Effects of Artificial Light Treatments on Growth, Mineral Composition, Physiology, and Pigment Concentration in Dieffenbachia maculata “Compacta” Plants

 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
2.2. Experimental Design and Treatments
2.3. Growth Parameters
2.4. Analysis of Mineral Elements
2.5. Physiological Parameters
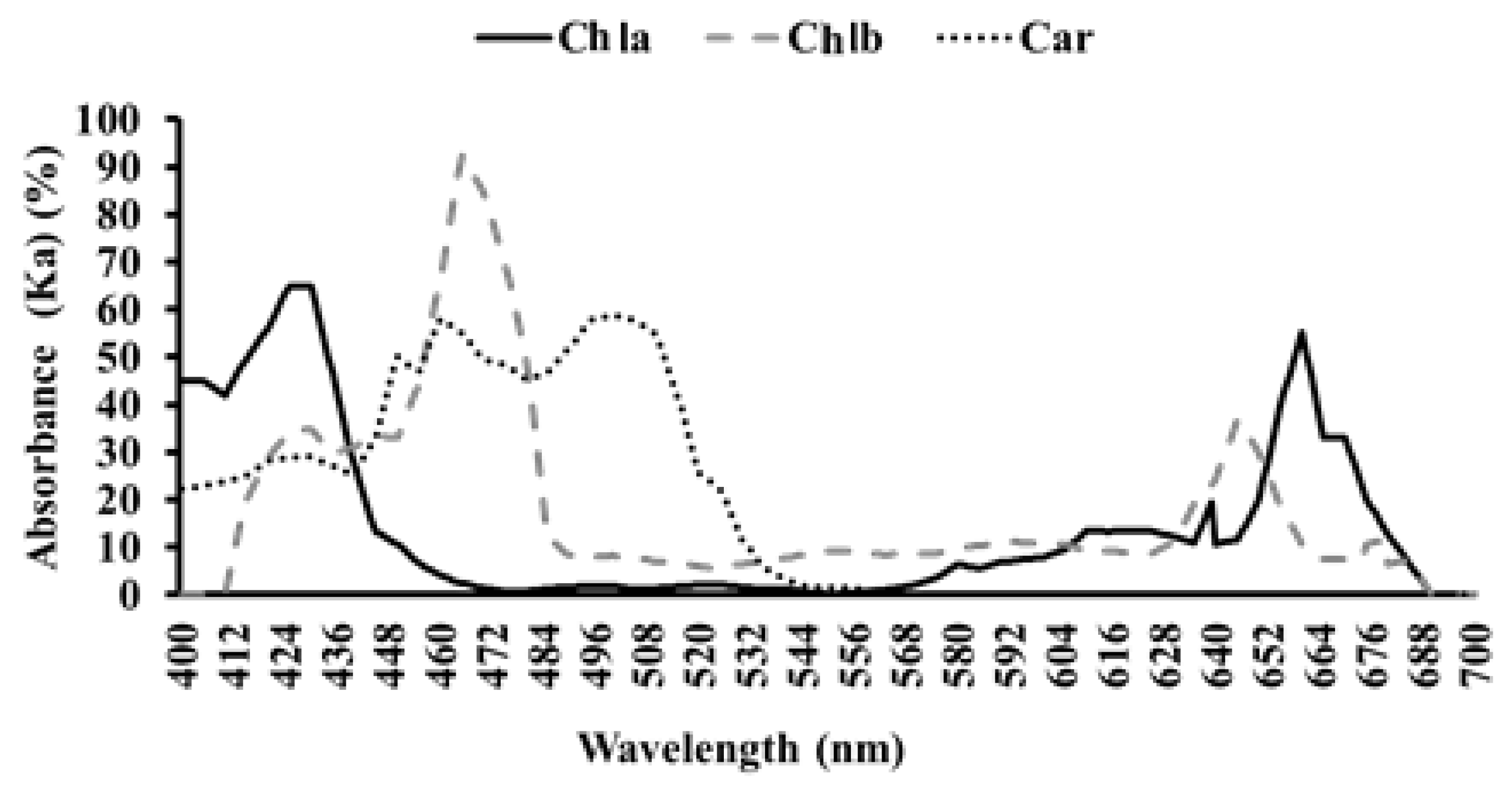
2.6. Pigment Concentrations
2.7. Plant´s Harvesting Lamp PAR Model
2.8. Statistical Analysis
3. Results
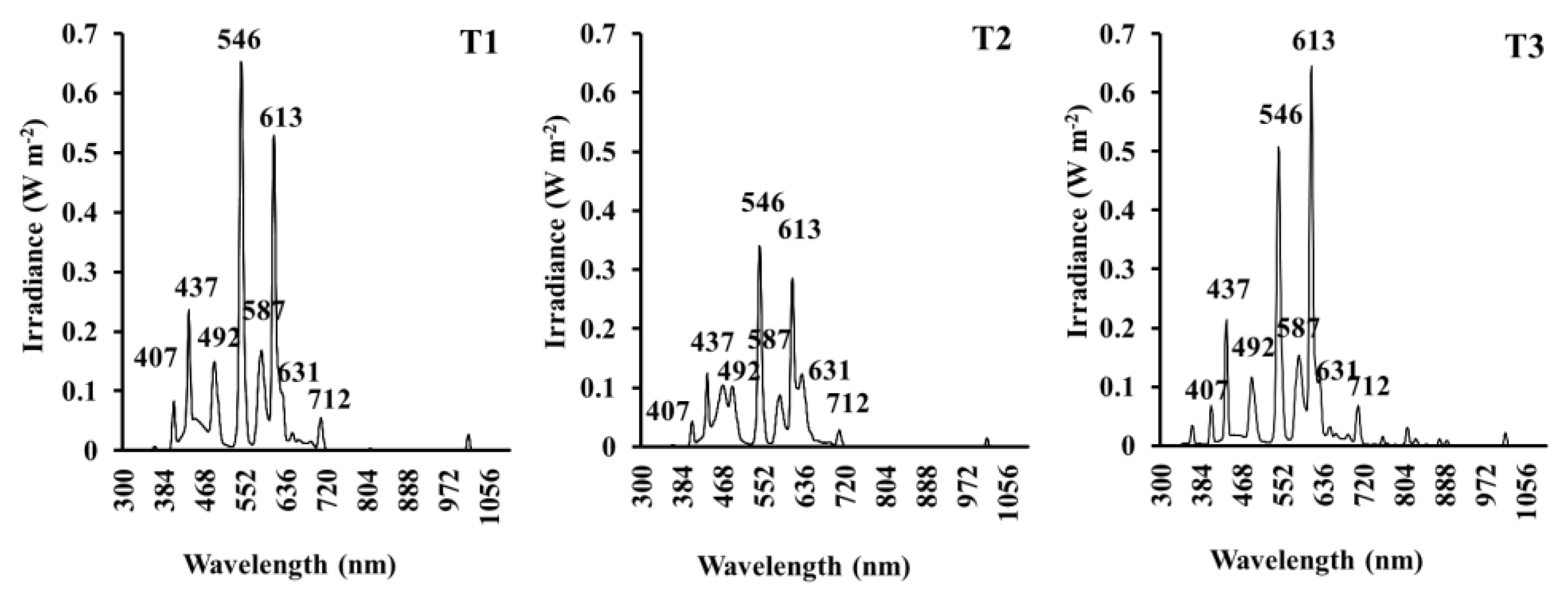
3.1. Spectral Distribution
3.2. Growth Parameters
3.3. Nutrient Concentrations
3.4. Physiological Parameters
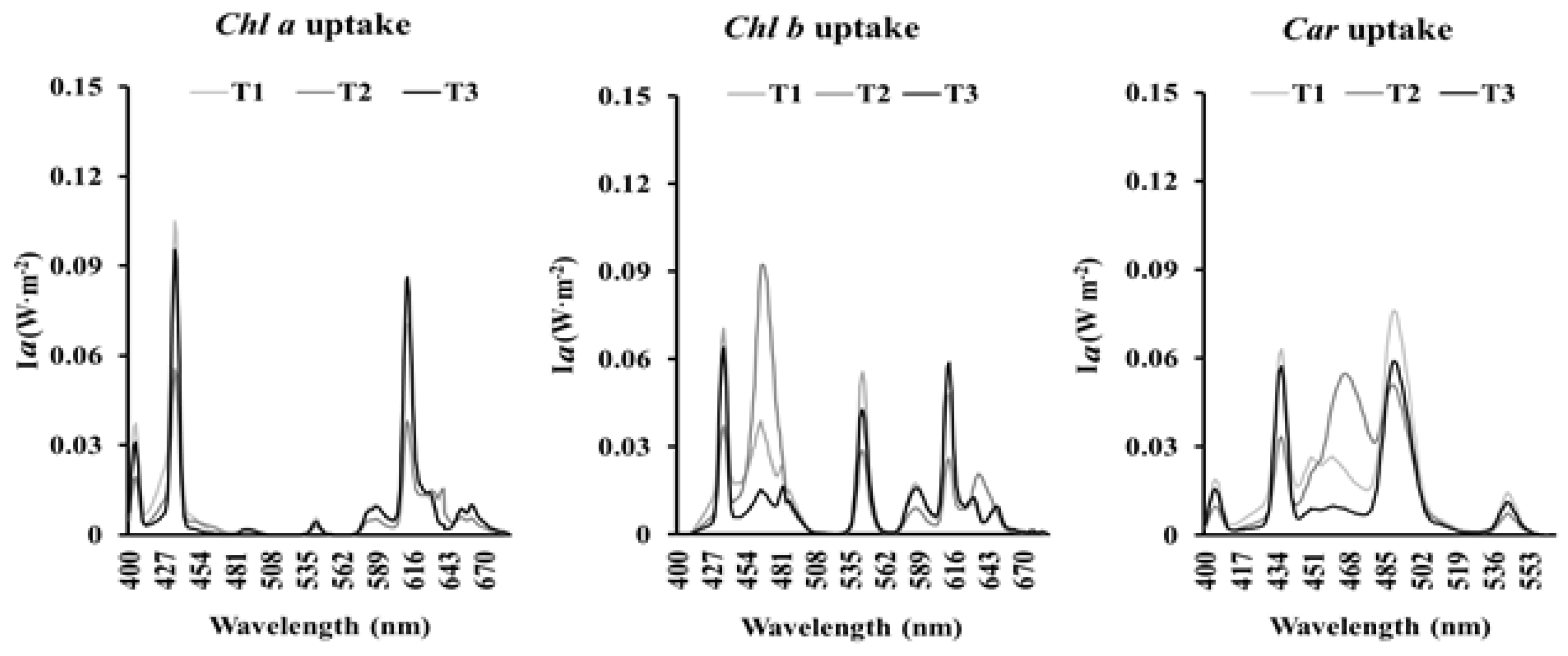
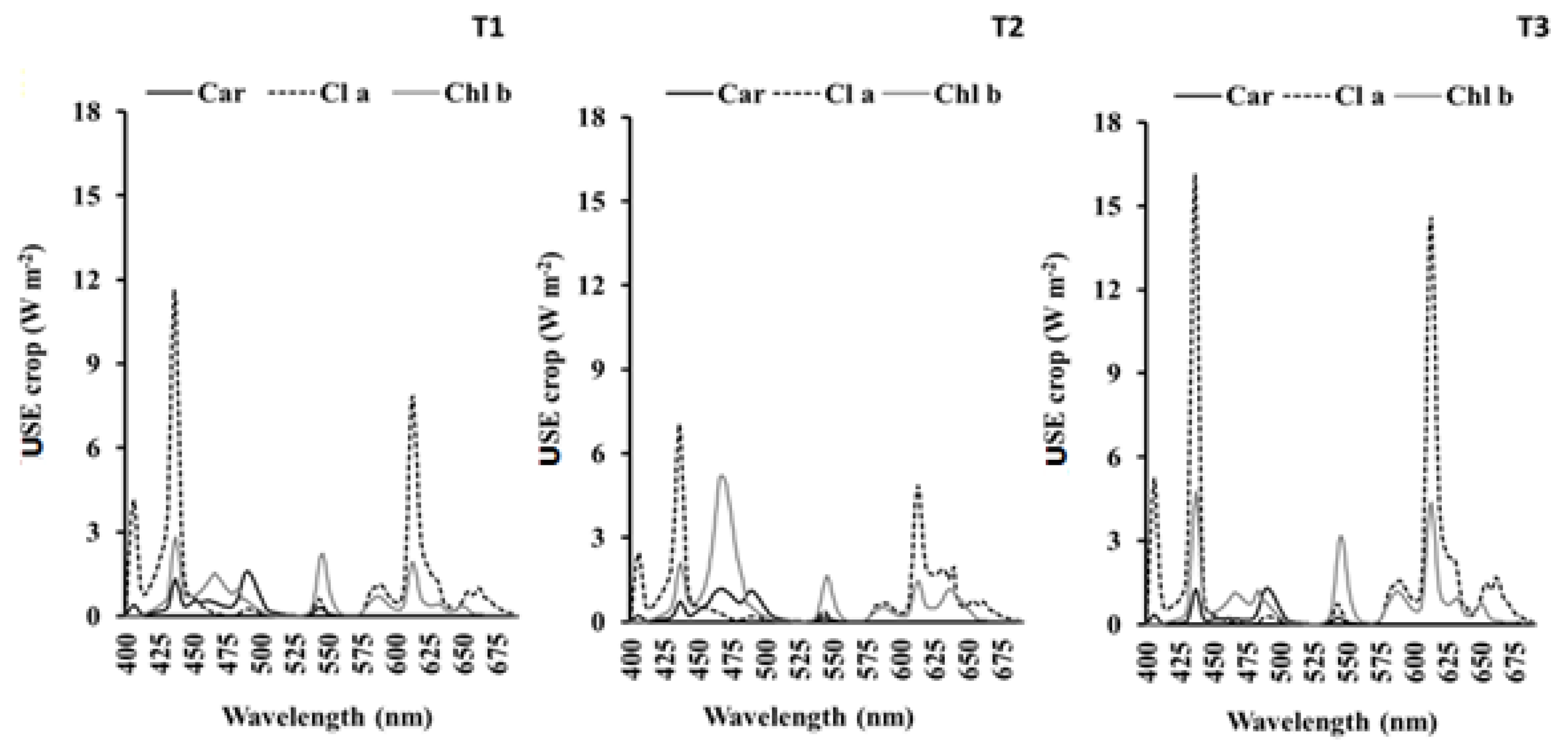
3.5. Photosynthetic Parameters
3.6. Plant’s Harvesting Lamp PAR Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lütken, H.; Clarke, J.L.; Müller, R. Genetic engineering and sustainable production of ornamentals: Current status and future directions. Plant Cell Rep. 2012, 31, 1141–1157. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Llanderal, A.; Pestana, M.; Correia, P.J.; Lao, M.T. Tolerance mechanisms of three potted ornamental plants grown under moderate salinity. Sci. Hortic. 2016, 201, 84–91. [Google Scholar] [CrossRef]
- Gangadhar, B.H.; Mishra, R.K.; Pandian, G.; Park, S.W. Comparative study of color, pungency, and biochemical composition in chili pepper (Capsicum annuum) under different light-emitting diode treatments. HortScience 2012, 47, 1729–1735. [Google Scholar] [CrossRef]
- Folta, K.M.; Childers, K.S. Light as a growth regulator: Controlling plant biology with narrow-bandwidth solid-state lighting systems. HortScience 2008, 43, 1957–1964. [Google Scholar] [CrossRef]
- Stutte, G.W. Light-emitting diodes for manipulating the phytochrome apparatus. HortScience 2009, 44, 231–234. [Google Scholar] [CrossRef]
- Bourget, M.C. An introduction to light emitting diodes. HortScience 2008, 43, 1944–1946. [Google Scholar] [CrossRef]
- Morrow, R.C. LED lighting in horticulture. HortScience 2008, 43, 1947–1950. [Google Scholar] [CrossRef]
- Samouliene, G.; Sirtautas, R.; Brazaitytė, A.; Sakalauskaitė, J.; Sakalauskienė, S.; Duchovskis, P. The impact of red and blue light-emitting diode illumination on radish physiological indices. Open Life Sci. 2011, 6, 821–828. [Google Scholar] [CrossRef]
- Pfündel, E.; Baake, E. A quantitative description of fluorescence excitation spectra in intact bean leaves greened under intermittent light. Photosynth. Res. 1990, 26, 19–28. [Google Scholar]
- Whitelam, G.; Halliday, K. Light and Plant Development; Blackwell Publishing: Oxford, UK, 2007. [Google Scholar]
- Sæbø, A.; Krekling, T.; Appelgren, M. Light quality affects photosynthesis and leaf anatomy of birch plantlets in vitro. Plant Cell Tissue Organ. Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Elsheikh, A.M.; Daffalla, H.M.; Khalfala, M.M. In vitro micropropagation of the ornamental plant Dieffenbachia: A review. Univers. J. Plant Sci. 2013, 1, 91–99. [Google Scholar]
- Heo, J.-W.; Lee, Y.-B.; Kim, D.-E.; Chang, Y.S.; Chun, C.-H. Effects of supplementary LED lighting on growth and biochemical parameters in Dieffenbachia amoena ‘Camella’ and Ficus elastica ‘Melany’. Korean J. Hortic. Sci. Technol. 2010, 1, 51–58. [Google Scholar]
- Baille, A.; González-Real, M.M.; López, J.C.; Cabrera, J.; Pérez-Parra, J. Characterization of the solar diffuse component under “Parral” plastic greenhouses. Acta Hortic. 2003, 614, 341–346. [Google Scholar] [CrossRef]
- Steiner, A. A universal method for preparing nutrient solution of a certain desired composition. Plant Soil 1961, 15, 134–154. [Google Scholar] [CrossRef]
- Ryser, P.; Lambers, H. Root and leaf attributes accounting for the performance of fast- and slow-growing grasses at different nutrient supply. Plant Soil 1995, 170, 251–265. [Google Scholar] [CrossRef]
- Ben Amor, N.; Ben Hamed, K.; Debez, A.; Grignon, C.; Abdelly, C. Physiological and antioxidant responses of the perennial halophyte Crithmum maritimum to salinity. Plant Sci. 2005, 168, 889–899. [Google Scholar] [CrossRef]
- Csáky, A.; Martínez-Grau, M.A. Técnicas Experimentales en Síntesis Orgánica; Síntesis: Madrid, Spain, 1998. [Google Scholar]
- Krom, M.D. Spectrophotometric determination of ammonia: Study of a modified Berthelot reaction using salicylate and dicholoroisocyanurate. Analyst 1980, 105, 305–316. [Google Scholar] [CrossRef]
- Hogue, E.; Wilcow, G.E.; Cantliffe, D.J. Effect of soil P on phosphate fraction in tomato leaves. J. Am. Soc. Hortic. Sci. 1970, 95, 174–176. [Google Scholar]
- Lachica, M.; Aguilar, A.; Yanez, J. Análisis foliar: Métodos utilizados en la estación experimental del Zaidín. Anal. Edafol. Agrobiol. 1973, 32, 1033–1047. [Google Scholar]
- Irigoyen, J.J.; Emerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Tien, T.M.; Gaskin, M.H.; Hubbell, D.H. Plant growth substance produced by Azospirillum brasilense and their effect on the growth of pearl millet (Pennisetum americanun L.). App. Environ. Microbiol. 1979, 37, 219–226. [Google Scholar]
- Wellburn, A. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvent with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Whitmarsh, J.; Govindjee, J.A. The photosynthetic process. In Concepts in Photobiology: Photosynthesis and Photomorphogenesis; Singhal, G.S., Renger, G., Sopory, S.K., Irrgang, K.D., Eds.; Narosa Publishers: New Delhi, India; Kluwer Academic: Norwell, MA, USA, 1999; pp. 11–51. [Google Scholar]
- McNellis, T.W.; Deng, X.W. Light control of seedling morphogenetic pattern. Plant Cell 1995, 7, 1749–1761. [Google Scholar] [CrossRef]
- Randall, W.C.; Lopez, R.G. Comparison of supplemental lighting from high-pressure sodium lamps and light-emitting diodes during bedding plant seedling production. HortScience 2014, 49, 589–595. [Google Scholar] [CrossRef]
- Terfa, M.T.; Poudel, M.S.; Roro, A.G.; Gislerød, H.R.; Olsen, J.E.; Torre, S. Light emitting diodes with a high proportion of blue light affects external and internal quality parameters of pot roses differently than the traditional high-pressure sodium lamp. Acta Hortic. 2012, 956, 635–642. [Google Scholar] [CrossRef]
- Terfa, M.T.; Solhaug, K.A.; Gislerød, H.R.; Olsen, J.E.; Torre, S. A high proportion of blue light increases the photosynthesis capacity and leaf formation rate of Rosa hybrid but does not affect time to flower opening. Physiol. Plant. 2012, 148, 146–159. [Google Scholar] [CrossRef]
- Zheng, L.; Van Labeke, M.C. Chrysanthemum morphology, photosynthetic efficiency and antioxidant capacity are differentially modified by light quality. J. Plant Physiol. 2017, 213, 66–74. [Google Scholar] [CrossRef]
- Currey, C.J.; Lopez, R.G. Biomass accumulation and allocation, photosynthesis, and carbohydrate status of new Guinea impatiens, Geranium, and Petunia cuttings are affected by photosynthetic daily light integral during root development. J. Am. Soc. Hortic. Sci. 2015, 140, 542–549. [Google Scholar] [CrossRef]
- Wollaeger, H.M.; Runkle, E.S. Growth and acclimation of impatiens, salvia, petunia, and tomato seedlings to blue and red light. HortScience 2015, 50, 522–529. [Google Scholar] [CrossRef]
- Jimenez, S.; Plaza, B.M.; Contreras, J.I.; Segura, M.L.; Lao, M.T. Water consumption modelling in Dieffenbachia amoena ‘Tropic Snow’. Acta Hortic. 2007, 813, 685–690. [Google Scholar] [CrossRef]
- Inoue, S.I.; Kinoshita, T. Blue light regulation of stomatal opening and the plasma membrane H+-ATPase. Plant Physiol. 2017, 174, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Mills, H.A.; Jones, J.J.B. Plant Analysis Handbook II; MicroMacro Publishing, Inc.: Athens, GA, USA, 1996; p. 422. [Google Scholar]
- Segura, M.L.; Rodríguez, J.M.; Jiménez, S.; López, J.; Carrasco, I. Dieffenbachia growth responses and N leaching from container-grown plants fertilized by several methods. Acta Hortic. 2004, 697, 387–391. [Google Scholar] [CrossRef]
- Evans, C.A.; Miller, E.K.; Friedland, A.J. Effect of nitrogen and light on nutrient concentrations and associated physiological responses in birch and fir seedlings. Plant Soil 2001, 236, 197–207. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Sams, C.E. Increases in shoot tissue pigments, glucosinolates, and mineral elements in sprouting broccoli after exposure to short-duration blue light from light emitting diodes. J. Am. Soc. Hortic. Sci. 2013, 138, 31–37. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Yanagisawa, S. Light signalling-induced regulation of nutrient acquisition and utilisation in plants. Semin. Cell Dev. Biol. 2018, 83, 123–132. [Google Scholar] [CrossRef]
- Fallovo, C.; Colla, G.; Schreiner, M.; Krumbein, A.; Schwarz, D. Effect of nitrogen form and radiation on growth and mineral concentration of two Brassica species. Sci. Hortic. 2009, 123, 170–177. [Google Scholar] [CrossRef]
- Almansa, E.M.; Espín, A.; Chica, R.M.; Lao, M.T. Nutritional response of seedling tomato plants under different lighting treatments. J. Plant Nutr. 2017, 40, 467–475. [Google Scholar] [CrossRef]
- Azuara, M.P.; Aparicio, P.J. Spectral dependence of photoregulation of inorganic nitrogen metabolism in Chlamydomonas reinhardtii. Plant Physiol. 1985, 77, 95–98. [Google Scholar] [CrossRef]
- Babourina, O.; Newman, I.; Shabala, S. Blue light-induced kinetics of H+ and Ca2+ fluxes in etiolated wild-type and phototropin mutant Arabidopsis seedlings. PNAS 2002, 99, 2433–2438. [Google Scholar] [CrossRef]
- Ben Rejeb, K.; Abdelly, C.; Savouré, A. How reactive oxygen species and proline face stress together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef]
- Zheng, L.; Van Labeke, M.C. Long-term effects of red-and blue-light emitting diodes on leaf anatomy and photosynthetic efficiency of three ornamental pot plants. Front. Plant Sci. 2017, 8, 917. [Google Scholar] [CrossRef]
- Almansa, E.M.; Espín, A.; Chica, R.M.; Lao, M. T Changes in endogenous auxin concentration in cultivars of tomato seedlings under artificial light. HortScience 2011, 46, 698–704. [Google Scholar] [CrossRef]
- Fukuda, N.; Yoshida, T.; Senaha, C.; Olsen, J.E.; Jikumaru, Y.; Kamiya, Y. Short main shoot length and inhibition of floral bud development under red light can be recovered by application of gibberellin and cytokinin. Acta Hortic. 2012, 956, 215–222. [Google Scholar] [CrossRef]
- Behringer, F.J.; Davies, P.J. Indole-3-acetic acid levels after phytochrome-mediated changes in the stem elongation rate of dark-and light-grown Pisum seedlings. Planta 1992, 188, 85–92. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Emery, R.J.N.; Pharis, R.P.; Reid, D.M. Uncoupling light quality from light irradiance effects in Helianthus annuus shoots: Putative roles for plant hormones in leaf and internode growth. J. Exp. Bot. 2007, 58, 2145–2157. [Google Scholar] [CrossRef]
- Halliday, K.J.; Martinez-Garcia, J.F.; Josse, E.M. Integration of light and auxin signaling. Cold Spring Harb. Perspect. Biol. 2009, 1, a001586. [Google Scholar] [CrossRef] [PubMed]
- Kowallik, W. Blue light effect on carbohydrate and protein metabolism. In Blue Light Responses: Phenomena and Occurrence in Plants and Microorganisms; Senger, H., Ed.; CRC Press: Boca Raton, FL, USA, 1987; Volume II, pp. 7–16. [Google Scholar]
- Bondada, B.R.; Syvertsen, J.P. Leaf chlorophyll, net gas exchange and chloroplast ultrastructure in citrus leaves of different nitrogen status. Tree Physiol. 2003, 23, 553–559. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z. The effects of different light qualities on rapeseed (Brassica napus L. ) plantlet growth and morphogenesis in vitro. Sci. Hortic. 2013, 150, 117–124. [Google Scholar]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2002; p. 690. [Google Scholar]
- Randall, W.C.; Lopez, R.G. Comparison of bedding plant seedlings grown under sole-source light-emitting diodes (LEDs) and greenhouse supplemental lighting from LEDs and high-pressure sodium lamps. HortScience 2015, 50, 705–713. [Google Scholar] [CrossRef]
- Bergstrand, K.J.; Mortensen, L.M.; Suthaparan, A.; Gislerød, H.R. Acclimatisation of greenhouse crops to differing light quality. Sci Hortic. 2016, 204, 1–7. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Paradiso, R.; Meinen, E.; Snel, J.F.; De Visser, P.; Van Ieperen, W.; Hogewoning, S.W.; Marcelis, L.F. Spectral dependence of photosynthesis and light absorptance in single leaves and canopy in rose. Sci. Hortic. 2011, 127, 548–554. [Google Scholar] [CrossRef]
- Young, A.J.; Pallett, K.E. Carotenoids. In Antioxidants in Higher Plants; CRC Press: Boca Raton, FL, USA, 2017; pp. 59–89. [Google Scholar]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2010, 62, 869–882. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Irradiance (W·m−2 and µmol·m−2·s−1) | |||
|---|---|---|---|
| Spectral Region (nm) | T1 | T2 | T3 |
| UV (300–400) | 0.18/0.49 | 0.09/0.25 | 0.41/1.22 |
| B (400–500) | 6.68/25.40 | 5.74/22.11 | 4.41/16.78 |
| R (600–700) | 7.57/39.32 | 5.75/30.02 | 8.67/44.99 |
| FR (700–800) | 0.64/3.81 | 0.33/1.94 | 0.90/5.45 |
| PAR (400–700) | 24.50/112.38 | 16.92/77.37 | 21.54/101.22 |
| NIR (700–1100) | 0.92/6.09 | 0.46/3.06 | 1.60/10.56 |
| TOTAL (300–1100) | 25.38/118.96 | 17.35/80.68 | 23.38/113.00 |
| PAR:TOTAL | 0.97/0.94 | 0.97/0.96 | 0.92/0.90 |
| PAR:NIR | 26.58/18.45 | 36.49/25.25 | 13.45/9.58 |
| B:R | 0.88/0.65 | 1.00/0.74 | 0.51/0.37 |
| B:FR | 10.45/6.66 | 17.62/11.39 | 4.87/3.08 |
| R:FR | 11.84/10.31 | 17.65/15.46 | 9.58/8.25 |
| Growth Parameters | Light Treatments | ||
|---|---|---|---|
| T1 | T2 | T3 | |
| RDW (g) | 0.90 ± 0.06 b | 1.14 ± 0.07 a | 0.86 ± 0.06 b |
| SDW (g) | 1.54 ± 0.08 a | 1.30 ± 0.07 b | 1.33 ± 0.07 b |
| LDW (g) | 2.61 ± 0.14 b | 2.05 ± 0.12 c | 3.03 ± 0.17 a |
| TDW (g) | 5.04 ± 0.25 a | 4.49 ± 0.20 b | 5.22 ± 0.23 a |
| RWR (-) | 0.18 ± 0.01 a | 0.25 ± 0.02 b | 0.16 ± 0.02 b |
| SWR (-) | 0.31 ± 0.02 a | 0.29 ± 0.01 b | 0.25 ± 0.01 c |
| LWR (-) | 0.52 ± 0.02 b | 0.46 ± 0.02 c | 0.58 ± 0.02 a |
| WCr (-) | 0.94 ± 0.01 a | 0.94 ± 0.01 a | 0.95 ± 0.01 a |
| WCs (-) | 0.92 ± 0.01 a | 0.92 ± 0.01 a | 0.93 ± 0.01 a |
| WCl (-) | 0.84 ± 0.01 a | 0.84 ± 0.01 a | 0.85 ± 0.01 a |
| Light Treatments | ||||
|---|---|---|---|---|
| T1 | T2 | T3 | ||
| N | R | 17.07 ± 1.66 a | 18.31 ± 1.78 a | 16.92 ± 1.58 a |
| S | 20.22 ± 1.96 a | 14.99 ± 1.44 b | 15.49 ± 1.51 b | |
| L | 19.30 ± 1.81 a | 16.25 ± 1.66 b | 16.01 ± 1.57 b | |
| P | R | 1.83 ± 0.16 a | 1.72 ± 0.16 a | 1.68 ± 0.17 a |
| S | 2.43 ± 0.21 a | 2.33 ± 0.20 a | 2.30 ± 0.20 a | |
| L | 2.78 ± 0.26 a | 2.60 ± 0.24 a | 2.56 ± 0.26 a | |
| K | R | 37.76 ± 3.63 a | 25.34 ± 2.46 b | 24.75 ± 2.37 b |
| S | 49.97 ± 4.82 a | 51.42 ± 4.97 a | 48.79 ± 4.72 a | |
| L | 54.17 ± 5.32 a | 43.35 ± 4.28 b | 43.17 ± 4.40 b | |
| Physiological Parameters | Light Treatments | ||
|---|---|---|---|
| T1 | T2 | T3 | |
| Proline (µg·g−1·FW) | 194.59 ± 18.53 a | 138.27 ± 12.79 b | 135.96 ± 12.53 b |
| TSS (mg·g−1·FW) | 3.99 ± 0.32 c | 5.26 ± 0.45 b | 8.16 ± 0.75 a |
| Starch (mg·g−1·FW) | 6.02 ± 0.51 c | 7.67 ± 0.75 b | 10.62 ± 1.01 a |
| IAA (mg·g−1·FW) | 4.11 ± 0.33 a | 3.09 ± 0.25 b | 2.45 ± 0.23 c |
| Parameter | Light Treatments | ||
|---|---|---|---|
| T1 | T2 | T3 | |
| Chl a (mg·g−1·FW) | 0.85 ± 0.08 b | 1.07 ± 0.04 a | 1.01 ± 0.03 a |
| Chl b (mg·g−1·FW) | 0.31 ± 0.03 b | 0.48 ± 0.03 a | 0.45 ± 0.03 a |
| Chl (a + b) (mg·g−1·FW) | 1.16 ± 0.15 b | 1.55 ± 0.10 a | 1.46 ± 0.10 a |
| Car (mg·g−1·FW) | 0.14 ± 0.01 a | ± 0.01 a | 0.13 ± 0.01 a |
| USEChla | USEChlb | USECar | |
|---|---|---|---|
| T1 | 3.10 ± 0.38 b | 1.46 ± 0.21 b | 0.54 ± 0.16 a |
| T2 | 2.75 ± 0.28 b | 2.49 ± 0.29 a | 0.72 ± 0.18 a |
| T3 | 5.04 ± 0.67 a | 2.52 ± 0.28 a | 0.62 ± 0.13 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Caparrós, P.; Almansa, E.M.; Chica, R.M.; Lao, M.T. Effects of Artificial Light Treatments on Growth, Mineral Composition, Physiology, and Pigment Concentration in Dieffenbachia maculata “Compacta” Plants. Sustainability 2019, 11, 2867. https://doi.org/10.3390/su11102867
García-Caparrós P, Almansa EM, Chica RM, Lao MT. Effects of Artificial Light Treatments on Growth, Mineral Composition, Physiology, and Pigment Concentration in Dieffenbachia maculata “Compacta” Plants. Sustainability. 2019; 11(10):2867. https://doi.org/10.3390/su11102867
Chicago/Turabian StyleGarcía-Caparrós, Pedro, Eva María Almansa, Rosa María Chica, and María Teresa Lao. 2019. "Effects of Artificial Light Treatments on Growth, Mineral Composition, Physiology, and Pigment Concentration in Dieffenbachia maculata “Compacta” Plants" Sustainability 11, no. 10: 2867. https://doi.org/10.3390/su11102867
APA StyleGarcía-Caparrós, P., Almansa, E. M., Chica, R. M., & Lao, M. T. (2019). Effects of Artificial Light Treatments on Growth, Mineral Composition, Physiology, and Pigment Concentration in Dieffenbachia maculata “Compacta” Plants. Sustainability, 11(10), 2867. https://doi.org/10.3390/su11102867