.; Palmeter, T. Fish Passage Mortality in a Tube Turbine; Fisheries and Oceans: Ottawa, ON, Canada, 1989. [Google Scholar]
- Larinier, M.; Travade, F. Downstream Migration: Problems and Facilities. Bull. Français Pêche Piscicult. 2002, 364, 181–207. [Google Scholar] [CrossRef]
- Larinier, M. Fish passage experience at small-scale hydro-electric power plants in France. Hydrobiologia 2008, 609, 97–108. [Google Scholar] [CrossRef]
- Coutant, C.C.; Whitney, R.R. Fish Behavior in Relation to Passage through Hydropower Turbines: A Review. Trans. Am. Fish. Soc. 2000, 129, 351–380. [Google Scholar] [CrossRef]
- Bell, M.C. Revised Compendium on the Success of Passage of Small Fish Through Turbines; Report to the U.S. Army Corps of Engineers, North Pacific Division: Portland, OR, USA, 1991. [Google Scholar]
- Turnpenny, A.W.; Clough, S.; Hanson, K.P.; Ramsey, R.; McEwan, D. Risk Assessment for Fish Passage Through Small Low-Head Turbines; Atomic Energy Research Establishment: Harwell, UK, 2000. [Google Scholar]
- Pavlov, D.S.; Lupandin, A.I.; Kostin, V. Downstream Migration of Fish Through Dams of Hydroelectric Power Plants. Oak Ridge National Laboratory Oak Ridge; Nauka: Moscow, Russia, 1999. [Google Scholar]
- Ploskey, G.R.; Carlson, T.J. Comparison of Blade-Strike Modeling Results with Empirical Data; EERE Publication and Product Library: Washington, DC, USA, 2004. [Google Scholar]
- Deng, Z.D.; Carlson, T.J.; Ploskey, G.R.; Richmond, M.C.; Dauble, D.D. Evaluation of blade-strike models for estimating the biological performance of Kaplan turbines. Ecol. Model. 2007, 208, 165–176. [Google Scholar] [CrossRef]
- Deng, Z.; Carlson, T.J.; Dauble, D.D.; Ploskey, G.R. Fish passage assessment of an advanced hydropower turbine and conventional turbine using blade-strike modeling. Energies 2011, 4, 57–67. [Google Scholar] [CrossRef]
- Ferguson, J.W.; Ploskey, G.R.; Leonardsson, K.; Zabel, R.W.; Lundqvist, H. Combining turbine blade-strike and life cycle models to assess mitigation strategies for fish passing dams. Can. J. Fish. Aquat. Sci. 2008, 65, 1568–1585. [Google Scholar] [CrossRef]
- Solomon, D.J. Fish Passage through Tidal Energy Barrages; Energy Technology Support Unit: Edinburgh, UK, 1988. [Google Scholar]
- Halttunen, E. Staying Alive: The Survival and Importance of Atlantic Salmon Post-Spawners. Ph.D. Thesis, University of Tromsø, Tromsø, Norway, 19 August 2011. [Google Scholar]
- Fu, T.; Deng, Z.D.; Duncan, J.P.; Zhou, D.; Carlson, T.J.; Johnson, G.E.; Hou, H. Assessing hydraulic conditions through Francis turbines using an autonomous sensor device. Renew. Energy 2016, 99, 1244–1252. [Google Scholar] [CrossRef]
- Leonardsson, K. Modellverktyg för Beräkning av Ålförluster vid Vattenkraftverk; ELFORSK-Rapport; Finns att hämta på ELFORSK: Stockholm, Sweden, 2012. [Google Scholar]
- Saylor, R.; Sterling, D.; Bevelhimer, M.; Pracheil, B.M. Within and Among Fish Species Differences in Simulated Turbine Blade Strike Mortality: Limits on the Use of Surrogacy for Untested Species. Water 2020, 12, 701. [Google Scholar] [CrossRef]
- Axén, C.; Koski, P. Laxdöden i Torneälven Lohikuolemat Tornionjoella Salmon Deaths in Torne river 2014—2016; Report of a Swedish-Finnish Survey; Havs-och Vattenmyndigheten: Göteborg, Sweden, 2017; p. 92. [Google Scholar]
- Axén, C.; Sturve, J.; Weichert, F. Fortsatta Undersökningar av Laxsjuklighet under 2018; Report of Continued Surveys 2018; Havs-och Vattenmyndigheten: Göteborg, Sweden, 2019; p. 43. [Google Scholar]
- Bevelhimer, M.S.; Pracheil, B.M.; Fortner, A.M.; Deck, K.L. An Overview of Experimental Efforts to Understand the Mechanisms of Fish Injury and Mortality Caused by Hydropower Turbine Blade Strike; Oak Ridge National Lab: Oak Ridge, TN, USA, 2017. [Google Scholar]
- Östergren, J. Migration and Genetic Structure of Salmo Salar and Salmo Trutta in Northern Swedish Rivers. Ph.D. Thesis, Faculty of Forest Sciences, Uppsala, Sweden, December 2006. [Google Scholar]
- Deng, Z.; Carlson, T.J.; Ploskey, G.R.; Richmond, M.C. Evaluation of Blade-Strike Models for Estimating the Biological Performance of Large Kaplan Hydro Turbines; Pacific Northwest National Lab: Richland, WA, USA, 2006. [Google Scholar]
- Klopries, E.-M.; Schüttrumpf, H. Mortality assessment for adult European eels (Anguilla Anguilla) during turbine passage using CFD modelling. Renew. Energy 2020, 147, 1481–1490. [Google Scholar] [CrossRef]
- Brown, R.S.; Colotelo, A.H.; Pflugrath, B.D.; Boys, C.; Baumgartner, L.J.; Deng, Z.; Silva, L.G.; Brauner, C.J.; Mallen-Cooper, M.; Phonekhampeng, O.; et al. Understanding Barotrauma in Fish Passing Hydro Structures: A Global Strategy for Sustainable Development of Water Resources. Fish 2014, 39, 108–122. [Google Scholar] [CrossRef]
- Čada, G.; Loar, J.; Garrison, L.; Fisher, R.; Neitzel, D. Efforts to Reduce Mortality to Hydroelectric Turbine-Passed Fish: Locating and Quantifying Damaging Shear Stresses. Environ. Manag. 2006, 37, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.S.; Carlson, T.J.; Gingerich, A.J.; Stephenson, J.R.; Pflugrath, B.D.; Welch, A.E.; Langeslay, M.J.; Ahmann, M.L.; Johnson, R.L.; Skalski, J.R.; et al. Quantifying Mortal Injury of Juvenile Chinook Salmon Exposed to Simulated Hydro-Turbine Passage. Trans. Am. Fish. Soc. 2012, 141, 147–157. [Google Scholar] [CrossRef]
- Nyqvist, D.; Bergman, E.; Calles, O.; Greenberg, L. Intake Approach and Dam Passage by Downstream-migrating Atlantic Salmon Kelts. River Res. Appl. 2017, 33, 697–706. [Google Scholar] [CrossRef]
- Östergren, J.; Rivinoja, P. Overwintering and downstream migration of sea trout (Salmo trutta L.) kelts under regulated flows—Northern Sweden. River Res. Appl. 2008, 24, 551–563. [Google Scholar] [CrossRef]
- De Leaniz, C.G.; Fleming, I.A.; Einum, S.; Verspoor, E.; Jordan, W.C.; Consuegra, S.; Aubin-Horth, N.; Lajus, D.L.; Letcher, B.H.; Youngson, A.F.; et al. A critical review of adaptive genetic variation in Atlantic salmon: implications for conservation. Boil. Rev. 2007, 82, 173–211. [Google Scholar] [CrossRef]
- Calles, O.; Greenberg, L. Connectivity is a two-way street-the need for a holistic approach to fish passage problems in regulated rivers. River Res. Appl. 2009, 25, 1268–1286. [Google Scholar] [CrossRef]
- Martinez, J.; Deng, Z.D.; Titzler, P.; Duncan, J.; Lu, J.; Mueller, R.; Tian, C.; Trumbo, B.; Ahmann, M.; Renholds, J. Hydraulic and biological characterization of a large Kaplan turbine. Renew. Energy 2019, 131, 240–249. [Google Scholar] [CrossRef]
- Ices Baltic Salmon and Trout Assessment Working Group. Available online: https://www.ices.dk/sites/pub/Publication%20Reports/Expert%20Group%20Report/Fisheries%20Resources%20Steering%20Group/2019/WGBAST/wgbast_2019.pdf (accessed on 30 June 2020).
- Ferguson, J.W.; Absolon, R.F.; Carlson, T.J.; Sandford, B.P. Evidence of Delayed Mortality on Juvenile Pacific Salmon Passing through Turbines at Columbia River Dams. Trans. Am. Fish. Soc. 2006, 135, 139–150. [Google Scholar] [CrossRef]


{kind=link}
| Parameter | Stornorrfors Francis Turbine | Lanforsen Kaplan Turbine |
|---|---|---|
| No. of turbine blades | Unit I: 15 | Unit I–IV: 4 |
| Unit II–IV: 16 | ||
| Blade rotational speed (r∙min−1) | Unit I–III: 125 | Unit I–IV: 93.8 |
| Unit IV: 155 | ||
| Overall turbine blades and hub | Unit I–III: 4.33 | Unit I–IV: 4.98 |
| (runner) diameter (m) | Unit IV: 4.75 | |
| Hub diameter (m) | NA | Unit I–IV: 4.5 |
| Height of the wicket gates (m) | Unit I–III: 1.56 | NA |
| Unit IV: 1.73 | ||
| Turbine discharge (m3 s−1) | Unit I–III: 250 | Unit I–IV: 155 |
| Unit: IV: 275 |
| Period | Location | Parameter | Mean TL (cm) | Capture Method | Tagging Method | No. Tagged | No. Passed |
|---|---|---|---|---|---|---|---|
| 6–7 June 2017 | Umeälven | Adult | |||||
| Atlantic salmon | 70.9 ± 8.7 (54.0–92.0) | net/rod | surg. + PIT | 10 | 3 | ||
| Brown trout | 59.6 ± 2.6 (52.0–68.0) | net/rod | surg. + PIT | 10 | 4 | ||
| 8–15 June 2017 | Umeälven | Juvenile | |||||
| Atlantic salmon | 16.0 ± 0.3 (13.6–18.6) | smolt trap | surg. + PIT | 50 | 43 | ||
| 19 July 2018–12 September 2018 | Umeälven | Adult | |||||
| Atlantic salmon | 62.0 ± 1.3 (54.0–69.0) | net/fish ladder | surg./gast. | 31 | 5 | ||
| 13 June 2018–21 June 2018 | Umeälven | Juvenile | |||||
| Atlantic salmon | 15.9 ± 0.4 (12.3–18.5) | smolt trap | surg. + PIT | 40 | 16 | ||
| 20 June2019–10 September 2019 | Umeälven | Adult | |||||
| Atlantic salmon | 61.2 ± 1.6 (52.0–94.0) | net/fish ladder | surg./gast. + PIT | 56 | 13 | ||
| 2 May 2019–20 May 2019 | Dalälven | Juvenile | |||||
| Atlantic salmon | 16.3 ± 0.4 (13.0–20.0) | electrofishing | surg. + PIT | 80 | 61 | ||
| Brown trout | 24.6 ± 0.7 (21.0–27.0) | electrofishing | surgically + PIT | 17 | 7 |
| Tag and Receiver | Specification | Umeälven 2017 | Umeälven 2018 | Umeälven 2019 | Dalälven 2019 |
|---|---|---|---|---|---|
| PIT | BIOMARK HPT12, 12.5 MM, 134.2 kHz ISO FDXB | 70 | 40 | 44 | 97 |
| V16 Acoustic transmitter | V16-4L-069k-1, 69 kHz | 20 | 31 | ||
| V9 Acoustic transmitter | V9-2L, 180 kHz | 20 | 57 | ||
| V5 Acoustic transmitter | V5-1H, 180 kHz | 50 | 20 | 97 | |
| V7TP Acoustic transmitter | V7TP-180 kHz | 20 | |||
| VR2W Acoustic receiver | Frequency 69 kHz and 180 kHz | 27 | 48 | 34 | 24 |
| VR2AR Acoustic receiver | Acoustic release. Receive transmissions at 69 kHz | 15 | |||
| HR Acoustic receiver | High Residence. Receive transmissions at 180 kHz | 24 | 23 | 19 |
| Group | Location | Turbine Type | Parameter | Mean TL (cm) | No. |
|---|---|---|---|---|---|
| 1 | Umeälven | Francis | Adult | ||
| Atlantic salmon | 62.1 ± 2.8 (55.0–83.0) | 21 | |||
| Brown trout | 58.0 ± 4.5 (52.0–63.0) | 4 | |||
| 2 | Umeälven | Francis | Adult | ||
| Atlantic salmon | 61.9 ± 1.8 (52.0–94.0) | 54 | |||
| Brown trout | 58.0 ± 4.5 (52.0–63.0) | 4 | |||
| 3 | Umeälven | Francis | Juvenile | ||
| Atlantic salmon | 15.9 ± 0.3 (13.5–18.6) | 59 | |||
| Brown trout | - | 0 | |||
| 4 | Dalälven | Kaplan | Juvenile | ||
| Atlantic salmon | 16.4 ± 0.4 (13.0–19.5) | 61 | |||
| Brown trout | 25.1 ± 1.0 (23.0–27.0) | 7 |
| Group | Turbine Type | Mean Total Length (cm) | Mortality (%) obs. | Mortality (%) Model | Abs. Bias |
|---|---|---|---|---|---|
| 1 Adult Francis | F × 4 | 61 ± 2.5 (52–83) | 56 ± 20.5 (36–76) | 33.2 (4–76) | −0.23 |
| 2 Adult Francis | F × 4 | 61.7 ± 1.8 (52–94) | 81 ± 10.3 (71–91) | 33.2 (12–59) | −0.48 |
| 3 Juvenile Francis | F × 4 | 15.9 ± 3.1 (13.5–18.6) | 11.9 ± 8.4 (3–20) | 8.5 (0–25) | −0.03 |
| 4 Juvenile Kaplan | K × 4 | 17.3 ± 7.4 (13–27) | 0 ± 0 (0–0) | 1.3 (0–7) | 0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vikström, L.; Leonardsson, K.; Leander, J.; Shry, S.; Calles, O.; Hellström, G. Validation of Francis–Kaplan Turbine Blade Strike Models for Adult and Juvenile Atlantic Salmon (Salmo Salar, L.) and Anadromous Brown Trout (Salmo Trutta, L.) Passing High Head Turbines. Sustainability 2020, 12, 6384. https://doi.org/10.3390/su12166384
Vikström L, Leonardsson K, Leander J, Shry S, Calles O, Hellström G. Validation of Francis–Kaplan Turbine Blade Strike Models for Adult and Juvenile Atlantic Salmon (Salmo Salar, L.) and Anadromous Brown Trout (Salmo Trutta, L.) Passing High Head Turbines. Sustainability. 2020; 12(16):6384. https://doi.org/10.3390/su12166384
Chicago/Turabian StyleVikström, Linda, Kjell Leonardsson, Johan Leander, Samuel Shry, Olle Calles, and Gustav Hellström. 2020. "Validation of Francis–Kaplan Turbine Blade Strike Models for Adult and Juvenile Atlantic Salmon (Salmo Salar, L.) and Anadromous Brown Trout (Salmo Trutta, L.) Passing High Head Turbines" Sustainability 12, no. 16: 6384. https://doi.org/10.3390/su12166384
APA StyleVikström, L., Leonardsson, K., Leander, J., Shry, S., Calles, O., & Hellström, G. (2020). Validation of Francis–Kaplan Turbine Blade Strike Models for Adult and Juvenile Atlantic Salmon (Salmo Salar, L.) and Anadromous Brown Trout (Salmo Trutta, L.) Passing High Head Turbines. Sustainability, 12(16), 6384. https://doi.org/10.3390/su12166384