Light Pollution, Circadian Photoreception, and Melatonin in Vertebrates

 , , ,
, , ,  , ,
, ,  , ,
, ,
Abstract
1. Introduction
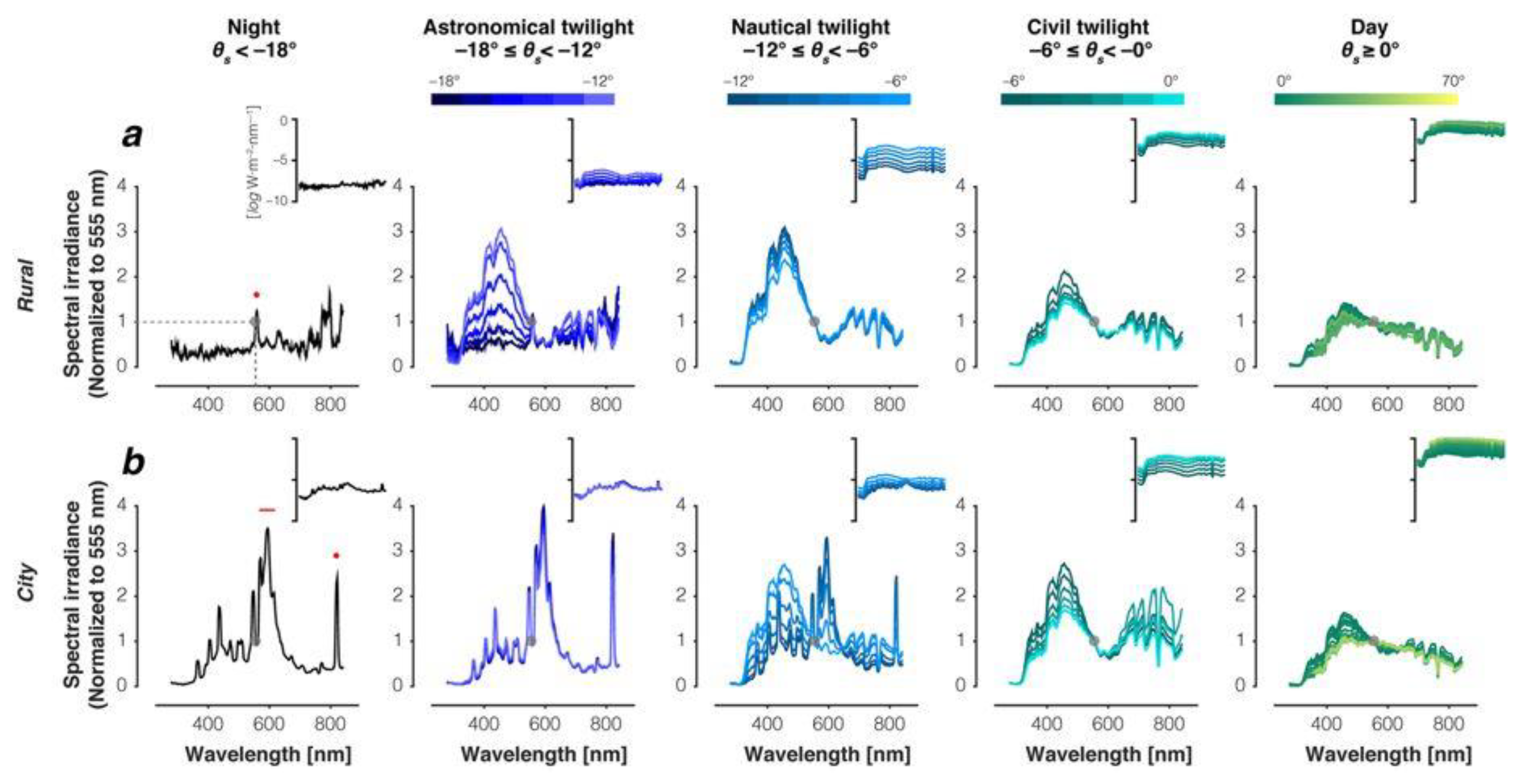
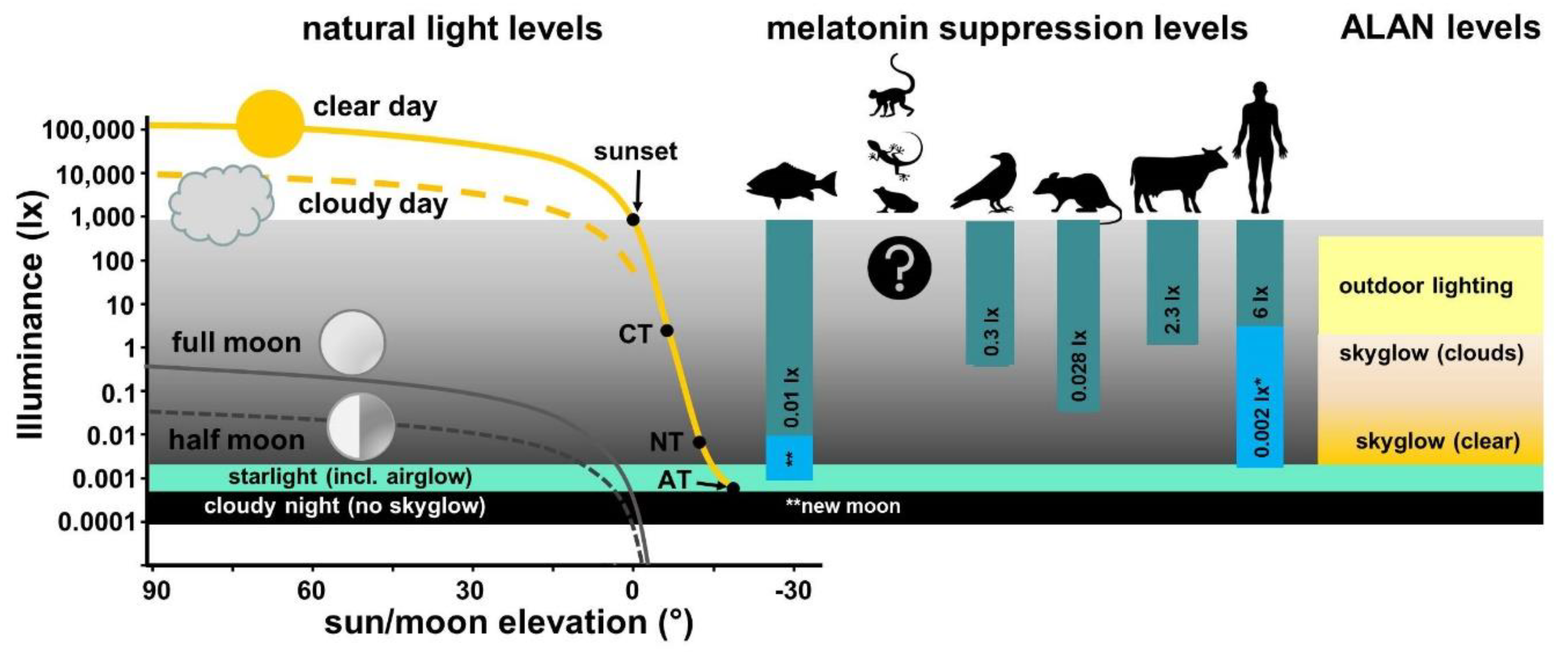
2. Natural Light and Light Pollution
2.1. Illuminance and Spectral Composition of Natural Light
2.2. Light Pollution
3. Photoreception and Circadian Systems in Vertebrates
3.1. Diversity of Photoreceptive Organs and Different Photoentrainment Pathways
3.2. Evolutionary Aspects of Diverse Photoreceptive Organs
4. Effects of Artificial Light at Night on Melatonin Production in Different Vertebrate Classes: A Systematic Review of the Literature
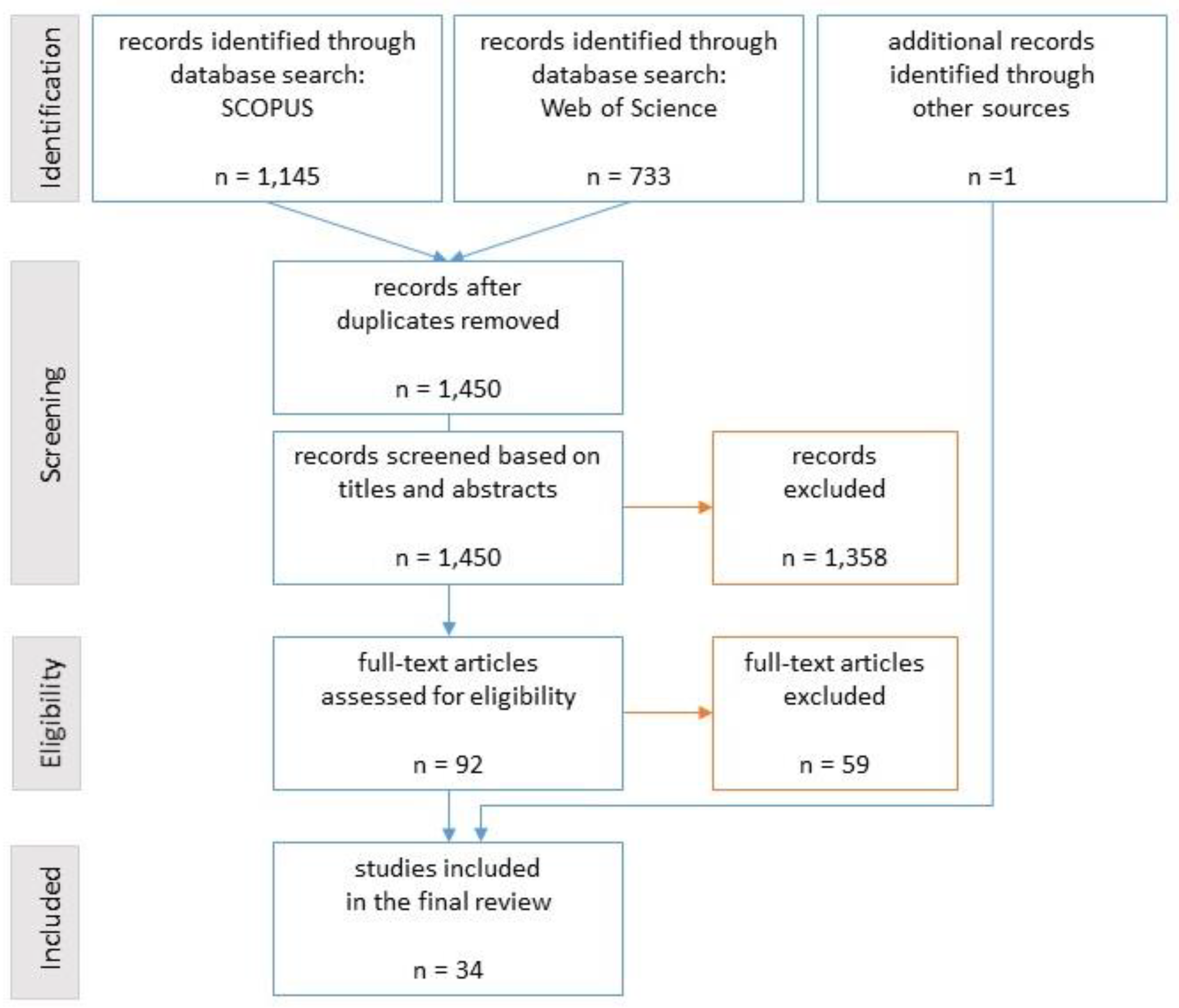
4.1. Search and Eligibility Criteria
4.2. Result Summary
4.2.1. Fishes
4.2.2. Amphibians
4.2.3. Reptiles
4.2.4. Birds
4.2.5. Non-Human Mammals (Rodents, Ungulates, and Primates)
4.2.6. Mammals (Humans)
5. Discussion
5.1. Fishes
5.2. Amphibians
5.3. Reptiles
5.4. Birds
5.5. Non-Human Mammals (Rodents, Ungulates, and Primates)
5.6. Mammals (Humans)
5.7. A Note on Experimental Design
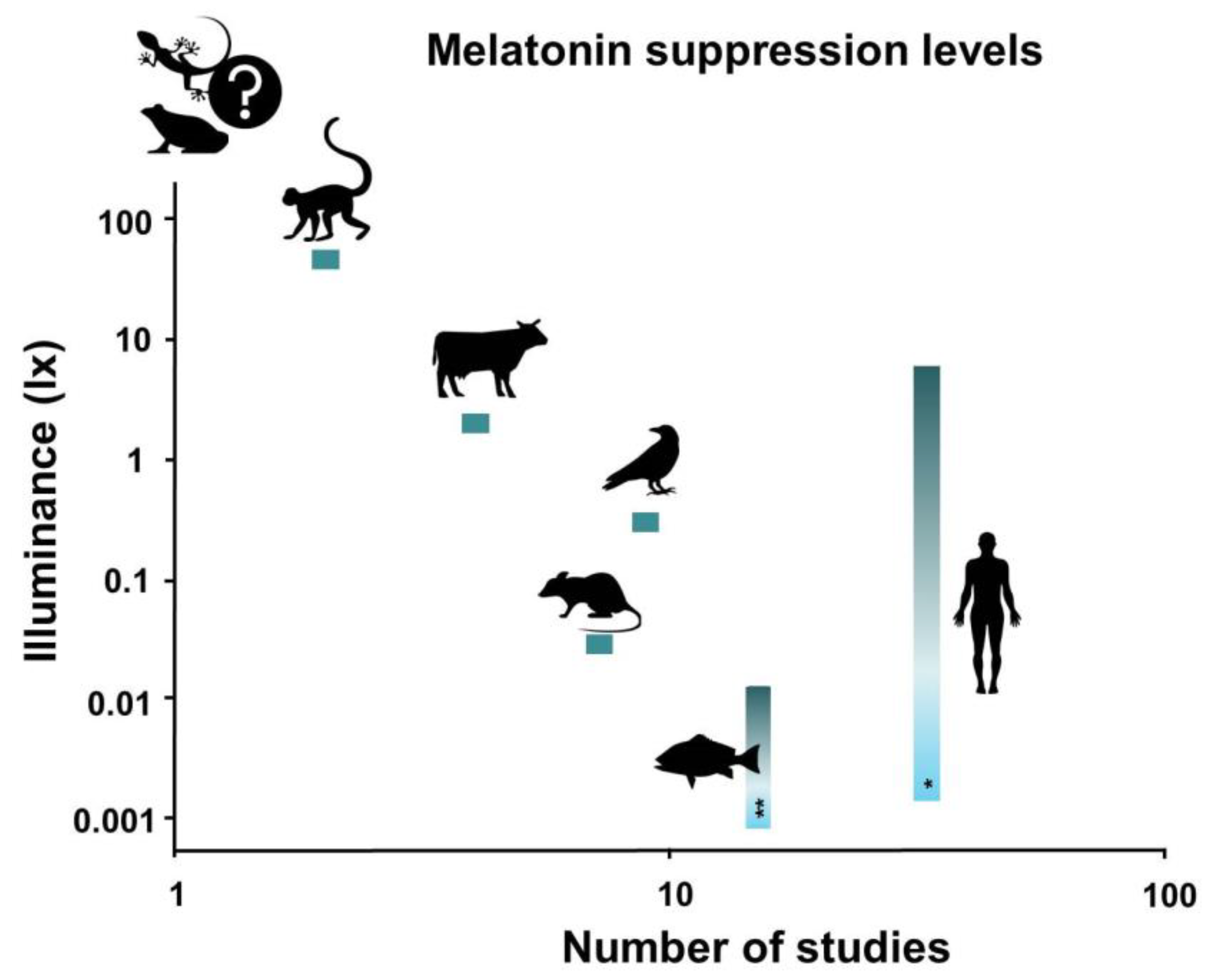
5.8. A Note on the Measurement of Light Intensities
5.9. Research Gaps and Recommendations for Future Research
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
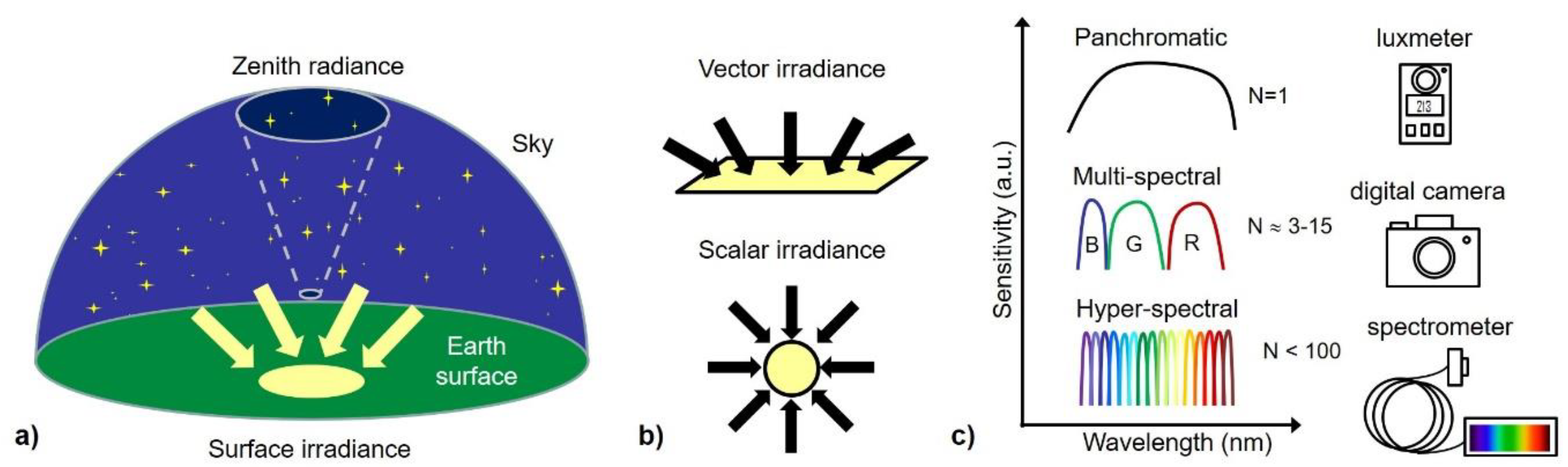
Appendix A. A Short Overview of Relevant Radiometry and Light Propagation Basics
Appendix A.1. Further Reading
Appendix A.2. General Radiometry and Geometry

Appendix A.3. Spectral Measurements in Single or Multiple Spectral Bands
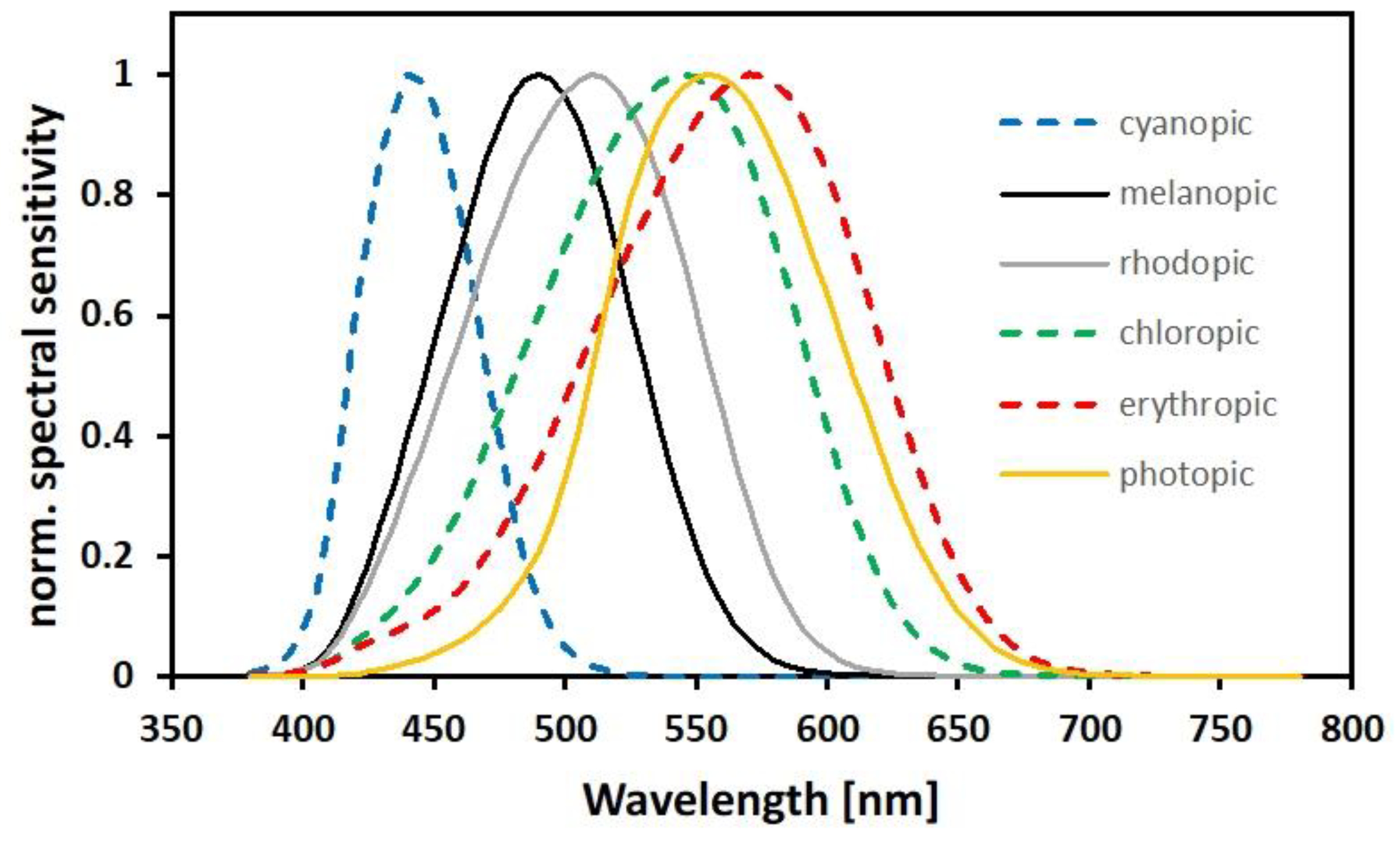
Appendix A.3.1. Photometric Quantities
Appendix A.3.2. Photosynthetically Active Radiation
Appendix A.3.3. Astronomical Magnitudes
Appendix A.3.4. Multi- and Hyperspectral Bands
Appendix A.4. Conversion between Units
Appendix A.4.1. Conversion between Different Geometries
Appendix A.4.2. Conversion of Photon Flux to Radiant Flux
Appendix A.4.3. Conversion between Spectral Bands

Appendix A.4.4. Example: Converting Photon Flux per Unit Area to Illuminance
Appendix A.5. Recommendations: What to Measure and How
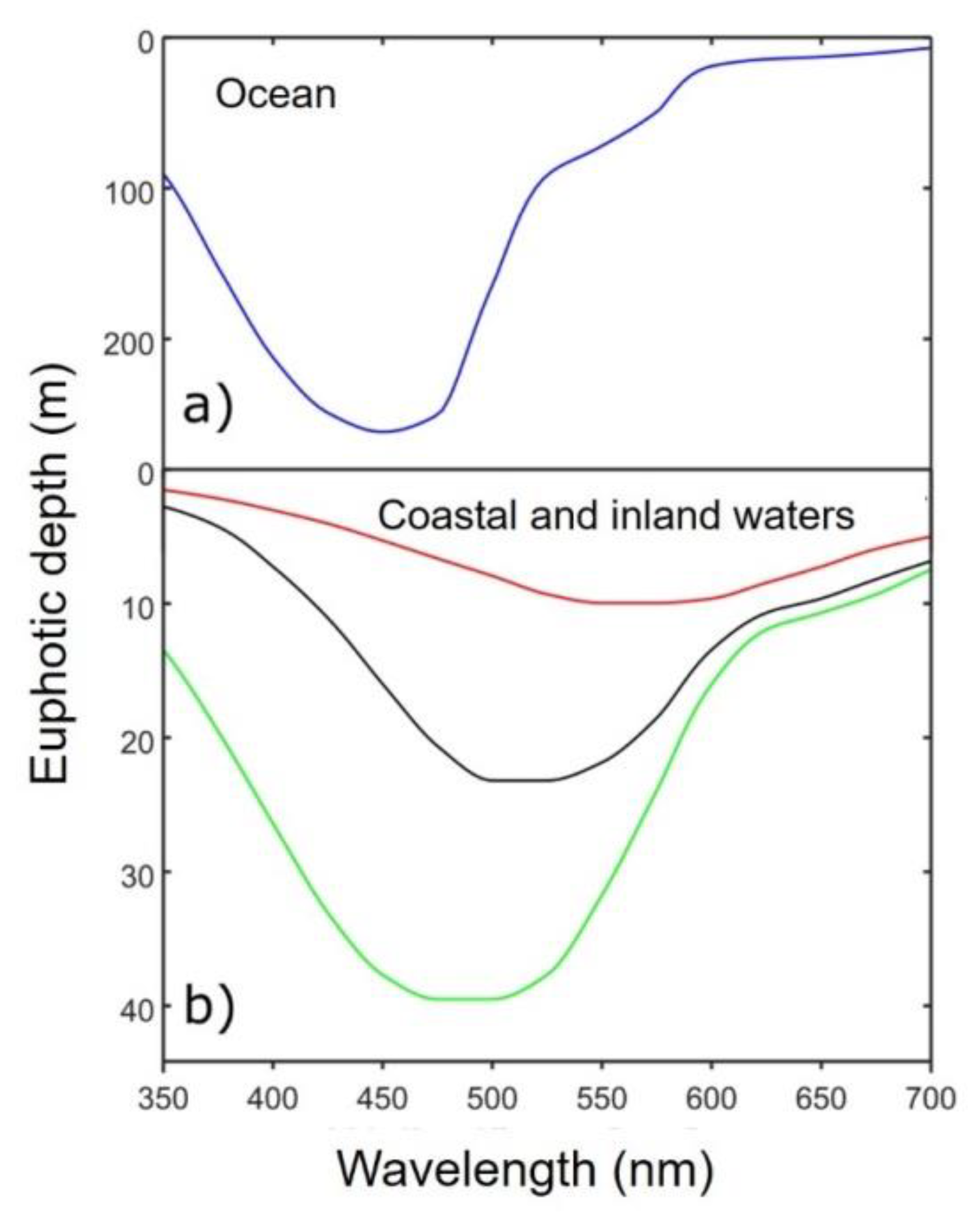
Appendix A6. Light Propagation in a Medium (Mainly Relevant for Water in the Context of This Paper)

References
- Navara, K.J.; Nelson, R.J. The dark side of light at night: Physiological, epidemiological, and ecological consequences. J. Pineal Res. 2007, 43, 215–224. [Google Scholar] [CrossRef]
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, N.; Hass, R.; Lerner, A.B.; Case, J.D.; Lee, T.H.; Mori, W.; et al. Isolation of melatonin, the pineal gland factor that lightens melanocytes. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar] [CrossRef]
- Vivien-Roels, B.; Pévet, P. Melatonin: Presence and formation in invertebrates. Experientia 1993, 49, 642–647. [Google Scholar] [CrossRef]
- Tal, O.; Haim, A.; Harel, O.; Gerchman, Y. Melatonin as an antioxidant and its semi-lunar rhythm in green macroalga Ulva sp. J. Exp. Bot. 2011, 62, 1903–1910. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin synthesis and function: Evolutionary history in animals and plants. Front. Endocrinol. 2019, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Hardeland, R.; Manchester, L.C.; Paredes, S.D.; Korkmaz, A.; Sainz, R.M.; Mayo, J.C.; Fuentes-Broto, L.; Reiter, R.J. The changing biological roles of melatonin during evolution: From an antioxidant to signals of darkness, sexual selection and fitness. Biol. Rev. 2010, 85, 607–623. [Google Scholar] [CrossRef] [PubMed]
- Lewy, A.J.; Wehr, T.A.; Goodwin, F.K.; Newsome, D.A.; Markey, S.P. Light suppresses melatonin secretion in humans. Science 1980, 210, 1267–1269. [Google Scholar] [CrossRef] [PubMed]
- Aschoff, J. Exogenous and endogenous components in circadian rhythms. Cold Spring Harb. Symp. Quant. Biol. 1960, 25, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Foster, R.G. Twilight times: Light and the circadian system. Photochem. Photobiol. 1997, 66, 549–561. [Google Scholar] [CrossRef]
- Falcón, J.; Besseau, L.; Fuentès, M.; Sauzet, S.; Magnanou, E.; Boeuf, G. Structural and functional evolution of the pineal melatonin system in vertebrates. Ann. N. Y. Acad. Sci. 2009, 1163, 101–111. [Google Scholar] [CrossRef]
- Vivid, D.; Bentley, G.E. Seasonal reproduction in vertebrates: Melatonin synthesis, binding, and functionality using tinbergen’s four questions. Molecules 2018, 23, 652. [Google Scholar] [CrossRef] [PubMed]
- Gern, W.A.; Duvall, D.; Nervina, J.M. Melatonin: A discussion of its evolution and actions in vertebrates. Am. Zool. 1986, 26, 985–996. [Google Scholar] [CrossRef]
- Wiechmann, A.F.; Sherry, D.M. Role of melatonin and its receptors in the vertebrate retina. Int. Rev. Cell Mol. Biol. 2013, 300, 211–242. [Google Scholar] [PubMed]
- Gaston, K.J.; Visser, M.E.; Hölker, F. The biological impacts of artificial light at night: The research challenge. Phil. Trans. R. Soc. B 2015, 370, 20140133. [Google Scholar] [CrossRef] [PubMed]
- De Jong, M.; Jeninga, L.; Ouyang, J.Q.; van Oers, K.; Spoelstra, K.; Visser, M.E. Dose-dependent responses of avian daily rhythms to artificial light at night. Physiol. Behav. 2016, 155, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Kayumov, L.; Casper, R.F.; Hawa, R.J.; Perelman, B.; Chung, S.A.; Sokalsky, S.; Shapiro, C.M. Blocking low-wavelength light prevents nocturnal melatonin suppression with no adverse effect on performance during simulated shift work. J. Clin. Endocrinol. Metab. 2005, 90, 2755–2761. [Google Scholar] [CrossRef]
- Kyba, C.C.M.; Kuester, T.; de Miguel, A.S.; Baugh, K.; Jechow, A.; Hölker, F.; Bennie, J.; Elvidge, C.D.; Gaston, K.J.; Guanter, L. Artificially lit surface of Earth at night increasing in radiance and extent. Sci. Adv. 2017, 3, e1701528. [Google Scholar] [CrossRef]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by retinal ganglion cells that set the circadian clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef]
- Takahashi, J.S.; DeCoursey, P.J.; Bauman, L.; Menaker, M. Spectral sensitivity of a novel photoreceptive system mediating entrainment of mammalian circadian rhythms. Nature 1984, 308, 186–188. [Google Scholar] [CrossRef]
- Barbier, D. The airglow. Vistas Astron. 1956, 2, 929–939. [Google Scholar] [CrossRef]
- Brown, D.R. Natural Illumination Charts: Research and Development Project NS 714-100; Deparment of the Navy, Bureau of Ships: Washington, DC, USA, 1952. [Google Scholar]
- Kyba, C.C.M.; Mohar, A.; Posch, T. How bright is moonlight? Astron. Geophys. 2017, 58, 31–32. [Google Scholar] [CrossRef]
- Hänel, A.; Posch, T.; Ribas, S.J.; Aubé, M.; Duriscoe, D.; Jechow, A.; Kollath, Z.; Lolkema, D.E.; Moore, C.; Schmidt, N.; et al. Measuring night sky brightness: Methods and challenges. J. Quant. Spectrosc. Radiat. Transf. 2018, 205, 278–290. [Google Scholar] [CrossRef]
- Jechow, A.; Hölker, F. Snowglow—The amplification of skyglow by snow and clouds can exceed full moon illuminance in suburban areas. J. Imaging 2019, 5, 69. [Google Scholar] [CrossRef]
- Spitschan, M.; Aguirre, G.K.; Brainard, D.H.; Sweeney, A.M. Variation of outdoor illumination as a function of solar elevation and light pollution. Sci. Rep. 2016, 6, 26756. [Google Scholar] [CrossRef] [PubMed]
- Lythgoe, J.N. The Ecology of Vision, 1st ed.; Clarendon Press: Oxford, UK, 1979; ISBN 978-0-19-854529-3. [Google Scholar]
- Riegel, K.W. Light Pollution: Outdoor lighting is a growing threat to astronomy. Science 1973, 179, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Schroer, S.; Hölker, F. Impact of lighting on flora and fauna. In Handbook of Advanced Lighting Technology; Karlicek, R., Sun, C.-C., Zissis, G., Ma, R., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–33. ISBN 978-3-319-00295-8. [Google Scholar]
- Cho, Y.; Ryu, S.-H.; Lee, B.R.; Kim, K.H.; Lee, E.; Choi, J. Effects of artificial light at night on human health: A literature review of observational and experimental studies applied to exposure assessment. Chronobiol. Int. 2015, 32, 1294–1310. [Google Scholar] [CrossRef] [PubMed]
- Longcore, T.; Rich, C. Ecological light pollution. Front. Ecol. Environ. 2004, 2, 191–198. [Google Scholar] [CrossRef]
- Hölker, F.; Wolter, C.; Perkin, E.K.; Tockner, K. Light pollution as a biodiversity threat. Trends Ecol. Evol. 2010, 25, 681–682. [Google Scholar] [CrossRef]
- Hölker, F.; Wurzbacher, C.; Weißenborn, C.; Monaghan, M.T.; Holzhauer, S.I.J.; Premke, K. Microbial diversity and community respiration in freshwater sediments influenced by artificial light at night. Phil. Trans. R. Soc. B 2015, 370, 20140130. [Google Scholar] [CrossRef]
- Knop, E.; Zoller, L.; Ryser, R.; Gerpe, C.; Hörler, M.; Fontaine, C. Artificial light at night as a new threat to pollination. Nature 2017, 548, 206–209. [Google Scholar] [CrossRef]
- Kurvers, R.H.J.M.; Drägestein, J.; Hölker, F.; Jechow, A.; Krause, J.; Bierbach, D. Artificial light at night affects emergence from a refuge and space use in guppies. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hölker, F.; Moss, T.; Griefahn, B.; Kloas, W.; Voigt, C.; Henckel, D.; Hänel, A.; Kappeler, P.; Völker, S.; Schwope, A.; et al. The dark side of light: A transdisciplinary research agenda for light pollution policy. Ecol. Soc. 2010, 15. [Google Scholar] [CrossRef]
- Kyba, C.C.M.; Hänel, A.; Hölker, F. Redefining efficiency for outdoor lighting. Energy Environ. Sci. 2014, 7, 1806–1809. [Google Scholar] [CrossRef]
- Rosebrugh, D.W. Sky-glow from large cities. JR Astron. Soc. Can. 1935, 29, 79. [Google Scholar]
- Falchi, F.; Cinzano, P.; Duriscoe, D.; Kyba, C.C.M.; Elvidge, C.D.; Baugh, K.; Portnov, B.A.; Rybnikova, N.A.; Furgoni, R. The new world atlas of artificial night sky brightness. Sci. Adv. 2016, 2, e1600377. [Google Scholar] [CrossRef] [PubMed]
- Jechow, A.; Kolláth, Z.; Ribas, S.J.; Spoelstra, H.; Hölker, F.; Kyba, C.C.M. Imaging and mapping the impact of clouds on skyglow with all-sky photometry. Sci. Rep. 2017, 7, 6741. [Google Scholar] [CrossRef] [PubMed]
- Menaker, M.; Moreira, L.F.; Tosini, G. Evolution of circadian organization in vertebrates. Braz. J. Med. Biol. Res. 1997, 30, 305–313. [Google Scholar] [CrossRef]
- Doyle, S.; Menaker, M. Circadian photoreception in vertebrates. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 499–508. [Google Scholar] [CrossRef]
- Pérez, J.H.; Tolla, E.; Dunn, I.C.; Meddle, S.L.; Stevenson, T.J. A comparative perspective on extra-retinal photoreception. Trends Endocrinol. Metab. 2019, 30, 39–53. [Google Scholar] [CrossRef]
- Peirson, S.N.; Halford, S.; Foster, R.G. The evolution of irradiance detection: Melanopsin and the non-visual opsins. Phil. Trans. R. Soc. B 2009, 364, 2849–2865. [Google Scholar] [CrossRef]
- Korf, H.W.; Schomerus, C.; Stehle, J.H. The pineal organ, its hormone melatonin, and the photoneuroendocrine system. Adv. Anat. Embryol. Cell Biol. 1998, 146, 1–100. [Google Scholar] [PubMed]
- Dodt, E.; Meissl, H. The pineal and parietal organs of lower vertebrates. Experientia 1982, 38, 996–1000. [Google Scholar] [CrossRef] [PubMed]
- Ganong, W.F.; Shepherd, M.D.; Wall, J.R.; Van Brunt, E.E.; Clegg, M.T. Penetration of light into the brain of mammals. Endocrinology 1963, 72, 962–963. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, H.G.; van Veen, T. Spectral characteristics of visible radiation penetrating into the brain and stimulating extraretinal photoreceptors. J. Comp. Physiol. 1979, 130, 277–282. [Google Scholar] [CrossRef]
- Dodt, E. The parietal eye (pineal and parietal organs) of lower vertebrates. In Visual Centers in the Brain; Handbook of Sensory Physiology; Berlucchi, G., Brindley, G.S., Brooks, B., Creutzfeldt, O.D., Dodt, E., Doty, R.W., Freund, H.-J., Gross, C.G., Jeffreys, D.A., Eds.; Springer: Berlin/Heidelberg, Germany, 1973; pp. 113–140. ISBN 978-3-642-65495-4. [Google Scholar]
- Klein, D.C.; Coon, S.L.; Roseboom, P.H.; Weller, J.L.; Bernard, M.; Gastel, J.A.; Zatz, M.; Iuvone, P.M.; Rodriguez, I.R.; Bégay, V.; et al. The melatonin rhythm-generating enzyme: Molecular regulation of serotonin N-acetyltransferase in the pineal gland. Recent Prog. Horm. Res. 1997, 52, 307–357. [Google Scholar] [PubMed]
- Falcón, J.; Migaud, H.; Muñoz-Cueto, J.A.; Carrillo, M. Current knowledge on the melatonin system in teleost fish. Gen. Comp. Endocr. 2010, 165, 469–482. [Google Scholar] [CrossRef]
- Erren, T.C.; Erren, M.; Lerchl, A.; Meyer-Rochow, V.B. Clockwork blue: On the evolution of non-image-forming retinal photoreceptors in marine and terrestrial vertebrates. Naturwissenschaften 2008, 95, 273–279. [Google Scholar] [CrossRef]
- Walmsley, L.; Hanna, L.; Mouland, J.; Martial, F.; West, A.; Smedley, A.R.; Bechtold, D.A.; Webb, A.R.; Lucas, R.J.; Brown, T.M. Colour as a signal for entraining the mammalian circadian clock. PLoS Biol. 2015, 13, e1002127. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Vera, L.M.; López-Olmeda, J.F.; Bayarri, M.J.; Madrid, J.A.; Sánchez-Vázquez, F.J. Influence of light intensity on plasma melatonin and locomotor activity rhythms in tench. Chronobiol. Int. 2005, 22, 67–78. [Google Scholar] [CrossRef]
- Nikaido, Y.; Ueda, S.; Takemura, A. Photic and circadian regulation of melatonin production in the Mozambique tilapia Oreochromis mossambicus. Comp. Biochem Physiol. A Mol. Integr. Physiol. 2009, 152, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Kim, N.N.; Choi, Y.-U.; Choi, C.Y. Changes in circadian parameters of humbug damselfish, Dascyllus aruanus according to lunar phase shifts in Micronesia. Biol. Rhythm Res. 2017, 48, 475–483. [Google Scholar] [CrossRef]
- Rahman, M.S.; Kim, B.H.; Takemura, A.; Park, C.B.; Lee, Y.D. Effects of moonlight exposure on plasma melatonin rhythms in the seagrass rabbitfish, Siganus canaliculatus. J. Biol. Rhythms 2004, 19, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Kim, B.-H.; Takemura, A.; Park, C.-B.; Lee, Y.-D. Influence of light–dark and lunar cycles on the ocular melatonin rhythms in the seagrass rabbitfish, a lunar-synchronized spawner. J. Pineal Res. 2004, 37, 122–128. [Google Scholar] [CrossRef]
- Liu, Q.; Manning, A.J.; Duston, J. Light intensity and suppression of nocturnal plasma melatonin in Arctic charr (Salvelinus alpinus). Comp. Biochem Physiol. A Mol. Integr. Physiol. 2019, 228, 103–106. [Google Scholar] [CrossRef]
- Park, Y.J.; Park, J.G.; Takeuchi, Y.; Hur, S.P.; Lee, Y.D.; Kim, S.J.; Takemura, A. Influence of moonlight on mRNA expression patterns of melatonin receptor subtypes in the pineal organ of a tropical fish. Mar. Genom. 2014, 14, 67–70. [Google Scholar] [CrossRef]
- Porter, M.J.R.; Duncan, N.J.; Handeland, S.O.; Stefansson, S.O.; Bromage, N.R. Temperature, light intensity and plasma melatonin levels in juvenile Atlantic salmon. J. Fish. Biol. 2001, 58, 431–438. [Google Scholar] [CrossRef]
- Bayarri, M.J.; Madrid, J.A.; Sánchez-Vázquez, F.J. Influence of light intensity, spectrum and orientaion on sea bass plasma and ocular melatonin. J. Pineal Res. 2002, 32, 34–40. [Google Scholar] [CrossRef]
- Brüning, A.; Kloas, W.; Preuer, T.; Hölker, F. Influence of artificially induced light pollution on the hormone system of two common fish species, perch and roach, in a rural habitat. Conserv. Physiol. 2018, 6, coy016. [Google Scholar] [CrossRef]
- Brüning, A.; Hölker, F.; Franke, S.; Preuer, T.; Kloas, W. Spotlight on fish: Light pollution affects circadian rhythms of European perch but does not cause stress. Sci. Tot. Environ. 2015, 511, 516–522. [Google Scholar] [CrossRef]
- Brüning, A.; Hölker, F.; Franke, S.; Kleiner, W.; Kloas, W. Influence of light intensity and spectral composition of artificial light at night on melatonin rhythm and mRNA expression of gonadotropins in roach Rutilus rutilus. Fish. Physiol. Biochem. 2018, 44, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kupprat, F.; Hölker, F.; Kloas, W. Can skyglow reduce nocturnal melatonin concentrations in European perch? Environ. Pollut. under review.
- Brüning, A.; Hölker, F.; Franke, S.; Kleiner, W.; Kloas, W. Impact of different colours of artificial light at night on melatonin rhythm and gene expression of gonadotropins in European perch. Sci. Tot. Environ. 2016, 543, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, T.; Park, Y.J.; Park, J.G.; Imamura, S.; Takeuchi, Y.; Hur, S.P.; Takemura, A. Moonlight affects mRNA abundance of arylalkylamine N-acetyltransferase in the retina of a lunar-synchronized spawner, the goldlined spinefoot. J. Exp. Zool. A Ecol. Genet. Physiol. 2013, 319, 505–516. [Google Scholar] [PubMed]
- Iigo, M.; Sato, M.; Ikeda, E.; Kawasaki, S.; Noguchi, F.; Nishi, G. Effects of photic environment on ocular melatonin contents in a labrid teleost, the wrasse Halichoeres tenuispinnis. Gen. Comp. Endocrinol. 2003, 133, 252–259. [Google Scholar] [CrossRef]
- Rich, C.; Longcore, T. (Eds.) Ecological Consequences of Artificial Night Lighting; Island Press: Washington, DC, USA, 2006. [Google Scholar]
- Wise, S. Studying the ecological impacts of light pollution on wildlife: Amphibians as models. In Starlight: A Common Heritage; Cipriano, M., Jafar, J., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 209–218. [Google Scholar]
- Perry, G.; Buchanan, B.W.; Fisher, R.N.; Salmon, M.; Wise, S.E. Effects of artificial night lighting on amphibians and reptiles in urban environments. In Urban Herpetology; Mitchell, J.C., Brown, R.E.J., Eds.; Society for the study of Amphibians and Reptiles: Salt Lake City, UT, USA, 2008. [Google Scholar]
- Dananay, K.L.; Benard, M.F. Artificial light at night decreases metamorphic duration and juvenile growth in a widespread amphibian. Proc. R. Soc. B. 2018, 285. [Google Scholar] [CrossRef]
- Touzot, M.; Teulier, L.; Lengagne, T.; Secondi, J.; Théry, M.; Libourel, P.-A.; Guillard, L.; Mondy, N. Artificial light at night disturbs the activity and energy allocation of the common toad during the breeding period. Conserv. Physiol. 2019, 7. [Google Scholar] [CrossRef]
- Dias, K.S.; Dosso, E.S.; Hall, A.S.; Schuch, A.P.; Tozetti, A.M. Ecological light pollution affects anuran calling season, daily calling period, and sensitivity to light in natural Brazilian wetlands. Sci. Nat. 2019, 106, 46. [Google Scholar] [CrossRef]
- Underhill, V.A.; Höbel, G. Mate choice behavior of female Eastern Gray Treefrogs (Hyla versicolor) is robust to anthropogenic light pollution. Ethology 2018, 124, 537–548. [Google Scholar] [CrossRef]
- Wright, M.L.; Bruni, N.K. Influence of the photocycle and thermocycle on rhythms of plasma thyroxine and plasma and ocular melatonin in late metamorphic stages of the bullfrog tadpole, Rana catesbeiana. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2004, 139, 33–40. [Google Scholar] [CrossRef]
- Wright, M.L.; Duffy, J.L.; Guertin, C.J.; Alves, C.D.; Szatkowski, M.C.; Visconti, R.F. Developmental and diel changes in plasma thyroxine and plasma and ocular melatonin in the larval and juvenile bullfrog, Rana catesbeiana. Gen. Comp. Endocrinol. 2003, 130, 120–128. [Google Scholar] [CrossRef]
- Wright, M.L.; Francisco, L.L.; Scott, J.L.; Richardson, S.E.; Carr, J.A.; King, A.B.; Noyes, A.G.; Visconti, R.F. Effects of bilateral and unilateral ophthalmectomy on plasma melatonin in Rana tadpoles and froglets under various experimental conditions. Gen. Comp. Endocrinol. 2006, 147, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Vanecek, J. Cellular mechanisms of melatonin action. Physiol. Rev. 1998, 78, 687–721. [Google Scholar] [CrossRef] [PubMed]
- Binkley, S.; Mosher, K.; Rubin, F.; White, B. Xenopus tadpole melanophores are controlled by dark and light and melatonin without influence of time of day. J. Pineal Res. 1988, 5, 87–97. [Google Scholar] [CrossRef]
- Lee, J.H.; Hung, C.F.; Ho, C.C.; Chang, S.H.; Lai, Y.S.; Chung, J.G. Light-induced changes in frog pineal gland N-acetyltransferase activity. Neurochem. Int. 1997, 31, 533–540. [Google Scholar] [CrossRef]
- Green, C.B.; Liang, M.Y.; Steenhard, B.M.; Besharse, J.C. Ontogeny of circadian and light regulation of melatonin release in Xenopus laevis embryos. Brain Res. Dev. Brain Res. 1999, 117, 109–116. [Google Scholar] [CrossRef]
- Foster, R.G.; Roberts, A. The pineal eye in Xenopus laevis embryos and larvae: A photoreceptor with a direct excitatory effect on behaviour. J. Comp. Physiol. 1982, 145, 413–419. [Google Scholar] [CrossRef]
- Loew, E.R.; Lythgoe, J.N. The ecology of colour vision. Endeavour 1985, 9, 170–174. [Google Scholar] [CrossRef]
- Cahill, G.M.; Parsons, S.E.; Besharse, J.C. Spectral sensitivity of melatonin synthesis suppression in Xenopus eyecups. Vis. Neurosci. 1998, 15, 499–502. [Google Scholar] [CrossRef]
- Firth, B.T.; Christian, K.A.; Belan, I.; Kennaway, D.J. Melatonin rhythms in the Australian freshwater crocodile (Crocodylus johnstoni): A reptile lacking a pineal complex? J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2010, 180, 67–72. [Google Scholar] [CrossRef]
- Roth, J.J.; Gern, W.A.; Roth, E.C.; Ralph, C.L.; Jacobson, E. Nonpineal melatonin in the alligator (Alligator mississippiensis). Science 1980, 210, 548–550. [Google Scholar] [CrossRef] [PubMed]
- Bertolucci, C.; Frigato, E.; Foà, A. The reptilian clock system: Circadian clock, extraretinal photoreception, and clock-dependent celestial compass orientation mechanisms in reptiles. In Biological Timekeeping: Clocks, Rhythms and Behaviour; Springer: New Delhi, India, 2017; pp. 223–239. [Google Scholar]
- Underwood, H. Pineal melatonin rhythms in the lizard Anolis carolinensis: Effects of light and temperature cycles. J. Comp. Physiol. A Sens Neural Behav. Physiol. 1985, 157, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Hyde, L.L.; Underwood, H. Effects of nightbreak, T-cycle, and resonance lighting schedules on the pineal melatonin rhythm of the lizard Anolis carolinensis: Correlations with the reproductive response. J. Pineal Res. 1993, 15, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Underwood, H.; Calaban, M. Pineal melatonin rhythms in the lizard Anolis carolinensis: I. Response to light and temperature cycles. J. Biol. Rhythms 1987, 2, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Firth, B.T.; Belan, I.; Kennaway, D.J.; Moyer, R.W. Thermocyclic entrainment of lizard blood plasma melatonin rhythms in constant and cyclic photic environments. Am. J. Physiol. 1999, 277, R1620–R1626. [Google Scholar] [CrossRef]
- Moore, A.F.; Kawasaki, M.; Menaker, M. Photic induction of locomotor activity is correlated with photic habitat in Anolis lizards. J. Comp. Physiol. A Sens Neural Behav. Physiol. 2012, 198, 193–201. [Google Scholar] [CrossRef]
- Vivien-Roels, B.; Pévet, P.; Claustrat, B. Pineal and circulating melatonin rhythms in the box turtle, Terrapene carolina triunguis: Effect of photoperiod, light pulse, and environmental temperature. Gen. Comp. Endocrinol. 1988, 69, 163–173. [Google Scholar] [CrossRef]
- Dominoni, D.M.; Goymann, W.; Helm, B.; Partecke, J. Urban-like night illumination reduces melatonin release in European blackbirds (Turdus merula): Implications of city life for biological time-keeping of songbirds. Front. Zool. 2013, 10, 60. [Google Scholar] [CrossRef]
- Schoech, S.J.; Bowman, R.; Hahn, T.P.; Goymann, W.; Schwabl, I.; Bridge, E.S. The effects of low levels of light at night upon the endocrine physiology of western scrub-jays (Aphelocoma californica). J. Exp. Zool. A Ecol. Genet. Physiol. 2013, 319, 527–538. [Google Scholar] [CrossRef]
- Kumar, J.; Malik, S.; Bhardwaj, S.K.; Rani, S. Bright light at night alters the perception of daylength in Indian weaver bird (Ploceus philippinus). J. Exp. Zool. A Ecol. Integr. Physiol. 2018, 329, 488–496. [Google Scholar] [CrossRef]
- Jiang, J.; He, Y.; Kou, H.; Ju, Z.; Gao, X.; Zhao, H. The effects of artificial light at night on Eurasian tree sparrow (Passer montanus): Behavioral rhythm disruption, melatonin suppression and intestinal microbiota alterations. Ecol. Indic. 2020, 108, 105702. [Google Scholar] [CrossRef]
- Cockrem, J.F. Circadian rhythms of plasma melatonin in the Adelie penguin (Pygoscelis adeliae) in constant dim light and artificial photoperiods. J. Pineal Res. 1991, 11, 63–69. [Google Scholar] [CrossRef]
- Singh, J.; Rani, S.; Kumar, V. Functional similarity in relation to the external environment between circadian behavioral and melatonin rhythms in the subtropical Indian weaver bird. Horm. Behav. 2012, 61, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Vakkuri, O.; Rintamäki, H.; Leppäluoto, J. Plasma and tissue concentrations of melatonin after midnight light exposure and pinealectomy in the pigeon. J. Endocrinol. 1985, 105, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.D.; Perry, G.C.; Morris, T.R.; English, J. Supplementary dim light differentially influences sexual maturity, oviposition time, and melatonin rhythms in pullets. Poult. Sci. 2001, 80, 1723–1728. [Google Scholar] [CrossRef] [PubMed]
- Meyer, W.E.; Millam, J.R. Plasma melatonin levels in Japanese quail exposed to dim light are determined by subjective interpretation of day and night, not light intensity. Gen. Comp. Endocrinol. 1991, 82, 377–385. [Google Scholar] [CrossRef]
- Kennaway, D.J.; Voultsios, A.; Varcoe, T.J.; Moyer, R.W. Melatonin in mice: Rhythms, response to light, adrenergic stimulation, and metabolism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, R358–R365. [Google Scholar] [CrossRef][Green Version]
- Dauchy, R.T.; Wren, M.A.; Dauchy, E.M.; Hoffman, A.E.; Hanifin, J.P.; Warfield, B.; Jablonski, M.R.; Brainard, G.C.; Hill, S.M.; Mao, L.; et al. The influence of red light exposure at night on circadian metabolism and physiology in Sprague-Dawley rats. J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 40–50. [Google Scholar]
- Travlos, G.S.; Wilson, R.E.; Murrell, J.A.; Chignell, C.F.; Boorman, G.A. The effect of short intermittent light exposures on the melatonin circadian rhythm and NMU-induced breast cancer in female F344/N rats. Toxicol. Pathol. 2001, 29, 126–136. [Google Scholar] [CrossRef]
- Xiang, S.; Dauchy, R.T.; Hauch, A.; Mao, L.; Yuan, L.; Wren, M.A.; Belancio, V.P.; Mondal, D.; Frasch, T.; Blask, D.E.; et al. Doxorubicin resistance in breast cancer is driven by light at night-induced disruption of the circadian melatonin signal. J. Pineal Res. 2015, 59, 60–69. [Google Scholar] [CrossRef]
- Glickman, G.L.; Harrison, E.M.; Elliott, J.A.; Gorman, M.R. Increased photic sensitivity for phase resetting but not melatonin suppression in Siberian hamsters under short photoperiods. Horm. Behav. 2014, 65, 301–307. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hoffmann, K.; Illnerová, H.; Vaněček, J. Effect of photoperiod and of one minute light at night-time on the pineal rhythm on N-acetyltransferase activity in the Djungarian hamster Phodopus sungorus. Biol. Reprod. 1981, 24, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Schwimmer, H.; Mursu, N.; Haim, A. Effects of light and melatonin treatment on body temperature and melatonin secretion daily rhythms in a diurnal rodent, the fat sand rat. Chronobiol. Int. 2010, 27, 1401–1419. [Google Scholar] [CrossRef] [PubMed]
- Asher, A.; Shabtay, A.; Brosh, A.; Eitam, H.; Agmon, R.; Cohen-Zinder, M.; Zubidat, A.E.; Haim, A. “Chrono-functional milk”: The difference between melatonin concentrations in night-milk versus day-milk under different night illumination conditions. Chronobiol. Int. 2015, 32, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Deveson, S.L.; Arendt, J.; Forsyth, I.A. Sensitivity of goats to a light pulse during the night as assessed by suppression of melatonin concentrations in the plasma. J. Pineal Res. 1990, 8, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Vivien-Roels, B.; Pévet, P.; Zarazaga, L.; Malpaux, B.; Chemineau, P. Daily and light-at-night induced variations of circulating 5-methoxytryptophol (5-ML) in ewes with respectively high and low nocturnal melatonin secretion. J. Pineal Res. 1999, 27, 230–236. [Google Scholar] [CrossRef]
- Walsh, C.M.; Prendergast, R.L.; Sheridan, J.T.; Murphy, B.A. Blue light from light-emitting diodes directed at a single eye elicits a dose-dependent suppression of melatonin in horses. Vet. J. 2013, 196, 231–235. [Google Scholar] [CrossRef]
- Hoban, T.M.; Lewy, A.J.; Fuller, C.A. Light suppression of melatonin in the squirrel monkey (Saimiri sciureus). J. Pineal Res. 1990, 9, 13–19. [Google Scholar] [CrossRef]
- Phillips, A.J.K.; Vidafar, P.; Burns, A.C.; McGlashan, E.M.; Anderson, C.; Rajaratnam, S.M.W.; Lockley, S.W.; Cain, S.W. High sensitivity and interindividual variability in the response of the human circadian system to evening light. Proc. Natl. Acad. Sci. USA 2019, 116, 12019–12024. [Google Scholar] [CrossRef]
- Le Tallec, T.; Théry, M.; Perret, M. Melatonin concentrations and timing of seasonal reproduction in male mouse lemurs (Microcebus murinus) exposed to light pollution. J. Mammal. 2016, 97, 753–760. [Google Scholar] [CrossRef]
- Obayashi, K.; Saeki, K.; Iwamoto, J.; Okamoto, N.; Tomioka, K.; Nezu, S.; Ikada, Y.; Kurumatani, N. Exposure to light at night, nocturnal urinary melatonin excretion, and obesity/dyslipidemia in the elderly: A cross-sectional analysis of the HEIJO-KYO study. J. Clin. Endocrinol. Metab. 2013, 98, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Herljevic, M.; Middleton, B.; Thapan, K.; Skene, D.J. Light-induced melatonin suppression: Age-related reduction in response to short wavelength light. Exp. Gerontol. 2005, 40, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Bues, M.; Pross, A.; Stefani, O.; Frey, S.; Anders, D.; Späti, J.; Wirz-Justice, A.; Mager, R.; Cajochen, C. LED-backlit computer screens influence our biological clock and keep us more awake. J. Soc. Inf. Disp. 2012, 20, 266. [Google Scholar] [CrossRef]
- Chellappa, S.L.; Steiner, R.; Blattner, P.; Oelhafen, P.; Götz, T.; Cajochen, C. Non-visual effects of light on melatonin, alertness and cognitive performance: Can blue-enriched light keep us alert? PLoS ONE 2011, 6, e16429. [Google Scholar] [CrossRef] [PubMed]
- Figueiro, M.G.; Bullough, J.D.; Parsons, R.H.; Rea, M.S. Preliminary evidence for spectral opponency in the suppression of melatonin by light in humans. Neuroreport 2004, 15, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S.; Nakamura, K.; Honma, S.; Tokura, H.; Honma, K. Melatonin rhythm is not shifted by lights that suppress nocturnal melatonin in humans under entrainment. Am. J. Physiol. 1996, 270, R1073–R1077. [Google Scholar] [CrossRef]
- Heo, J.Y.; Kim, K.; Fava, M.; Mischoulon, D.; Papakostas, G.I.; Kim, M.J.; Kim, D.J.; Chang, K.A.J.; Oh, Y.; Yu, B.H.; et al. Effects of smartphone use with and without blue light at night in healthy adults: A randomized, double-blind, cross-over, placebo-controlled comparison. J. Psychiatr. Res. 2017, 87, 61–70. [Google Scholar] [CrossRef]
- Higuchi, S.; Fukuda, T.; Kozaki, T.; Takahashi, M.; Miura, N. Effectiveness of a red-visor cap for preventing light-induced melatonin suppression during simulated night work. J. Physiol. Anthropol. 2011, 30, 251–258. [Google Scholar] [CrossRef]
- Kozaki, T.; Koga, S.; Toda, N.; Noguchi, H.; Yasukouchi, A. Effects of short wavelength control in polychromatic light sources on nocturnal melatonin secretion. Neurosci. Lett. 2008, 439, 256–259. [Google Scholar] [CrossRef]
- Trinder, J.; Armstrong, S.M.; O’Brien, C.; Luke, D.; Martin, M.J. Inhibition of melatonin secretion onset by low levels of illumination. J. Sleep Res. 1996, 5, 77–82. [Google Scholar] [CrossRef]
- Van De Werken, M.; Giménez, M.C.; De Vries, B.; Beersma, D.G.M.; Gordijn, M.C.M. Short-wavelength attenuated polychromatic white light during work at night: Limited melatonin suppression without substantial decline of alertness. Chronobiol. Int. 2013, 30, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Figueiro, M.G.; Wood, B.; Plitnick, B.; Rea, M.S. The impact of light from computer monitors on melatonin levels in college students. Neuroendocrinol. Lett. 2011, 32, 158–163. [Google Scholar] [PubMed]
- Gooley, J.J.; Chamberlain, K.; Smith, K.A.; Khalsa, S.B.S.; Rajaratnam, S.M.W.; Van Reen, E.; Zeitzer, J.M.; Czeisler, C.A.; Lockley, S.W. Exposure to room light before bedtime suppresses melatonin onset and shortens melatonin duration in humans. J. Clin. Endocrinol. Metab. 2011, 96, E463–E472. [Google Scholar] [CrossRef] [PubMed]
- Vartanian, G.V.; Li, B.Y.; Chervenak, A.P.; Walch, O.J.; Pack, W.; Ala-Laurila, P.; Wong, K.Y. Melatonin suppression by light in humans is more sensitive than previously reported. J. Biol. Rhythms 2015, 30, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Wood, B.; Rea, M.S.; Plitnick, B.; Figueiro, M.G. Light level and duration of exposure determine the impact of self-luminous tablets on melatonin suppression. Appl. Ergon. 2013, 44, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Zeitzer, J.M.; Dijk, D.; Kronauer, R.E.; Brown, E.N.; Czeisler, C.A. Sensitivity of the human circadian pacemaker to nocturnal light: Melatonin phase resetting and suppression. J. Physiol. 2000, 526, 695–702. [Google Scholar] [CrossRef]
- Crowley, S.J.; Cain, S.W.; Burns, A.C.; Acebo, C.; Carskadon, M.A. Increased sensitivity of the circadian system to light in early/mid-puberty. J. Clin. Endocrinol. Metab. 2015, 100, 4067–4073. [Google Scholar] [CrossRef]
- Nagare, R.; Plitnick, B.; Figueiro, M.G. Effect of exposure duration and light spectra on nighttime melatonin suppression in adolescents and adults. Light Res. Technol 2019, 51, 530–543. [Google Scholar] [CrossRef]
- Nagare, R.; Rea, M.S.; Plitnick, B.; Figueiro, M.G. Nocturnal melatonin suppression by adolescents and adults for different levels, spectra, and durations of light exposure. J. Biol. Rhythms 2019, 34, 178–194. [Google Scholar] [CrossRef]
- Rüger, M.; Gordijn, M.C.M.; Beersma, D.G.M.; de Vries, B.; Daan, S. Weak relationships between suppression of melatonin and suppression of sleepiness/fatigue in response to light exposure. J. Sleep Res. 2005, 14, 221–227. [Google Scholar] [CrossRef]
- Chang, A.-M.; Aeschbach, D.; Duffy, J.F.; Czeisler, C.A. Evening use of light-emitting eReaders negatively affects sleep, circadian timing, and next-morning alertness. Proc. Natl. Acad. Sci. USA 2015, 112, 1232–1237. [Google Scholar] [CrossRef] [PubMed]
- Chinoy, E.D.; Duffy, J.F.; Czeisler, C.A. Unrestricted evening use of light-emitting tablet computers delays self-selected bedtime and disrupts circadian timing and alertness. Physiol. Rep. 2018, 6, e13692. [Google Scholar] [CrossRef] [PubMed]
- Kraneburg, A.; Franke, S.; Methling, R.; Griefahn, B. Effect of color temperature on melatonin production for illumination of working environments. Appl. Ergon. 2017, 58, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Münch, M.; Regente, J.; Bes, F.; De Zeeuw, J.; Hädel, S.; Wahnschaffe, A.; Kunz, D. Blue-enriched morning light as a countermeasure to light at the wrong time: Effects on cognition, sleepiness, sleep, and circadian phase. Neuropsychobiology 2017, 74, 207–218. [Google Scholar] [CrossRef]
- Nathan, P.J.; Wyndham, E.L.; Burrows, G.D.; Norman, T.R. The effect of gender on the melatonin suppression by light: A dose response relationship. J. Neural Transm. 2000, 107, 271–279. [Google Scholar] [CrossRef]
- Thapan, K.; Arendt, J.; Skene, D.J. An action spectrum for melatonin suppression: Evidence for a novel non-rod, non-cone photoreceptor system in humans. J. Physiol. 2001, 535, 261–267. [Google Scholar] [CrossRef]
- McIntyre, I.M.; Norman, T.R.; Burrows, G.D.; Armstrong, S.M. Quantal melatonin suppression by exposure to low intensity light in man. Life Sci. 1989, 45, 327–332. [Google Scholar] [CrossRef]
- Souman, J.L.; Borra, T.; de Goijer, I.; Schlangen, L.J.M.; Vlaskamp, B.N.S.; Lucassen, M.P. Spectral tuning of white light allows for strong reduction in melatonin suppression without changing illumination level or color temperature. J. Biol. Rhythms 2018, 33, 420–431. [Google Scholar] [CrossRef]
- Wahnschaffe, A.; Haedel, S.; Rodenbeck, A.; Stoll, C.; Rudolph, H.; Kozakov, R.; Schoepp, H.; Kunz, D. Out of the lab and into the bathroom: Evening short-term exposure to conventional light suppresses melatonin and increases alertness perception. Int. J. Mol. Sci. 2013, 14, 2573–2589. [Google Scholar] [CrossRef]
- Wright, H.R.; Lack, L.C. Effect of light wavelength on suppression and phase delay of the melatonin rhythm. Chronobiol. Int. 2001, 18, 801–808. [Google Scholar] [CrossRef]
- Whitmore, J.N.; French, J.; Fischer, J.R. Psychophysiological effects of a brief nocturnal light exposure. J. Hum. Ergol. 2001, 30, 267–272. [Google Scholar]
- Figueiro, M.G.; Rea, M.S.; Bullough, J.D. Circadian effectiveness of two polychromatic lights in suppressing human nocturnal melatonin. Neurosci. Lett. 2006, 406, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Nathan, P.J.; Burrows, G.D.; Norman, T.R. The effect of dim light on suppression of nocturnal melatonin in healthy women and men. J. Neural Transm. 1997, 104, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Zachmann, A.; Ali, M.A.; Falcón, J. Melatonin and its effects in fishes: An overview. In Rhythms in Fishes; Springer: Boston, MA, USA, 1992; pp. 149–165. [Google Scholar]
- Kummu, M.; de Moel, H.; Ward, P.J.; Varis, O. How close do we live to water? A global analysis of population distance to freshwater bodies. PLoS ONE 2011, 6, e20578. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Ortega, A.; López-Olmeda, J.F.; Vera, L.M.; Sánchez-Vázquez, F.J. Influence of constant light and darkness, light intensity, and light spectrum on plasma melatonin rhythms in Senegal sole. Chronobiol. Int. 2007, 24, 615–627. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Imamura, S.; Sawada, Y.; Hur, S.P.; Takemura, A. Effects of different colors of light on melatonin suppression and expression analysis of Aanat1 and melanopsin in the eye of a tropical damselfish. Gen. Comp. Endocrinol. 2014, 204, 158–165. [Google Scholar] [CrossRef]
- Carazo, I.; Norambuena, F.; Oliveira, C.; Sánchez-Vázquez, F.J.; Duncan, N.J. The effect of night illumination, red and infrared light, on locomotor activity, behaviour and melatonin of Senegalese sole (Solea senegalensis) broodstock. Physiol. Behav. 2013, 118, 201–207. [Google Scholar] [CrossRef]
- Oliveira, C.C.V.; Figueiredo, F.; Soares, F.; Pinto, W.; Dinis, M.T. Meagre’s melatonin profiles under captivity: Circadian rhythmicity and light sensitiveness. Fish. Physiol. Biochem. 2018, 44, 885–893. [Google Scholar] [CrossRef]
- Zubidat, A.E.; Nelson, R.J.; Haim, A. Spectral and duration sensitivity to light-at-night in ‘blind’ and sighted rodent species. J. Exp. Biol. 2011, 214, 3206–3217. [Google Scholar] [CrossRef]
- Dimovski, A.M.; Robert, K.A. Artificial light pollution: Shifting spectral wavelengths to mitigate physiological and health consequences in a nocturnal marsupial mammal. J. Exp. Zool. A Ecol. Integr. Physiol. 2018, 329, 497–505. [Google Scholar] [CrossRef]
- Reiter, R.J.; King, T.S.; Richardson, B.A.; Hurlbut, E.C. Studies on pineal melatonin levels in a diurnal species, the Eastern chipmunk (Tamias striatus): Effects of light at night, propranolol administration or superior cervical ganglionectomy. J. Neural Transm. 1982, 54, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Peters, J.F. Non-suppressibility by room light of pineal N-acetyltransferase activity and melatonin levels in two diurnally active rodents, the Mexican ground squirrel (Spermophilus mexicanus) and the eastern chipmunk (Tamias striatus). Endocr. Res. 1984, 10, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Reiter, M.N.; Hattori, A.; Yaga, K.; Herbert, D.C.; Barlow-Walden, L. The pineal melatonin rhythm and its regulation by light in a subterranean rodent, the valley pocket gopher (Thomomys bottae). J. Pineal Res. 1994, 16, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Haim, A.; Zubidat, A.E. Artificial light at night: Melatonin as a mediator between the environment and epigenome. Phil. Trans. R. Soc. B 2015, 370, 20140121. [Google Scholar] [CrossRef] [PubMed]
- Giudice, A.; Crispo, A.; Grimaldi, M.; Polo, A.; Bimonte, S.; Capunzo, M.; Amore, A.; D’Arena, G.; Cerino, P.; Budillon, A.; et al. The effect of light exposure at night (LAN) on carcinogenesis via decreased nocturnal melatonin synthesis. Molecules 2018, 23, 1308. [Google Scholar] [CrossRef] [PubMed]
- Wever, R. A The Circadian System of Man: Results of Experiments Under Temporal Isolation; Springer: New York, NY, USA, 1979. [Google Scholar]
- McGlashan, E.M.; Poudel, G.R.; Vidafar, P.; Drummond, S.P.A.; Cain, S.W. Imaging individual differences in the response of the human suprachiasmatic area to light. Front. Neurol. 2018, 9, 1022. [Google Scholar] [CrossRef]
- Prayag, A.S.; Najjar, R.P.; Gronfier, C. Melatonin suppression is exquisitely sensitive to light and primarily driven by melanopsin in humans. J. Pineal Res. 2019, 66, e12562. [Google Scholar] [CrossRef]
- Oliveira, C.; Duncan, N.J.; Pousão-Ferreira, P.; Mañanós, E.; Sánchez-Vázquez, F.J. Influence of the lunar cycle on plasma melatonin, vitellogenin and sex steroids rhythms in Senegal sole, Solea senegalensis. Aquaculture 2010, 306, 343–347. [Google Scholar] [CrossRef]
- Rea, M.; Figueiro, M. Light as a circadian stimulus for architectural lighting. Light Res. Technol. 2018, 50, 497–510. [Google Scholar] [CrossRef]
- Raap, T.; Casasole, G.; Costantini, D.; AbdElgawad, H.; Asard, H.; Pinxten, R.; Eens, M. Artificial light at night affects body mass but not oxidative status in free-living nestling songbirds: An experimental study. Sci. Rep. 2016, 6, 35626. [Google Scholar] [CrossRef]
- Johnsen, S. The Optics of Life: A Biologist’s Guide to Light in Nature; Princeton University Press: Princeton, NJ, USA, 2012. [Google Scholar]
- Lucas, R.J.; Peirson, S.; Berson, D.M.; Brown, T.M.; Cooper, H.M.; Czeisler, C.A.; Figueiro, M.G.; Gamlin, P.D.; Lockley, S.W.; O’Hagan, J.B.; et al. Measuring and using light in the melanopsin age. Trends Neurosci. 2014, 37, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jechow, A.; Hölker, F. How dark is a river? Artificial light at night in aquatic systems and the need for comprehensive night-time light measurements. WIREs Water 2019, e1388. [Google Scholar] [CrossRef]
- Smith, R.C.; Wilson, W.H. Photon scalar irradiance. Appl. Opt. 1972, 11, 934–938. [Google Scholar] [CrossRef] [PubMed]
- Thimijan, R.; Heins, R. Photometric, radiometric, and quantum light units of measure: A review of procedures for interconversion. Hortscience 1983, 18, 818–822. [Google Scholar]
- Jechow, A.; Hölker, F.; Kyba, C.C.M. Using all-sky differential photometry to investigate how nocturnal clouds darken the night sky in rural areas. Sci. Rep. 2019, 9, 1391. [Google Scholar] [CrossRef] [PubMed]
- Kocifaj, M.; Posch, T.; Solano Lamphar, H.A. On the relation between zenith sky brightness and horizontal illuminance. Mon. Not. R. Astron. Soc. 2015, 446, 2895–2901. [Google Scholar] [CrossRef]
- Mobley, C.D. Light and Water: Radiative Transfer in Natural Waters; Academic Press: San Diego, CA, USA, 1994. [Google Scholar]
- Hölker, F.; Jechow, A.; Schroer, S.; Gessner, M.O. Nächtliches Licht und Lichtverschmutzung in und um Gewässer. In Handbuch Angewandte Limnologie: Grundlagen-Gewässerbelastung-Restaurierung-Aquatische Ökotoxikologie-Bewertung-Gewässerschutz; WILEY-VCH Verlag GmbH & Co.: KGaA, Weinheim, 2018; pp. 1–26. ISBN 978-3-527-67848-8. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parapineal Organ | Pineal Organ | Deep-Brain Photoreceptors | Retina (Including Melanopsin) | |
|---|---|---|---|---|
| Fishes | + | + | + | + |
| Amphibians | + | + | + | + |
| Reptiles | + | + | + | + |
| Birds | − | + | + | + |
| Mammals | − | − | − | + |
| Fishes | Amphibians | Reptiles | Birds | Mammals (Rodents and Ungulates) | Mammals (Primates) | Mammals (Humans) | |
|---|---|---|---|---|---|---|---|
| Number of studies identified in the keyword search in Scopus | 357 | 56 | 15 | 155 | 125 | 17 | 1145 |
| Number of studies identified in the keyword search in Web of Science | 367 | 34 | 32 | 169 | 133 | 25 | 733 |
| Number of studies included in the review | 15 | 0 | 0 | 10 | 11 | 2 | 34 |
| (3A) Plasma melatonin at exposure to nocturnal white light in fishes | ||||||||
| Species | Taxonomic family | Temperature | Light intensities | Light color/light type | Control | Methodology and timing of sampling | Melatonin levels relative to control | Reference |
| European sea bass (Dicentrarchus labrax) | Moranidae | 23 °C | 1.4 lx (0.6 µW/cm2) 5.7 lx (2.4 µW/cm2) 14.2 lx (6 µW/cm2) 141.8 lx (60 µW/cm2) | White | Dark | Plasma sample N/A | −12.5% −25% −75% * −83% * | (Bayarri et al., 2002) [62] |
| Humbug damselfish (Dascyllus aruanus) | Pomacentridae | N/A | First-quarter moon Full moon (0.3 lx) Last-quarter moon | Moonlight | New moon | Plasma sample N/A | −29% * −55% * −35% * | (Choi et al., 2017) [56] |
| Mozambique tilapia (Oreochromis mossambicus) | Cichlidae | N/A | New moon Full moon 0.1 lx 1 lx | Moonlight White | Dark | Plasma sample 01:00 a.m. | −25% * −56% * −33% * −31% * | (Nikaido et al., 2009) [55] |
| Seagrass rabbitfish (Signaus canaliculatus) | Siganidae | N/A | First-quarter moon Full moon Last-quarter moon 12 lx | Moonlight White | New moon Dark | Plasma sample N/A | −28% N/A −59% N/A −7% N/A −62% * (mean) | (Rahman et al., 2004) [57] |
| Goldlined spinefoot (Signaus guttatus) | Siganidae | N/A for sampling (19.8–30.9 °C) | Full moon | Moonlight | New moon | Plasma sample N/A | −41% * | (Park et al., 2014) [60] |
| Arctic charr (Salvelinus alpinus) | Salmonidae | 10 °C | 0.1–0.3 lx (mid-water) 50–65 lx (mid-water) | White | Dark | Plasma sample 10:00 p.m. | −57% * −74% * | (Liu et al., 2019) [59] |
| Atlantic salmon (Salmo salar) | Salmonidae | N/A for sampling (1–17 °C) | 1 lx 20 lx 100 lx | White | Dark | Plasma sample 9:00 p.m. | −11% N/A −23% N/A −30% N/A | (Porter et al., 2001) [61] |
| Tench (Tinca tinca) | Cyprinidae | N/A | 0.3 lx (3.3 µW/cm2) 1 lx (5.3 µW/cm2) 3 lx (10.5 µW/cm2) | White | Dark | Plasma sample N/A | −67% * −65% * −67% * | (Vera et al., 2005) [54] |
| Roach (Rutilus rutilus) | Cyprinidae | N/A | 15 lx | White | Half-moon (0.02 lx) | Plasma sample N/A | +14% for F +900% for M | (Brüning et al., 2018) [63] |
| Roach (Rutilus rutilus) | Cyprinidae | 15 °C | 1 lx 10 lx 100 lx | White | Dark | Water-based measurement 8:00 a.m. | −97% * −97% * −99% * | (Brüning et al., 2018) [65] |
| Eurasian perch (Perca fluviatilis) | Percidae | 16 °C | 1 lx 10 lx 100 lx | White | Dark | Water-based measurement 5:00 a.m. | −73% */79 % N/A (rel) −79% */75 % N/A (rel) −84% */82% N/A (rel) | (Brüning et al., 2015) [64] |
| Eurasian perch (Perca fluviatilis) | Percidae | N/A | 15 lx | White | Half-moon (0.02 lx) | Plasma sample N/A | 0% | (Brüning et al., 2018) [63] |
| Eurasian perch (Perca fluviatilis) | Percidae | 16 °C | 0.01 lx 0.1 lx 1 lx | White | Dark | Water-based measurement 5:00 a.m. | −48% */−15% (rel) −57% */−21% * (rel) −78% */−49% * (rel) | Kupprat et al. under revision [66] |
| (3B) Plasma melatonin at exposure to nocturnal colored light in fishes | ||||||||
| Species | Taxonomic family | Temperature | Light intensities | Light color/light type | Control | Methodology | Melatonin levels relative to control | Reference |
| European sea bass (Dicentrarchus labrax) | Moranidae | 23 °C | 2 lx (2.4 µW/cm2) 11 lx (2.4 µW/cm2) 3 lx (2.4 µW/cm2) 5 lx (6 µW/cm2) 28 lx (6 µW/cm2) 7 lx (6 µW/cm2) | Blue Green Red Blue Green Red | Dark | Plasma sample N/A | −55% * −58% * −14% −84% * −56% * −78% * | (Bayarri et al., 2002) [62] |
| Roach (Rutilus rutilus) | Cyprinidae | 15 °C | 0.15 lx (5.6 mW/m2) 2.2 lx (4.8 mW/m2) 0.65 lx (3.9 mW/m2) (all 0.021 µmol/s/m2) | Blue Green Red | Dark | Water-based measurement 8:00 a.m. | −95% * −95% * −95% * | (Brüning et al., 2018) [65] |
| Eurasian perch (Perca fluviatilis) | Percidae | 16 °C | 0.15 lx (5.6 mW/m2) 2.2 lx (4.8 mW/m2) 0.65 lx (3.9 mW/m2) (all 0.021 µmol/s/m2) | Blue Green Red | Dark | Water-based measurement 08:00 a.m. | −68% */−10% N/A (rel) −84% */−50% N/A (rel) −80% */−33% N/A (rel) | (Brüning et al., 2016) [67] |
| (3C) Ocular melatonin at exposure to nocturnal white light in fishes | ||||||||
| Species | Taxonomic family | Temperature | Light intensities | Light color/light type | Control | Methodology | Melatonin levels relative to control | Reference |
| European sea bass (Dicentrarchus labrax) | Moranidae | 23 °C | 1.4 lx (0.6 µW/cm2) 6 lx (2.4 µW/cm2) 14 lx (6 µW/cm2) 140 lx (60 µW/cm2) | White | Dark | Ocular sample N/A | +40% +76% * +152% * 0% | (Bayarri et al., 2002) [62] |
| Wrasse (Halichaoeres tenuispinnis) | Labridae | 25 °C | 0.13 lx 1.3 lx 13 lx 130 lx | White | Dark | Ocular sample N/A | −24% −65% * −66% * −74% * | (Iigo et al., 2003) [69] |
| Seagrass rabbitfish (Signaus canaliculatus) | Siganidae | N/A | First-quarter moon | Moonlight | New moon | Ocular sample N/A | −34% N/A | (Rahman et al., 2004) [58] |
| Full moon | −42% N/A | |||||||
| Last-quarter moon | −16% N/A | |||||||
| 12 lx | White | Dark | −49% * (mean) | |||||
| Full moon | Moonlight | New moon | sgAanat1 expression (Ocular sample) N/A | −66% * (continuous) | ||||
| Goldlineds spinefoot (Signaus guttatus) | Siganidae | 25 °C | −58% (before pulse, 12:00 p.m.) | (Kashiwagi et al., 2013) [68] | ||||
| Full moon | Moonlight | Dark | −72% * (2-h pulse) | |||||
| (3D) Ocular melatonin at exposure to nocturnal colored light in fishes | ||||||||
| Species | Taxonomic family | Temperature | Light intensities | Light color/light type | Control | Methodology | Melatonin levels relative to control | Reference |
| European sea bass (Dicentrarchus labrax) | Moranidae | 23 °C | 2 lx (2.4 µW/cm2) 11 lx (2.4 µW/cm2) 3 lx (2.4 µW/cm2) 5 lx (6 µW/cm2) 28 lx (6 µW/cm2) 7 lx (6 µW/cm2) | Blue Green Red Blue Green Red | Dark | Ocular sample N/A | +717% * +400% * +567% * +1733% * +650% * +233% | (Bayarri et al., 2002) [62] |
| Species | Taxonomic Family | Temperature | Light Intensities and Duration | Light Color/Light Type | Control | Methodology | Melatonin Relative to Control | Reference |
|---|---|---|---|---|---|---|---|---|
| Great tit (Parus major) | Paridae | 10–14 °C | 0.15, 0.5, 1.5, 5 lx for entire night | Warm white LED | 0.05 lx | Plasma sample | −50% under 5 lx * | (de Jong et al., 2016) [15] |
| Eurasian blackbird (Turdus merula) | Turdidae | 20 °C | 0.3 lx for entire night | Incandescent light bulb | 0.001 lx | Plasma sample | ~−50% winter (dusk and dawn) * ~−30% summer (dusk, midnight, dawn) * | (Dominoni et al., 2013) [96] |
| Eurasian tree sparrow (Passer montanus) | Passeridae | 22 °C | 8 lx for entire night | Cool white fluorescent lamp | 0 lx | Plasma sample | −70% at midnight * | (Jiang et al., 2020) [99] |
| Indian weaver bird (Ploceus philippinus) | Ploceidae | 24 °C | 2 lx for entire night or 4 h | Cool white fluorescent lamp | 0.1 lx | Plasma sample | −66% at midnight * −50% at dawn and dusk * | (Kumar et al., 2018) [98] |
| Western scrub-jay (Aphelocoma californica) | Corvidae | 20 °C | 0.3 lx for entire night | Incandescent light bulb | 0.01 lx | Plasma sample | +50% * | (Schoech et al., 2013) [97] |
| (5A) Melatonin at exposure to nocturnal light in mammals: rodents | ||||||||
| Species | Number, gender, and age of subjects | Light intensity/treatments | Light duration | CCT, wavelength, light color/light type | Control light intensities | Methodology | Melatonin relative to control | Reference |
| Sprague–Dawley rats | n = 12 M adults (3–4 weeks) | 8.07 ± 0.95 lx (3.31 ± 0.38 μW/cm2) | Overnight (12-h light/12-h dark), over 4 weeks | >620 nm, red light | 0 lx | Plasma MEL | ~−95% for peak dark-phase MEL | (Dauchy et al., 2015) [106] |
| Siberian hamsters (Phodopus sungorus) | n = 6–8 M adults | 1. 0.003 μW/cm2 2. 0.03 μW/cm2 3. 0.3 μW/cm2 4. 1.31 μW/cm2 5. 4.8 μW/cm2 6. 68 μW/cm2 | 15-min light pulse, 60% into the scotophase, LP: 14-h light/10-h dark SP: 10-h light/14-h dark | 480 nm LED | 0 lx | Plasma MEL | MEL SUP similar for both photoperiods: 1. NS 2. ~−60% * 3. ~−90% * 4. ~−90% * 5. ~−90% * 6. ~−90% * | (Glickman et al., 2014) [109] |
| Siberian hamsters (Phodopus sungorus) | n = 132–156 M adults | SP (8-h light/16-h dark) + light pulse (200–800 lx) | 1-min light pulse, in the middle of the scotophase, 3–4 weeks | Fluorescent light | 1. LP (16-h light/8-h dark) 2. SP (8-h light/16-h dark) Only | Pineal N-acetyltransferase (MEL formation) | Treatment to CON 1: NS Treatment to CON 2: 50% shorter enzyme activity time under short photoperiod only | (Hoffmann et al., 1981) |
| Laboratory mice | n = 10–14 M adults | 200 lx | 15-min light pulse: 1. 4 h into scotophase, 2. 2 h before end of scotophase | N/A | 0 lx | Plasma and pineal MEL | Similar MEL SUP after light pulse at both times: ~−85% * pineal MEL ~−70% * plasma MEL 1: delay of morning decline of MEL production 2: advance of morning decline of MEL production (>2 h) | (Kennaway et al., 2002) [105] |
| Fat sand rat (Psammomys obesus) | n = 8 M N/A | 1. Acute light pulse 2. SP (8-h light/16-h dark) 2. SP + chronic light pulse 4. Constant dim blue light (30 lx, 470 nm) | Acute light pulse: 30 min, 5 h into the scotophase Chronic light pulse: 3 × 15 min at 4-h intervals Constant light: over 24 h for 3 weeks | Cool white fluorescent | Control photoperiod 12-h light/12-h dark, no light at night | aMT6s in urine | 1: −63% * 2: −73% * (6 h into scotophase) 3: +71−89% * compared to SP only 4: +71% * | (Schwimmer et al., 2010) [111] |
| Rats | n = 3−6 F 5 weeks (50 ± 3 days) | 61, 81, 128, 193 lx | 1. 5× 1-min light pulse, 2 h into scotophase, 1 day 2. 2 weeks 3. 10 weeks | Incandescent light | 12-h light/12-h dark, dim red light at night (λ ≥ 650 nm), <0.1 lx | Serum MEL | 1: ~−65% * 2: ~−35% * (high sensitivity mid-scotophase ~−39%), phase advance 3. ~−25% *, stronger phase advance | (Travlos et al., 2001) [107] |
| Nude rats | n = 12 F 1–2 weeks | 0.2 lx dim light | Throughout the scotophase (12-h light/12-h dark) | 0 lx | Plasma MEL | ~−88% * for peak dark-phase MEL | (Xiang et al., 2015) [108] | |
| (5B) Melatonin at exposure to nocturnal light in mammals: ungulates | ||||||||
| Species | Number, gender, and age of subjects | Light intensity/treatments | Light duration | CCT, wavelength, light color/light type | Control light intensities | Methodology | Melatonin relative to control | Reference |
| Israeli Holstein cows | n = 14 F mean age 3.85 years | 105 ± 3.91 lx | Overnight | ALAN: 482 ± 5.12 nm Control: 648 ± 5.12 nm | 5.08 ± 0.04 lx | MEL in milk | −44% * in night-time sample −38.4 * in daytime sample | (Asher et al., 2015) [112] |
| Goats | n = 6 F 2 years | 1. 0.87 ± 0.14 lx (0.26 ± 0.004 µW/cm2) 2. 2.3 ± 0.3 lx (0.68 ± 0.09 µW/cm2) 3. 14.2 ± 2.1 lx (4.22 ± 0.62 µW/cm2) 4. 137 ± 14 lx (40.70 ± 4.16 µW/cm2) | 1 h, 3 h into scotophase (8-h light/16-h dark). Increasing intensity treatments, 1–3 days between the treatments | Cool white fluorescent lamps | 0 lx | Plasma MEL | 1: 0% 2. −43.1% * 3: −71.1% * 4: −81.2% * | (Deveson et al. 1990) [113] |
| Sheep | n = 6–7 F N/A | N/A | 1 h at midnight | N/A | Dim red light (<1 lx) | Plasma MEL | ~−92% * for the high-MEL group ~−88% * for the low-MEL group | (Vivien-Roels et al. 1999) [114] |
| Thoroughbred horse (Equus caballus) | n = 4 M 5 years | 3, 10, 50, 100 | 1 h | Blue LED, 468 nm | <0.1 lx | Serum MEL | 3 lx: NS 10 lx: ~−60% * 50 lx: ~−80% * 100 lx: ~−70% * | (Walsh et al. 2013) [115] |
| (5C) Melatonin at exposure to nocturnal light in mammals: primates | ||||||||
| Species | Number, gender, and age of subjects | Light intensity/treatments | Light duration and timing | CCT, wavelength, light color/light type | Control light intensities | Methodology | Melatonin relative to control | Reference |
| Mouse lemur (Microcebus murinus) | n = 12 M adults | 51.5 lx | 3–5 h, 5 weeks during the nights | White LED Yellow LED | ~0.3 lx | aMT6s in urine | −50% * | (Le Tallec, Théry, and Perret 2016) [118] |
| Squirrel monkey (Saimiri sciureus) | n = 9 M adults | 200 lx | 2 h (12:00–02:00 a.m.) | Fluorescent light | No light at night | MEL in plasma, aMT6s in urine | −4.8% * | (Hoban, Lewy, and Fuller 1990) [116] |
| Number, Gender (M, F), Age of Subjects | Light Intensity/Light Type | Duration and Timing of Light Exposure | CCT, Wavelength, Color of Light | Control Lighting Conditions | Location | Methodology | MEL Relative to CON | Reference |
|---|---|---|---|---|---|---|---|---|
| n = 13 M = 13 mean age 23.8 ± 5.0 years | ~75 lx | 5 h (7:30 p.m.–12:30 a.m.) | 1. LED screen—6953 K 2. No LED screen—4775 K | <8 lx | Laboratory | MEL in saliva | SIGN effects for screen, time of day, interaction screen vs. time of day | (Bues et al. 2012) [121] |
| n = 12 M = 6, F = 6 mean age 24.92 ± 2.87 years | Printed book 0.91 lx LED ebook 31.73 lx | 4 h before bed time every day (6:00–10:00 p.m.) 5 days each session | N/A | ~3 lx ambient room light level | Laboratory | MEL in plasma | LED ebook: −55.12 ± 20.12% * Printed book: 18.77 ± 39.57% NS between sessions * | (Chang et al. 2015) [139] |
| n = 16 M = 16 (20–28 years) | 40 lx | 2 h (9:30–11:30 p.m.) | Fluorescent lamps: 6500 K 2500 K incandescent lamp: 3000 K | 0 lx | Laboratory | MEL in saliva | 6500 K: * MEL increase to pre-light exposure levels: 6500 K: 29.5 ± 5% 3000 K: 49 ± 7.6% 2500 K: 42 ± 8.6% | (Chellappa et al. 2011) [122] |
| n = 9 M = 6; F = 3 mean age 25.7 ± 3.0 years | LED tablet: 38.4 ± 23.2 lx (min–max: 1.5–148.2 lx) | 2× 5 consecutive evenings (6:00–8:45 p.m.) | N/A | Printed book: 0.7 ± 0.2 lx (min–max: 0.2–1.4 lx) | Laboratory | MEL in plasma | LED tablet: 54.17 ± 18%; printed: 9.75 ± 22.75%; LED tablet delayed bedtime for 30 min, MEL secretion for ~1 h and sleep onset for 30 min | (Chinoy, Duffy, and Czeisler 2018) [140] |
| n = 67 M = 38; F = 29; pre- to mid-pubertal group (9.1–14.7 years); late- to post-pubertal group (11.5–15.9 years) | 15 lx 150 lx 500 lx | 4 consecutive nights 1 h evening group (11:00 p.m.–12:00 a.m.) morning group (3:00–4:00 a.m.) | N/A | 0.1 lx | Laboratory | MEL in saliva | Evening light exposure: pre- to mid-pubertal group MEL SUP: 15 lx: −9.2 ± 20.5% 150 lx: −26 ± 17.7% 500 lx: −36.9 ± 11.4% late- to post-pubertal group MEL SUP 15 lx: −5.3 ± 17.7% 150 lx: −12.5 ± 17.3% 500 lx: −23.9 ± 21.7% Evening light exposure between age groups: SIG Morning light exposure between age groups: NS | (Crowley et al. 2015) [135] |
| n = 4 M = 4 (20–21 years) | 1. 18 lx (29 μW/cm2) for blue LED 2. 450 lx (170 μW/cm2) for mercury vapor | 1 h (1:00–2:00 a.m. and 5:00–6:00 a.m.) | Blue LED, mercury vapor lamp | Dimly illuminated area | Laboratory | MEL in plasma | 1: −34 ± 9% * 2: −18 ± 6% * | (Figueiro, et al. 2004) [123] |
| n = 40 M = 22 (18–54 years, median = 21 years) F = 18 (18–35 years, median = 23 years) | 1. 30 lx 2. 100 lx 3. 300 lx 4. 1000 lx (at the cornea) | 40 min, 2 experimental sessions (2:00–2:40 a.m., 3:40–4:20 a.m.) | 4100 K 8000 K | Dim light (red LED) | Laboratory | MEL in plasma | SIGN effect of light level and spectrum 4100 K: 1: 3 ± 11% 2: 10 ± 4% 3: 38 ± 7% 4: 38 ± 6% 8000 K: 1: 10 ± 8% 2: 32 ± 7% 3: 47 ± 4% 4: 34 ± 9% | (Figueiro, Rea, and Bullough 2006) [150] |
| n = 21 mean age 28.0 ± 9.9 years | 1. Computer monitor 7 lx 2. Computer monitor 7 lx + 40 lx blue safety glasses (peak λ ~470 nm) | 2 h (11:00 p.m.–1:00 a.m.) | N/A | Computer monitor 7 lx + orange-tinted glasses (<525 nm ~0) | Laboratory | MEL in saliva | 1: −11% NS 2: −30% * | (Figueiro et al. 2011) [130] |
| n = 116 study 1 n = 104, study 2 n = 12 (18–30 years) | <200 lx | overnight illumination Study 1: 9–10 days Study 2: 14 days | 4100 K | ˂3 lx | Laboratory | MEL in plasma | Exposure during sleep: ~−50% in 85% trial exposure before bedtime: SUP MEL, later MEL onset in 99.0% of individuals and shortened MEL duration by ~90 min | (Gooley et al. 2011) [131] |
| n = 9 M = 9 mean age 22.11 ± 0.69 years | 200 lx 500 lx 1000 lx 5000 lx 10,000 lx | 3 h (6:00–9:00 a.m.) | N/A | ˂200 lx | Laboratory | MEL in plasma | 200 lx NS ˃500 lx *, first hour of exposure ~35–50% | (Hashimoto et al. 1996) [124] |
| n = 22 M=22 mean age 30.95 ± 4.15 years | 2 types of smartphones: LED with blue light, LED with suppressed blue light | 150 min (7:30–10:00 p.m.) | N/A | <3 lx | Laboratory | MEL in blood serum | LED with blue light: lower levels and later onset of MEL secretion than LED with SUP blue light MEL between groups: NS | (Heo et al. 2017) [125] |
| n = 34, F young (n = 13), mean age 24 ± 3 years F postmenopausal (n = 21), mean age 57 ± 5 years | 456 nm: ~11.4 lx (3.8 μW/cm2) 29.4 lx (9.8 μW/cm2) 548 nm: 84 lx (28 μW/cm2) 186 lx (62 μW/cm2) | 30 min during the rising phase of melatonin synthesis | Monochromatic light: 456 nm 548 nm | 0 lx | Laboratory | MEL in plasma | 456 nm, postmenopausal women: ~11.4 lx: −1.4 ± 5% NS 29.4 lx: −12.5 ± 4% NS Young women ~11.4 lx: −18.4 ± 4.8% * 29.4 lx: −25.5 ± 6.4% * 548 nm: SIGN SUP in both age group, NS difference between age groups | (Herljevic et al. 2005) [120] |
| n = 11 M = 11 mean age 21.2 ± 0.9 years | 1. Non-visor: 500 lx 2. red visor: 160 lx 3. blue visor: 160 lx | 2–4 h (11:00 p.m.–3:00 a.m.) | 4200 K | ˂15 lx | Laboratory | MEL in saliva | 2-h exposure: 1: −57.0 ± 26.6% 2: -20.3 ± 32.1% * to non-visor 3: −31.1 ± 41.5% 4 h exposure: 1: −52.6 ± 22.4% 2: −7.7 ± 3.3% * 3: −3 ± 67.6% * | (Higuchi et al. 2011) [126] |
| n = 12 M = 12 mean age 21.3 ± 1.1 years | 200 lx | 1.5 h (1:00–2:30 a.m.) | Fluorescent lamps: 2300 K 3000 K 5000 K | <10 lx | Laboratory | MEL in saliva | 5000 K: * 3000 K: * 5000 K more than 3000 K 2300 K: NS | (Kozaki et al. 2008) [127] |
| (Experiment 2) n = 16 M = 9, F = 7 (19–28 years) | 200 lx | 4 h (10:00 p.m.–2:00 a.m.) | L1—1600 K L2—1950 K L3—2750 K L4—3900 KL5—6100 K L6—7100 K L7—14,000 K | <0.1 lx | Laboratory | MEL in saliva | Small MEL SUP under CCT <2000 K and strong MEL SUP with CCT ≥3900 K AUC under L4–L7 were SIGN lower than under L1 and AUCs under L3-L7 were SIGN smaller than under L2 | (Kraneburg et al. 2017) [141] |
| n = 6 M = 4, F = 2 mean age 32.3 + 6.2 years | 200 lx 400 lx 600 lx | 3 h (12:00–03:00 a.m.) | N/A | ˂10 lx | Laboratory | MEL in plasma | 200 lx: −21% NS 400 lx: −50% * 600 lx: −65% * | (McIntyre et al. 1989) [145] |
| n = 18 M = 6; F = 12; mean age 23.2 ± 3.3 years | Evening light: orange (OL) ~100 lx; blue-enriched (BL) ~500 lx Morning light: mixed blue-enriched (MBL) 750 lx; control (CON) 40 lx | EL: 30 min (10:00–10:30 p.m.) ML: 3 h (8:00–11:00 a.m.) | OL—1500 K BL—6500 K DL—2700 K MBL—3,537 K CON—2600 K | Dim light (DL) <7 lx | Laboratory and home settings | MEL in saliva | Evening light: BL to D: * OL to DL: * OL to BL: * Phase shifts greater under CON than MBL, phase delays after 3 days: MBL to CON * phase advances: NS | (Münch et al. 2017) [142] |
| n = 24 12 adolescents: M = 3, F = 9 mean age 16.5 ± 1.9 years 12 adults: M = 2, F = 10 mean age 46 ± 5.2 years | 295 lx for 2700 K 209 lx for 5600 K (CS at the eye level = 0.25) | 4 h (11:00 p.m.–3:00 a.m.) | Blue light, 2700 K 5600 K | <5 lx | Laboratory | MEL in saliva | Greater MEL SUP for longer exposure duration, SIGN interaction between light spectrum and age Adolescents: 5600 K: −43 ± 4% * 2700 K: −29 ± 4% * Adults: 5600 K: −33% ± 3% * 2700 K: −33% ± 2% * | (Nagare, Plitnick and Figueiro 2019) [136] |
| n = 41 18 adolescents (13–18 years) 23 adults (24–55 years) | 40–1000 lx (CS = 0.07, 0.14, 0.30, 0.50) | 0.5–3.0 h (11:00 p.m.–2:00 a.m.) | White light, 2700K 6500 K | <5 lx | Laboratory | MEL in saliva | SIGN effects of light intensity, spectrum, exposure duration; effect of light diminishes with increasing exposure duration for age groups and spectra. 6500 K: −24.7 ± 1% * 2700 K: −18.4 ± 1% * | (Nagare et al. 2019) [137] |
| n = 54, study 1: M = 22 (19–34 years), F = 21 (20–56 years) study 2: M = 4 (27–58 years), F = 7 (25–51 years) | Study 1: 200 lx study 2: 500 lx | 1h (12:00–1:00 a.m.) | 5000 K | 10–20 lx | Laboratory | MEL in plasma | Study 1: M: 16,9% F: 14,5% study 2: M: 35% F: 45% NS differences between the sexes | (Nathan, Burrows, and Norman 1997) [151] |
| n = 10 M = 5, F = 5 mean age 25.0 ± 2.5 years | 200 lx 500 lx 1000 lx 3000 lx | 1h (12:00–1:00 a.m.) | 5000 K | 10–20 lx | Laboratory | MEL in plasma | 200 lx: −17% NS 500 lx: −40% * 1000 lx: −56% * 3000 lx: −74% * No differences between the sexes | (Nathan et al. 2000) [143] |
| n = 528; dark LAN group n = 383 LAN group n = 145 Seniors (min 60 years) mean age 72.8 years | LAN group ≥ 3 lx | Continuous overnight exposure | N/A | Dark LAN <3 lx | Home settings | aMT6s in urine | LAN 6.8 g (4.3–10.5); Dark LAN group: 6.5 g (3.9–9.5); NS | (Obayashi et al. 2013) [119] |
| n = 56 M = 27, F = 29 mean age 20.8 ± 2.6 years | 10, 30, 50, 100, 200, 400, and 2000 lx | 5 h (4 h before sleep and 1 h after sleep start) | 4100 K | <1 lx | Laboratory | MEL in saliva | −50% * at group level ~24.6 lx; high variability for individuals: −50% * from 6 to 350 lx | (Phillips et. al. 2019) [117] |
| n = 36, each experiment n = 12 M = 29, F = 7 mean age 21.91 ± 2.03 years | 100 lx, partial illumination of the retina | 4 h (12:00–4:00 a.m.) | N/A | <10 lx | Laboratory | MEL in saliva | −22% * after temporal and −58% * after nasal exposure | (Rüger et al. 2005) [138] |
| n = 16 M = 9; F = 7 mean age 36.2 ± 8.6 years | ~175 lx | 3 h 2 h before and 1 h after habitual bedtime | White light, similar CCT 2700 K 2 spectral powers: low 52.03 µW/cm2 high 90.85 µW/cm2 | <5 lx | Laboratory | MEL in saliva | High MEF: ~−50% * low MEF: NS | (Souman et al. 2018) [146] |
| n = 22 M = 18, F = 4 mean age 27.0 ± 7.0 years | 0.70–65 µW/cm2 | 30 min (11:30 p.m.–2:30 a.m.) CT 16–18 | Monochromatic light, λmax 424, 456, 472, 496, 520, and 548 nm | No light exposure | Laboratory | MEL in plasma | MEL concentrations SUP in a dose-dependent manner. No SIGN MEL SUP at the lowest irradiances (0.70–3.3 µW/cm2) of 520 nm | (Thapan, Arendt, and Skene 2001) [144] |
| n = 6 M = 6 (18–22 years, mean age 20.2 years) | 250 lx 500 lx 1000 lx 2500 lx | 3 h individually determined 1h before MEL onset | N/A | <50 lx | Laboratory | MEL in plasma | Complete MEL SUPPR compared to CON in all treatments | (Trinder et al. 1996) [128] |
| n = 33 M = 33 study 1 n = 17 study 2 n = 16 (22.6 ± 2.2 years) | 1. Short-wavelength attenuated polychromatic white light (SWL): 193 lx 2. Full-spectrum light (FWL): 256 lx | 3 nights, overnight exposure (11:00 p.m.–7:00 a.m.) | 3000 K | 3 lx | Laboratory and home settings | MEL in saliva aMT6s in urine | SWL: −6 ± 4% NS FWL: −45 ± 6% * SWA to CON * | (Van De Werken et al. 2013) [129] |
| n = 6 (19–37 years) | 8.1 log photons/cm2·s 9.2 log photons/cm2·s 10.3 log photons/cm2·s | 1 h (10:00–11:00 p.m.) | 460 nm | Complete darkness | Laboratory | MEL in saliva | Threshold for human melatonin suppression ~10 log photons/cm2·s at 460 nm | (Vartanian et al. 2015) [132] |
| n = 9 M = 6; F = 3 mean age 26.3 ± 4.2 (22–33 years) | 1. 130 lx 2. 130 lx 3. 500 lx 4. 500 lx 5. 500 lx | 30 min (10:30–11:00 p.m.) | 1. 2000 K 2. 6000 K 3. 6000 K 4. 5000 K 5. 2800 K | <10 lx | Laboratory | MEL in saliva | 1: NS 2–5: * | (Wahnschaffe et al. 2013) [147] |
| n = 10 M = 6, F = 4 18–41 years | 500–1000 lx full spectrum 500–1000 lx green light | 1 h (2:00–3:00 a.m.) | N/A | 20–50 lx full spectrum (dim) | Laboratory | MEL in saliva | All light treatments SIGN different from CON. None of the treatment conditions SIGN different from each other | (Whitmore, French, and Fischer 2001) [149] |
| n = 13 mean age 18.9 ± 5.2 years | Tablet + blue LED 59 ± 5.0 lx tablet only 18 ± 3. 8 lx | 1–2 h (11:00 p.m.–1:00 a.m.) 3 weekends, 1 week apart | blue light λmax ~470 nm | Tablet + orange-tinted glasses 9.8 ± 1.9 used as control | Laboratory | MEL in saliva | 1 h exposure: Tablet + blue LED: −48 ± 4% * Tablet only: −7 ± 4% NS 2 h exposure: Tablet + blue LED: −66 ± 4% * Tablet only: −23 ± 6% * | (Wood et al. 2013) [133] |
| n = 15 M = 5, F = 10 mean age 31.8 ± 11.9 years | 130 µW/cm2 (received by each eye) | 2 h (12:00–2:00 a.m.) | 1. 660 nm 2. 595 nm 3. 525 nm 4. 497 nm 5. 470 nm | No light | Laboratory | MEL in saliva | 1: −5% NS 2: −10% NS 3: −70% * 4: −80% * 5: −65% * | (Wright and Lack 2001) [148] |
| n = 23 mean age 27.8 ± 8.91 (18–44 years) | 3 lx 106 lx 9100 lx | 6–5-h pulse | Cool light | Dim light | Laboratory | MEL in plasma | 3 lx: −11% N/A 106 lx: −88% N/A 9100 lx: −98% N/A | (Zeitzer et al. 2000) [134] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grubisic, M.; Haim, A.; Bhusal, P.; Dominoni, D.M.; Gabriel, K.M.A.; Jechow, A.; Kupprat, F.; Lerner, A.; Marchant, P.; Riley, W.; et al. Light Pollution, Circadian Photoreception, and Melatonin in Vertebrates. Sustainability 2019, 11, 6400. https://doi.org/10.3390/su11226400
Grubisic M, Haim A, Bhusal P, Dominoni DM, Gabriel KMA, Jechow A, Kupprat F, Lerner A, Marchant P, Riley W, et al. Light Pollution, Circadian Photoreception, and Melatonin in Vertebrates. Sustainability. 2019; 11(22):6400. https://doi.org/10.3390/su11226400
Chicago/Turabian StyleGrubisic, Maja, Abraham Haim, Pramod Bhusal, Davide M. Dominoni, Katharina M. A. Gabriel, Andreas Jechow, Franziska Kupprat, Amit Lerner, Paul Marchant, William Riley, and et al. 2019. "Light Pollution, Circadian Photoreception, and Melatonin in Vertebrates" Sustainability 11, no. 22: 6400. https://doi.org/10.3390/su11226400
APA StyleGrubisic, M., Haim, A., Bhusal, P., Dominoni, D. M., Gabriel, K. M. A., Jechow, A., Kupprat, F., Lerner, A., Marchant, P., Riley, W., Stebelova, K., van Grunsven, R. H. A., Zeman, M., Zubidat, A. E., & Hölker, F. (2019). Light Pollution, Circadian Photoreception, and Melatonin in Vertebrates. Sustainability, 11(22), 6400. https://doi.org/10.3390/su11226400