The Role of Different Earthworm Species (Metaphire Hilgendorfi and Eisenia Fetida) on CO2 Emissions and Microbial Biomass during Barley Decomposition



Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Characteristics and Earthworms
2.2. Experimental Design
2.3. Measurements of Soil Chemical Properties
2.4. Measurements of Soil Enzyme Activities
2.5. Measurements of Soil Respiration
2.6. Data Analysis
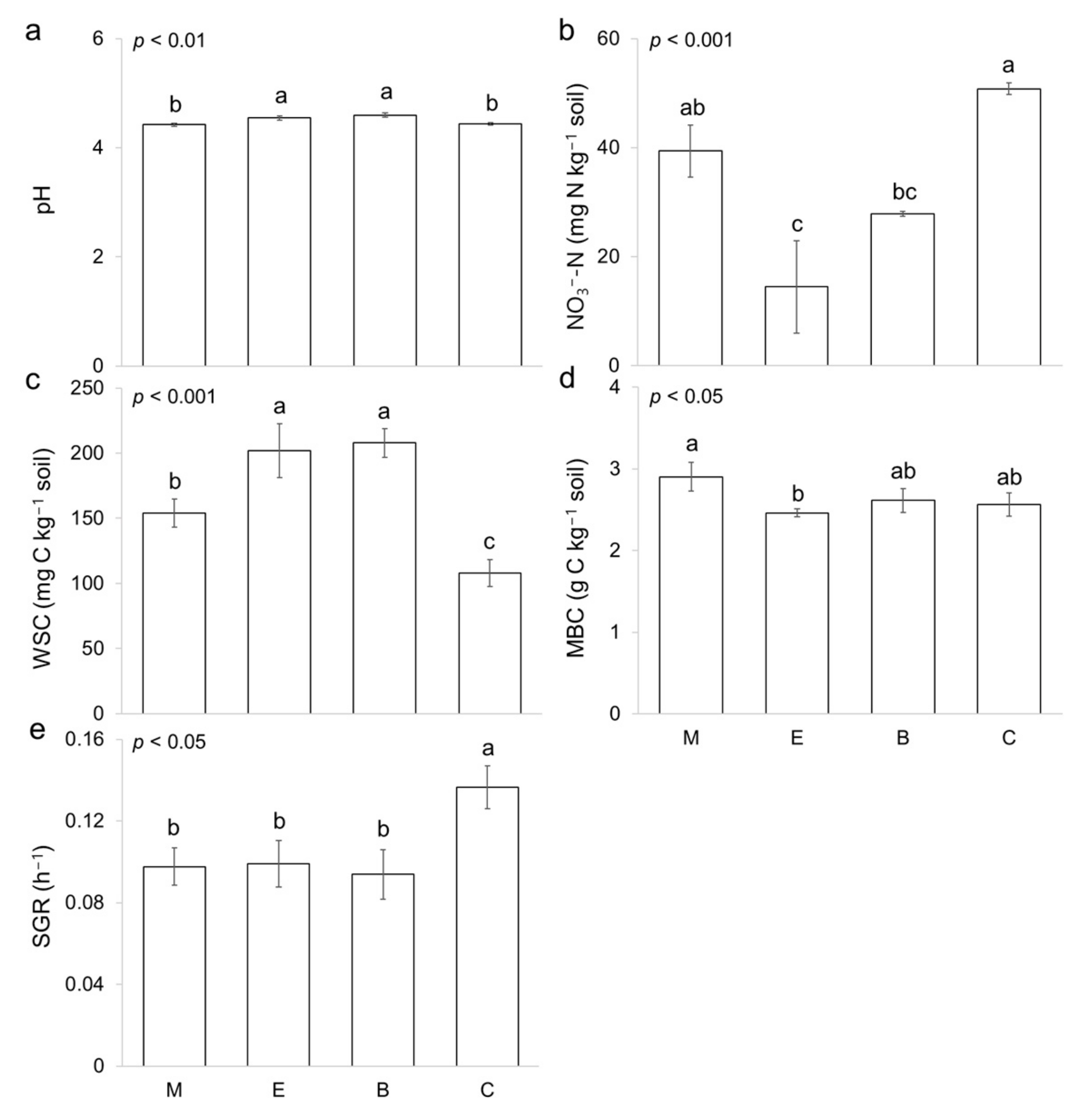
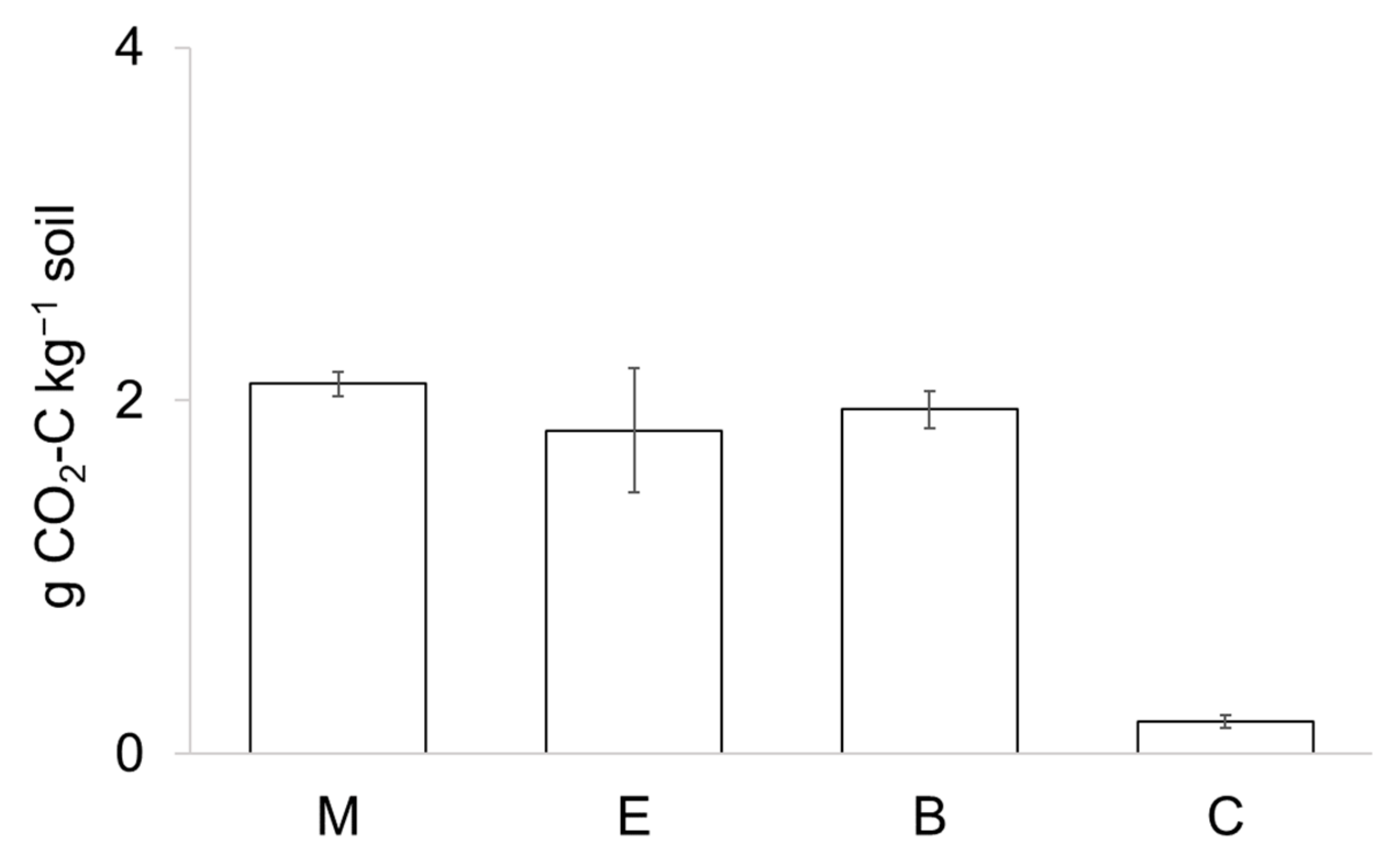
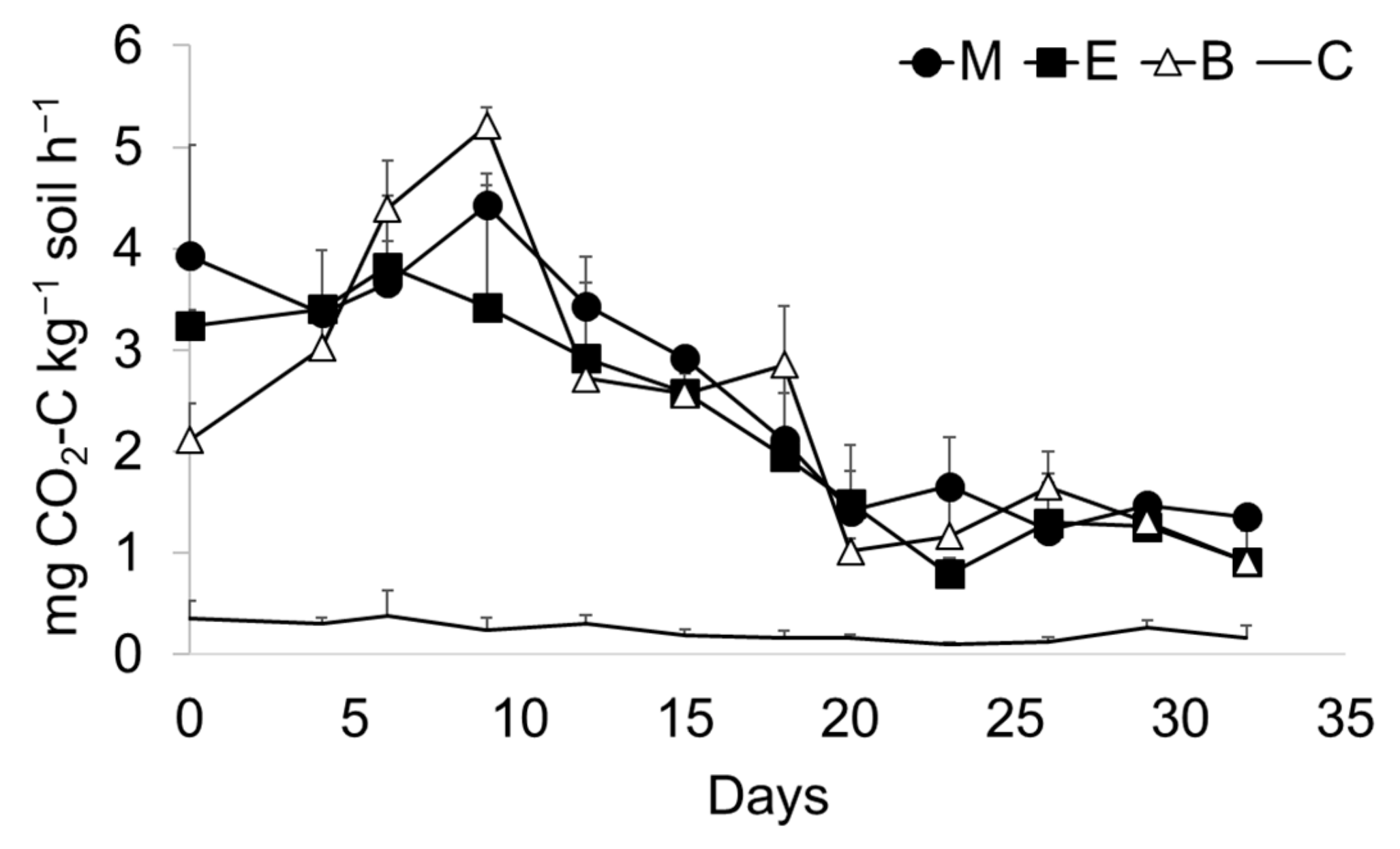
3. Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Luo, Z.; Wang, E.; Sun, O.J. Soil carbon change and its responses to agricultural practices in Australian agro-ecosystems: A review and synthesis. Geoderma 2010, 155, 211–223. [Google Scholar] [CrossRef]
- Li, X.G.; Jia, B.; Lv, J.; Ma, Q.; Kuzyakov, Y.; Li, F. Nitrogen fertilization decreases the decomposition of soil organic matter and plant residues in planted soils. Soil Biol. Biochem. 2017, 112, 47–55. [Google Scholar] [CrossRef]
- Fonte, S.J.; Six, J. Earthworms and litter management contributions to ecosystem services in a tropical agroforestry system. Ecol. Appl. 2010, 20, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Brandon, M.; Lores, M.; Dominguez, J. Species-specific effects of epigeic earthworms on microbial community structure during first stages of decomposition of organic matter. PLoS ONE 2012, 7, e31895. [Google Scholar] [CrossRef]
- Massey, P.A.; Creamer, R.E.; Schulte, R.P.O.; Whelan, M.J.; Ritz, K. The effects of earthworms, botanical diversity and fertiliser type on the vertical distribution of soil nutrients and plant nutrient acquisition. Biol. Fertil. Soils 2013, 49, 1189–1201. [Google Scholar] [CrossRef]
- Griffiths, B.; Faber, J.; Bloem, J. Applying soil health indicators to encourage sustainable soil use: The transition from scientific study to practical application. Sustainability 2018, 10, 3021. [Google Scholar] [CrossRef]
- Groffman, P.M.; Fahey, T.J.; Fisk, M.C.; Yavitt, J.B.; Sherman, R.E.; Bohlen, P.J.; Maerz, J.C. Earthworms increase soil microbial biomass carrying capacity and nitrogen retention in northern hardwood forests. Soil Biol. Biochem. 2015, 87 (Suppl. C), 51–58. [Google Scholar] [CrossRef]
- Aira, M.; Sampedro, L.; Monroy, F.; Dominguez, J. Detritivorous earthworms directly modify the structure, thus altering the functioning of a microdecomposer food web. Soil Biol. Biochem. 2008, 40, 2511–2516. [Google Scholar] [CrossRef]
- Aira, M.; Dominguez, J. Earthworm Effects without earthworms: Inoculation of raw organic matter with worm-worked substrates alters microbial community functioning. PLoS ONE 2011, 6, e16354. [Google Scholar] [CrossRef]
- Sampedro, L.; Dominguez, J. Stable isotope natural abundances (Δ13C and Δ15N) of the earthworm Eisenia fetida and other soil fauna living in two different vermicomposting environments. Appl. Soil Ecol. 2008, 38, 91–99. [Google Scholar] [CrossRef]
- Uchida, T.; Kaneko, N.; Ito, M.T.; Futagami, K.; Sasaki, T.; Sugimoto, A. Analysis of the feeding ecology of earthworms (Megascolecidae) in Japanese forests using gut content fractionation and Δ15N and Δ13C stable isotope natural abundances. Appl. Soil Ecol. 2004, 27, 153–163. [Google Scholar] [CrossRef]
- Chang, C.-H.; Szlavecz, K.; Filley, T.; Buyer, J.S.; Bernard, M.J.; Pitz, S.L. Belowground competition among invading detritivores. Ecology 2016, 97, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Enami, Y.; Okano, S.; Yada, H.; Nakamura, Y. Influence of earthworm activity and rice straw application on the soil microbial community structure analyzed by PLFA pattern. Eur. J. Soil Biol. 2001, 37, 269–272. [Google Scholar] [CrossRef]
- Caravaca, F.; Pera, A.; Masciandaro, G.; Ceccanti, B.; Roldan, A. A Microcosm approach to assessing the effects of earthworm inoculation and oat cover cropping on CO2 fluxes and biological properties in an amended semiarid soil. Chemosphere 2005, 59, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.-G.; Li, G.-T.; Shen, T.-S.; Wang, J.-K.; Sun, Z. Changes in microbial biomass C, N, and P and enzyme activities in soil incubated with the earthworms Metaphire guillelmi or Eisenia fetida. Soil Biol. Biochem. 2000, 32, 2055–2062. [Google Scholar] [CrossRef]
- Aira, M.; Monroy, F.; Domínguez, J. Eisenia fetida (Oligochaeta: Lumbricidae) modifies the structure and physiological capabilities of microbial communities improving carbon mineralization during vermicomposting of pig manure. Microb. Ecol. 2007, 54, 662–671. [Google Scholar] [CrossRef]
- Tao, J.; Griffiths, B.; Zhang, S.; Chen, X.; Liu, M.; Hu, F.; Li, H. Effects of earthworms on soil enzyme activity in an organic residue amended rice-wheat Rotation Agro-Ecosystem. Appl. Soil Ecol. 2009, 42, 221–226. [Google Scholar] [CrossRef]
- D’Haene, K.; Moreels, E.; De Neve, S.; Chaves Daguilar, B.; Boeckx, P.; Hofman, G.; van Cleemput, O. Soil properties influencing the denitrification potential of flemish agricultural soils. Biol. Fertil. Soils 2003, 38, 358–366. [Google Scholar] [CrossRef]
- Jin, T.; Shimizu, M.; Marutani, S.; Desyatkin, A.R.; Iizuka, N.; Hata, H.; Hatano, R. Effect of chemical fertilizer and manure application on N2O emission from reed canary grassland in Hokkaido, Japan. Soil Sci. Plant. Nutr. 2010, 56, 53–65. [Google Scholar] [CrossRef]
- Wu, F.; Wan, J.H.C.; Wu, S.; Wong, M. Effects of earthworms and plant growth-promoting rhizobacteria (PGPR) on availability of nitrogen, phosphorus, and potassium in soil. J. Plant Nutr. Soil Sci. 2012, 175, 423–433. [Google Scholar] [CrossRef]
- Hoang, D.T.T.; Razavi, B.S.; Kuzyakov, Y.; Blagodatskaya, E. Earthworm burrows: Kinetics and spatial distribution of enzymes of C-, N- and P- cycles. Soil Biol. Biochem. 2016, 99, 94–103. [Google Scholar] [CrossRef]
- Berry, E.C.; Jordan, D. Temperature and soil moisture content effects on the growth of Lumbricus terrestris (Oligochaeta: Lumbricidae) under laboratory conditions. Soil Biol. Biochem. 2001, 33, 133–136. [Google Scholar] [CrossRef]
- Van Gestel, M.; Merckx, R.; Vlassak, K. Microbial biomass responses to soil drying and rewetting: The fate of fast- and slow-growing microorganisms in soils from different climates. Soil Biol. Biochem. 1993, 25, 109–123. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Haney, R.L.; Honeycutt, C.W.; Schomberg, H.H.; Hons, F.M. Flush of carbon dioxide following rewetting of dried soil relates to active organic pools. Soil Sci. Soc. Am. J. 2000, 64, 613–623. [Google Scholar] [CrossRef]
- Datta, R.; Vranova, V.; Pavelka, M.; Rejšek, K.; Formanek, P. Effect of soil sieving on respiration induced by low-molecular-weight substrates. Int. Agrophysics 2014, 28, 119–124. [Google Scholar] [CrossRef]
- Vidal, A.; Quenea, K.; Alexis, M.; Nguyen Tu, T.T.; Mathieu, J.; Vaury, V.; Derenne, S. Fate of 13C labelled root and shoot residues in soil and anecic earthworm casts: A mesocosm experiment. Geoderma 2017, 285 (Suppl. C), 9–18. [Google Scholar] [CrossRef]
- Fründ, H.-C.; Butt, K.; Capowiez, Y.; Eisenhauer, N.; Emmerling, C.; Ernst, G.; Potthoff, M.; Schadler, M.; Schrader, S. Using earthworms as model organisms in the laboratory: Recommendations for experimental implementations. Pedobiologia 2010, 53, 119–125. [Google Scholar] [CrossRef]
- Hamamoto, T.; Uchida, Y. Sodium contents in dairy cow urine and soil aggregate sizes influence the amount of nitrogen lost from soil. Appl. Environ. Soil Sci. 2015, 275985. [Google Scholar] [CrossRef]
- Uchida, Y.; Nishimura, S.; Akiyama, H. The relationship of water-soluble carbon and hot-water-soluble carbon with soil respiration in agricultural fields. Agric. Ecosyst. Environ. 2012, 156, 116–122. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Toda, M.; Uchida, Y. Long-term use of green manure legume and chemical fertiliser affect soil bacterial community structures but not the rate of soil nitrate decrease when excess carbon and nitrogen are applied. Soil Res. 2017, 55, 524. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Ito, E.; Ono, K.; Ito, Y.M.; Araki, M. A Neural network approach to simple prediction of soil nitrification potential: A case study in Japanese temperate forests. Ecol. Model. 2008, 219, 200–211. [Google Scholar] [CrossRef]
- Mogi, H.; Anzai, M.; Uchida, Y. Heterogeneity of nitrification potentials within a paddock of a sheep farming system. Grassl. Sci. 2017, 63, 132–138. [Google Scholar] [CrossRef]
- Bastida, F.; Jindo, K.; Moreno, J.L.; Hernandez, T.; Garcia, C. Effects of organic amendments on soil carbon fractions, enzyme activity and humus-enzyme complexes under semi-arid conditions. Eur. J. Soil Biol. 2012, 53, 94–102. [Google Scholar] [CrossRef]
- Khalil, M.I.; Hossain, M.B.; Schmidhalter, U. Carbon and nitrogen mineralization in different upland soils of the subtropics treated with organic materials. Soil Biol. Biochem. 2005, 37, 1507–1518. [Google Scholar] [CrossRef]
- Aira, M.; Monroy, F.; Dominguez, J.; Mato, S. How earthworm density affects microbial biomass and activity in pig manure. Eur. J. Soil Biol. 2002, 38, 7–10. [Google Scholar] [CrossRef]
- Ruz-Jerez, B.E.; Ball, P.R.; Tillman, R.W. Laboratory assessment of nutrient release from a pasture soil receiving grass or clover residues, in the presence or absence of Lumbricus rubellus or Eisenia fetida. Soil Biol. Biochem. 1992, 24, 1529–1534. [Google Scholar] [CrossRef]
- Hendriksen, N.B. Leaf litter selection by detritivore and geophagous earthworms. Biol. Fertil. Soils 1990, 10, 17–21. [Google Scholar]
- Nakamura, Y. The effect of soil management on the soil faunal makeup of a cropped Andosol in central Japan. Soil Tillage. Res. 1988, 12, 177–186. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Kyoshima, T.; Kaneko, N. Mineral nitrogen dynamics in the casts of epigeic earthworms (Metaphire hilgendorfi: Megascolecidae). Soil Sci. Plant. Nutr. 2011, 57, 387–395. [Google Scholar] [CrossRef]
- Greiner, H.G.; Kashian, D.R.; Tiegs, S.D. Impacts of invasive Asian (Amynthas hilgendorfi) and European (Lumbricus rubellus) earthworms in a north American temperate deciduous forest. Biol. Invasions 2012, 14, 2017–2027. [Google Scholar] [CrossRef]
- Hartenstein, R.; Hartenstein, F. Physicochemical changes effected in activated sludge by the earthworm Eisenia foetida. J. Environ. Qual. 1981, 10, 377. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Parkinson, C.M.; Hamilton, W.E.; Dindal, D.L. Role of the earthworm, Eisenia foetida, in affecting organic matter decomposition in microcosms of sludge-amended Soil. J. Appl. Ecol. 1982, 19, 805. [Google Scholar] [CrossRef]
- Subler, S.; Kirsch, A.S. Spring Dynamics of soil carbon, nitrogen, and microbial activity in earthworm middens in a no-till cornfield. Biol. Fertil. Soils 1998, 26, 243–249. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar]
- Kruck, S.; Joschko, M.; Schultz-Sternberg, R.; Kroschewski, B.; Tessmann, J. A classification scheme for earthworm populations (Lumbricidae) in cultivated agricultural soils in Brandenburg, Germany. J. Plant Nutr. Soil Sci. 2006, 169, 651–660. [Google Scholar] [CrossRef]
- Lubbers, I.M.; van Groenigen, K.J.; Fonte, S.J.; Six, J.; Brussaard, L.; van Groenigen, J.W. Greenhouse-Gas emissions from soils increased by earthworms. Nat. Clim. Chang. 2013, 3, 187–194. [Google Scholar] [CrossRef]
- Lubbers, I.M.; Pulleman, M.M.; van Groenigen, J.W. Can earthworms simultaneously enhance decomposition and stabilization of plant residue carbon? Soil Biol. Biochem. 2017, 105, 12–24. [Google Scholar] [CrossRef]
- Zhu, X.; Chang, L.; Li, J.; Liu, J.; Feng, L.; Wu, D. Interactions between earthworms and mesofauna affect CO2 and N2O emissions from soils under long-term conservation tillage. Geoderma 2018, 332, 153–160. [Google Scholar] [CrossRef]
- Caravaca, F.; Roldán, A. Effect of Eisenia foetida earthworms on mineralization kinetics, microbial biomass, enzyme activities, respiration and labile C fractions of three soils treated with a composted organic residue. Biol. Fertil. Soils 2003, 38, 45–51. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| M | E | B | C | |
|---|---|---|---|---|
| NH4+-N (mg N kg−1 soil) | 24.1 ± 10.6 | 14.7 ± 9.2 | 21.0 ± 2.9 | 20.0 ± 1.7 |
| TN (g N kg−1 soil) | 5.43 ± 0.25 | 5.28 ± 0.37 | 4.52 ± 1.29 | 5.58 ± 0.26 |
| HWSC (mg C kg−1 soil) | 815 ± 24 | 867 ± 123 | 800 ± 8 | 843 ± 68 |
| TC (g C kg−1 soil) | 45.8 ± 6.29 | 39.6 ± 0.37 | 39.4 ± 0.2 | 40.7 ± 0.08 |
| CN ratio | 8.49 ± 1.54 | 7.53 ± 0.45 | 9.69 ± 3.39 | 7.31 ± 0.34 |
| UA (mg NH4+-N kg−1 soil h−1) | 7.61 ± 6.82 | 23.5 ± 4.04 | 19.5 ± 10.11 | 17.5 ± 9.31 |
| NEA (µg NO3−-N kg−1 soil h−1) | 194 ± 42 | 233 ± 59 | 91 ± 58 | 121 ± 41 |
| DEA (µg N2O-N kg−1 soil h−1) | 158 ± 97 | 85 ± 7 | 93 ± 18 | 66 ± 16 |
| SIR (mg CO2 kg−1 soil h−1) | 3.34 ± 0.73 | 3.14 ± 0.44 | 3.50 ± 0.60 | 2.15 ± 0.15 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamamoto, T.; Uchida, Y. The Role of Different Earthworm Species (Metaphire Hilgendorfi and Eisenia Fetida) on CO2 Emissions and Microbial Biomass during Barley Decomposition. Sustainability 2019, 11, 6544. https://doi.org/10.3390/su11236544
Hamamoto T, Uchida Y. The Role of Different Earthworm Species (Metaphire Hilgendorfi and Eisenia Fetida) on CO2 Emissions and Microbial Biomass during Barley Decomposition. Sustainability. 2019; 11(23):6544. https://doi.org/10.3390/su11236544
Chicago/Turabian StyleHamamoto, Toru, and Yoshitaka Uchida. 2019. "The Role of Different Earthworm Species (Metaphire Hilgendorfi and Eisenia Fetida) on CO2 Emissions and Microbial Biomass during Barley Decomposition" Sustainability 11, no. 23: 6544. https://doi.org/10.3390/su11236544
APA StyleHamamoto, T., & Uchida, Y. (2019). The Role of Different Earthworm Species (Metaphire Hilgendorfi and Eisenia Fetida) on CO2 Emissions and Microbial Biomass during Barley Decomposition. Sustainability, 11(23), 6544. https://doi.org/10.3390/su11236544