Growth and Distribution of Maize Roots in Response to Nitrogen Accumulation in Soil Profiles after Long-Term Fertilization Management on a Calcareous Soil

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Fertilization and Treatments
2.3. Sampling and Measurement
2.4. Calculation
2.5. Statistical Analysis
3. Results
3.1. Grain Yield, Dry Matter Accumulation, and N Uptake in Maize Shoots
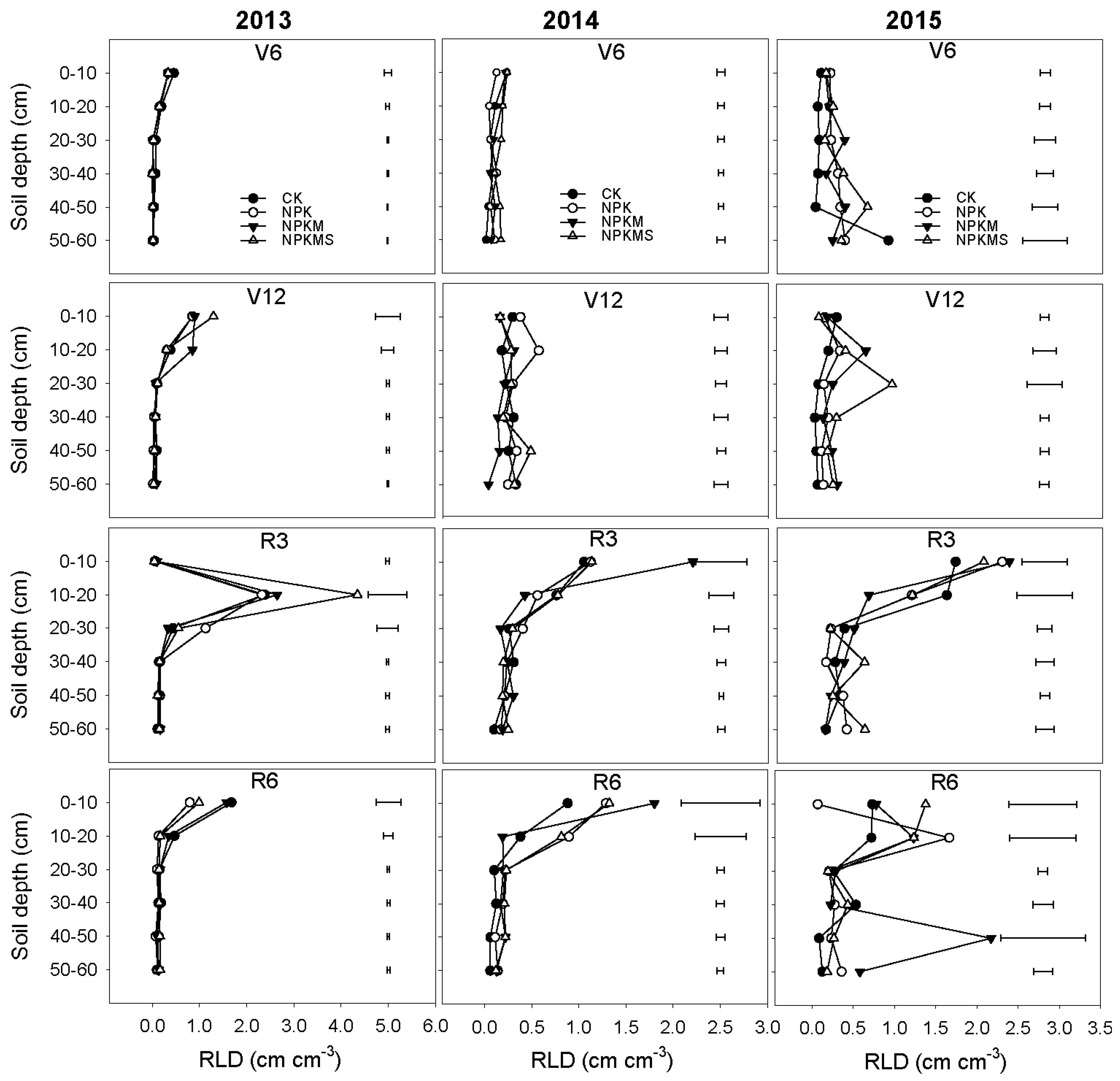
3.2. Distribution of Total Root Length Density
3.3. Root Distributions in Diameter Classes
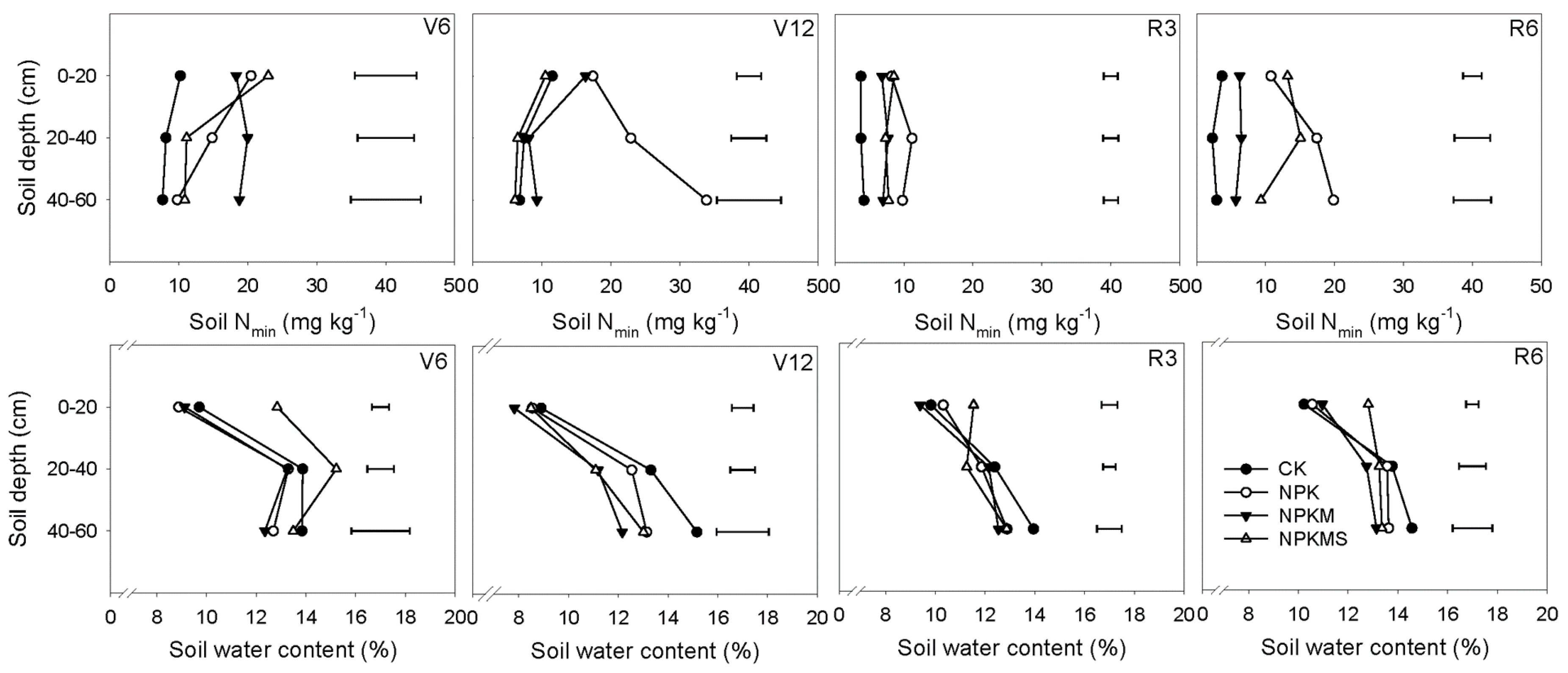
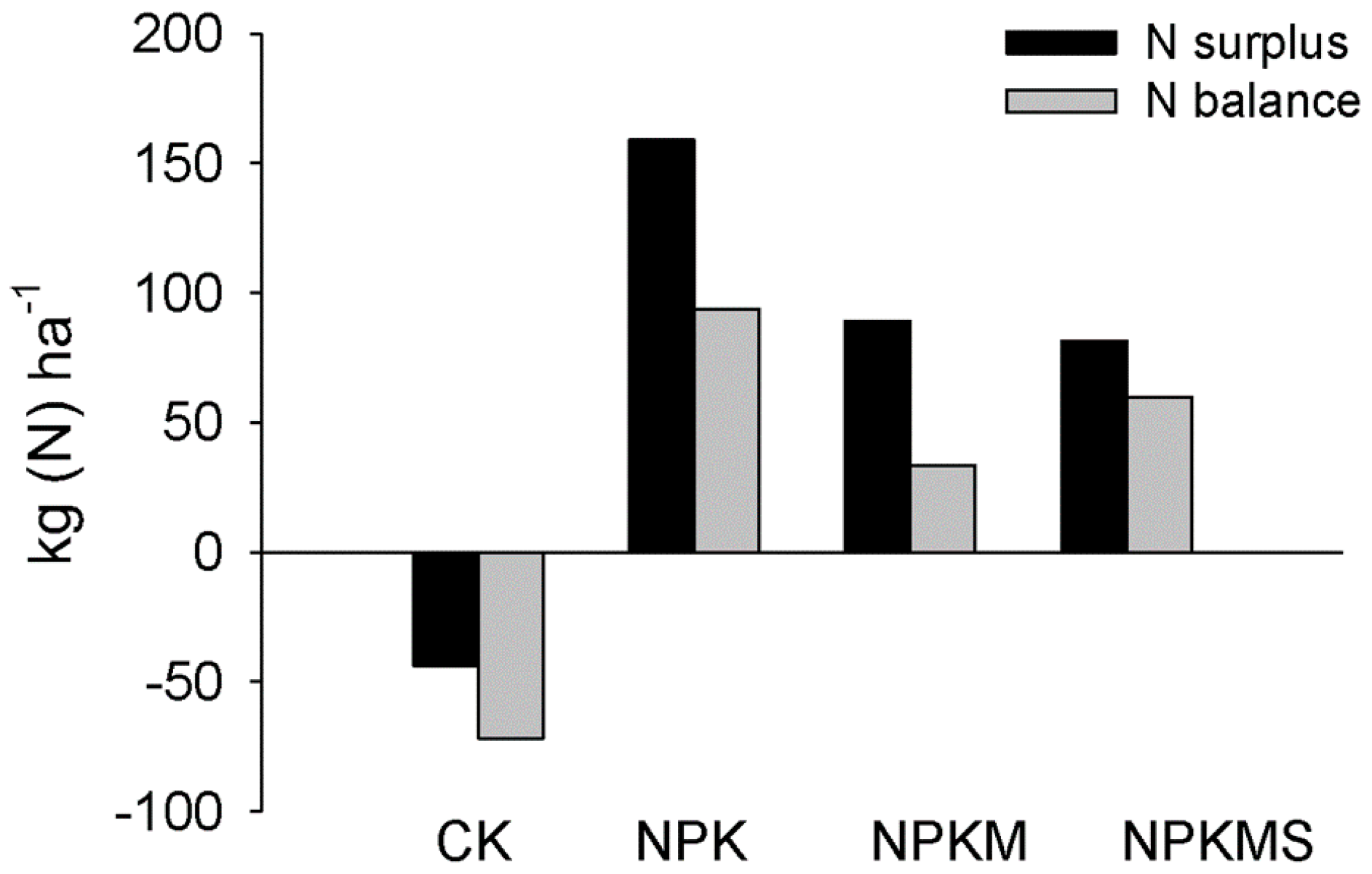
3.4. Soil Nmin, Water Content, and N Balances
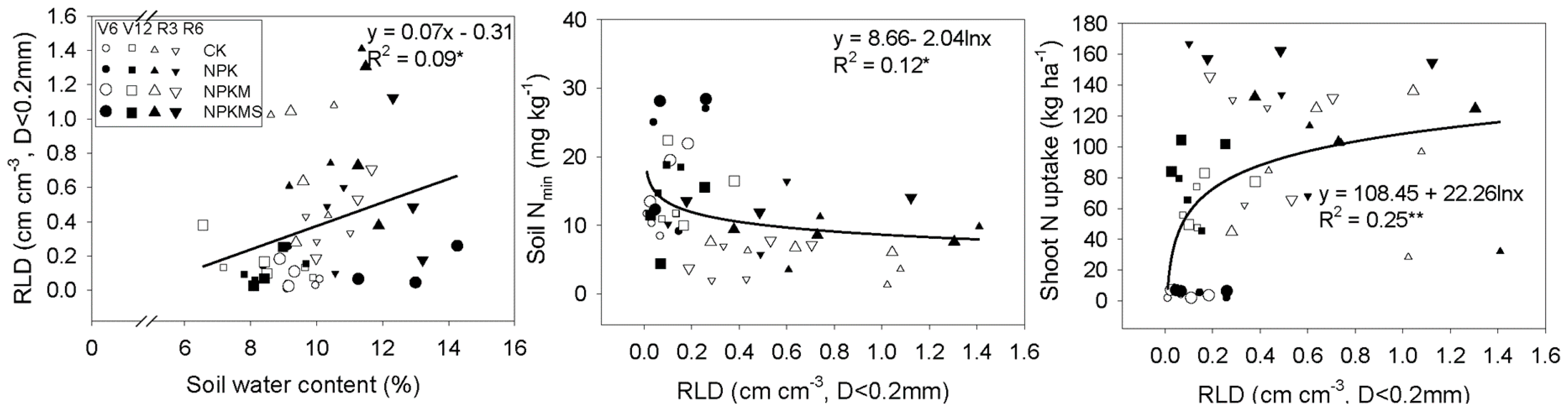
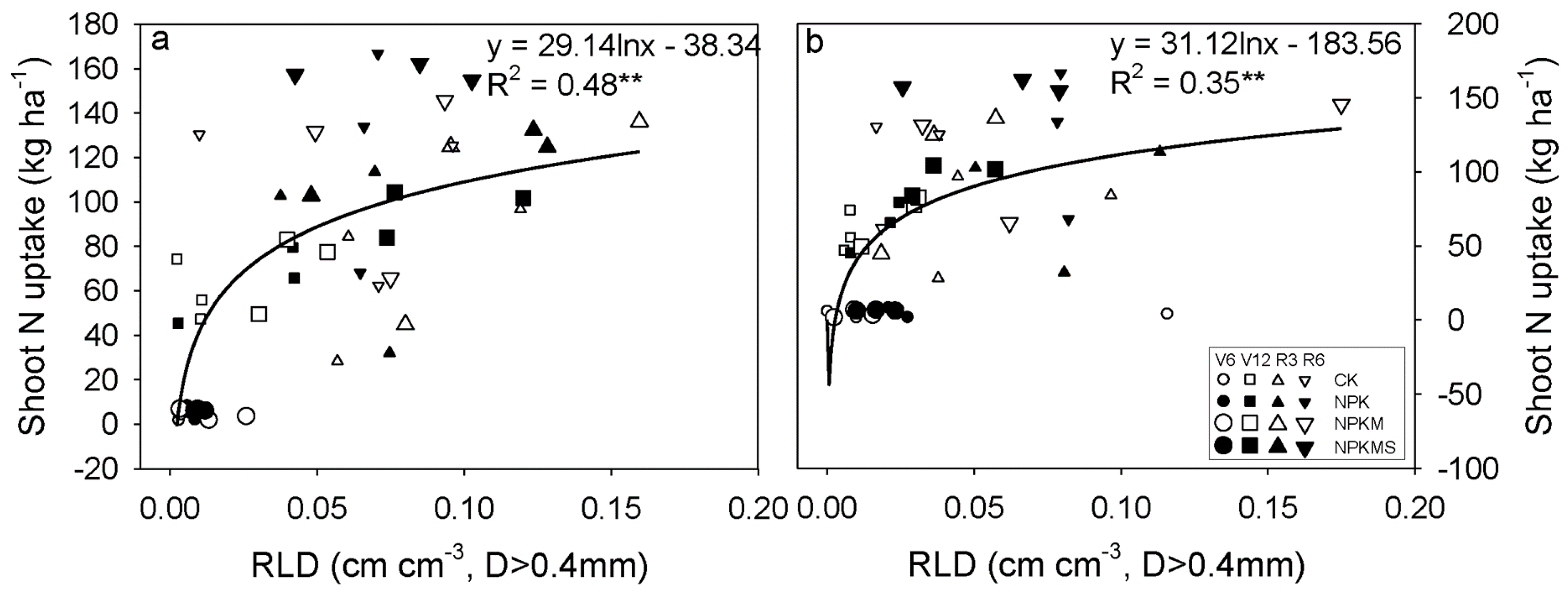
3.5. Correlations between Soil Water Content, RLD, Soil Nmin, and Shoot N Uptake
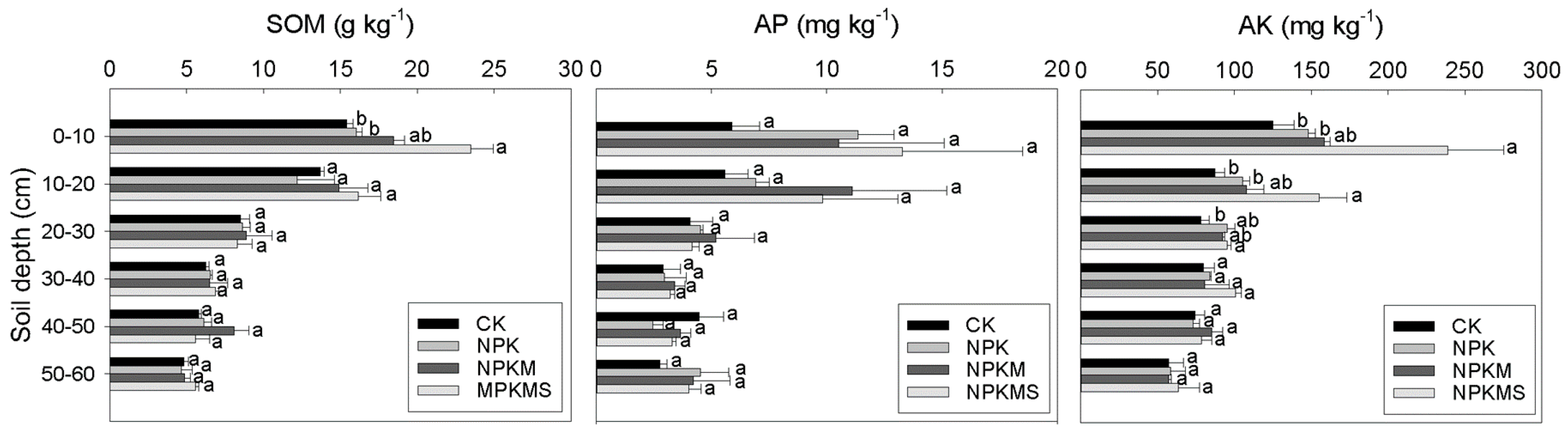
3.6. Soil Chemical Properties
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hawkesford, M.J. Reducing the reliance on nitrogen fertilizer for wheat production. J. Cereal Sci. 2014, 9, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.J.; Zhang, Y.; Han, W.X.; Tang, A.H.; Shen, J.L.; Cui, Z.L.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; de Vries, W.; Liu, X.J.; Hao, T.X.; Zeng, M.F.; Shen, J.B.; Zhang, F.S. Enhanced acidification in Chinese croplands as derived from element budgets in the period 1980–2010. Sci Total Environ. 2017, 618, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Mi, G.H.; Chen, F.J.; Wu, Q.P.; Lai, N.W.; Yuan, L.X.; Zhang, F.S. Ideotype root architecture for efficient nitrogen acquisition by maize in intensive cropping systems. Sci. China Life Sci. 2010, 53, 1369–1373. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, B.; Zhang, J.; Müller, C.; Cai, Z. Mechanisms of soil N dynamics following long-term application of organic fertilizers to subtropical rain-fed purple soil in China. Soil Biol. Biochem. 2015, 91, 222–231. [Google Scholar] [CrossRef]
- Caires, E.F.; Zardo Filho, R.; Barth, G.; Joris, H.A. Optimizing nitrogen use efficiency for no-till corn production by improving root growth and capturing NO3−N in subsoil. Pedosphere 2016, 26, 474–485. [Google Scholar] [CrossRef]
- Olmo, M.; Villar, R.; Salazar, P.; Alburquerque, J.A. Changes in soil nutrient availability explain biochar’s impact on wheat root development. Plant Soil 2016, 399, 333–343. [Google Scholar] [CrossRef]
- Kou, L.; Guo, D.; Yang, H.; Gao, W.; Li, S. Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China. Plant Soil 2015, 391, 207–218. [Google Scholar] [CrossRef]
- Tian, Q.Y.; Chen, F.J.; Zhang, F.S.; Mi, G.H. Genotypic Difference in nitrogen acquisition ability in maize plants is related to the coordination of leaf and root growth. J. Plant Nutr. 2006, 29, 317–330. [Google Scholar] [CrossRef]
- Gao, K.; Chen, F.; Yuan, L.; Zhang, F.S.; Mi, G.H. A comprehensive analysis of root morphological changes and nitrogen allocation in maize in response to low nitrogen stress. Plant Cell Environ. 2015, 38, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Postma, J.A.; Schurr, U.; Fiorani, F. Dynamic root growth and architecture responses to limiting nutrient availability: Linking physiological models and experimentation. Biotechnol. Adv. 2014, 32, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Li, C.; Mi, G.; Li, L.; Yuan, L.X.; Jiang, R.F.; Zhang, F.S. Maximizing root/rhizosphere efficiency to improve crop productivity and nutrient use efficiency in intensive agriculture of China. J. Exp. Bot. 2012, 64, 1181–1192. [Google Scholar] [CrossRef] [PubMed]
- Mi, G.H.; Liu, J.; Chen, F.J.; Zhang, F.S.; Cui, Z.L.; Liu, X.J. Nitrogen uptake and remobilization in maize hybrids differing leaf senescence. J. Plant Nutr. 2003, 26, 237–247. [Google Scholar] [CrossRef]
- Meng, L.; Ding, W.X.; Cai, Z.C. Long-term application of organic manure and nitrogen fertilizer on N2O emissions, soil quality and crop production in a sandy loam soil. Soil Biol. Biochem. 2005, 37, 2017–2045. [Google Scholar] [CrossRef]
- Soltani, S.; Khoshgoftarmanesh, A.H.; Afyuni, M.; Shrivani, M.; Schulin, R. The effect of preceding crop on wheat grain zinc concentration and its relationship to total amino acids and dissolved organic carbon in rhizosphere soil solution. Biol. Fertil. Soils 2014, 50, 239–247. [Google Scholar] [CrossRef]
- Hati, K.M.; Mandal, K.G.; Misra, A.K.; Ghosh, P.K.; Bandyopadhyay, K.K. Effect of inorganic fertilizer and farmyard manure on soil physical properties, root distribution, and water-use efficiency of soybean in Vertisols of central India. Bioresour. Technol. 2006, 97, 2182–2188. [Google Scholar] [CrossRef] [PubMed]
- Han, D.R.; Wiesmeier, M.; Conant, R.T.; Kühnel, N.; Sun, Z.G.; Kögel-Knabner, I.; Hou, R.X.; Cong, P.F.; Liang, R.B.; Zhu, O.Y. Large soil organic carbon increase due to improved agronomic management in the North China Plain from 1980s to 2010s. Glob. Chang. Biol. 2018, 24, 987–1000. [Google Scholar] [CrossRef] [PubMed]
- De Vries, J.W.; Hoogmoed, W.B.; Groenestein, C.M.; Schröder, J.J.; Sukkel, W.; De Boer, I.J.M.; Koerkamp, P.G. Integrated manure management to reduce environmental impact: I. Structured design of strategies. Agric. Syst. 2015, 139, 29–37. [Google Scholar] [CrossRef]
- Sáez, J.A.; Belda, R.M.; Bernal, M.P.; Fornes, F. Biochar improves agro-environmental aspects of pig slurry compost as a substrate for crops with energy and remediation uses. Ind. Crops Prod. 2016, 94, 97–106. [Google Scholar] [CrossRef]
- Yang, Q.; Tian, H.; Li, X.; Ren, W.; Zhang, B.; Zhang, X.; Wolf, J. Spatiotemporal patterns of livestock manure nutrient production in the conterminous United States from 1930 to 2012. Sci. Total Environ. 2016, 541, 1592–1602. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Huang, S.; Li, J.; Guo, D.; Lin, S.; Lu, G. Long-term manure amendments and chemical fertilizers enhanced soil organic carbon sequestration in a wheat (Triticum aestivum, L.)–maize (Zea mays, L.) rotation system. J. Sci. Food Agric. 2017, 97, 2575–2581. [Google Scholar] [CrossRef] [PubMed]
- Yazdanpanah, N.; Mahmoodabadi, M.; Cerdà, A. The impact of organic amendments on soil hydrology, structure and microbial respiration in semiarid lands. Geoderma 2016, 266, 58–65. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Li, C.H.; Wang, Y.W.; Hu, Y.M.; Christie, P.; Zhang, J.L.; Li, X.L. Maize yield and soil fertility with combined use of compost and inorganic fertilizers on a calcareous soil on the North China Plain. Soil Tillage Res. 2016, 155, 85–94. [Google Scholar] [CrossRef]
- Böhm, W. Methods of Studying Root Systems; Springer: New York, NY, USA, 1979. [Google Scholar]
- Niu, J.; Peng, Y.F.; Li, C.J.; Zhang, F.S. Changes in root length at the reproductive stage of maize plants own in the field and quartz sand. J. Plant Nutr. Soil Sci. 2010, 173, 306–314. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Chen, X.C.; Zhang, J.; Chen, Y.L.; Li, Q.; Chen, F.J.; Yuan, L.X.; Mi, G.H. Changes in root size and distribution in relation to nitrogen accumulation during maize breeding in China. Plant Soil. 2014, 374, 121–130. [Google Scholar] [CrossRef]
- Linkohr, B.I.; Williamson, L.C.; Fitter, A.H.; Leyser, H.M. Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis. Plant J. 2002, 29, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Inukai, Y.; Yamauchi, A. Root development and nutrient uptake. Crit. Rev. Plant Sci. 2006, 25, 279–301. [Google Scholar] [CrossRef]
- Lynch, J.P.; Brown, K.M. Topsoil foraging—An architectural adaptation of plants to low phosphorus availability. Plant Soil 2001, 237, 225–237. [Google Scholar] [CrossRef]
- Peng, Y.; Li, X.; Li, C. Temporal and spatial profiling of root growth revealed novel response of maize roots under various nitrogen supplies in the field. PLoS ONE 2012, 7, e37726. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Niu, J.; Peng, Z.; Zhang, F.S.; Li, C.J. Shoot growth potential drives N uptake in maize plants and correlates with root growth in the soil. Field Crop Res. 2010, 115, 85–93. [Google Scholar] [CrossRef]
- Fitter, A.; Williamson, L.; Linkohr, B.; Leyser, O. Root system architecture determines fitness in an Arabidopsis mutant in competition for immobile phosphate ions but not for nitrate ions. Proc. R. Soc. Lond. B Biol. Sci. 2002, 269, 2017–2022. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.H.; Shen, J.B.; Blackwell, M.; Li, H.G.; Zhao, B.Q.; Yuan, H.M. Combined applications of nitrogen and phosphorus fertilizers with manure increase maize yield and nutrient uptake via stimulating root growth in a long-term experiment. Pedosphere 2016, 26, 62–73. [Google Scholar] [CrossRef]
- Ning, P.; Liao, C.S.; Li, S.; Yu, P.; Zhang, Y.; Li, X.X.; Li, C.J. Maize cob plus husks mimics the grain sink to stimulate nutrient uptake by roots. Field Crop Res. 2012, 130, 38–45. [Google Scholar] [CrossRef]
- Wang, C.; Liu, W.; Li, Q.X.; Ma, D.Y.; Lu, H.F.; Wei, F.; Xie, Y.X.; Zhu, Y.J.; Guo, T.C. Effects of different irrigation and nitrogen regimes on root growth and its correlation with above-ground plant parts in high-yielding wheat under field conditions. Field Crop Res. 2014, 165, 138–149. [Google Scholar] [CrossRef]
- Marschner, P. Mineral Nutrition of Higher Plants, 3nd ed.; Academic Press: London, UK, 2011. [Google Scholar]
- Chen, Y.L.; Xiao, C.X.; Chen, X.C.; Li, Q.; Fang, J.; Chen, F.J.; Yuan, L.X.; Mi, G.H. Characterization of the plant traits contributed to high grain yield and high grain nitrogen concentration in maize. Field Crop Res. 2014, 159, 1–9. [Google Scholar] [CrossRef]
- Popescu, G.C.; Popescu, M. Yield, berry quality and physiological response of grapevine to foliar humic acid application. Bragantia. 2018, 77, 273–282. [Google Scholar] [CrossRef]
- Domingo-Olivé, F.; Bosch-Serra, À.D.; Yagüe, M.R.; Poch, R.M.; Boixadera, J. Long term application of dairy cattle manure and pig slurry to winter cereals improves soil quality. Nutr. Cycl. Agroecosyst. 2016, 104, 39–51. [Google Scholar] [CrossRef]
- Mi, W.H.; Wu, L.Q.; Brookes, P.C.; Liu, Y.L.; Zhang, X.; Yang, B. Changes in soil organic carbon fractions under integrated management systems in a low-productivity paddy soil given different organic amendments and chemical fertilizers. Soil Tillage Res. 2016, 163, 64–70. [Google Scholar] [CrossRef]
- Hartmann, T.E.; Yue, S.C.; Schulz, R.; Chen, X.P.; Zhang, F.S.; Müller, T. Nitrogen dynamics, apparent mineralization and balance calculations in a maize–wheat double cropping system of the North China Plain. Field Crop Res. 2014, 160, 22–30. [Google Scholar] [CrossRef]
- Benbi, D.K.; Kiranvir, B.; Sharma, S. Sensitivity of labile soil organic carbon pools to long-term fertilizer, straw and manure management in rice-wheat system. Pedosphere 2015, 25, 534–545. [Google Scholar] [CrossRef]
- Liu, E.K.; Yan, C.R.; Mei, X.R.; He, W.Q.; Bing, S.H.; Ding, L.P.; Liu, Q.; Liu, S.; Fan, T.L. Long-term effect of chemical fertilizer, straw, and manure on soil chemical and biological properties in northwest China. Geoderma 2010, 158, 173–180. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Treatments | Grain Yield (Mg ha−1) | Yield Components | ||
|---|---|---|---|---|---|
| Spikes ha−1 | Grains spike−1 | TKW (g per 1000 grains) | |||
| 2013 | CK | 5.72 ± 0.46b | 51,708 ± 846a | 433.8 ± 25.1a | 282.7 ± 5.7a |
| NPK | 6.44 ± 0.25ab | 50,433 ± 327a | 452.2 ± 14.7a | 284.5 ± 11.7a | |
| NPKM | 7.10 ± 0.61a | 51,567 ± 732a | 443.3 ± 6.3a | 275.9 ± 8.8a | |
| NPKMS | 6.89 ± 0.27a | 52,471 ± 366a | 449.2 ± 10.6a | 280.2 ± 8.0a | |
| 2014 | CK | 5.73 ± 0.16c | 67,292 ± 1296a | 325.3 ± 32.6b | 276.0 ± 6.5b |
| NPK | 8.54 ± 0.30b | 65,450 ± 818a | 449.4 ± 39.1ab | 314.2 ± 5.8a | |
| NPKM | 9.25 ± 0.57ab | 64,317 ± 1097a | 444.5 ± 54.6ab | 295.4 ± 13.7ab | |
| NPKMS | 9.80 ± 0.38a | 65,167 ± 1388a | 541.8 ± 36.7a | 288.9 ± 6.9ab | |
| 2015 | CK | 4.39 ± 0.27b | 50,000 ± 1416a | 207.8 ± 39.3b | 255.1 ± 2.5b |
| NPK | 7.67 ± 0.34a | 51,806 ± 731a | 381.3 ± 44.1a | 276.2 ± 12.8a | |
| NPKM | 7.63 ± 0.37a | 48,750 ± 765a | 398.9 ± 9.1a | 260.0 ± 11.2ab | |
| NPKMS | 8.35 ± 0.32a | 49,722 ± 1332a | 417.3 ± 47.3a | 261.6 ± 9.3ab | |
| Year | Treatments | V6 | V12 | R3 | R6 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Biomass Accumulation (Mg ha−1) | N Accumulation (Mg ha−1) | Biomass Accumulation (Mg ha−1) | N Accumulation (Mg ha−1) | Biomass Accumulation (Mg ha−1) | N Accumulation (Mg ha−1) | Biomass Accumulation (Mg ha−1) | N Accumulation (Mg ha−1) | ||
| 2013 | CK | 0.31 ± 0.05a | 6.19 ± 0.96a | 2.30 ± 0.43a | 31.95 ± 7.29b | 7.29 ± 1.21b | 58.97 ± 8.73b | 9.78 ± 0.83b | 78.13 ± 6.84b |
| NPK | 0.33 ± 0.03a | 7.37 ± 0.45a | 2.99 ± 0.34a | 41.86 ± 4.60ab | 9.76 ± 0.35ab | 111.15 ± 4.11a | 11.87 ± 1.07ab | 117.66 ± 14.76a | |
| NPKM | 0.37 ± 0.04a | 7.48 ± 0.55a | 2.90 ± 0.19a | 44.65 ± 4.68ab | 9.17 ± 0.55ab | 102.13 ± 11.36a | 12.73 ± 0.83a | 118.81 ± 4.84a | |
| NPKMS | 0.40 ± 0.05a | 8.12 ± 1.38a | 2.98 ± 0.21a | 50.93 ± 3.18a | 10.33 ± 1.19a | 118.28 ± 15.59a | 13.37 ± 0.78a | 127.69 ± 9.77a | |
| 2014 | CK | 0.15 ± 0.02b | 3.28 ± 0.50b | 2.40 ± 0.32b | 27.71 ± 6.24b | 5.86 ± 0.24b | 54.45 ± 3.57b | 11.43 ± 1.37b | 78.99 ± 10.44b |
| NPK | 0.27 ± 0.03a | 8.64 ± 0.98a | 3.61 ± 0.13a | 61.84 ± 2.49a | 8.14 ± 0.67a | 104.73 ± 7.16a | 17.53 ± 0.46a | 153.58 ± 13.28a | |
| NPKM | 0.26 ± 0.03a | 7.97 ± 0.59a | 3.21 ± 0.4ab | 59.16 ± 7.15a | 7.63 ± 0.55a | 98.84 ± 9.41a | 15.30 ± 0.65a | 157.07 ± 5.90a | |
| NPKMS | 0.29 ± 0.03a | 9.36 ± 0.64a | 3.74 ± 0.24a | 70.43 ± 5.81a | 8.33 ± 0.48a | 106.10 ± 8.24a | 17.89 ± 1.02a | 166.78 ± 14.25a | |
| 2015 | CK | 0.08 ± 0.00b | 1.92 ± 0.09b | 2.20 ± 0.51b | 38.50 ± 8.94b | 4.99 ± 0.24b | 34.29 ± 3.62b | 12.17 ± 0.71b | 71.75 ± 6.54b |
| NPK | 0.22 ± 0.02a | 6.81 ± 0.95a | 4.05 ± 0.32a | 82.24 ± 6.69a | 7.91 ± 0.65a | 110.16 ± 10.33a | 17.88 ± 0.99a | 156.36 ± 8.73a | |
| NPKM | 0.19 ± 0.02a | 5.69 ± 0.70a | 3.95 ± 0.57a | 78.57 ± 8.57a | 9.02 ± 0.56a | 116.37 ± 7.83a | 17.71 ± 0.82a | 141.67 ± 7.72a | |
| NPKMS | 0.24 ± 0.02a | 6.84 ± 0.49a | 4.21 ± 0.49a | 84.56 ± 7.91a | 8.55 ± 0.27a | 105.37 ± 7.27a | 18.29 ± 0.54a | 149.23 ± 6.01a | |
| Year | Treatments | V6 | V12 | R3 | R6 | ||||
|---|---|---|---|---|---|---|---|---|---|
| RLD (cm cm−3) | REN (mg m−1) | RLD (cm cm−3) | REN (mg m−1) | RLD (cm cm−3) | REN (mg m−1) | RLD (cm cm−3) | REN (mg m−1) | ||
| 2013 | CK | 0.16 ± 0.04a | 0.72 ± 0.11a | 0.26 ± 0.08a | 2.11 ± 0.11a | 0.56 ± 0.13a | 2.37 ± 0.95b | 0.45 ± 0.06a | 3.02 ± 0.67a |
| NPK | 0.10 ± 0.03a | 1.37 ± 0.33a | 0.24 ± 0.06a | 3.20 ± 0.32a | 0.68 ± 0.15a | 2.99 ± 0.69a | 0.22 ± 0.07a | 12.59 ± 5.37a | |
| NPKM | 0.11 ± 0.01a | 1.18 ± 0.19a | 0.36 ± 0.02a | 1.96 ± 0.32a | 0.61 ± 0.08a | 2.78 ± 0.49a | 0.41 ± 0.09a | 5.07 ± 1.03a | |
| NPKMS | 0.10 ± 0.01a | 1.59 ± 0.42a | 0.32 ± 0.10a | 3.25 ± 0.75a | 0.91 ± 0.13a | 2.29 ± 0.25a | 0.30 ± 0.11a | 9.07 ± 2.71a | |
| 2014 | CK | 0.09 ± 0.02a | 0.83 ± 0.31a | 0.26 ± 0.05ab | 1.46 ± 0.24c | 0.44 ± 0.14a | 2.65 ± 1.01a | 0.27 ± 0.03a | 4.45 ± 0.78a |
| NPK | 0.09 ± 0.02a | 1.93 ± 0.73a | 0.34 ± 0.03a | 3.13 ± 0.16b | 0.44 ± 0.09a | 4.04 ± 0.54a | 0.48 ± 0.08a | 5.47 ± 0.48a | |
| NPKM | 0.12 ± 0.03a | 1.34 ± 0.44a | 0.16 ± 0.03b | 5.92 ± 0.72a | 0.58 ± 0.09a | 2.75 ± 0.56a | 0.45 ± 0.09a | 6.29 ± 1.63a | |
| NPKMS | 0.17 ± 0.04a | 1.03 ± 0.26a | 0.28 ± 0.03ab | 4.11 ± 0.39b | 0.47 ± 0.06a | 3.49 ± 0.55a | 0.49 ± 0.16a | 7.78 ± 3.86a | |
| 2015 | CK | 0.19 ± 0.16a | 0.67 ± 0.30a | 0.10 ± 0.01a | 7.84 ± 0.98a | 0.74 ± 0.13a | 0.84 ± 0.15b | 0.41 ± 0.07a | 2.89 ± 0.66a |
| NPK | 0.26 ± 0.04a | 0.43 ± 0.12a | 0.16 ± 0.01a | 8.89 ± 0.53a | 0.76 ± 0.08a | 2.33 ± 0.38ab | 0.47 ± 0.10a | 6.37 ± 2.06a | |
| NPKM | 0.24 ± 0.08a | 0.44 ± 0.09a | 0.28 ± 0.06a | 4.96 ± 1.37a | 0.71 ± 0.13a | 2.98 ± 0.65a | 0.88 ± 0.14a | 2.74 ± 0.41a | |
| NPKMS | 0.31 ± 0.03a | 0.36 ± 0.02a | 0.35 ± 0.13a | 5.86 ± 2.59a | 0.82 ± 0.19a | 2.67 ± 0.65a | 0.61 ± 0.24a | 6.09 ± 3.03a | |
| Growth Period | Treatments | 0–10 cm | 10–20 cm | 20–30 cm | 30–40 cm | 40–50 cm | 50–60 cm |
|---|---|---|---|---|---|---|---|
| V6 | CK | 0.05 ± 0.03a | 0.02 ± 0.00a | 0.05 ± 0.03a | 0.03 ± 0.01a | 0.01 ± 0.00a | 0.87 ± 0.84a |
| NPK | 0.26 ± 0.23a | 0.12 ± 0.09a | 0.17 ± 0.13a | 0.24 ± 0.22a | 0.23 ± 0.13a | 0.25 ± 0.09a | |
| NPKM | 0.13 ± 0.07a | 0.13 ± 0.09a | 0.30 ± 0.28a | 0.13 ± 0.06a | 0.30 ± 0.17a | 0.14 ± 0.05a | |
| NPKMS | 0.10 ± 0.04a | 0.15 ± 0.10a | 0.08 ± 0.06a | 0.28 ± 0.21a | 0.50 ± 0.23a | 0.24 ± 0.12a | |
| V12 | CK | 0.14 ± 0.03a | 0.08 ± 0.02a | 0.03 ± 0.02b | 0.01 ± 0.00a | 0.02 ± 0.00a | 0.04 ± 0.00b |
| NPK | 0.07 ± 0.05a | 0.14 ± 0.01a | 0.06 ± 0.02b | 0.10 ± 0.07a | 0.05 ± 0.01a | 0.07 ± 0.03ab | |
| NPKM | 0.10 ± 0.04a | 0.33 ± 0.15a | 0.13 ± 0.04ab | 0.06 ± 0.01a | 0.14 ± 0.08a | 0.18 ± 0.05a | |
| NPKMS | 0.03 ± 0.02a | 0.20 ± 0.12a | 0.56 ± 0.27a | 0.16 ± 0.05a | 0.08 ± 0.04a | 0.12 ± 0.02ab | |
| R3 | CK | 0.80 ± 0.03a | 0.89 ± 0.40a | 0.17 ± 0.04a | 0.13 ± 0.03a | 0.13 ± 0.02a | 0.06 ± 0.02b |
| NPK | 1.05 ± 0.08a | 0.79 ± 0.57a | 0.09 ± 0.01a | 0.07 ± 0.03a | 0.19 ± 0.04a | 0.18 ± 0.07ab | |
| NPKM | 1.11 ± 0.47a | 0.20 ± 0.04a | 0.24 ± 0.01a | 0.19 ± 0.07a | 0.11 ± 0.05a | 0.06 ± 0.02b | |
| NPKMS | 0.98 ± 0.35a | 0.62 ± 0.29a | 0.09 ± 0.03a | 0.32 ± 0.14a | 0.10 ± 0.05a | 0.27 ± 0.05a | |
| R6 | CK | 0.37 ± 0.11a | 0.33 ± 0.06a | 0.11 ± 0.05a | 0.30 ± 0.20a | 0.04 ± 0.01b | 0.06 ± 0.03b |
| NPK | 0.05 ± 0.04a | 0.76 ± 0.29a | 0.09 ± 0.01a | 0.14 ± 0.04a | 0.11 ± 0.01a | 0.18 ± 0.04ab | |
| NPKM | 0.37 ± 0.25a | 0.58 ± 0.35a | 0.15 ± 0.05a | 0.11 ± 0.02a | 0.14 ± 0.06a | 0.32 ± 0.12a | |
| NPKMS | 0.66 ± 0.55a | 0.53 ± 0.23a | 0.09 ± 0.05a | 0.22 ± 0.10a | 0.14 ± 0.08a | 0.08 ± 0.03b |
| Growth Period | Treatments | 0–10 cm | 10–20 cm | 20–30 cm | 30–40 cm | 40–50 cm | 50–60 cm |
|---|---|---|---|---|---|---|---|
| V6 | CK | 0.01 ± 0.01a | 0.01 ± 0.00a | 0.12 ± 0.06a | 0.01 ± 0.00a | 0.01 ± 0.00b | 0.35 ± 0.34a |
| NPK | 0.02 ± 0.01a | 0.04 ± 0.03a | 0.31 ± 0.15a | 0.04 ± 0.03a | 0.07 ± 0.03ab | 0.09 ± 0.03a | |
| NPKM | 0.05 ± 0.03a | 0.03 ± 0.01a | 0.58 ± 0.05a | 0.06 ± 0.03a | 0.06 ± 0.05ab | 0.07 ± 0.02a | |
| NPKMS | 0.03 ± 0.01a | 0.04 ± 0.02a | 0.36 ± 0.20a | 0.07 ± 0.03a | 0.12 ± 0.04a | 0.08 ± 0.04a | |
| V12 | CK | 0.06 ± 0.00a | 0.05 ± 0.03a | 0.02 ± 0.01b | 0.01 ± 0.00b | 0.01 ± 0.00a | 0.03 ± 0.01b |
| NPK | 0.02 ± 0.01b | 0.08 ± 0.00a | 0.03 ± 0.01b | 0.05 ± 0.03ab | 0.03 ± 0.01a | 0.03 ± 0.01b | |
| NPKM | 0.02 ± 0.01b | 0.15 ± 0.06a | 0.05 ± 0.01b | 0.03 ± 0.01ab | 0.05 ± 0.02a | 0.08 ± 0.02a | |
| NPKMS | 0.02 ± 0.01b | 0.11 ± 0.07a | 0.26 ± 0.13a | 0.07 ± 0.02a | 0.05 ± 0.02a | 0.06 ± 0.01ab | |
| R3 | CK | 0.64 ± 0.05a | 0.54 ± 0.25a | 0.10 ± 0.02a | 0.06 ± 0.02a | 0.10 ± 0.02a | 0.04 ± 0.01bc |
| NPK | 0.70 ± 0.09a | 0.34 ± 0.23a | 0.04 ± 0.01a | 0.03 ± 0.01a | 0.09 ± 0.02a | 0.13 ± 0.04ab | |
| NPKM | 0.66 ± 0.28a | 0.13 ± 0.03a | 0.11 ± 0.05a | 0.09 ± 0.03a | 0.06 ± 0.03a | 0.03 ± 0.01c | |
| NPKMS | 0.69 ± 0.26a | 0.37 ± 0.19a | 0.05 ± 0.00a | 0.15 ± 0.06a | 0.06 ± 0.04a | 0.17 ± 0.04a | |
| R6 | CK | 0.23 ± 0.03a | 0.19 ± 0.04a | 0.08 ± 0.04a | 0.17 ± 0.12a | 0.03 ± 0.01b | 0.03 ± 0.01b |
| NPK | 0.02 ± 0.02a | 0.53 ± 0.19a | 0.03 ± 0.01a | 0.07 ± 0.01a | 0.05 ± 0.01b | 0.09 ± 0.02ab | |
| NPKM | 0.19 ± 0.15a | 0.36 ± 0.24a | 0.06 ± 0.03a | 0.04 ± 0.01a | 0.56 ± 0.26a | 0.16 ± 0.07a | |
| NPKMS | 0.42 ± 0.37a | 0.33 ± 0.14a | 0.04 ± 0.03a | 0.11 ± 0.06a | 0.07 ± 0.05b | 0.04 ± 0.02ab |
| Growth Period | Treatments | 0–10 cm | 10–20 cm | 20–30 cm | 30–40 cm | 40–50 cm | 50–60 cm |
|---|---|---|---|---|---|---|---|
| V6 | CK | 0.02 ± 0.01a | 0.01 ± 0.00a | 0.00 ± 0.00a | 0.00 ± 0.00a | 0.01 ± 0.01a | 0.12 ± 0.11a |
| NPK | 0.01 ± 0.00a | 0.03 ± 0.02a | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.02 ± 0.01a | 0.03 ± 0.01a | |
| NPKM | 0.03 ± 0.00a | 0.02 ± 0.00a | 0.01 ± 0.01a | 0.02 ± 0.02a | 0.01 ± 0.01a | 0.01 ± 0.00a | |
| NPKMS | 0.02 ± 0.01a | 0.04 ± 0.02a | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.02 ± 0.01a | 0.01 ± 0.00a | |
| V12 | CK | 0.08 ± 0.03a | 0.04 ± 0.02b | 0.01 ± 0.00b | 0.00 ± 0.00a | 0.01 ± 0.00b | 0.01 ± 0.00b |
| NPK | 0.05 ± 0.03a | 0.09 ± 0.02ab | 0.04 ± 0.02b | 0.02 ± 0.01a | 0.02 ± 0.01ab | 0.02 ± 0.01b | |
| NPKM | 0.05 ± 0.02a | 0.15 ± 0.04a | 0.06 ± 0.01b | 0.03 ± 0.00a | 0.03 ± 0.01ab | 0.03 ± 0.00ab | |
| NPKMS | 0.02 ± 0.01a | 0.07 ± 0.03ab | 0.13 ± 0.02a | 0.05 ± 0.01a | 0.04 ± 0.01a | 0.04 ± 0.01a | |
| R3 | CK | 0.27 ± 0.08a | 0.18 ± 0.07a | 0.10 ± 0.03a | 0.06 ± 0.01a | 0.08 ± 0.02a | 0.04 ± 0.02b |
| NPK | 0.52 ± 0.14a | 0.18 ± 0.05a | 0.07 ± 0.01a | 0.05 ± 0.01a | 0.08 ± 0.01a | 0.09 ± 0.03b | |
| NPKM | 0.39 ± 0.11a | 0.12 ± 0.03a | 0.13 ± 0.03a | 0.09 ± 0.02a | 0.04 ± 0.01a | 0.04 ± 0.02b | |
| NPKMS | 0.37 ± 0.08a | 0.20 ± 0.07a | 0.06 ± 0.02a | 0.14 ± 0.03a | 0.06 ± 0.03a | 0.17 ± 0.02a | |
| R6 | CK | 0.13 ± 0.08a | 0.20 ± 0.03a | 0.06 ± 0.03a | 0.05 ± 0.02a | 0.02 ± 0.01b | 0.03 ± 0.01a |
| NPK | 0.04 ± 0.03a | 0.37 ± 0.09a | 0.08 ± 0.00a | 0.06 ± 0.00a | 0.07 ± 0.01ab | 0.09 ± 0.00a | |
| NPKM | 0.21 ± 0.16a | 0.29 ± 0.15a | 0.08 ± 0.01a | 0.07 ± 0.01a | 0.17 ± 0.06a | 0.10 ± 0.04a | |
| NPKMS | 0.29 ± 0.19a | 0.37 ± 0.15a | 0.06 ± 0.03a | 0.09 ± 0.02a | 0.05 ± 0.03ab | 0.06 ± 0.03a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Li, T.; Bei, S.; Zhang, J.; Li, X. Growth and Distribution of Maize Roots in Response to Nitrogen Accumulation in Soil Profiles after Long-Term Fertilization Management on a Calcareous Soil. Sustainability 2018, 10, 4315. https://doi.org/10.3390/su10114315
Zhang Y, Li T, Bei S, Zhang J, Li X. Growth and Distribution of Maize Roots in Response to Nitrogen Accumulation in Soil Profiles after Long-Term Fertilization Management on a Calcareous Soil. Sustainability. 2018; 10(11):4315. https://doi.org/10.3390/su10114315
Chicago/Turabian StyleZhang, Yunlong, Tengteng Li, Shuikuan Bei, Junling Zhang, and Xiaolin Li. 2018. "Growth and Distribution of Maize Roots in Response to Nitrogen Accumulation in Soil Profiles after Long-Term Fertilization Management on a Calcareous Soil" Sustainability 10, no. 11: 4315. https://doi.org/10.3390/su10114315
APA StyleZhang, Y., Li, T., Bei, S., Zhang, J., & Li, X. (2018). Growth and Distribution of Maize Roots in Response to Nitrogen Accumulation in Soil Profiles after Long-Term Fertilization Management on a Calcareous Soil. Sustainability, 10(11), 4315. https://doi.org/10.3390/su10114315