Sustainable Valorization of Halophytes from the Mediterranean Area: A Comprehensive Evaluation of Their Fatty Acid Profile and Implications for Human and Animal Nutrition

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Procedure and Processing of Biomass
2.2. Extraction and Fatty Acid Methyl Esters (FAME) Preparation
2.3. Determination of FAME Profile by GC-MS
2.4. Statistical Analysis
3. Results and Discussion
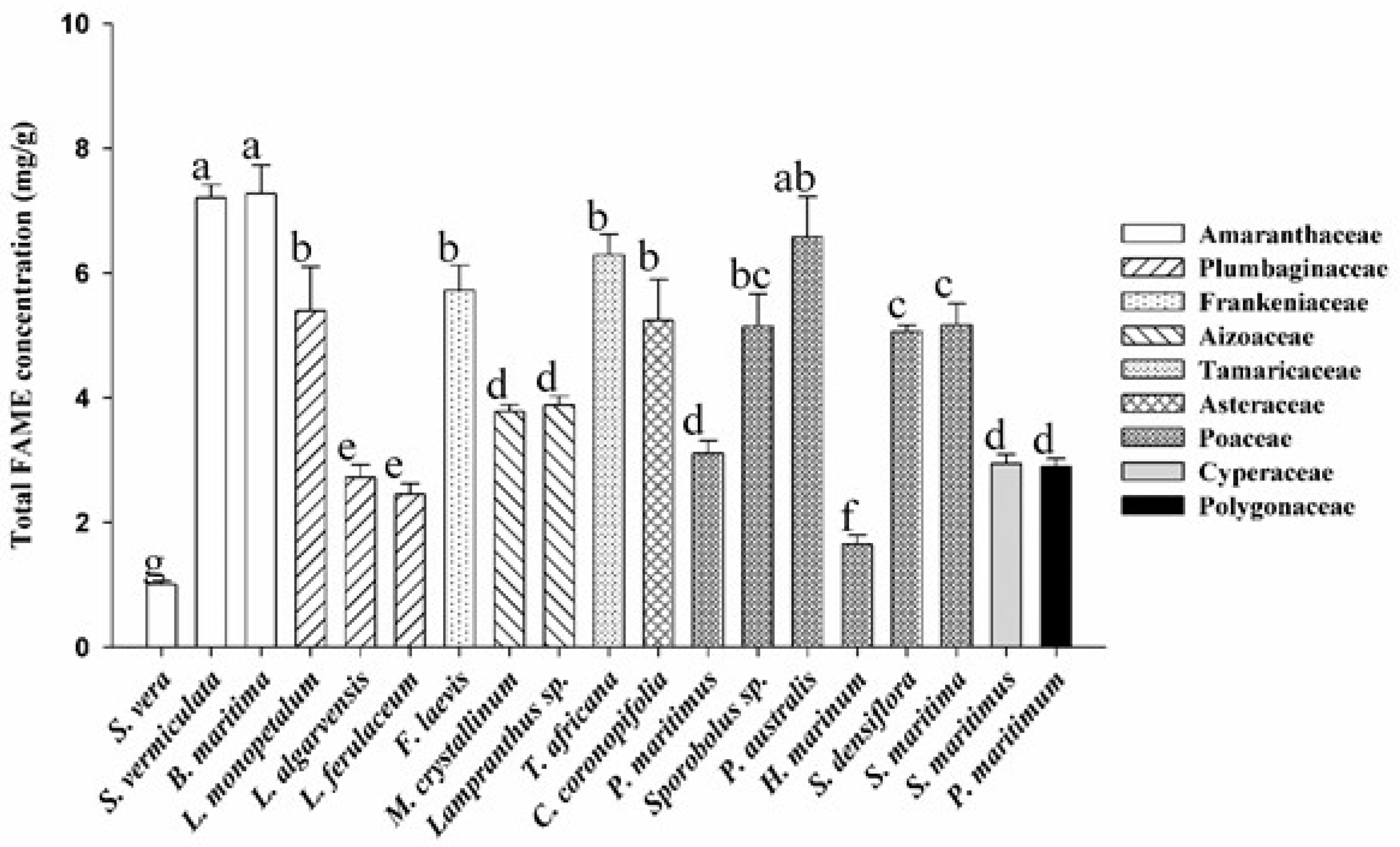
3.1. Total FAME Concentration
3.2. FAME Profiles
3.3. Less Common FAME
3.4. Potential Implications for Human and Animal Nutrition
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The threat of soil salinity: A European scale review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef]
- FAO. Available online: http://www.grida.no/resources/5906 (accessed on 7 September 2018).
- Branco, J.; Oliveria, M.; Ferreira, R.; Povoa, O. Desertification in Portugal: Causes, consequences and possible solutions. In Atas do I Congresso de Estudos Rurais do Norte Alentejano; Instituto Politécnico de Portalegre: Portalegre, Portugal, 2012. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Panta, S.; Flowers, T.; Lane, P.; Doyle, R.; Haros, G.; Shabala, S. Halophyte agriculture: Success stories. Environ. Exp. Bot. 2014, 107, 71–83. [Google Scholar] [CrossRef]
- Ventura, Y.; Eshel, A.; Pasternak, D.; Sagi, M. The development of halophyte-based agriculture: Past and present. Ann. Bot. 2015, 115, 529–540. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Debez, A.; Falleh, H.; Grignon, C.; Abdelly, C. Salinity effects on polyphenol content and antioxidant activities in leaves of the halophyte Cakile maritima. Plant Physiol. Biochem. 2007, 45, 244–249. [Google Scholar] [CrossRef]
- Cheeseman, J.M. The evolution of halophytes, glycophytes and crops, and its implications for food security under saline conditions. New Phytol. 2015, 206, 557–570. [Google Scholar] [CrossRef]
- Barreira, L.; Resek, E.; Rodrigues, M.J.; Rocha, M.I.; Pereira, H.; Bandarra, N.; da Silva, M.M.; Varela, J.; Custodio, L. Halophytes: Gourmet food with nutritional health benefits? J. Food Compos. Anal. 2017, 59, 35–42. [Google Scholar] [CrossRef]
- El Shaer, H.M. Halophytes and salt-tolerant plants as potential forage for ruminants in the Near East region. Small Rumin. Res. 2010, 91, 3–12. [Google Scholar] [CrossRef]
- Zárate, R.; El Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.A.; Rodríguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6, 25. [Google Scholar] [CrossRef]
- Abedi, E.; Sahari, M.A. Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sci. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef]
- Conway, M.C.; Mulhern, M.S.; McSorley, E.M.; van Wijngaarden, E.; Strain, J.J.; Myers, G.J.; Davidson, P.W.; Shamlaye, C.F.; Yeates, A.J. Dietary determinants of polyunsaturated fatty acid (PUFA) status in a high fish-eating cohort during pregnancy. Nutrients 2018, 10, 927. [Google Scholar] [CrossRef]
- Van Tran, L.; Malla, B.A.; Kumar, S.; Tyagi, A.K. Polyunsaturated fatty acids in male ruminant reproduction—A Review. Asian Australas. J. Anim. Sci. 2017, 30, 622–637. [Google Scholar] [CrossRef]
- Castro, L.F.C.; Monroig, O.; Leaver, M.J.; Wilson, J.; Cunha, I.; Tocher, D.R. Functional desaturase Fads1 (Delta 5) and Fads2 (Delta 6) orthologues evolved before the origin of jawed vertebrates. PLoS ONE 2012, 7, e31950. [Google Scholar] [CrossRef]
- Wu, D.; Lewis, E.D.; Pae, M.; Meydani, S.N. Nutritional modulation of immune function: analysis of evidence, mechanisms, and clinical relevance. Front. Immunol. 2018, 9, 3160. [Google Scholar] [CrossRef]
- Ben Hamed, K.; Ben Youssef, N.; Ranieri, A.; Zarrouk, M.; Abdelly, C. Changes in content and fatty acid profiles of total lipids and sulfolipids in the halophyte Crithmum maritimum under salt stress. J. Plant Physiol. 2005, 162, 599–602. [Google Scholar] [CrossRef]
- Nouairi, I.; Ghnaya, T.; Ben Youssef, N.; Zarrouk, M.; Habib Ghorbel, M. Changes in content and fatty acid profiles of total lipids of two halophytes: Sesuvium portulacastrum and Mesembryanthemum crystallinum under cadmium stress. J. Plant Physiol. 2006, 163, 1198–1202. [Google Scholar] [CrossRef]
- Custódio, L.; Ferreira Ana, C.; Pereira, H.; Silvestre, L.; Vizetto-Duarte, C.; Barreira, L.; Rauter Amélia, P.; Alberício, F.; Varela, J. The marine halophytes Carpobrotus edulis L. and Arthrocnemum macrostachyum L. are potential sources of nutritionally important PUFAs and metabolites with antioxidant, metal chelating and anticholinesterase inhibitory activities. Bot. Mar. 2012, 55, 281–288. [Google Scholar] [CrossRef]
- Tsydendambaev, V.D.; Ivanova, T.V.; Khalilova, L.A.; Kurkova, E.B.; Myasoedov, N.A.; Balnokin, Y.V. Fatty acid composition of lipids in vegetative organs of the halophyte Suaeda altissima under different levels of salinity. Russ. J. Plant Physiol. 2013, 60, 661–671. [Google Scholar] [CrossRef]
- Duarte, B.; Carreiras, J.; Pérez-Romero, J.A.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Matos, A.R.; Marques, J.C.; Caçador, I. Halophyte fatty acids as biomarkers of anthropogenic-driven contamination in Mediterranean marshes: Sentinel species survey and development of an integrated biomarker response (IBR) index. Ecol. Indic. 2018, 87, 86–96. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved recovery of fatty-acid through direct trans-esterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar]
- Vizetto-Duarte, C.; Custodio, L.; Acosta, G.; Lago, J.H.G.; Morais, T.R.; de Sousa, C.B.; Gangadhar, K.N.; Rodrigues, M.J.; Pereira, H.; Lima, R.T.; et al. Can macroalgae provide promising anti-tumoral compounds? A closer look at Cystoseira tamariscifolia as a source for antioxidant and anti-hepatocarcinoma compounds. PeerJ 2016, 4, e1704. [Google Scholar] [CrossRef]
- Meziane, T.; Bodineau, L.; Retiere, C.; Thoumelin, G. The use of lipid markers to define sources of organic matter in sediment and food web of the intertidal salt-marsh-flat ecosystem of Mont-Saint-Michel Bay, France. J. Sea Res. 1997, 38, 47–58. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Muylaert, K.; Foubert, I. Optimization of an analytical procedure for extraction of lipids from microalgae. J. Am. Oil Chem. Soc. 2012, 89, 189–198. [Google Scholar] [CrossRef]
- Arayana, G.L.; Rao, K.S.; Pantulu, A.J.; Thyagarajan, G. Composition of lipids in roots, stalks, leaves and flowers of Eichhornia crassipes (Mart.) Solms. Aquat. Bot. 1984, 20, 219–227. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Rajion, M.A.; Goh, Y.M. Effects of oils rich in linoleic and α-linolenic acids on fatty acid profile and gene expression in goat meat. Nutrients 2014, 6, 3913–3928. [Google Scholar] [CrossRef]
- Ren, J.; Chung, S.H. Anti-inflammatory effect of alpha-linolenic acid and its mode of action through the inhibition of nitric oxide production and inducible nitric oxide synthase gene expression via NF-kappaB and mitogen-activated protein kinase pathways. J. Agric. Food Chem. 2007, 55, 5073–5080. [Google Scholar] [CrossRef]
- Ren, J.; Han, E.J.; Chung, S.H. In vivo and in vitro anti-inflammatory activities of alpha-linolenic acid isolated from Actinidia polygama fruits. Arch. Pharmacal Res. 2007, 30, 708–714. [Google Scholar] [CrossRef]
- Connor, W.E. Importance of n-3 fatty acids in health and disease. Am. J. Clin. Nutr. 2000, 71, 171s–175s. [Google Scholar] [CrossRef]
- Lauritzen, I.; Blondeau, N.; Heurteaux, C.; Widmann, C.; Romey, G.; Lazdunski, M. Polyunsaturated fatty acids are potent neuroprotectors. EMBO J. 2000, 19, 1784–1793. [Google Scholar] [CrossRef]
- Walters, D.; Raynor, L.; Mitchell, A.; Walker, R.; Walker, K. Antifungal activities of four fatty acids against plant pathogenic fungi. Mycopathologia 2004, 157, 87–90. [Google Scholar]
- Pariza, M.W.; Park, Y.; Cook, M.E. Mechanisms of action of conjugated linoleic acid: Evidence and speculation. Proc. Soc. Exp. Biol. Med. 2000, 223, 8–13. [Google Scholar] [CrossRef]
- Kether, F.B.H.; Mahjoub, M.A.; Mahjoub, S.A.; Salah, K.B.; Helal, A.N.; Mighri, Z. Chemical composition, in vitro antifungal and antioxidant activities of essential oil from Cotula coronopifolia L. growing in Tunisia. Afr. J. Microbiol. Res. 2012, 6, 4388–4395. [Google Scholar]
- Chen, S.; Ju, M.L.; Luo, Y.; Chen, Z.J.; Zhao, C.P.; Zhou, Y.; Fu, J. Hepatoprotective and antioxidant activities of the aqueous extract from the rhizome of Phragmites australis. Z. Nat. C 2013, 68, 439–444. [Google Scholar]
- Sellal, A.; Belattar, R.; Bouzidi, A. Heavy metals chelating ability and antioxidant activity of Phragmites australis stems extracts. J. Ecol. Eng. 2019, 20, 116–123. [Google Scholar]
- Mohammed, H.A.; ESBelkhaer, A.; Abdellatif, A.G. Phytochemical and antioxidant screening of Suaeda vera L. growing in Libya. Int. J. Pharm. Res. Sch. 2014, 3, 891–896. [Google Scholar]
- Uncini Manganelli, R.E.; Camangi, F.; Tomei, P.E. Curing animals with plants: traditional usage in Tuscany (Italy). J. Ethnopharmacol. 2001, 78, 171–191. [Google Scholar] [CrossRef]
- Viegi, L.; Pieroni, A.; Guarrera, P.M.; Vangelisti, R. A review of plants used in folk veterinary medicine in Italy as basis for a databank. J. Ethnopharmacol. 2003, 89, 221–244. [Google Scholar] [CrossRef]
- Menale, B.; Muoio, R. Use of medicinal plants in the South-Eastern area of the Partenio Regional Park (Campania, Southern Italy). J. Ethnopharmacol. 2014, 153, 297–307. [Google Scholar] [CrossRef]
- Mongrand, S.; Bessoule, J.J.; Cabantous, F.; Cassagne, C. The C16:3\C18:3 fatty acid balance in photosynthetic tissues from 468 plant species. Phytochemistry 1998, 49, 1049–1064. [Google Scholar] [CrossRef]
- Ivanov, D.S.; Levic´, J.D.; Sredanovic´, S.A. Fatty acid composition of various soybean products. Food Feed Res. 2010, 37, 65–70. [Google Scholar]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Scollan, N.; Hocquette, J.F.; Nuernberg, K.; Dannenberger, D.; Richardson, I.; Moloney, A. Innovations in beef production systems that enhance the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 2006, 74, 17–33. [Google Scholar] [CrossRef]
- Bourre, J.M. Where to find omega-3 fatty acids and how feeding animals with diet enriched in omega-3 fatty acids to increase nutritional value of derived products for human: What is actually useful? J. Nutr. Health Aging 2005, 9, 232–242. [Google Scholar]
- Abd El-Hack, M.E.; Samak, D.H.; Noreldin, A.E.; Arif, M.; Yaqoob, H.S.; Swelum, A.A. Towards saving freshwater: Halophytes as unconventional feedstuffs in livestock feed: A review. Environ. Sci. Pollut. Res. 2018, 25, 14397–14406. [Google Scholar] [CrossRef]
- Inácio, M.; Pereira, V.; Pinto, M. The Soil Geochemical Atlas of Portugal: Overview and applications. J. Geochem. Explor. 2008, 98, 22–33. [Google Scholar] [CrossRef]


{kind=link}
| Amaranthaceae | Plumbaginaceae | ||||||
|---|---|---|---|---|---|---|---|
| FA (%) | Common Name | S. vera | S. vermiculata | B. maritima | L. monopetalum | L. algarvense | L. ferulaceum |
| C12:0 | Lauric acid | * | 0.2 ± 0.0 | * | 1.4 ± 0.2 | 0.6 ± 0.0 | 0.8 ± 0.0 |
| C14:0 | Myristic acid | 1.9 ± 0.0 | 1.0 ± 0.1 | 0.6 ± 0.0 | 0.7 ± 0.1 | 1.8 ± 0.1 | 3.3 ± 0.1 |
| C15:0 | Pentadecanoic acid | * | 0.3 ± 0.0 | 0.3 ± 0.0 | * | * | 0.7 ± 0.0 |
| C16:0 | Palmitic acid | 27.4 ± 0.1 | 20.9 ± 0.8 | 25.4 ± 0.3 | 19.4± 2.3 | 33.0 ± 0.2 | 35.0 ± 1.0 |
| C17:0 | Margaric acid | * | 0.5 ± 0.0 | 0.3 ± 0.0 | 0.5 ± 0.0 | * | 0.6 ± 0.0 |
| C18:0 | Stearic acid | 3.5 ± 0.0 | 3.3 ± 0.1 | 2.7 ± 0.0 | 11.6 ± 9.7 | 4.3 ± 0.1 | 3.7 ± 0.1 |
| C20:0 | Arachidic acid | 3.5 ± 0.1 | 3.4 ± 0.1 | 1.1 ± 1.0 | 23.3 ± 3.0 | 3.2 ± 0.1 | 5.5 ± 0.2 |
| C21:0 | Henicosanoic acid | * | 0.4 ± 0.1 | * | 1.2 ± 0.2 | * | * |
| C22:0 | Behenic acid | 6.0 ± 0.1 | 5.0 ± 0.1 | 2.5 ± 0.1 | 16.4 ± 1.9 | 2.8 ± 0.0 | 8.3 ± 0.6 |
| C23:0 | Tricosylic acid | * | 1.6 ± 0.0 | 0.5 ± 0.0 | 1.7 ± 0.2 | * | * |
| C24:0 | Lignoceric acid | * | 2.6 ± 0.1 | 1.6 ± 0.3 | 4.7 ± 0.4 | 2.6 ± 0.3 | 4.8 ± 0.4 |
| C26:0 | Cerotic acid | * | 1.2 ± 0.1 | * | * | * | * |
| ∑ SFA | 42.2 ± 0.2 | 40.6 ± 0.9 | 34.9 ± 1.1 | 81.0 ± 10.6 | 48.3 ± 0.4 | 62.9 ± 1.2 | |
| C16:1 | Palmitoleic acid | * | 2.0 ± 0.0 | 0.4 ± 0.3 | * | 0.5 ± 0.4 | 1.7 ± 0.1 |
| C18:1 | Oleic acid | * | 9.8 ± 0.3 | 12.6 ± 0.1 | 3.8 ± 0.4 | 8.8 ± 0.1 | 7.3 ± 0.2 |
| C20:1 | Eicosenoic acid | * | * | 0.4 ± 0.01 | * | * | * |
| ∑ MUFA | * | 11.8 ± 0.2 | 13.4 ± 0.3 | 3.8 ± 0.4 | 9.4 ± 0.4 | 9.0 ± 0.2 | |
| C18:2(n-6) | Linoleic acid | 22.0 ± 0.7 | 47.6 ± 1.0 | 51.4 ± 0.8 | 15.2 ± 1.6 | 42.3 ± 0.2 | 28.2 ± 0.3 |
| C16:3(n-3) | Hexadecatrienoic acid | * | * | 0.2 ± 0.0 | * | * | * |
| C18:3(n-3) | α-linolenic acid | 35.8 ± 0.6 | * | * | * | * | * |
| ∑ PUFA | 57.8 ± 1.0 | 47.6 ± 1.0 | 51.7 ± 0.8 | 15.2 ± 1.6 | 42.3 ± 0.2 | 28.2 ± 0.3 | |
| ∑ PUFA/∑ SFA | 1.4 | 1.2 | 1.5 | 1.9 | 0.9 | 0.4 | |
| n-6/n-3 | 0.6 | - | - | - | - | - |
| Frankeniaceae | Aizoaceae | Tamaricaceae | Asteraceae | |||
|---|---|---|---|---|---|---|
| FA (%) | Common Name | F. laevis | M. crystallinum | Lampranthus sp. | T. africana | C. coronopifolia |
| C12:0 | Lauric acid | 2.7 ±. 0.1 | 0.3 ± 0.0 | 1.4 ± 0.1 | 1.3 ± 0.1 | 0.2 ± 0.1 |
| C14:0 | Myristic acid | 3.7 ± 0.1 | 2.3 ± 0.1 | 3.4 ± 0.1 | 4.0 ± 0.2 | 1.0 ± 0.1 |
| C15:0 | Pentadecanoic acid | 0.2 ± 0.2 | * | 0.5 ± 0.3 | 0.3 ± 0.0 | 0.3 ± 0.0 |
| C16:0 | Palmitic acid | 18.7 ± 0.4 | 26.1 ± 0.4 | 14.3 ± 0.5 | 25.6 ± 0.6 | 17.5 ± 0.6 |
| C17:0 | Margaric acid | 0.5 ± 0.0 | 0.5 ± 0.0 | 0.5 ± 0.3 | 0.4 ± 0.0 | 0.2 ± 0.2 |
| C18:0 | Stearic acid | 2.5 ± 0.1 | 3.3 ± 0.1 | 3.0 ± 0.1 | 3.9 ± 0.0 | 1.6 ± 0.1 |
| C20:0 | Arachidic acid | 5.2 ± 0.6 | 10.5 ± 0.1 | 18.9 ± 0.9 | 3.6 ± 0.2 | 0.6 ± 0.5 |
| C21:0 | Henicosanoic acid | * | 0.8 ± 0.0 | 1.2 ± 0.1 | * | * |
| C22:0 | Behenic acid | 9.8 ± 0.5 | 12.0 ± 0.1 | 18.9 ± 0.9 | 6.1 ± 0.5 | 1.4 ± 0.1 |
| C23:0 | Tricosylic acid | * | 1.6 ± 0.0 | * | 2.1 ±1.0 | * |
| C24:0 | Lignoceric acid | 17.6 ± 1.1 | 5.9 ± 1.2 | 6.2 ± 1.1 | 4.1 ±2.0 | 1.9 ± 0.1 |
| C26:0 | Cerotic acid | 2.5 ± 0.1 | * | * | * | * |
| ∑ SFA | 70.1 ± 1.5 | 63.5 ± 1.3 | 68.3 ± 1.3 | 51.6 ± 2.4 | 24.8 ± 0.8 | |
| C16:1 | Palmitoleic acid | 1.6 ± 0.5 | 0.9 ± 0.0 | * | 0.9 ± 0.8 | 0.6 ± 0.0 |
| C18:1 | Oleic acid | 8.8 ± 0.3 | 13.8 ± 0.1 | 6.9 ± 0.2 | 11.1 ± 0.2 | * |
| C20:1 | Eicosenoic acid | 0.3 ± 0.0 | * | * | 0.3 ± 0.0 | * |
| ∑ MUFA | 10.8 ± 0.6 | 14.7 ± 0.1 | 6.9 ± 0.2 | 12.3 ± 0.8 | 0.6 ± 0.0 | |
| C18:2(n-6) | Linoleic acid | 19.1 ± 1.0 | 19.6 ± 0.3 | 24.8 ± 1.0 | 36.2 ± 0.3 | 34.7 ± 1.0 |
| C16:3(n-3) | Hexadecatrienoic acid | * | 2.2 ± 0.1 | * | * | * |
| C18:3(n-3) | α-linolenic acid | * | * | * | * | 39.9 ± 0.2 |
| ∑ PUFA | 19.1 ± 1.0 | 21.8 ± 0.3 | 24.8 ± 1.0 | 36.2 ± 0.3 | 74.6 ± 1.0 | |
| ∑ PUFA/∑ SFA | 0.3 | 0.3 | 0.4 | 0.7 | 3.0 | |
| n-6/n-3 | - | - | - | - | 0.9 |
| Poaceae | Cyperaceae | Polygonaceae | |||||||
|---|---|---|---|---|---|---|---|---|---|
| FA (%) | Common name | P. maritimus | Sporobolus sp. | P. australis | H. marinum | S. densiflora | S. maritima | S. maritimus | P. maritimum |
| C12:0 | Lauric acid | 0.8 ± 0.0 | 1.2 ± 0.0 | 0.8 ± 0.0 | 0.5 ± 0.0 | 0.7 ± 0.0 | 0.7 ± 0.0 | 3.1 ± 0.1 | 0.7 ± 0.0 |
| C14:0 | Myristic acid | 2.0 ± 0.1 | 1.8 ± 0.1 | 2.0 ± 0.1 | 1.6 ± 0.1 | 1.5 ± 0.0 | 1.5 ± 0.1 | 5.4 ± 0.2 | 4.2 ± 0.2 |
| C15:0 | Pentadecanoic acid | 1.0 ± 0.0 | 0.4 ± 0.0 | 0.3 ± 0.0 | *. | 0.5 ± 0.0 | 0.5 ± 0.0 | 1.1 ± 0.0 | 0.5 ± 0.0 |
| C16:0 | Palmitic acid | 36.4 ± 1.1 | 19.7 ± 1.0 | 20.7 ± 0.4 | 21.6 ± 0.3 | 17.9 ± 0.0 | 17.6 ± 0.6 | 40.1 ± 0.9 | 27.8 ± 0.1 |
| C17:0 | Margaric acid | 0.9 ± 0.0 | * | 0.5 ± 0.0 | * | 0.5 ± 0.0 | 0.5 ± 0.0 | 1.1 ± 0.0 | 0.4 ± 0.3 |
| C18:0 | Stearic acid | 4.6 ± 0.2 | 2.5 ± 0.3 | 0.8 ± 1.3 | 1.8 ± 0.1 | 2.4 ± 0.0 | 2.2 ± 0.1 | 4.4 ± 0.1 | 3.3 ± 0.0 |
| C20:0 | Arachidic acid | 2.4 ± 0.0 | 4.2 ± 0.4 | 2.9 ± 0.1 | 2.4 ± 0.1 | 6.7 ± 0.1 | 7.0 ± 0.3 | 3.5 ± 0.1 | 7.5 ± 0.2 |
| C21:0 | Henicosanoic acid | 0.4 ± 0.3 | *. | 0.5 ± 0.0 | * | * | * | * | 0.7 ± 0.0 |
| C22:0 | Behenic acid | 2.1 ± 0.4 | 3.8 ± 0.2 | 2.3 ± 0.0 | 1.8 ± 0.2 | 2.6 ± 0.4 | 2.6 ± 0.1 | 3.4 ± 0.2 | 10.5 ± 0.1 |
| C23:0 | Tricosylic acid | * | * | 0.7 ± 0.0 | * | 2.0 ± 0.6 | 1.1 ± 0.7 | 1.7 ± 0.1 | 1.9 ± 0.3 |
| C24:0 | Lignoceric acid | 3.5 ± 0.4 | 3.3 ± 0.4 | 2.5 ± 0.4 | * | 3.9 ± 0.0 | 4.4 ± 0.7 | 5.9 ± 0.4 | 6.6 ± 0.3 |
| C26:0 | Cerotic acid | * | * | *. | * | 1.5 ± 0.0 | 0.8 ± 0.9 | 2.5 ± 0.2 | * |
| ∑ SFA | 54.2 ± 2.7 | 36.9 ± 1.2 | 34.0 ± 1.5 | 29.7 ± 0.4 | 40.2 ± 0.7 | 38.9 ± 1.5 | 72.3 ± 1.0 | 64.2 ± 0.6 | |
| C16:1 | Palmitoleic acid | 2.4 ± 0.2 | * | 1.5 ± 0.1 | 1.1 ± 0.0 | 1.5 ± 0.0 | 1.6 ± 0.2 | * | 2.1 ± 0.0 |
| C18:1 | Oleic acid | 10.7 ± 0.3 | 0.8 ± 0.0 | * | 0.8 ± 0.0 | 14.7 ± 0.2 | 14.0 ± 0.7 | 6.8 ± 0.2 | 9.8 ± 0.0 |
| C20:1 | Eicosenoic acid | * | * | 0.1 ± 0.1 | * | 0.7 ± 0.0 | 0.9 ± 0.1 | * | * |
| ∑ MUFA | 13.1 ± 0.3 | 0.7 ± 0.0 | 1.6 ± 0.1 | 1.9 ± 0.0 | 16.9 ± 0.1 | 16.5 ± 0.6 | 6.8 ± 0.2 | 11.8 ± 0.0 | |
| C18:2(n-6) | Linoleic acid | 32.7 ± 0.8 | 28.8 ± 0.6 | 26.5 ± 0.4 | 34.8 ± 0.7 | 42.4 ± 0.7 | 44.1 ± 0.6 | 20.9 ± 1.7 | 24.0 ± 0.1 |
| C20:2(n-6) | Eicosadienoic acid | * | * | * | * | 0.5 ± 0.0 | 0.5 ± 0.0 | * | * |
| C18:3(n-3) | α-linolenic acid | * | 33.5 ± 1.7 | 37.9 ±1.0 | 33.8 ± 0.3 | * | * | * | * |
| ∑ PUFA | 32.7 ± 0.8 | 62.3 ± 1.8 | 64.4 ± 1.1 | 68.4 ± 0.7 | 42.9 ± 0.7 | 44.6 ± 0.6 | 20.9 ± 1.8 | 24.0 ± 0.1 | |
| ∑ PUFA/∑ SFA | 0.6 | 1.7 | 1.9 | 2.3 | 1.1 | 1.1 | 0.3 | 0.4 | |
| n-6/n-3 | - | 0.9 | 0.7 | 1.0 | - | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizetto-Duarte, C.; Figueiredo, F.; Rodrigues, M.J.; Polo, C.; Rešek, E.; Custódio, L. Sustainable Valorization of Halophytes from the Mediterranean Area: A Comprehensive Evaluation of Their Fatty Acid Profile and Implications for Human and Animal Nutrition. Sustainability 2019, 11, 2197. https://doi.org/10.3390/su11082197
Vizetto-Duarte C, Figueiredo F, Rodrigues MJ, Polo C, Rešek E, Custódio L. Sustainable Valorization of Halophytes from the Mediterranean Area: A Comprehensive Evaluation of Their Fatty Acid Profile and Implications for Human and Animal Nutrition. Sustainability. 2019; 11(8):2197. https://doi.org/10.3390/su11082197
Chicago/Turabian StyleVizetto-Duarte, Catarina, Filipe Figueiredo, Maria João Rodrigues, Cristina Polo, Eva Rešek, and Luísa Custódio. 2019. "Sustainable Valorization of Halophytes from the Mediterranean Area: A Comprehensive Evaluation of Their Fatty Acid Profile and Implications for Human and Animal Nutrition" Sustainability 11, no. 8: 2197. https://doi.org/10.3390/su11082197
APA StyleVizetto-Duarte, C., Figueiredo, F., Rodrigues, M. J., Polo, C., Rešek, E., & Custódio, L. (2019). Sustainable Valorization of Halophytes from the Mediterranean Area: A Comprehensive Evaluation of Their Fatty Acid Profile and Implications for Human and Animal Nutrition. Sustainability, 11(8), 2197. https://doi.org/10.3390/su11082197