Beyond Assuming Co-Benefits in Nature-Based Solutions: A Human-Centered Approach to Optimize Social and Ecological Outcomes for Advancing Sustainable Urban Planning


Abstract
:1. Introduction
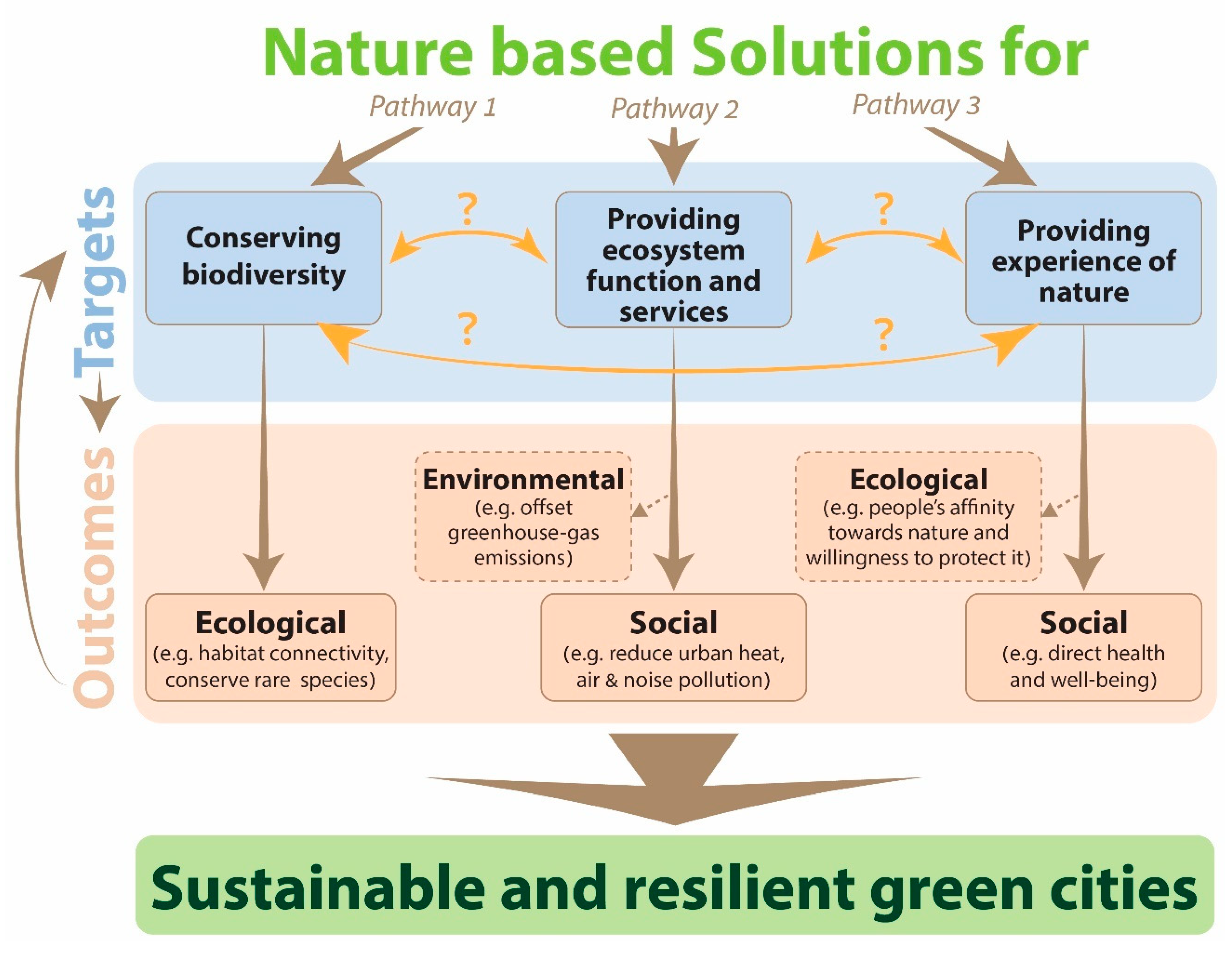
2. Synthesis of the Literature: Pathways, Synergies, and Trade-offs
2.1. Complexities within the Three Pathways for Greening Cities
2.2. Trade-Offs and Synergies among the Three Pathways
3. Towards a More Integrated Approach: A Novel Framework
3.1. Predicting Where Species Will Be
3.2. Predicting Where People Will Be Healthy, Happy, and Connected to Nature
3.3. Identifying Optimal NbS Implementation That Maximizes Co-Benefits
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Commission. Nature-Based Solutions. Available online: https://ec.europa.eu/research/environment/index.cfm?pg=nbs (accessed on 6 May 2019).
- Faivre, N.; Fritz, M.; Freitas, T.; de Boissezon, B.; Vandewoestijne, S. Nature-Based Solutions in the EU: Innovating with nature to address social, economic and environmental challenges. Environ. Res. 2017, 159, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Raymond, C.M.; Frantzeskaki, N.; Kabisch, N.; Berry, P.; Breil, M.; Nita, M.R.; Geneletti, D.; Calfapietra, C. A framework for assessing and implementing the co-benefits of nature-based solutions in urban areas. Environ. Sci. Policy 2017, 77, 15–24. [Google Scholar] [CrossRef]
- Aronson, M.F.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the city: Key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef]
- Shwartz, A.; Turbé, A.; Julliard, R.; Simon, L.; Prévot, A.-C. Outstanding challenges for urban conservation research and action. Glob. Environ. Chang. 2014, 28, 39–49. [Google Scholar] [CrossRef]
- Ossola, A.; Hahs, A.K.; Livesley, S.J. Habitat complexity influences fine scale hydrological processes and the incidence of stormwater runoff in managed urban ecosystems. J. Environ. Manag. 2015, 159, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Urban Nature Atlas. Available online: https://naturvation.eu/atlas (accessed on 17 July 2019).
- Yang, J.; McBride, J.; Zhou, J.; Sun, Z. The urban forest in Beijing and its role in air pollution reduction. Urban For. Urban Green. 2005, 3, 65–78. [Google Scholar] [CrossRef]
- Van den Bosch, M.; Sang, Å.O. Urban natural environments as nature-based solutions for improved public health—A systematic review of reviews. Environ. Res. 2017, 158, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Dearborn, D.C.; Kark, S. Motivations for Conserving Urban Biodiversity. Conserv. Biol. 2010, 24, 432–440. [Google Scholar] [CrossRef]
- Beatley, T. Biophilic Cities; Island Press: Washington, DC, USA, 2010. [Google Scholar]
- Matsuoka, R.H.; Kaplan, R. People needs in the urban landscape: Analysis of Landscape and Urban Planning contributions. Landsc. Urban Plan. 2008, 84, 7–19. [Google Scholar] [CrossRef]
- Maes, J.; Paracchini, M.L.; Zulian, G.; Dunbar, M.B.; Alkemade, R. Synergies and trade-offs between ecosystem service supply, biodiversity, and habitat conservation status in Europe. Biol. Conserv. 2012, 155, 1–12. [Google Scholar] [CrossRef]
- Howe, C.; Suich, H.; Vira, B.; Mace, G.M. Creating win-wins from trade-offs? Ecosystem services for human well-being: A meta-analysis of ecosystem service trade-offs and synergies in the real world. Glob. Environ. Chang. 2014, 28, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Knapp, S.; Haase, D.; Klotz, S.; Schwarz, N. Do Urban Biodiversity and Urban Ecosystem Services Go Hand in Hand, or Do We Just Hope It Is That Easy? In Urban Transformations; Future City, Springer: Cham, Germany, 2018; pp. 301–312. [Google Scholar]
- Jax, K.; Heink, U. Searching for the place of biodiversity in the ecosystem services discourse. Biol. Conserv. 2015, 191, 198–205. [Google Scholar] [CrossRef]
- Pett, T.J.; Shwartz, A.; Irvine, K.N.; Dallimer, M.; Davies, Z.G. Unpacking the People–Biodiversity Paradox: A Conceptual Framework. BioScience 2016, 66, 576–583. [Google Scholar] [CrossRef]
- Shanahan, D.F.; Lin, B.B.; Bush, R.; Gaston, K.J.; Dean, J.H.; Barber, E.; Fuller, R.A. Toward improved public health outcomes from urban nature. Am. J. Public Health 2015, 105, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Knapp, S.; Kühn, I.; Bakker, J.P.; Kleyer, M.; Klotz, S.; Ozinga, W.A.; Poschlod, P.; Thompson, K.; Thuiller, W.; Römermann, C. How species traits and affinity to urban land use control large-scale species frequency. Divers. Distrib. 2009, 15, 533–546. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: a meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Shwartz, A.; Muratet, A.; Simon, L.; Julliard, R. Local and management variables outweigh landscape effects in enhancing the diversity of different taxa in a big metropolis. Biol. Conserv. 2013, 157, 285–292. [Google Scholar] [CrossRef]
- Battin, J. When Good Animals Love Bad Habitats: Ecological Traps and the Conservation of Animal Populations. Conserv. Biol. 2004, 18, 1482–1491. [Google Scholar] [CrossRef]
- Mänd, R.; Tilgar, V.; Lõhmus, A.; Leivits, A. Providing nest boxes for hole-nesting birds—Does habitat matter? Biodivers. Conserv. 2005, 14, 1823–1840. [Google Scholar] [CrossRef]
- Lin, B.B.; Fuller, R.A. FORUM: Sharing or sparing? How should we grow the world’s cities? J. Appl. Ecol. 2013, 50, 1161–1168. [Google Scholar]
- Sushinsky, J.R.; Rhodes, J.R.; Possingham, H.P.; Gill, T.K.; Fuller, R.A. How should we grow cities to minimize their biodiversity impacts? Glob. Chang. Biol. 2013, 19, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Pataki, D.E.; Carreiro, M.M.; Cherrier, J.; Grulke, N.E.; Jennings, V.; Pincetl, S.; Pouyat, R.V.; Whitlow, T.H.; Zipperer, W.C. Coupling biogeochemical cycles in urban environments: ecosystem services, green solutions, and misconceptions. Front. Ecol. Environ. 2011, 9, 27–36. [Google Scholar] [CrossRef]
- Haase, D.; Larondelle, N.; Andersson, E.; Artmann, M.; Borgström, S.; Breuste, J.; Gomez-Baggethun, E.; Gren, Å.; Hamstead, Z.; Hansen, R.; et al. A Quantitative Review of Urban Ecosystem Service Assessments: Concepts, Models, and Implementation. AMBIO 2014, 43, 413–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabisch, N.; Frantzeskaki, N.; Pauleit, S.; Naumann, S.; Davis, M.; Artmann, M.; Haase, D.; Knapp, S.; Korn, H.; Stadler, J.; et al. Nature-based solutions to climate change mitigation and adaptation in urban areas: perspectives on indicators, knowledge gaps, barriers, and opportunities for action. Ecol. Soc. 2016, 21. [Google Scholar] [CrossRef] [Green Version]
- Grote, R.; Samson, R.; Alonso, R.; Amorim, J.H.; Cariñanos, P.; Churkina, G.; Fares, S.; Thiec, D.L.; Niinemets, Ü.; Mikkelsen, T.N.; et al. Functional traits of urban trees: Air pollution mitigation potential. Front. Ecol. Environ. 2016, 14, 543–550. [Google Scholar] [CrossRef]
- Soga, M.; Gaston, K.J. Extinction of experience: the loss of human–nature interactions. Front. Ecol. Environ. 2016, 14, 94–101. [Google Scholar] [CrossRef]
- Fischer, L.K.; Honold, J.; Botzat, A.; Brinkmeyer, D.; Cvejić, R.; Delshammar, T.; Elands, B.; Haase, D.; Kabisch, N.; Karle, S.J.; et al. Recreational ecosystem services in European cities: Sociocultural and geographical contexts matter for park use. Ecosyst. Serv. 2018, 31, 455–467. [Google Scholar] [CrossRef]
- Eizenberg, E.; Orenstein, D.E.; Zimroni, H. Back to the (Visualization) Laboratory, Back to the (Visualization) Laboratory: Using Focus Groups to Generate Qualitative and Quantitative Results, Using Focus Groups to Generate Qualitative and Quantitative Results. J. Plan. Educ. Res. 2018, 38, 345–358. [Google Scholar]
- Gaston, K.J.; Soga, M.; Duffy, J.P.; Garrett, J.K.; Gaston, S.; Cox, D.T.C. Personalised Ecology. Trends Ecol. Evol. 2018, 33, 916–925. [Google Scholar] [CrossRef] [Green Version]
- Wu, J. Urban sustainability: An inevitable goal of landscape research. Landsc. Ecol. 2010, 25, 1–4. [Google Scholar] [CrossRef]
- Soga, M.; Yamaura, Y.; Koike, S.; Gaston, K.J. Land sharing vs. land sparing: Does the compact city reconcile urban development and biodiversity conservation? J. Appl. Ecol. 2014, 51, 1378–1386. [Google Scholar] [CrossRef]
- Soga, M.; Yamaura, Y.; Aikoh, T.; Shoji, Y.; Kubo, T.; Gaston, K.J. Reducing the extinction of experience: Association between urban form and recreational use of public greenspace. Landsc. Urban Plan. 2015, 143, 69–75. [Google Scholar] [CrossRef]
- Shanahan, D.F.; Fuller, R.A.; Bush, R.; Lin, B.B.; Gaston, K.J. The Health Benefits of Urban Nature: How Much Do We Need? BioScience 2015, 65, 476–485. [Google Scholar] [CrossRef] [Green Version]
- Fischer, L.K.; Honold, J.; Cvejić, R.; Delshammar, T.; Hilbert, S.; Lafortezza, R.; Nastran, M.; Nielsen, A.B.; Pintar, M.; van der Jagt, A.P.N.; et al. Beyond green: Broad support for biodiversity in multicultural European cities. Glob. Environ. Chang. 2018, 49, 35–45. [Google Scholar] [CrossRef]
- Clayton, S.; Colléony, A.; Conversy, P.; Maclouf, E.; Martin, L.; Torres, A.-C.; Truong, M.-X.; Prévot, A.-C. Transformation of experience: Toward a new relationship with nature. Conserv. Lett. 2017, 10, 645–651. [Google Scholar] [CrossRef]
- Nassauer, J.I. Messy Ecosystems, Orderly Frames. Landsc. J. 1995, 14, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Lindemann-Matthies, P.; Matthies, D. The influence of plant species richness on stress recovery of humans. Web Ecol. 2018, 18, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Fuller, R.A.; Irvine, K.N.; Devine-Wright, P.; Warren, P.H.; Gaston, K.J. Psychological benefits of greenspace increase with biodiversity. Biol. Lett. 2007, 3, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Dallimer, M.; Irvine, K.N.; Skinner, A.M.J.; Davies, Z.G.; Rouquette, J.R.; Maltby, L.L.; Warren, P.H.; Armsworth, P.R.; Gaston, K.J. Biodiversity and the Feel-Good Factor: Understanding Associations between Self-Reported Human Well-being and Species Richness. BioScience 2012, 62, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Shwartz, A.; Turbé, A.; Simon, L.; Julliard, R. Enhancing urban biodiversity and its influence on city-dwellers: An experiment. Biol. Conserv. 2014, 171, 82–90. [Google Scholar] [CrossRef]
- Chawla, L. Children’s concern for the natural environment. Child. Environ. Q. 1988, 5, 13–20. [Google Scholar]
- Colléony, A.; White, R.; Shwartz, A. The influence of spending time outside on experience of nature and environmental attitudes. Landsc. Urban Plan. 2019, 187, 96–104. [Google Scholar] [CrossRef]
- Reynolds, S.J.; Galbraith, J.A.; Smith, J.A.; Jones, D.N. Garden Bird Feeding: Insights and Prospects from a North-South Comparison of This Global Urban Phenomenon. Front. Ecol. Evol. 2017, 5, 24. [Google Scholar] [CrossRef]
- Robb, G.N.; McDonald, R.A.; Chamberlain, D.E.; Bearhop, S. Food for thought: supplementary feeding as a driver of ecological change in avian populations. Front. Ecol. Environ. 2008, 6, 476–484. [Google Scholar] [CrossRef]
- McKinney, M.L. Urban futures. In Urban Ecology; Cambridge University Press: Cambridge, UK, 2010; pp. 287–308. [Google Scholar]
- Shwartz, A.; Cheval, H.; Simon, L.; Julliard, R. Virtual Garden Computer Program for use in Exploring the Elements of Biodiversity People Want in Cities. Conserv. Biol. 2013, 27, 876–886. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.U.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blouin, D.; Pellerin, S.; Poulin, M. Increase in non-native species richness leads to biotic homogenization in vacant lots of a highly urbanized landscape. Urban Ecosyst. 2019. [Google Scholar] [CrossRef]
- White, R.L.; Strubbe, D.; Dallimer, M.; Davies, Z.G.; Davis, A.J.S.; Edelaar, P.; Groombridge, J.; Jackson, H.A.; Menchetti, M.; Mori, E.; et al. Assessing the ecological and societal impacts of alien parrots in Europe using a transparent and inclusive evidence-mapping scheme. NeoBiota 2019, 48, 45–69. [Google Scholar] [CrossRef] [Green Version]
- Luna, A.; Edelaar, P.; Shwartz, A. Assessment of social perception of an invasive parakeet using a novel visual survey method. NeoBiota 2019, 46, 71–89. [Google Scholar] [CrossRef] [Green Version]
- Toger, M.; Benenson, I.; Wang, Y.; Malkinson, D.; Czamanski, D. Pigs in space: an agent-based model of the penetration of wild boars into cities. Available online: https://www.academia.edu/28814735/Pigs_in_space_an_agent-based_model_of_the_penetration_of_wild_boars_into_cities_Working_draft (accessed on 25 May 2019).
- Yirga, G.; Leirs, H.; Iongh, H.H.D.; Asmelash, T.; Gebrehiwot, K.; Deckers, J.; Bauer, H. Spotted hyena (Crocuta crocuta) concentrate around urban waste dumps across Tigray, northern Ethiopia. Wildl. Res. 2015, 42, 563–569. [Google Scholar] [CrossRef]
- Linklater, W.; Chapman, H.; Gregor, A.; Calder-Flynn, R.; Gouws, J.; Quigan, O.; Rustandi, A.; Brian-Molitaviti, J.; Ying, Y. Initiating a conflict with wildlife – the reintroduction and feeding of kākā, Wellington City, New Zealand. Pac. Conserv. Biol. 2018, 24, 360–370. [Google Scholar] [CrossRef]
- Naidoo, R.; Balmford, A.; Costanza, R.; Fisher, B.; Green, R.E.; Lehner, B.; Malcolm, T.R.; Ricketts, T.H. Global mapping of ecosystem services and conservation priorities. Proc. Natl. Acad. Sci. USA 2008, 105, 9495–9500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyytimäki, J.; Petersen, L.K.; Normander, B.; Bezák, P. Nature as a nuisance? Ecosystem services and disservices to urban lifestyle. Environ. Sci. 2008, 5, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Lovasi, G.S.; Quinn, J.W.; Neckerman, K.M.; Perzanowski, M.S.; Rundle, A. Children living in areas with more street trees have lower asthma prevalence. J. Epidemiol. Community Health 2008, 62, 647–649. [Google Scholar] [CrossRef] [PubMed]
- Lovasi, G.S.; O’Neil-Dunne, J.P.M.; Lu, J.W.T.; Sheehan, D.; Perzanowski, M.S.; Macfaden, S.W.; King, K.L.; Matte, T.; Miller, R.L.; Hoepner, L.A.; et al. Urban tree canopy and asthma, wheeze, rhinitis, and allergic sensitization to tree pollen in a New York City birth cohort. Environ. Health Perspect. 2013, 121, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Bodnaruk, E.W.; Kroll, C.N.; Yang, Y.; Hirabayashi, S.; Nowak, D.J.; Endreny, T.A. Where to plant urban trees? A spatially explicit methodology to explore ecosystem service tradeoffs. Landsc. Urban Plan. 2017, 157, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, N.; Moretti, M.; Bugalho, M.N.; Davies, Z.G.; Haase, D.; Hack, J.; Hof, A.; Melero, Y.; Pett, T.J.; Knapp, S. Understanding biodiversity-ecosystem service relationships in urban areas: A comprehensive literature review. Ecosyst. Serv. 2017, 27 Pt A, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Anguelovski, I.; Connolly, J.J.T.; Masip, L.; Pearsall, H. Assessing green gentrification in historically disenfranchised neighborhoods: a longitudinal and spatial analysis of Barcelona. Urban Geogr. 2017, 39, 458–491. [Google Scholar] [CrossRef]
- Wolch, J.R.; Byrne, J.; Newell, J.P. Urban green space, public health, and environmental justice: The challenge of making cities ‘just green enough. ’ Landsc. Urban Plan. 2014, 125, 234–244. [Google Scholar] [CrossRef]
- Dooling, S. Ecological Gentrification: A Research Agenda Exploring Justice in the City. Int. J. Urban Reg. Res. 2009, 33, 621–639. [Google Scholar] [CrossRef]
- Buckley, R.C.; Brough, P. Economic Value of Parks via Human Mental Health: An Analytical Framework. Front. Ecol. Evol. 2017. [Google Scholar] [CrossRef]
- Zhang, X.; Estoque, R.C.; Murayama, Y. An urban heat island study in Nanchang City, China based on land surface temperature and social-ecological variables. Sustain. Cities Soc. 2017, 32, 557–568. [Google Scholar] [CrossRef]
- Pan, H.; Zhang, L.; Cong, C.; Deal, B.; Wang, Y. A dynamic and spatially explicit modeling approach to identify the ecosystem service implications of complex urban systems interactions. Ecol. Indic. 2019, 102, 426–436. [Google Scholar] [CrossRef]
- Pan, H.; Page, J.; Zhang, L.; Chen, S.; Cong, C.; Destouni, G.; Kalantari, Z.; Deal, B. Using comparative socio-ecological modeling to support Climate Action Planning (CAP). J. Clean. Prod. 2019, 232, 30–42. [Google Scholar] [CrossRef]
- Ball, I.R.; Possingham, H.P.; Watts, M. Marxan and relatives: Software for spatial conservation prioritisation. In Spatial Conservation Prioritisation: Quantitative Methods and Computational Tools; Oxford University Press: Oxford, UK, 2009; pp. 185–195. [Google Scholar]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Araújo, M.B.; Peterson, A.T. Uses and misuses of bioclimatic envelope modeling. Ecology 2012, 93, 1527–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Kattwinkel, M.; Biedermann, R.; Kleyer, M. Temporary conservation for urban biodiversity. Biol. Conserv. 2011, 144, 2335–2343. [Google Scholar] [CrossRef]
- Kattwinkel, M.; Strauss, B.; Biedermann, R.; Kleyer, M. Modelling multi-species response to landscape dynamics: mosaic cycles support urban biodiversity. Landsc. Ecol. 2009, 24, 929–941. [Google Scholar] [CrossRef]
- Davis, A.Y.; Belaire, J.A.; Farfan, M.A.; Milz, D.; Sweeney, E.R.; Loss, S.R.; Minor, E.S. Green infrastructure and bird diversity across an urban socioeconomic gradient. Ecosphere 2012, 3, 105. [Google Scholar] [CrossRef]
- Louarn, M.L.; Clergeau, P.; Strubbe, D.; Deschamps-Cottin, M. Dynamic species distribution models reveal spatiotemporal habitat shifts in native range-expanding versus non-native invasive birds in an urban area. J. Avian Biol. 2018, 49, jav-01527. [Google Scholar] [CrossRef]
- Landau, K.I.; van Leeuwen, W.J.D. Fine scale spatial urban land cover factors associated with adult mosquito abundance and risk in Tucson, Arizona. J. Vector Ecol. J. Soc. Vector Ecol. 2012, 37, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Rödder, D.; Nekum, S.; Cord, A.F.; Engler, J.O. Coupling Satellite Data with Species Distribution and Connectivity Models as a Tool for Environmental Management and Planning in Matrix-Sensitive Species. Environ. Manag. 2016, 58, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Kabisch, N.; Korn, H.; Stadler, J.; Bonn, A. Nature-based Solutions to Climate Change Adaptation in Urban Areas —Linkages between Science, Policy and Practice. In Theory and Practice of Urban Sustainability Transitions; SpringerOpen: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Casalegno, S.; Anderson, K.; Cox, D.T.C.; Hancock, S.; Gaston, K.J. Ecological connectivity in the three-dimensional urban green volume using waveform airborne lidar. Sci. Rep. 2017, 7, 45571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayanu, Y.Z.; Conrad, C.; Nauss, T.; Wegmann, M.; Koellner, T. Quantifying and Mapping Ecosystem Services Supplies and Demands: A Review of Remote Sensing Applications. Environ. Sci. Technol. 2012, 46, 8529–8541. [Google Scholar] [CrossRef] [PubMed]
- Schröter, M.; Kraemer, R.; Mantel, M.; Kabisch, N.; Hecker, S.; Richter, A.; Neumeier, V.; Bonn, A. Citizen science for assessing ecosystem services: Status, challenges and opportunities. Ecosyst. Serv. 2017, 28, 80–94. [Google Scholar] [CrossRef]
- Feld, C.K.; Martins da Silva, P.; Sousa, J.P.; Bello, F.D.; Bugter, R.; Grandin, U.; Hering, D.; Lavorel, S.; Mountford, O.; Pardo, I.; et al. Indicators of biodiversity and ecosystem services: A synthesis across ecosystems and spatial scales. Oikos 2009, 118, 1862–1871. [Google Scholar] [CrossRef]
- Crawford, J.R.; Henry, J.D. The Positive and Negative Affect Schedule (PANAS): Construct validity, measurement properties and normative data in a large non-clinical sample. Br. J. Clin. Psychol. 2004, 43, 245–265. [Google Scholar] [CrossRef] [PubMed]
- Luck, G.W.; Davidson, P.; Boxall, D.; Smallbone, L. Relations between urban bird and plant communities and human well-being and connection to nature. Conserv. Biol. 2011, 25, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Tennant, R.; Hiller, L.; Fishwick, R.; Platt, S.; Joseph, S.; Weich, S.; Parkinson, J.; Secker, J.; Stewart-Brown, S. The Warwick-Edinburgh Mental Well-being Scale (WEMWBS): development and UK validation. Health Qual. Life Outcomes 2007, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Tam, K.-P. Concepts and measures related to connection to nature: Similarities and differences. J. Environ. Psychol. 2013, 34, 64–78. [Google Scholar] [CrossRef]
- Nisbet, E.K.; Zelenski, J.M.; Murphy, S.A. The Nature Relatedness Scale: Linking Individuals’ Connection With Nature to Environmental Concern and Behavior. Environ. Behav. 2009, 41, 715–740. [Google Scholar] [CrossRef]
- Clayton, S. Environmental Identity: A Conceptual and an Operational Definition. In Identity and the Natural Environment; MIT Press: Cambridge, MA, USA, 2003; pp. 45–65. [Google Scholar]
- Shanahan, D.F.; Bush, R.; Gaston, K.J.; Lin, B.B.; Dean, J.; Barber, E.; Fuller, R.A. Health Benefits from Nature Experiences Depend on Dose. Sci. Rep. 2016, 6, 28551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartig, T.; Evans, G.W.; Jamner, L.D.; Davis, D.S.; Gärling, T. Tracking restoration in natural and urban field settings. J. Environ. Psychol. 2003, 23, 109–123. [Google Scholar] [CrossRef]
- Keniger, L.E.; Gaston, K.J.; Irvine, K.N.; Fuller, R.A. What are the Benefits of Interacting with Nature? Int. J. Environ. Res. Public. Health 2013, 10, 913–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altshuler, B. Modeling of dose-response relationships. Environ. Health Perspect. 1981, 42, 23–27. [Google Scholar] [CrossRef]
- Barton, J.; Pretty, J. What is the Best Dose of Nature and Green Exercise for Improving Mental Health? A Multi-Study Analysis. Environ. Sci. Technol. 2010, 44, 3947–3955. [Google Scholar] [CrossRef]
- Cox, D.T.C.; Shanahan, D.F.; Hudson, H.; Fuller, R.; Anderson, K.; Hancock, S.; Gaston, K.; Cox, D.T.C.; Shanahan, D.F.; Hudson, H.L.; et al. Doses of Nearby Nature Simultaneously Associated with Multiple Health Benefits. Int. J. Environ. Res. Public. Health 2017, 14, 172. [Google Scholar] [CrossRef]
- Andersson, E.; McPhearson, T.; Kremer, P.; Gomez-Baggethun, E.; Haase, D.; Tuvendal, M.; Wurster, D. Scale and context dependence of ecosystem service providing units. Ecosyst. Serv. 2015, 12, 157–164. [Google Scholar] [CrossRef]
- Luck, G.W.; Daily, G.C.; Ehrlich, P.R. Population diversity and ecosystem services. Trends Ecol. Evol. 2003, 18, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Reid, C.E.; Clougherty, J.E.; Shmool, J.L.C.; Kubzansky, L.D. Is All Urban Green Space the Same? A Comparison of the Health Benefits of Trees and Grass in New York City. Int. J. Environ. Res. Public. Health 2017, 14, 1411. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, D.F.; Lin, B.B.; Gaston, K.J.; Bush, R.; Fuller, R.A. What is the role of trees and remnant vegetation in attracting people to urban parks? Landsc. Ecol. 2015, 30, 153–165. [Google Scholar] [CrossRef]
- Zardo, L.; Geneletti, D.; Pérez-Soba, M.; Van Eupen, M. Estimating the cooling capacity of green infrastructures to support urban planning. Ecosyst. Serv. 2017, 26, 225–235. [Google Scholar] [CrossRef]
- Sæbø, A.; Popek, R.; Nawrot, B.; Hanslin, H.M.; Gawronska, H.; Gawronski, S.W. Plant species differences in particulate matter accumulation on leaf surfaces. Sci. Total Environ. 2012, 427–428, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Shwartz, A.; Shirley, S.; Kark, S. How do habitat variability and management regime shape the spatial heterogeneity of birds within a large Mediterranean urban park? Landsc. Urban Plan. 2008, 84, 219–229. [Google Scholar] [CrossRef]
- Kearney, M.; Porter, W. Mechanistic niche modelling: combining physiological and spatial data to predict species’ ranges. Ecol. Lett. 2009, 12, 334–350. [Google Scholar] [CrossRef] [PubMed]
- Dormann, C.F.; Schymanski, S.J.; Cabral, J.; Chuine, I.; Graham, C.; Hartig, F.; Kearney, M.; Morin, X.; Römermann, C.; Schröder, B.; et al. Correlation and process in species distribution models: bridging a dichotomy. J. Biogeogr. 2012, 39, 2119–2131. [Google Scholar] [CrossRef]
- Gordon, A.; Simondson, D.; White, M.; Moilanen, A.; Bekessy, S.A. Integrating conservation planning and landuse planning in urban landscapes. Landsc. Urban Plan. 2009, 91, 183–194. [Google Scholar] [CrossRef]
- Lindemann-Matthies, P.; Keller, D.; Li, X.; Schmid, B. Attitudes toward forest diversity and forest ecosystem services—A cross-cultural comparison between China and Switzerland. J. Plant Ecol. 2014, 7, 1–9. [Google Scholar] [CrossRef]
- Espinosa, S.; Jacobson, S.K. Human-Wildlife Conflict and Environmental Education: Evaluating a Community Program to Protect the Andean Bear in Ecuador. J. Environ. Educ. 2012, 43, 55–65. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Cadenasso, M.L.; Childers, D.L.; Mcdonnell, M.J.; Zhou, W. Evolution and future of urban ecological science: ecology in, of, and for the city. Ecosyst. Health Sustain. 2016, 2, e01229. [Google Scholar] [CrossRef]
- Grimm, N.B.; Grove, J.G.; Pickett, S.T.A.; Redman, C.L. Integrated Approaches to Long-Term Studies of Urban Ecological SystemsUrban ecological systems present multiple challenges to ecologists—pervasive human impact and extreme heterogeneity of cities, and the need to integrate social and ecological approaches, concepts, and theory. BioScience 2000, 50, 571–584. [Google Scholar]
- Childers, D.L.; Cadenasso, M.L.; Grove, J.M.; Marshall, V.; McGrath, B.; Pickett, S.T.A. An Ecology for Cities: A Transformational Nexus of Design and Ecology to Advance Climate Change Resilience and Urban Sustainability. Sustainability 2015, 7, 3774–3791. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Components of Nature | Ecological Indicators | Suggested Method |
|---|---|---|
| Extent of lawn | The extent of lawn per unit area or grid square | The cover of visible lawn can be measured in a combined field and Geographic Information System (GIS) survey using transects or grid cells, depending on the scale and extent of lawn cover [101]. |
| Trees | Density of trees; tree cover and richness; tree cooling effect; tree uptake of air pollutants | Tree cover/density/richness can be estimated using GIS and using the tree layer of the considered area [102]; the tree cooling effect can be estimated using GIS survey using grid cells, depending on the scale and number of trees [103]; the tree uptake of air pollutants can be measured through chemical analyses [104]. |
| Plants and specific flowers | Plant or flower richness, abundance, and evenness; number of rainbow colors; vegetative heterogeneity | Field survey using (1) transects to sample shrubs and visible morphs of flowers; (2) quadrats to sample small herbaceous plants. The standard deviation of Normalized Difference Vegetation Index (NDVI) can be used to estimate the vegetative heterogeneity [93]. |
| Charismatic and colorful animals | Abundance of charismatic and colorful species | Field survey using transects or point counts [105]. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colléony, A.; Shwartz, A. Beyond Assuming Co-Benefits in Nature-Based Solutions: A Human-Centered Approach to Optimize Social and Ecological Outcomes for Advancing Sustainable Urban Planning. Sustainability 2019, 11, 4924. https://doi.org/10.3390/su11184924
Colléony A, Shwartz A. Beyond Assuming Co-Benefits in Nature-Based Solutions: A Human-Centered Approach to Optimize Social and Ecological Outcomes for Advancing Sustainable Urban Planning. Sustainability. 2019; 11(18):4924. https://doi.org/10.3390/su11184924
Chicago/Turabian StyleColléony, Agathe, and Assaf Shwartz. 2019. "Beyond Assuming Co-Benefits in Nature-Based Solutions: A Human-Centered Approach to Optimize Social and Ecological Outcomes for Advancing Sustainable Urban Planning" Sustainability 11, no. 18: 4924. https://doi.org/10.3390/su11184924
APA StyleColléony, A., & Shwartz, A. (2019). Beyond Assuming Co-Benefits in Nature-Based Solutions: A Human-Centered Approach to Optimize Social and Ecological Outcomes for Advancing Sustainable Urban Planning. Sustainability, 11(18), 4924. https://doi.org/10.3390/su11184924