Agroforestry and Biodiversity



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
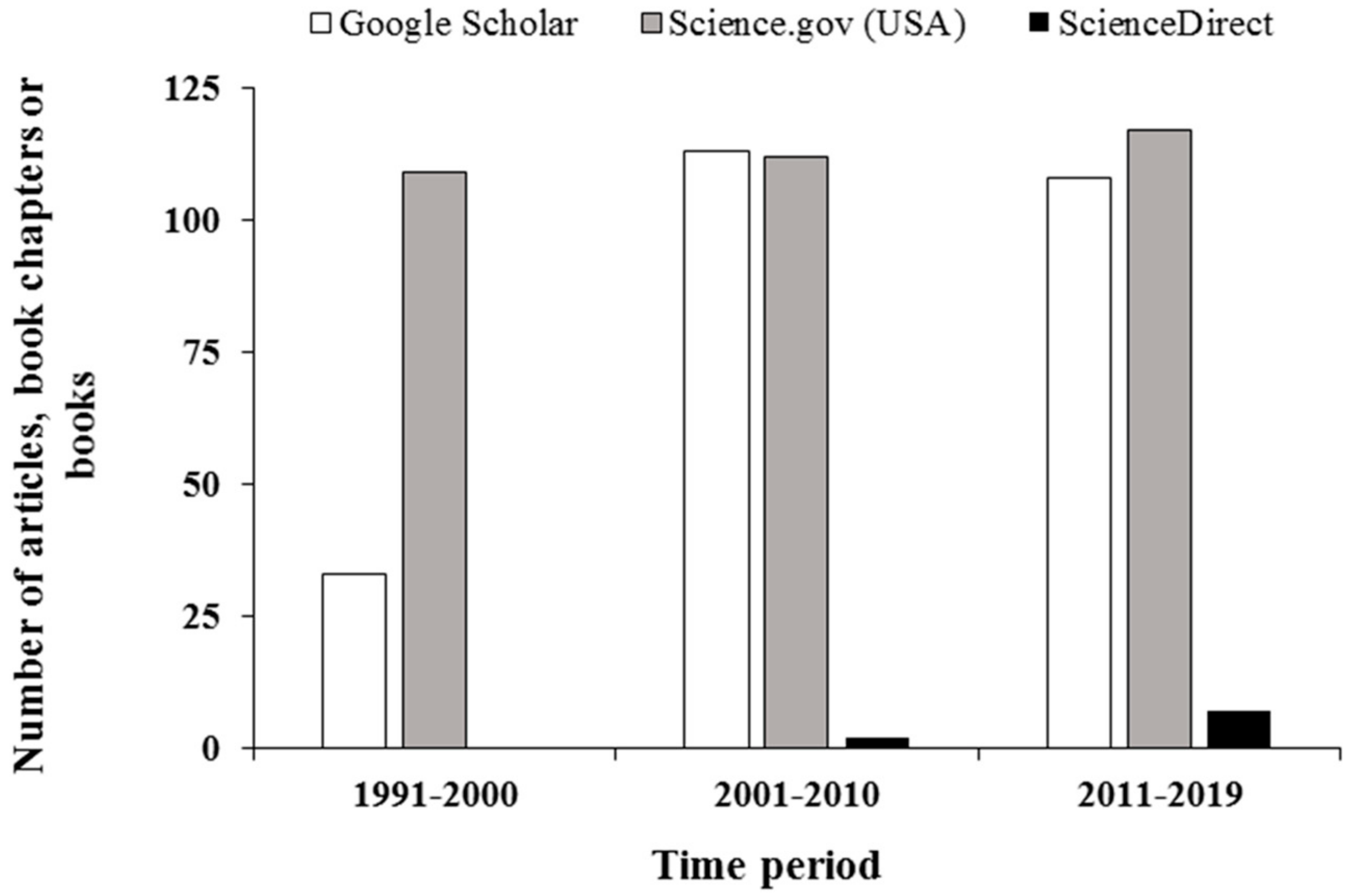
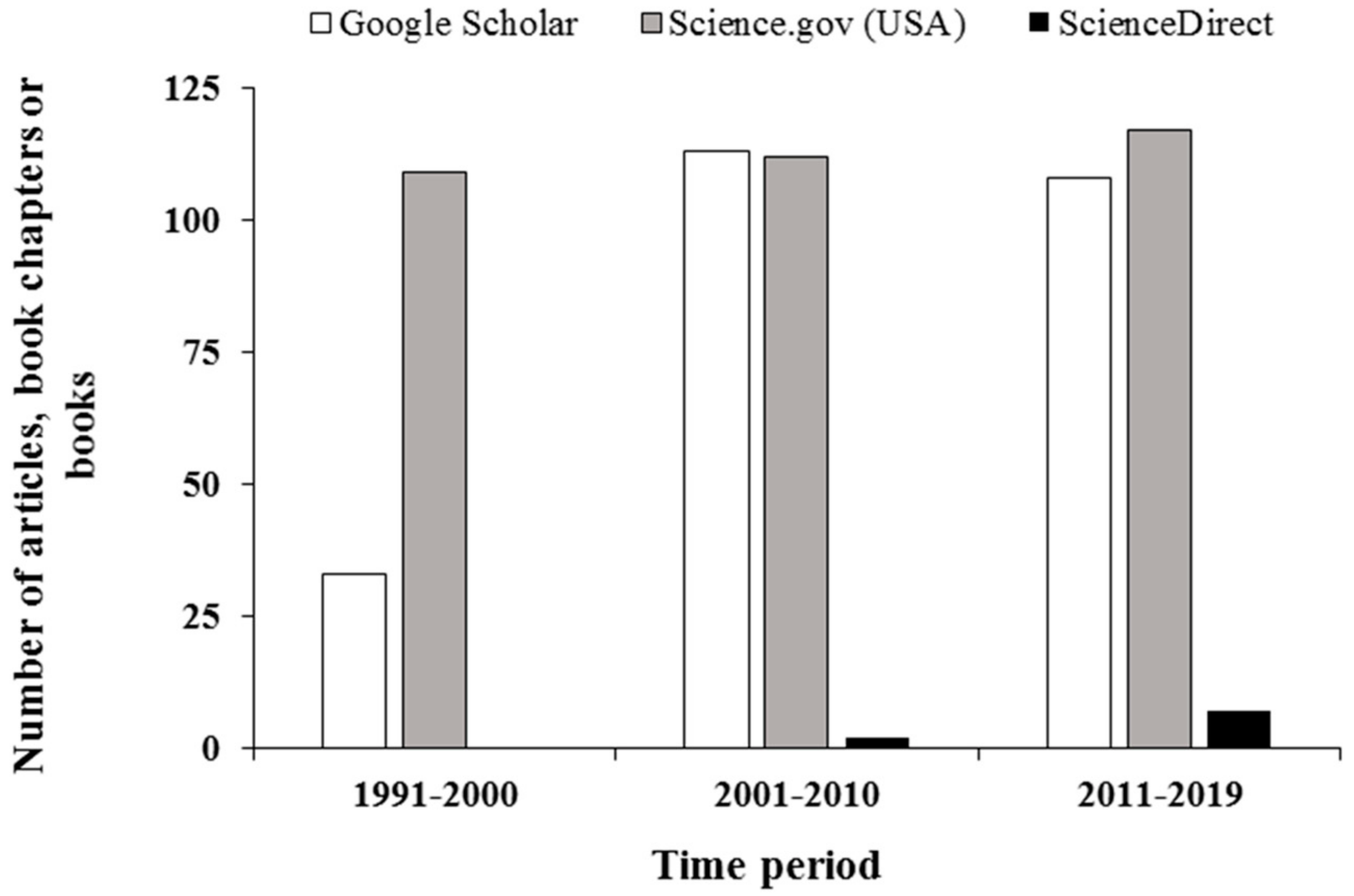
:1. Introduction
2. Materials and Methods
3. Agroforestry and Diversity
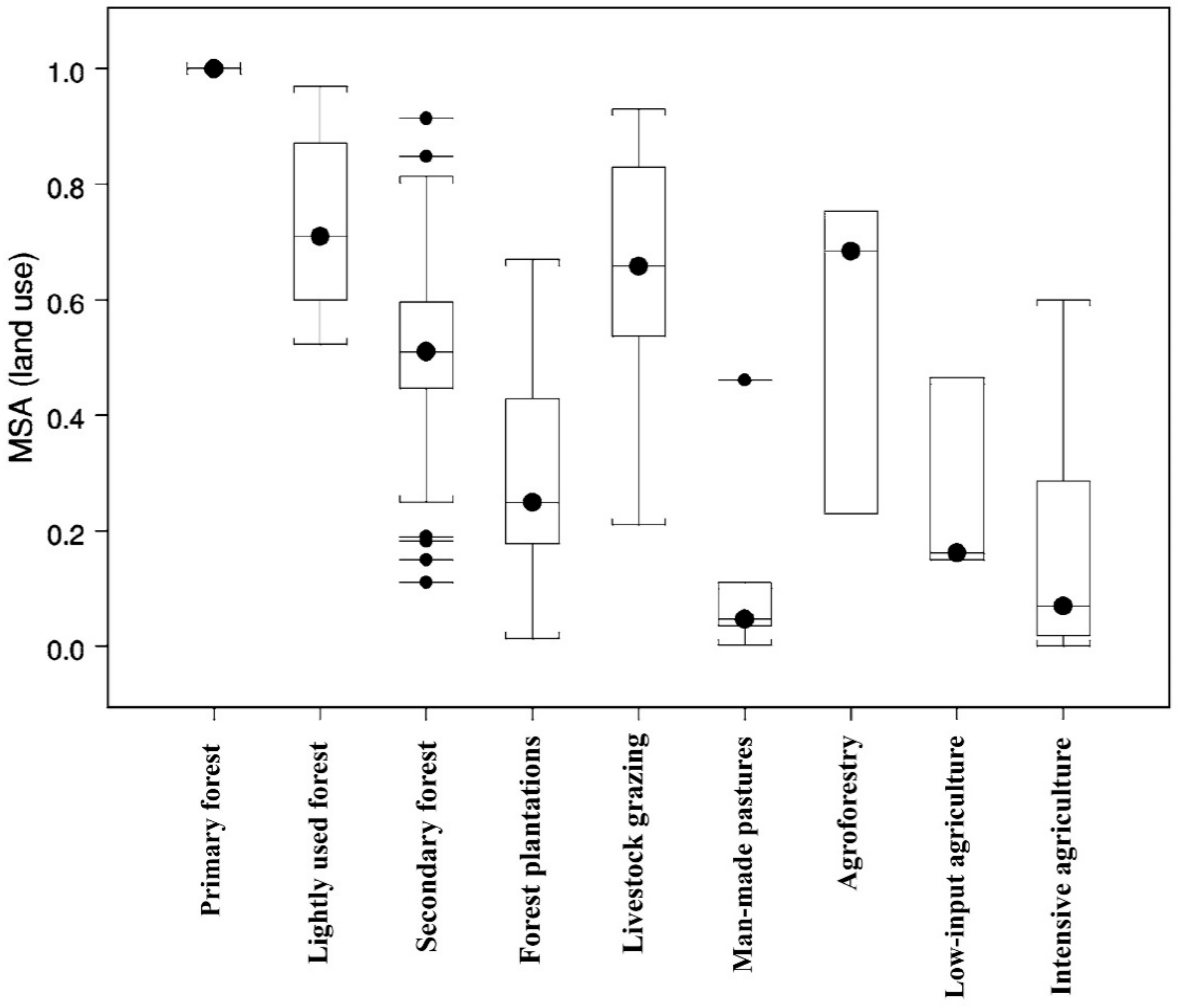
3.1. Agroforestry and Floral Diversity
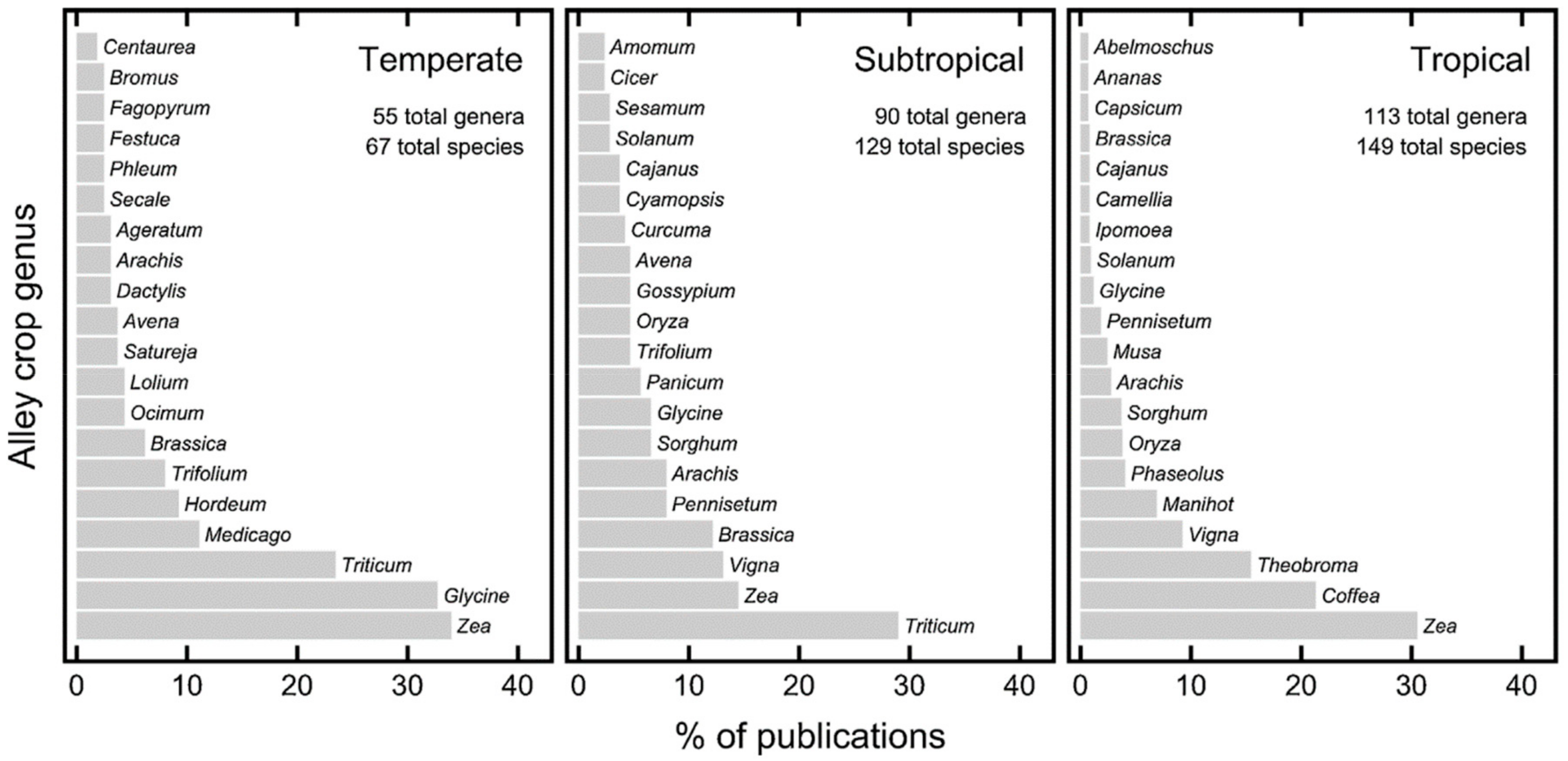
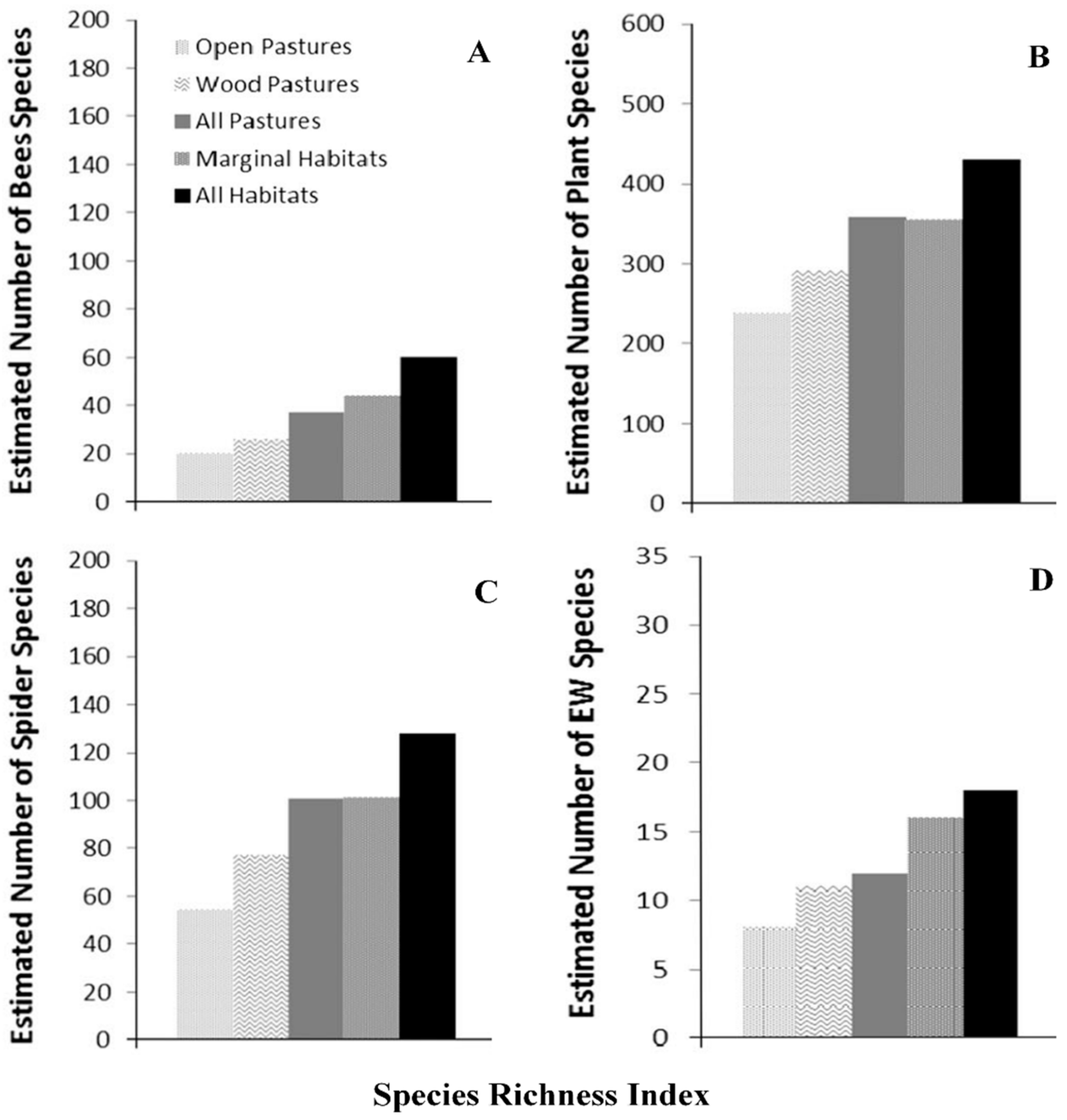
3.2. Agroforestry and Faunal Diversity
3.3. Agroforestry and Soil Microbial Diversity
4. Practical Implication of Agroforestry on Biodiversity
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AF | agroforestry |
| AMF | arbuscular mycorrhizae fungi |
| BD | biodiversity |
| SOC | soil organic carbon |
References
- Pimm, S.L.; Russell, G.J.; Gittleman, J.L.; Brooks, T.M. The future of biodiversity. Science 1995, 269, 269–350. [Google Scholar] [CrossRef]
- Jose, S. Agroforestry for conserving and enhancing biodiversity. Agrofor. Syst. 2012, 85, 1–8. [Google Scholar] [CrossRef]
- Pimm, S.L. Biodiversity: Climate change or habitat loss-which will kill more species? Curr. Biol. 2008, 18, R117–R119. [Google Scholar] [CrossRef]
- Rands, M.R.; Adams, W.M.; Bennun, L.; Butchart, S.H.; Clements, A.; Coomes, D.; Entwistle, A.; Hodge, I.; Kapos, V.; Scharlemann, J.P.; et al. Biodiversity conservation: Challenges beyond 2010. Science 2010, 329, 1298–1303. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Kessler, M.; Barkmann, J.; Bos, M.M.; Buchori, D.; Erasmi, S.; Faust, H.; Gerold, G.; Glenk, K.; Gradstein, S.R.; et al. Tradeoffs between income, biodiversity, and ecosystem functioning during tropical rainforest conversion and agroforestry intensification. Proc. Natl. Acad. Sci. USA 2007, 104, 4973–4978. [Google Scholar] [CrossRef]
- Culman, S.W.; Young-Mathews, A.; Hollander, A.D.; Ferris, H.; Sanchez-Moreno, S.; O’Green, A.T.; Jackson, L.E. Biodiversity is associated with indicators of soil ecosystem functions over a landscape gradient of agricultural intensification. Landsc. Ecol. 2010, 25, 1333–1348. [Google Scholar] [CrossRef]
- Vigouroux, Y.; Barnaud, A.; Scarcelli, N.; Thuillet, A.C. Biodiversity, evolution and adaptation of cultivated crops. C. R. Biol. 2011, 334, 450–457. [Google Scholar] [CrossRef]
- Thrupp, L.A. Linking agricultural biodiversity and food security: The valuable role of agrobiodiversity for sustainable agriculture. Int. Aff. 2000, 76, 283–297. [Google Scholar] [CrossRef]
- Sistla, S.A.; Roddy, A.B.; Williams, N.E.; Kramer, D.B.; Stevens, K.; Allison, S.D. Agroforestry practices promote biodiversity and natural resource diversity in Atlantic Nicaragua. PLoS ONE 2016, 11, e0162529. [Google Scholar] [CrossRef]
- Kleijn, D.; Sutherland, W.J. How effective are European agri-environment schemes in conserving and promoting biodiversity? J. Appl. Ecol. 2003, 40, 947–969. [Google Scholar] [CrossRef]
- Opermmann, R.; Beaufoy, G.; Jones, G. High Nature Value Farming in Europe: 35 European Countries—Experiences and Perspectives; Verlag Regionanlkultur: Ubstadt-Weiher, Germany, 2012. [Google Scholar]
- Kristensen, P. EEA Core Set of Indicators; Revised Version April 2003; European Environment Agency: Copenhagen, Denmark, 2003. [Google Scholar]
- McNeely, J.A.; Schroth, G. Agroforestry and biodiversity conservation—Traditional practices, present dynamics, and lessons for the future. Biodivers. Conserv. 2006, 15, 549–554. [Google Scholar] [CrossRef]
- Sanchez, P.A. Science in agroforestry. Agrofor. Syst. 1995, 30, 5–55. [Google Scholar] [CrossRef]
- Dobson, A.P.; Bradshaw, A.D.; Baker, A.J.M. Hopes for the future: Restoration ecology and conservation biology. Science 1997, 277, 515–522. [Google Scholar] [CrossRef]
- Huang, W.; Kanninen, M.; Xu, Q.; Huang, B. Agroforestry in China: Present state and future potential. Ambio 1997, 26, 393–397. [Google Scholar]
- Huang, W. Productive coexistence and gain in agroforestry systems. Acta For. Fenn. 1998, 260, 1–72. [Google Scholar] [CrossRef]
- Leakey, R.R.B. Agroforestry for biodiversity in farming systems. In Biodiversity in Agroecosystems; Collins, W.W., Qualset, C.O., Eds.; CRC Press: New York, NY, USA, 1999; pp. 127–145. [Google Scholar]
- Boffa, J.M. Agroforestry Parklands in Sub-Saharan Africa. In FAO Conservation Guide 34; Food and Agriculture Organization of the United Nations: Rome, Italy, 1999; Available online: http://www.fao.org/DOCREP/005/X3940E/X3940E00.htm (accessed on 7 May 2019).
- Huang, W.; Luukkanen, O.; Johanson, S.; Kaarakka, V.; Räisänen, S.; Vihemäki, H. Agroforestry for biodiversity conservation of nature reserves: Functional group identification and analysis. Agrofor. Syst. 2002, 55, 65–72. [Google Scholar] [CrossRef]
- Buck, L.E.; Gavin, T.A.; Lee, D.R.; Uphoff, N.T.; Behr, D.C.; Drinkwater, L.E.; Hively, W.D.; Werner, F.R. Ecoagriculture: A Review and Assessment of its Scientific Foundations; Cornell University: Ithaca, NY, USA, 2004. [Google Scholar]
- Méndez, V.E.; Lok, R.; Somarriba, E. Interdisciplinary analysis of home gardens in Nicaragua: Micro- zonation, plant use and socioeconomic importance. Agrofor. Syst. 2001, 51, 85–96. [Google Scholar] [CrossRef]
- Hemp, A. The banana forests of Kilimanjaro: Biodiversity and conservation of the Chagga home gardens. In Forest Diversity and Management. Topics in Biodiversity and Conservation; Hawksworth, D.L., Bull, A.T., Eds.; Springer: Amsterdam, The Netherlands, 2006; pp. 133–155. [Google Scholar]
- Borkhataria, R.R.; Collazo, J.A.; Groom, M.J. Species abundance and potential biological control services in shade vs. sun coffee in Puerto Rico. Agric. Ecosyst. Environ. 2012, 151, 1–5. [Google Scholar] [CrossRef]
- Noble, I.R.; Dirzo, R. Forests as human-dominated ecosystems. Science 1997, 277, 522–525. [Google Scholar] [CrossRef]
- Thevathasan, N.V.; Gordon, A.M. Ecology of tree intercropping system in the North temperate region: Experience from southern Ontario, Canada. Agrofor. Syst. 2004, 61, 257–268. [Google Scholar]
- Dollinger, J.; Jose, S. Agroforestry for soil health. Agrofor. Syst. 2018, 92, 213–219. [Google Scholar] [CrossRef]
- Bhagwat, S.A.; Willis, K.J.; Briks, H.J.B.; Whittaker, R.J. Agroforestry: A refuge for tropical biodiversity? Trends Ecol. Evol. 2008, 23, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Torralba, M.; Fagerholm, N.; Burgess, P.J.; Moreno, G.; Plieninger, T. Do European agroforestry systems enhance biodiversity and ecosystem services? A meta-analysis. Agric. Ecosyst. Environ. 2016, 230, 150–161. [Google Scholar] [CrossRef]
- Perfecto, I.; Rice, R.A.; Greenberg, R.; Van der Voort, M.E. Shade coffee: A disappearing refuge for biodiversity. Bioscience 1996, 46, 598–608. [Google Scholar] [CrossRef]
- Lawton, J.H.; Bignell, D.E.; Bolton, B.; Bloemers, G.F.; Eggleton, P.; Hammond, P.M.; Hodda, M.; Holt, R.D.; Larsen, T.B.; Mawdsley, N.A.; et al. Biodiversity inventories, indicator taxa and effects of habitat modification in tropical forest. Nature 1998, 391, 72–76. [Google Scholar] [CrossRef]
- Schroth, G.; da Fonseca, G.A.B.; Harvey, C.A. Agroforestry and Biodiversity Conservation in Tropical Landscapes; Island Press: Washington, DC, USA, 2004; p. 523. [Google Scholar]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
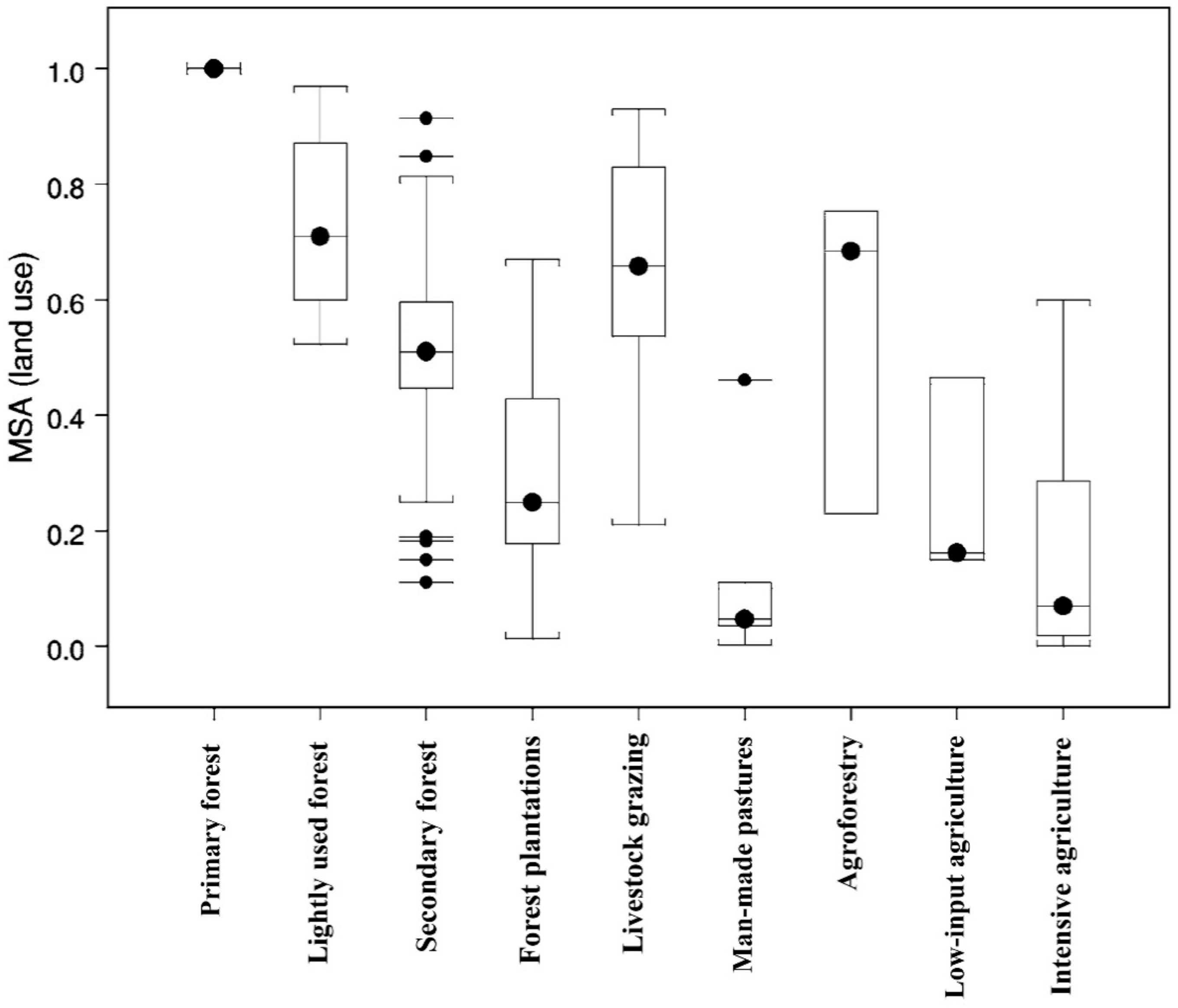
- Alkemade, R.; Van Oorschot, M.; Miles, L.; Nellemann, C.; Bakkenes, M.; Brink, B. GLOBIO3: A framework to investigate options for reducing global terrestrial biodiversity loss. Ecosystems 2009, 12, 374–390. [Google Scholar] [CrossRef]
- Barton, J.; Pretty, J. What is the best dose of nature and green exercise for improving mental health? A multi-study analysis. Environ. Sci. Technol. 2010, 44, 947–3955. [Google Scholar] [CrossRef]
- Leakey, R.; Tchoundjeu, Z.; Schreckenberg, K.; Simons, T.; Shackleton, S.; Mander, M.; Wynberg, R.; Shackleton, C.; Sullivan, C. Trees and markets for agroforestry tree products: Targeting poverty reduction and enhanced livelihoods. In World Agroforestry into the Future; Garrity, D., Okono, A., Grayson, M., Parrott, S., Eds.; World Agroforestry Centre: Nairobi, Kenya, 2006; pp. 11–22. [Google Scholar]
- Jose, S.; Gillespie, A.R.; Pallardy, S.G. Interspecific interactions in temperate agroforestry. Agrofor. Syst. 2004, 61, 237–255. [Google Scholar]
- Schulze, C.H.; Waltert, M.; Kessler, P.J.; Pitopang, R.; Veddeler, D.; Mühlenberg, M.; Gradstein, S.R.; Leuschner, C.; Steffan-Dewenter, I.; Tscharntke, T. Biodiversity indicator groups of tropical land-use systems: Comparing plants, birds, and insects. Ecol. Appl. 2004, 14, 1321–1333. [Google Scholar] [CrossRef]
- Udawatta, R.P.; Garrett, H.E.; Kallenbach, R.L. Agroforestry buffers for nonpoint source pollution reductions from agricultural watersheds. J. Environ. Qual. 2011, 40, 800–806. [Google Scholar] [CrossRef]
- Benayas, J.M.R.; Bullock, J.M. Restoration of biodiversity and ecosystem services on agricultural land. Ecosystems 2012, 15, 883–889. [Google Scholar] [CrossRef]
- Swallow, B.; Boffa, J. The potential for agroforestry to contribute to the conservation and enhancement of landscape biodiversity. In World Agroforestry into the Future; Garrity, D., Okono, A., Grayson, M., Parrott, S., Eds.; World Agroforestry Centre: Nairobi, Kenya, 2006; pp. 95–101. [Google Scholar]
- Varah, A.; Jones, H.; Smith, J.; Potts, S.G. Enhanced biodiversity and pollination in UK agroforestry systems. J. Sci. Food Agric. 2013, 93, 2073–2075. [Google Scholar] [CrossRef]
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multilayered relationship. Trends. Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef]
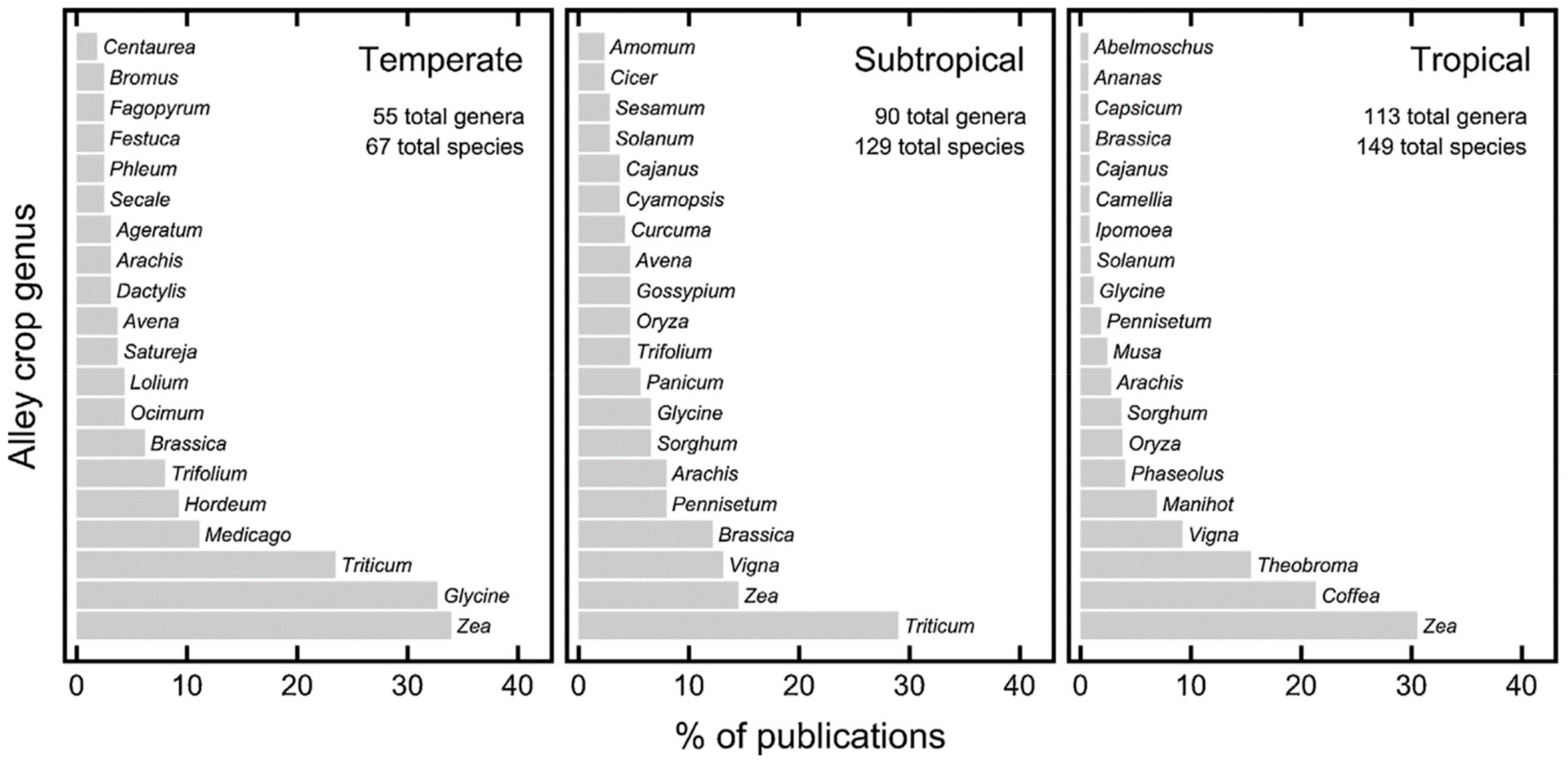
- Wolz, K.J.; DeLucia, E.H. Alley cropping: Global patterns of species composition and functions. Agric. Ecosyst. Environ. 2018, 252, 61–68. [Google Scholar] [CrossRef]
- Garrett, H.E.; McGraw, R.L.; Walter, W.D. Alley Cropping Practices. In North American Agroforestry: An Integrated Science and Practice, 2nd ed.; Garrett, H.E., Ed.; American Society of Agronomy: Madison, WI, USA, 2009; pp. 133–162. [Google Scholar]
- Chamberlain, J.L.; Mitchell, D.; Bringham, T.; Hobby, T.; Zabek, L.; Davis, J. Forest framing practices. In North American Agroforestry: An Integrated Science and Practice, 2nd ed.; Garrett, H.E., Ed.; American Society of Agronomy: Madison, WI, USA, 2009; pp. 219–256. [Google Scholar]
- Elliot, K.; Vose, J.M. Riparian area harvesting impacts on vegetation composition and diversity: Headwaters to Estuaries. In Advances in Watershed Science and Management; Younos, T., Parece, T.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; p. 104. [Google Scholar]
- Naiman, R.J.; Décamps, H.; McClain, M.E. Structutal pattern. In Riparia; Naiman, R.J., Décamps, H., McClain, M.E., Eds.; Elsevier Academic Press: New York, NY, USA, 2005; pp. 79–123. [Google Scholar]
- Sharrow, S.H.; Brauer, D.; Clason, T.R. Silvopastu practices. In North American Agroforestry: An Integrated Science and Practice, 2nd ed.; Garrett, H.E., Ed.; American Society of Agronomy: Madison, WI, USA, 2009; pp. 105–131. [Google Scholar]
- Orefice, J.; Carroll, J.; Conroy, D.; Ketner, L. Silvopasture practices and perspectives in the Northeasten United States. Agrofor. Syst. 2017, 91, 149–160. [Google Scholar] [CrossRef]
- Kort, J.; Turnock, R. Carbon Reservoir and Biomass in Canadian Prairie Shelterbelts. Agrofor. Syst. 1999, 44, 175–186. [Google Scholar] [CrossRef]
- Schultz, R.C.; Isenhart, T.M.; Colletti, J.P.; Simpkins, W.W.; Udawatta, R.P.; Schultz, P.L. Riparian and upland buffer practices. In North American Agroforestry: An Integrated Science and Practice, 2nd ed.; Garrett, H.E., Ed.; American Society of Agronomy Inc.: Madison, WI, USA, 2009; pp. 163–218. [Google Scholar]
- Afta.org. Available online: https://www.aftaweb.org/about/what-is-agroforestry/silvopasture.html (accessed on 7 May 2019).
- Bucheli, V.J.P.; Bokelmann, W. Agroforestry systems for biodiversity and ecosystem services: The case of the Sibundoy Valley in the Colombian province of Putumayo. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2017, 13, 380–397. [Google Scholar]
- Kumar, B.M.; Nair, P.K.R. Introduction, Tropical Home Gardens: A Time-Tested Example of Sustainable Agroforestry; Springer: Berlin/Heidelberg, Germany, 2006; pp. 1–12. [Google Scholar]
- Kabir, M.E.; Webb, E.L. Can home gardens conserve biodiversity in Bangladesh? Biotropica 2008, 40, 95–103. [Google Scholar]
- Kumari, M.A.S.; Kansuntisukmongkol, K.; Brockelman, W.Y. Plant diversity in home gardens and its contribution to household economy in suburban areas in Sri Lanka. Environ. Nat. Resour. J. 2009, 7, 12–30. [Google Scholar]
- Bardhan, S.; Jose, S.; Biswas, S.; Kabir, K.; Rogers, W. Home garden agroforestry systems: An intermediary for biodiversity conservation in Bangladesh. Agrofor. Syst. 2012, 85, 29–34. [Google Scholar] [CrossRef]
- Murdiyarso, D.; Van Noordwijk, M.; Wasrin, U.M.; Tomich, T.P.; Gillison, A.N. Environmental benefits and sustainable land-use options in the Jambi transect, Sumatra, Indonesia. J. Veg. Sci. 2002, 13, 429–438. [Google Scholar] [CrossRef]
- Jose, S. Agroforestry for ecosystem services and environmental benefits: An overview. Agrofor. Syst. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Ewel, J.J. Natural systems as models for the design of sustainable systems of land use. Agrofor. Syst. 1999, 45, 1–21. [Google Scholar] [CrossRef]
- Kindt, R. Methodology for Tree Species Diversification Planning for African Agroecosystems. Ph.D. Thesis, University of Ghent, Ghent, Belgium, 2002. [Google Scholar]
- Donald, P.F. Biodiversity impacts of some agricultural commodity production systems. Conserv. Biol. 2004, 18, 17–37. [Google Scholar] [CrossRef]
- Gillison, A.N.; Liswanti, N.; Budidarsono, S.; Noordwijk, M.V.; Tomich, T.P. Impact of cropping methods on biodiversity in coffee agroecosystems in Sumatra, Indonesia. Ecol. Soc. 2004, 9, 7. [Google Scholar] [CrossRef]
- Khan, M.L.; Arunachalam, A. Traditional agroforestry as a viable choice to conserve agro-biodiversity in the northeast India. In Agroforestry: Potentials and Opportunities; Pathak, P.S., Newaj, R., Eds.; Agrobios (India) and Indian Society of Agroforestry: Jodhpur, India, 2003; pp. 95–105. [Google Scholar]
- Michon, G.; De Foresta, H. The Indonesian agro-forest model. Forest resource management and biodiversity conservation. In Conserving Biodiversity Outside Protected Areas. The Role of Traditional Agro-Ecosystems; Halladay, P., Gilmour, D.A., Eds.; World Conservation Union, Forest Conservation Programme: Andalucía, Spain, 1995; pp. 90–106. [Google Scholar]
- Fanaye, A.O.; Adeyemi, E.A.; Olaiya, A.O. Spacing experiments in cocoa/kola/citrus intercrop. In Proceedings of the 14th International Cocoa Research Conference, Accra, Ghana, 13–18 October 2003. [Google Scholar]
- Asare, R. Cocoa Agroforests in West Africa: A Look at Activities on Preferred Trees in the Farming Systems; Forest & Landscape Working Papers No. 6-2005; Forest & Landscape Denmark (FLD): Hørsholm, Denmark, 2005. [Google Scholar]
- Osei-Bonsu, K.; Ameyaw-Oduro, C.; Tetteh, J.P. Traditional cocoa agroforestry: 1. Species encountered in the cocoa ecosystem of a typical cocoa-growing district in Ghana. In Proceedings of the 14th International Cocoa Research Conference, Accra, Ghana, 13–18 October 2003; pp. 13–18. [Google Scholar]
- Tomich, T.P.; Van Noordwijk, M.; Vosti, S.A.; Witcover, J. Agricultural development with rainforest conservation: Methods for seeking best bet alternatives to slash-and-burn, with applications to Brazil and Indonesia. Agric. Econ. 1998, 19, 159–174. [Google Scholar] [CrossRef]
- Tomich, T.P.; Van Noordwijk, M.; Budidarsono, S.; Gillison, A.; Kusumanto, T.; Murdiyarso, M.; Stolle, F.; Fagi, A.M. Agricultural intensification, deforestation, and the environment: Assessing tradeoffs in Sumatra, Indonesia. In Tradeoffs or Synergies? Agricultural Intensification, Economic Development and the Environment; Lee, D.R., Barrett, C.B., Eds.; CAB International: Wallingford, UK, 2001. [Google Scholar]
- Kearns, C.A.; Inouye, D.W. Pollinators, flowering plants, and conservation biology. Bioscience 1997, 47, 297–307. [Google Scholar] [CrossRef]
- Klein, A.M.; Steffan–Dewenter, I.; Tscharntke, T. Fruit set of highland coffee increases with the diversity of pollinating bees. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 955–961. [Google Scholar] [CrossRef]
- Reitsma, R.; Parrish, J.D.; McLarney, W. The role of cacao plantations in maintaining forest avian diversity in southeastern Costa Rica. Agrofor. Syst. 2001, 53, 185–193. [Google Scholar] [CrossRef]
- Gibbs, S.; Koblents, H.; Coleman, B.; Gordon, A.; Thevathasan, N.; Williams, P. Avian diversity in a temperate tree-based intercropping system from inception to now. Agrofor. Syst. 2016, 90, 905–916. [Google Scholar] [CrossRef]
- Williams, P.; Koblents, H.; Gordon, A. Bird use of an intercropped corn field and old field in southern Ontario, Canada. In Proceedings of the 4th North American Agroforestry Conference, Boise, ID, USA, 23–28 July 1995; Ehrenreich, J.H., Ehrenreich, D.L., Lee, H.W., Eds.; University of Idaho: Moscow, ID, USA, 1995; pp. 158–162. [Google Scholar]
- Harvey, C.A.; Gonzalez-Villalobos, J.A. Agroforestry systems conserve species-rich but modified assemblages of tropical birds and bats. Biodivers. Conserv. 2007, 16, 2257–2292. [Google Scholar] [CrossRef]
- Naidoo, R. Species richness and community composition of songbirds in a tropical forest-agriculture landscape. Anim. Conserv. 2004, 7, 93–105. [Google Scholar] [CrossRef]
- Soini, E. Dynamics of livelihood strategies in the changing landscapes of the southern slopes of Mt. Kilimanjaro, Tanzania. Agric. Syst. 2005, 85, 306–323. [Google Scholar] [CrossRef]
- Griffith, D.M. Agroforestry: A refuge for tropical biodiversity after fire. Conserv. Biol. 2000, 14, 325–326. [Google Scholar] [CrossRef]
- Zomer, R.J.; Neufeldt, H.; Xu, J.; Ahrends, A.; Bossio, D.; Trabucco, A.; Van Noordwijk, M.; Wang, M. Global Tree Cover and Biomass Carbon on Agricultural Land: The contribution of agroforestry to global and national carbon budgets. Sci. Rep. 2016, 6, 29987. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.J.; Beck, M.M. Influences of shelterbelts on wildlife management and biology. Agric. Ecosyst. Environ. 1998, 22, 301–335. [Google Scholar]
- Söderström, B.O.; Pärt, T. Influence of landscape scale on farmland birds breeding in semi-natural pastures. Conserv. Biol. 2000, 14, 522–533. [Google Scholar]
- Barrios, E.; Valencia, V.; Jonsson, M.; Brauman, A.; Hairiah, K.; Mortimer, P.E.; Okubo, S. Contribution of trees to the conservation of biodiversity and ecosystem services in agricultural landscapes. Int. J. Biodiv. Sci. Ecosyst. Serv. Manag. 2017, 14, 1–16. [Google Scholar] [CrossRef]
- Donaldson, J.; Nänni, I.; Zachariades, C.; Kemper, J. Effects of habitat fragmentation on pollinator diversity and plant reproductive success in Renosterveld shrub-lands of South Africa. Conserv. Biol. 2002, 5, 1267–1276. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.M.; Steffan-Dewenter, I.; Tscharntke, T. Rain forest promotes trophic interactions and diversity of trap-nesting Hymenoptera in adjacent agroforestry. J. Anim. Ecol. 2006, 75, 315–323. [Google Scholar] [CrossRef]
- Karp, D.S.; Mendenhall, C.D.; Sandi, R.F.; Chaumont, N.; Ehrlich, P.R.; Hadly, E.A.; Daily, G.C. Forest bolsters bird abundance, pest control and coffee yield. Ecol. Lett. 2013, 16, 1339–1347. [Google Scholar] [CrossRef]
- Brandle, J.R.; Hodges, L.; Zhou, X. Windbreaks in sustainable agriculture. Agrofor. Syst. 2004, 61, 65–78. [Google Scholar]
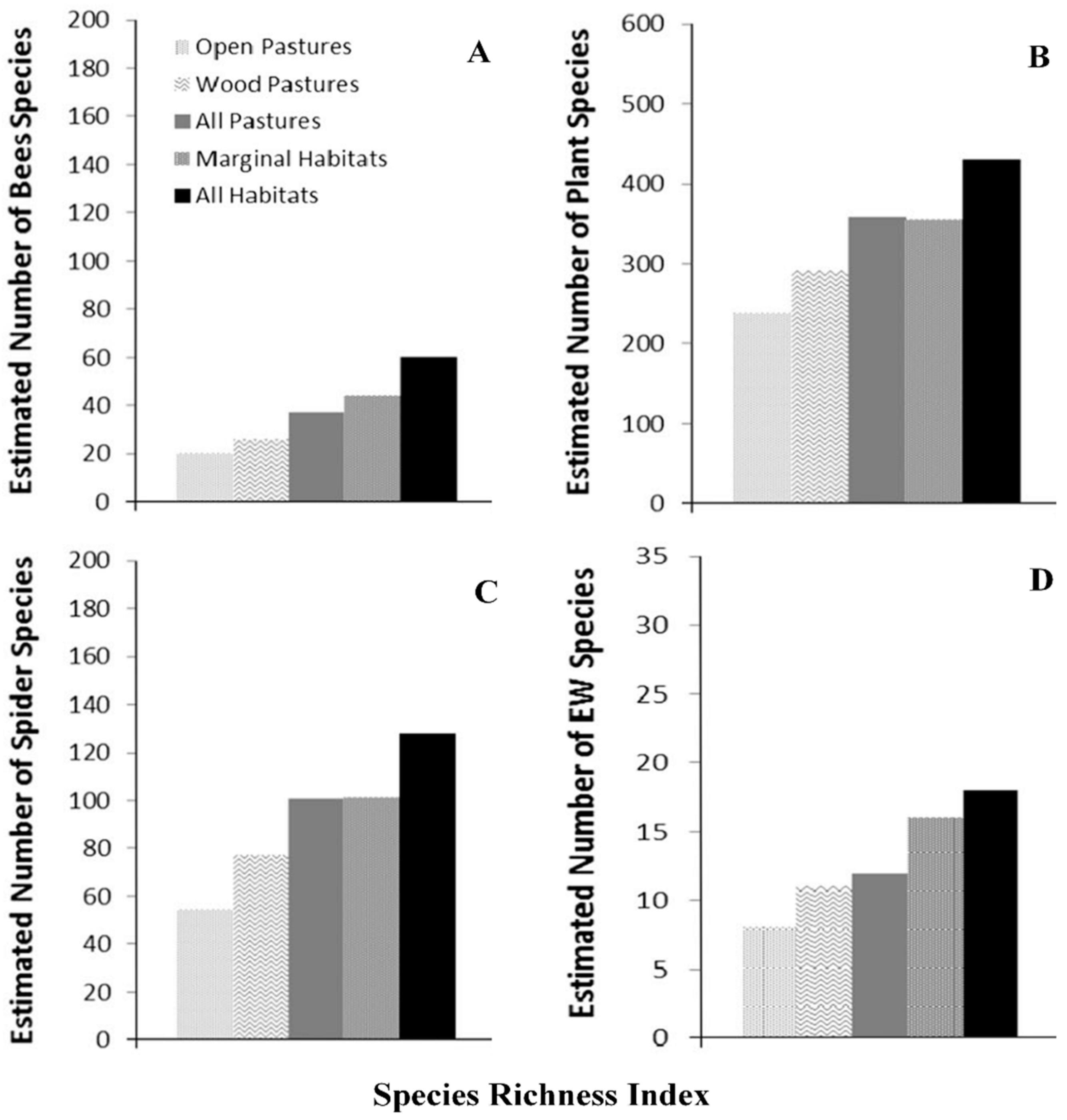
- Moreno, G.; Gonzalez-Bornay, G.; Pulido, F.; Lopez-Diaz, M.L.; Bertomeu, M.; Jua’rez, E.; Diaz, M. Exploring the causes of high biodiversity of Iberian dehesas: The importance of wood pastures and marginal habitats. Agrofor. Syst. 2016, 90, 87–105. [Google Scholar] [CrossRef]
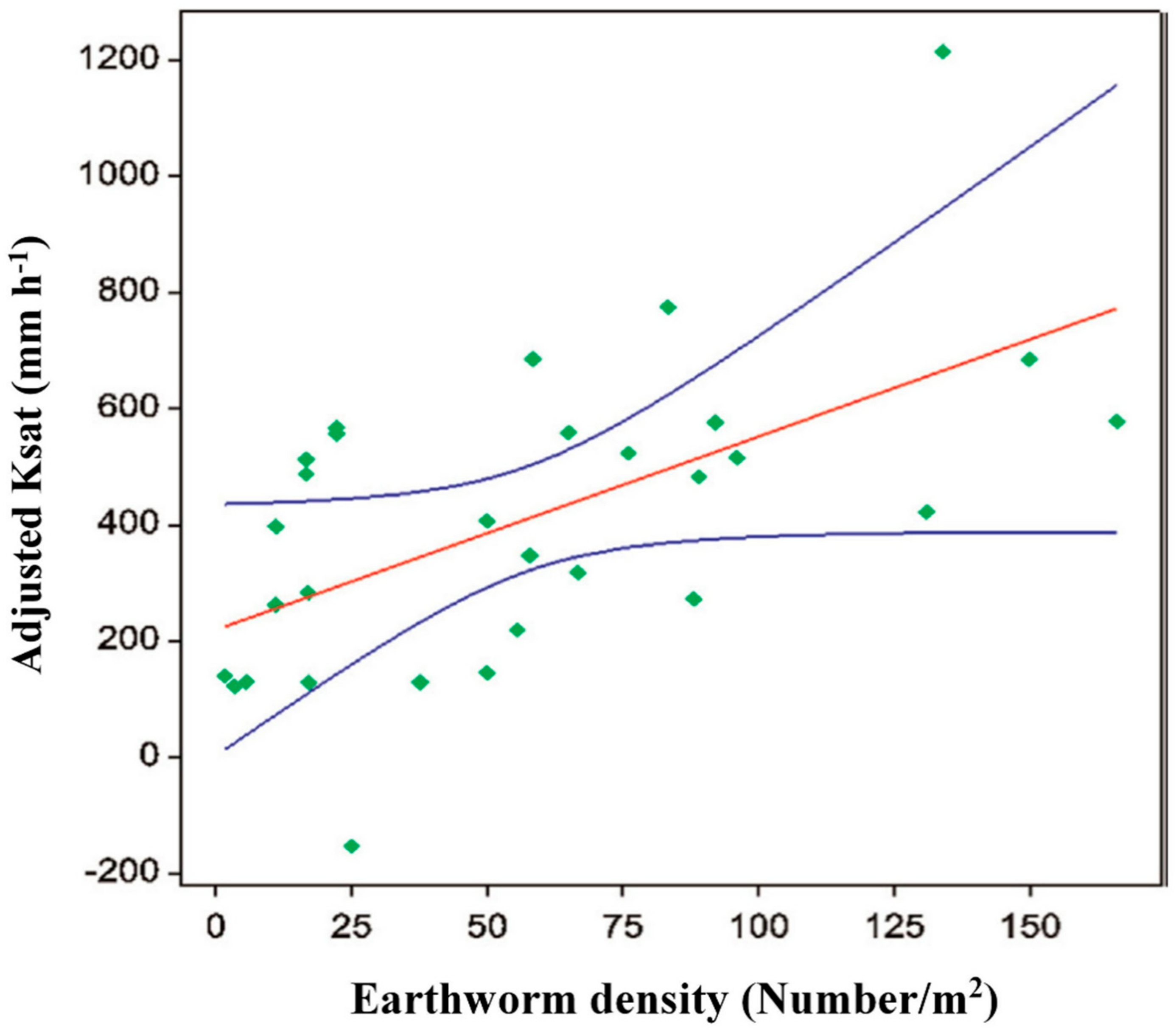
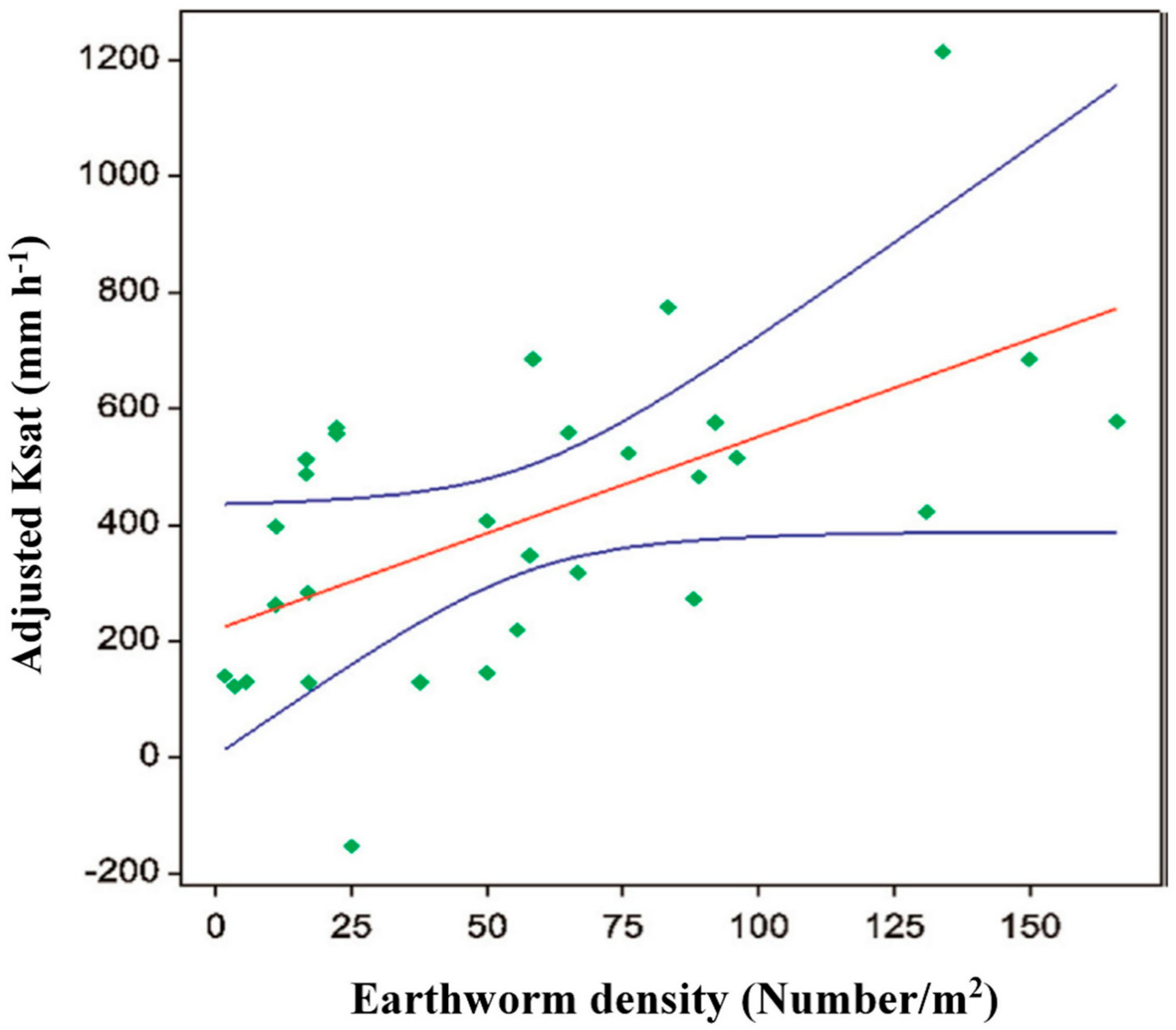
- Price, G.W.; Gordon, A.M. Spatial and temporal distribution of earthworms in a temperate intercropping system in southern Ontario, Canada. Agrofor. Syst. 1999, 44, 141–149. [Google Scholar] [CrossRef]
- Pauli, N.; Oberthur, T.; Barrios, E.; Conacher, A.J. Fine-scale spatial and temporal variation in earthworm surface casting activity in agroforestry fields, western Honduras. Pedobiologia 2010, 53, 127–139. [Google Scholar] [CrossRef]
- Cardinael, R.; Guenet, B.; Chevallier, T.; Dupraz, C.; Cozzi, T.; Chenu, C. High organic inputs explain shallow and deep SOC storage in a long-term agroforestry system–combining experimental and modeling approaches. Biogeosciences 2018, 15, 297–317. [Google Scholar] [CrossRef]
- Spurgeon, D.J.; Keith, A.M.; Schmidt, O.; Lammertsma, D.R.; Faber, J.H. Land-use and land-management change: Relationships with earthworm and fungi communities and soil structural properties. BMC Ecol. 2013, 13, 46. [Google Scholar] [CrossRef] [PubMed]
- Keesstra, S.D.; Bouma, J.; Wallinga, J.; Tittonell, P.; Smith, P.; Cerdà, A.; Montanarella, L.; Quinton, J.N.; Pachepsky, Y.; Van Der Putten, W.H.; et al. The significance of soils and soil science towards realization of the United Nations Sustainable Development Goals. Soil 2016, 2, 111–128. [Google Scholar] [CrossRef]
- Bainard, L.D.; Klironomos, J.N.; Gordon, A.M. Arbuscular mycorrhizal fungi in tree-based intercropping systems: A review of their abundance and diversity. Pedobiologia 2011, 54, 57–61. [Google Scholar] [CrossRef]
- Beuschel, R.; Piepho, H.P.; Joergensen, R.G.; Wachendorf, C. Similar spatial patterns of soil quality indicators in three poplar-based silvo-arable alley cropping systems in Germany. Biol. Fertil. Soils 2019, 55, 1–14. [Google Scholar] [CrossRef]
- Lacombe, S.; Bradley, R.L.; Hamel, C.; Beaulieu, C. Do tree-based intercropping systems increase the diversity and stability of soil microbial communities? Agric. Ecosyst. Environ. 2009, 131, 25–31. [Google Scholar] [CrossRef]
- Chifflot, V.; Rivest, D.; Olivier, A.; Cogliastro, A.; Khasa, D. Molecular analysis of arbuscular mycorrhizal community structure and spores distribution in tree-based intercropping and forest systems. Agric. Ecosyst. Environ. 2009, 131, 32–39. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, M.; Zhou, P.; Fang, Y.; Ji, Y. Impact of tree species on barley rhizosphere-associated fungi in an agroforestry ecosystem as revealed by 18S rDNA PCRDGGE. Agrofor. Syst. 2018, 92, 541–554. [Google Scholar]
- Kremer, R.J.; Kussman, R.D. Soil quality in a pecan–kura clover alley cropping system in the Midwestern USA. Agrofor. Syst. 2011, 83, 213–223. [Google Scholar] [CrossRef]
- Mutabaruka, R.; Mutabaruka, C.; Fernandez, I. Diversity of arbuscular mycorrhizal fungi associated to tree species in semiarid areas of Machakos, Kenya. Arid Land Res. Manag. 2002, 16, 385–390. [Google Scholar] [CrossRef]
- Pande, M.; Tarafdar, J.C. Arbuscular mycorrhizal fungal diversity in neem-based agroforestry systems in Rajasthan. Appl. Soil Ecol. 2004, 26, 233–241. [Google Scholar] [CrossRef]
- Banarjee, S.; Baah-Acheamfour, M.; Carlyle, C.M.; Bissett, A.; Richardon, A.E.; Siddique, T.; Bork, E.W.; Chang, S.X. Determinants of bacterial communities in Canadian agroforestry systems. Environ. Microbiol. 2015, 18, 1805–1816. [Google Scholar] [CrossRef] [PubMed]
- Kremer, R.J.; Hezel, L.F. Soil quality improvement under an ecologically based farming system in northwest Missouri. Renew. Agric. Food Syst. 2013, 28, 245–254. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function: Are there any links? Ecology 2003, 84, 2042–2050. [Google Scholar] [CrossRef]
- Keith, A.M.; Van der Wal, R.; Brooker, R.W.; Osler, G.H.R.; Chapman, S.J.; Burslem, D.F.R.P.; Elson, D.A. Increasing litter species richness reduces variability in a terrestrial decomposer system. Ecology 2008, 89, 2657–2664. [Google Scholar] [CrossRef] [PubMed]
- Rivest, D.; Lorente, M.; Olivier, A.; Messier, C. Soil biochemical properties and microbial resilience in agroforestry systems: Effects on wheat growth under controlled drought and flooding conditions. Sci. Total Environ. 2013, 463, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.; Gupta, S.R.; Singh, G. Soil carbon, microbial activity, and nitrogen availability in a agroforestry systems on moderately alkaline soils in northern India. Appl. Soil Ecol. 2000, 15, 283–294. [Google Scholar] [CrossRef]
- Wojewoda, D.; Russel, S. The impact of a shelterbelt on soil properties an microbial activity in an adjacent corp field. Pol. J. Ecol. 2003, 51, 291–307. [Google Scholar]
- Amador, J.A.; Glucksman, A.M.; Lyons, J.B.; Gorres, J.H. Spatial distribution of soil phosphatase activity within a riparian forest. Soil Sci. 1997, 162, 808–825. [Google Scholar] [CrossRef]
- Boerner, R.E.J.; Decker, K.L.M.; Sutherland, E.K. Prescribed burning effects on soil enzyme activity in a southern Ohio hardwood forest: A landscape-scale analysis. Soil Biol. Biochem. 2000, 32, 899–908. [Google Scholar] [CrossRef]
- Mungai, N.W.; Motavalli, P.P.; Kremer, R.J.; Nelson, K.A. Spatial variation of soil enzyme activities and microbial functional diversity in temperate alley cropping practices. Biol. Fertil. Soils 2006, 42, 129–136. [Google Scholar] [CrossRef]
- Dornbush, M.E. Grasses, litter and their interaction affects microbial biomass and soil enzyme activity. Soil Biol. Biochem. 2007, 39, 2241–2249. [Google Scholar] [CrossRef]
- Udawatta, R.P.; Kremer, R.J.; Adamson, B.W.; Anderson, S.H. Variations in soil aggregate stability and enzyme activities in a temperate agroforestry practice. Appl. Soil Ecol. 2008, 39, 153–160. [Google Scholar] [CrossRef]
- Helgason, B.L.; Walley, F.L.; Germida, J.J. No-till and soil management increases microbial biomass and alters community profiles in soil aggregates. App. Soil Ecol. 2010, 46, 390–397. [Google Scholar] [CrossRef]
- Adhikari, K.; Hartemink, A.E.; Minasny, B.; BouKheir, R.; Greve, M.B.; Greve, M.H. Digital mapping of soil organic carbon contents and stocks in Denmark. PLoS ONE 2014, 9, e105519. [Google Scholar] [CrossRef] [PubMed]
- Udawatta, R.P.; Anderson, S.H. CT-measured pore characteristics of surface and subsurface soils as influenced by agroforestry and grass buffers. Geoderma 2008, 145, 381–389. [Google Scholar] [CrossRef]
- Udawatta, R.P.; Kremer, R.J.; Garrett, H.E.; Anderson, S.H. Soil enzyme activities and physical properties in a watershed managed under agroforestry and row-crop system. Agric. Ecosyst. Environ. 2009, 131, 98–104. [Google Scholar] [CrossRef]
- Paudel, B.; Udawatta, R.P.; Kremer, R.J.; Anderson, S.H. Agroforestry and grass buffer effects on soil quality parameters for grazed pasture and row-crop systems. Appl. Soil Ecol. 2011, 48, 125–132. [Google Scholar] [CrossRef]
- Paudel, B.; Udawatta, R.P.; Kremer, R.J.; Anderson, S.H. Soil quality indicator responses to row crop, grazed pasture, and agroforestry buffer management. Agrofor. Syst. 2012, 84, 311–323. [Google Scholar] [CrossRef]
- Meyers, R.T.; Zak, D.R.; White, D.C.; Peacock, A. Landscape-level patterns of microbial community composition and substrate use in upland forest ecosystems. Soil Sci. Soc. Am. J. 2001, 65, 359–367. [Google Scholar] [CrossRef]
- Weerasekara, C.; Udawatta, R.P.; Jose, S.; Kremer, R.J.; Weerasekara, C. Soil quality differences in a row-crop watershed with agroforestry and grass buffers. Agrofor. Syst. 2016, 90, 829–838. [Google Scholar] [CrossRef]
- Bambrick, A.D.; Whallen, J.K.; Bradley, R.L.; Cogliastro, A.; Gordon, A.M.; Olivier, A.; Thevathasan, N.V. Spatial heterogeneity of organic carbon in tree-based inter cropping systems in Quebec and Ontario, Canada. Agrofor. Syst. 2010, 79, 343–353. [Google Scholar] [CrossRef]
- Bardhan, S.; Jose, S.; Udawatta, R.P.; Fritschi, F. Microbial community diversity in a 21-year old temperate alley cropping system. Agrofor. Syst. 2013, 87, 1031–1041. [Google Scholar] [CrossRef]
- Nii-Annang, S.; Grünewald, H.; Freese, D.; Hüttl, R.F.; Dilly, O. Microbial activity, organic C accumulation and 13C abundance in soils under alley cropping systems after 9 years of recultivation of quaternary deposits. Biol. Fertil. Soils 2009, 45, 531–538. [Google Scholar] [CrossRef]
- Foley, J.A.; Defries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Burgess, P.J. Effects of agroforestry on farm biodiversity in the UK. Scott. For. 1999, 53, 24–27. [Google Scholar]
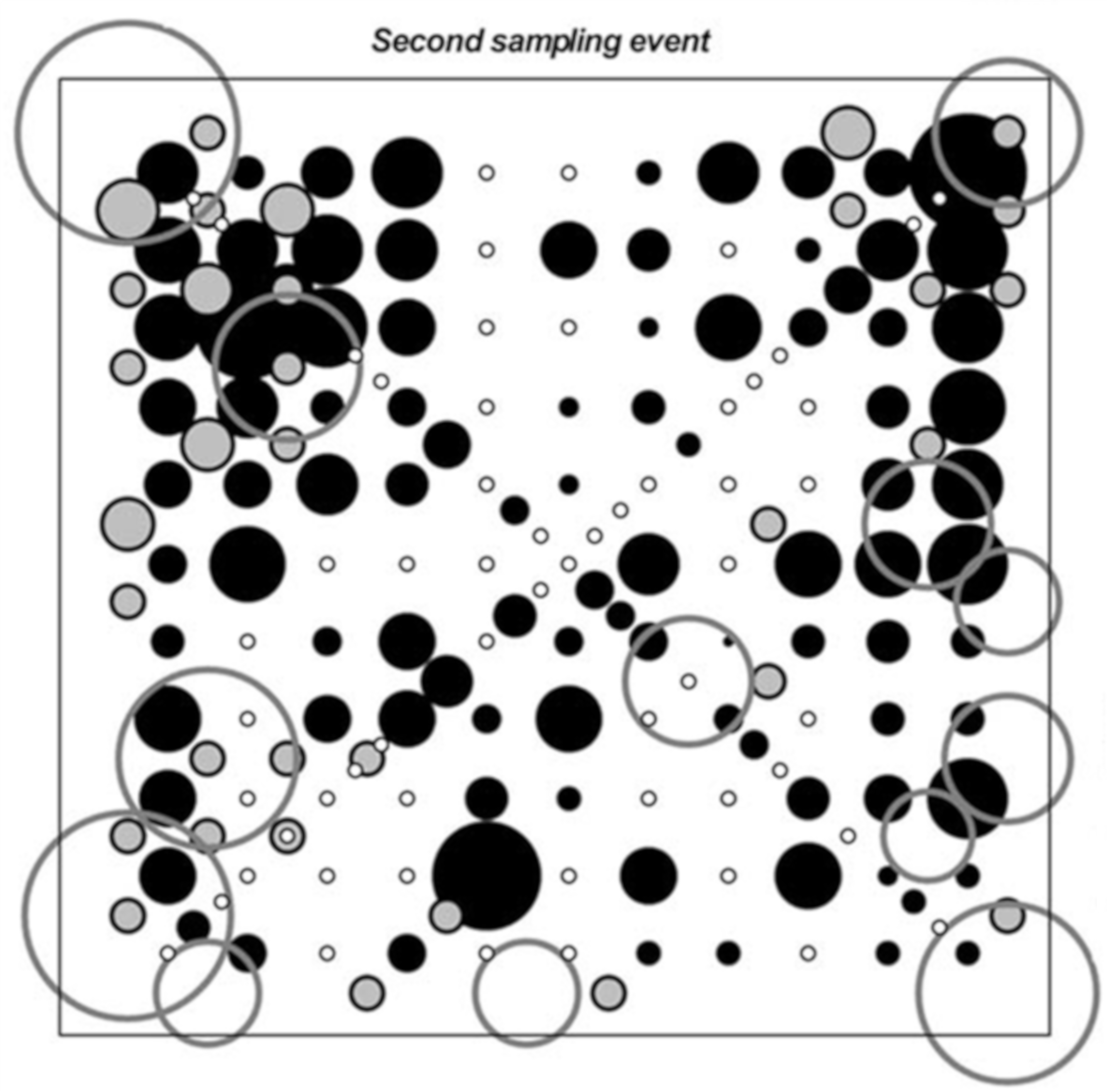
- Stamps, W.T.; Woods, T.W.; Linit, M.J.; Garrett, H.E. Arthropod diversity in alley cropped black walnut (Juglans nigra L.) stands in eastern Missouri, USA. Agrofor. Syst. 2002, 56, 167–175. [Google Scholar] [CrossRef]
- McNeely, J.A. Nature vs. nurture: Managing relationships between forests, agroforestry and wild biodiversity. Agrofor. Syst 2004, 61, 155–165. [Google Scholar]
- Seobi, T.; Anderson, S.H.; Udawatta, R.P.; Gantzer, C.J. Influence of grass and agroforestry buffer strips on soil hydraulic properties for an Albaqualf. Soil Sci. Soc. Am. J. 2005, 69, 893–901. [Google Scholar] [CrossRef]
- Kumar, S.; Anderson, S.H.; Bricknell, L.G.; Udawatta, R.P.; Gantzer, C.J. Soil hydraulic properties influenced by agroforestry and grass buffers for grazed pasture systems. J. Soil Water Conserv. 2008, 63, 224–232. [Google Scholar] [CrossRef]
- Svoma, B.M.; Fox, N.I.; Pallardy, Q.; Udawatta, R.P. Evapotranspiration differences between agroforestry and grass buffer systems. Agric. Water Manag. 2016, 176, 214–221. [Google Scholar] [CrossRef]
- Prober, S.M.; Left, J.W.; Bates, S.T.; Borer, E.T.; Firn, J.; Harpole, W.S.; Lind, E.M.; Seabloom, E.W.; Adler, P.B.; Bakker, J.D.; et al. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 2015, 18, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Kleijn, D.; Kohler, F.; Báldi, A.; Batáry, P.; Concepción, E.D.; Clough, Y.; Díaz, M.; Gabriel, D.; Holzschuh, A.; Knop, E.; Kovács, A. On the relationship between farmland biodiversity and land-use intensity in Europe. Proc. R. Soc. Lond. Biol. Sci. 2008, 276, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Simmons, B.L.; Coleman, D.C. Microbial community response to transition from conventional to conservation tillage in cotton fields. App. Soil Ecol. 2008, 40, 518–528. [Google Scholar] [CrossRef]
- Mathimaran, N.; Ruh, R.; Jama, B.; Verchot, L.; Frossard, E.; Jansa, J. Impact of agricultural management on arbuscular mycorrhizal fungal communities in Ken-yan ferra sol. Agric. Ecosyst. Environ. 2007, 119, 22–32. [Google Scholar] [CrossRef]
- Thiele-Bruhn, S.; Bloem, J.; De Vries, F.T.; Kalbitz, K.; Wagg, C. Linking soil biodiversity and agricultural soil management. Curr. Opin. Environ. Sustain. 2012, 4, 523–528. [Google Scholar] [CrossRef]
- Swallow, B.; Russell, D.; Fay, C. Agroforestry and environmental governance. In World Agroforestry into the Future; Garrity, D., Okono, A., Grayson, M., Parrott, S., Eds.; World Agroforestry Centre: Nairobi, Kenya, 2006; pp. 85–94. [Google Scholar]
- Williams-Guillén, K.; McCann, C.; Sanchez, J.M.; Koontz, F. Resource availability and habitat use by mantled howling monkeys in a Nicaraguan coffee plantation: Can agroforests serve as core habitat for a forest mammal? Anim. Conserv. 2006, 9, 331–338. [Google Scholar] [CrossRef]
- Round, P.D.; Gale, G.A.; Brockelman, W.Y. A comparison of bird communities in mixed fruit orchards and natural forest at Khao Luang, southern Thailand. Biodivers. Conserv. 2006, 15, 2873–2891. [Google Scholar] [CrossRef]
- Bainard, L.D.; Koch, A.M.; Gordon, A.M.; Klironomos, J.N. Temporal and compositional differences of arbuscular mycorrhizal fungal communities in conventional monocropping and tree-based intercropping systems. Soil Biol. Biochem. 2012, 45, 172–180. [Google Scholar] [CrossRef]
- Bainard, L.D.; Kochb, A.M.; Gordon, A.M.; Newmaster, S.G.; Thevathasan, N.V.; Klironomosb, J.N. Influence of trees on the spatial structure of arbuscular mycorrhizal communities in a temperate tree-based intercropping system. Agric. Ecosyst. Environ. 2011, 144, 13–20. [Google Scholar] [CrossRef]
- Bergmeier, E.; Petermann, J.; Schröder, E. Geobotanical survey of wood-pasture habitats in Europe: Diversity, threats and conservation. Biodivers. Conserv. 2010, 19, 2995–3014. [Google Scholar] [CrossRef]
- Fischer, J.; Stott, J.; Law, B.S. The disproportionate value of paddock trees. Biol. Conserv. 2010, 143, 1564–1567. [Google Scholar] [CrossRef]
- Barrios, E.; Sileshi, G.W.; Shepherd, K.; Ferguson, S. Agroforestry and Soil Health: Linking Trees, Soil Biota, and Ecosystem Services. In Soil Ecology and Ecosystem Services, 1st ed.; Wall, D.H., Bardgett, R.D., Behan-Pelletier, V., Herrick, J.E., Jones, H., Ritz, K., Six, J., Strong, D.R., van der Putten, W.H., Eds.; Oxford University Press: Oxford, England, 2012; pp. 315–330. [Google Scholar]
- Sileshi, G.; Mafongoya, P.L. Quantity and quality of organic inputs from coppicing leguminous trees influence abundance of soil macrofauna in maize crops in eastern Zambia. Biol. Fertil. Soils 2007, 43, 333–340. [Google Scholar] [CrossRef]
- Angelsen, A.; Kaimowitz, D. Is agroforestry likely to reduce deforestation? In Agroforestry and Biodiversity Conservation in Tropical Landscapes; Schroth, G., da Fonseca, G.A.B., Harvey, C.A., Gascon, C., Vasconcelos, H.L., Izac, A.M.N., Eds.; Island Press: Washington, DC, USA, 2004; Chapter 5; pp. 87–106. [Google Scholar]
- Lin, C.H.; Lerch, R.N.; Goyne, K.W.; Garrett, H.E. Reducing herbicides and veterinary antibiotic losses from agroecosystems using vegetative buffers. J. Environ. Qual. 2011, 40, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Chu, B.; Goyne, K.W.; Anderson, S.H.; Lin, C.H.; Udawatta, R.P. Veterinary antibiotic sorption to agroforestry buffer, grass buffer, and cropland soils. Agrofor. Syst. 2010, 79, 67–80. [Google Scholar] [CrossRef]
- Lerat, S.; Lapointe, L.; Gutjahr, S.; Piche, Y.; Vierheilig, H. Carbon partitioning in a split-root system of arbuscular mycorrhizal plants is fungal and plant species dependent. New Phytol. 2003, 157, 589–595. [Google Scholar] [CrossRef]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
P. Udawatta, R.; Rankoth, L.; Jose, S. Agroforestry and Biodiversity. Sustainability 2019, 11, 2879. https://doi.org/10.3390/su11102879
P. Udawatta R, Rankoth L, Jose S. Agroforestry and Biodiversity. Sustainability. 2019; 11(10):2879. https://doi.org/10.3390/su11102879
Chicago/Turabian StyleP. Udawatta, Ranjith, Lalith Rankoth, and Shibu Jose. 2019. "Agroforestry and Biodiversity" Sustainability 11, no. 10: 2879. https://doi.org/10.3390/su11102879
APA StyleP. Udawatta, R., Rankoth, L., & Jose, S. (2019). Agroforestry and Biodiversity. Sustainability, 11(10), 2879. https://doi.org/10.3390/su11102879