
Structural Characterization and Hypoglycemic Function of Polysaccharides from Cordyceps cicadae

Abstract
1. Introduction
2. Results and Discussion
2.1. Purified Polysaccharides
2.2. Chemical Compositions and Molecular Weights of SPP, BSP, and PPP
2.3. FT-IR and UV-Vis Spectra
2.4. Methylation Status
2.5. Nuclear Magnetic Resonance (NMR) Spectra
2.6. In Vitro Hypoglycemic Activity
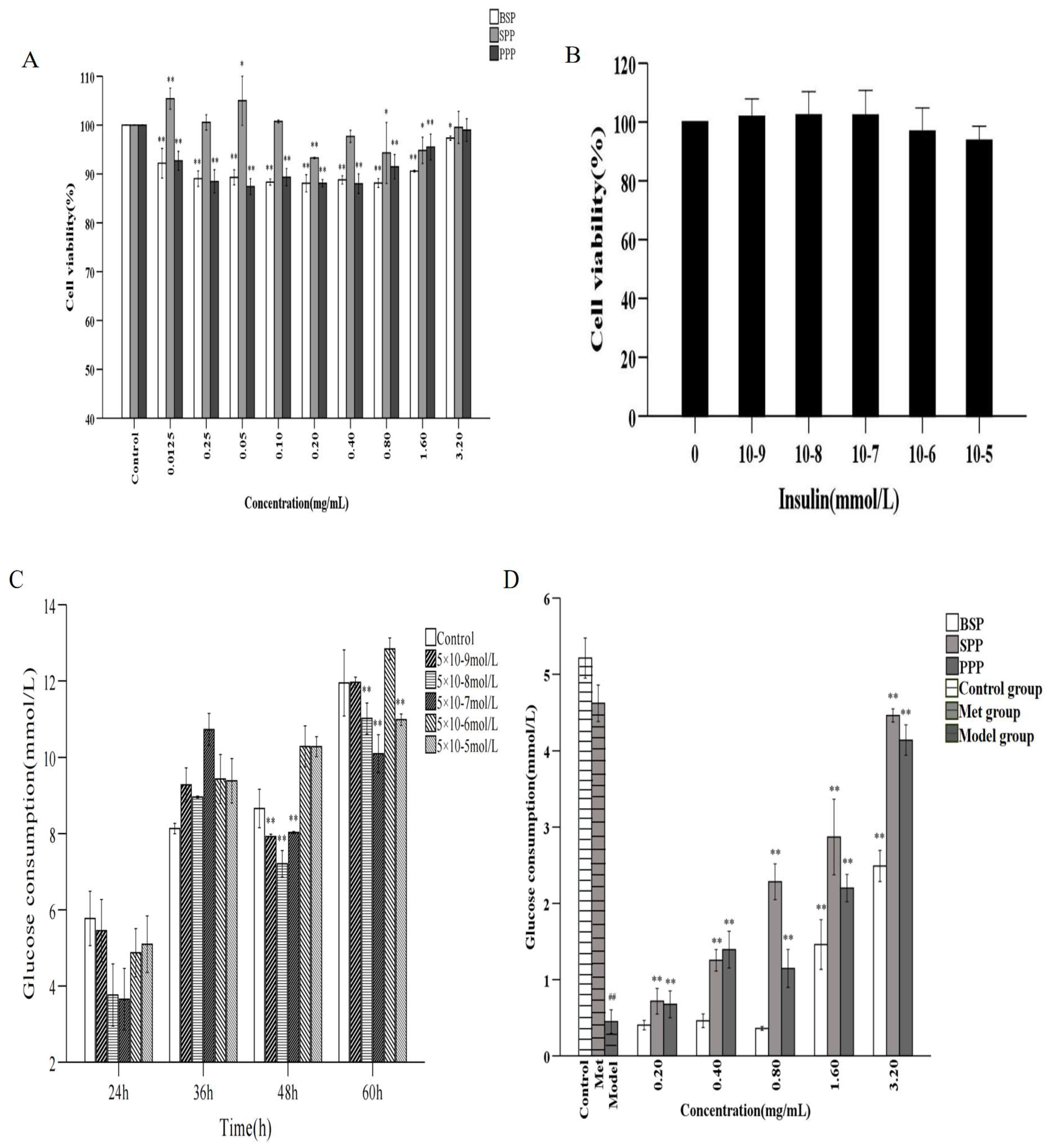
2.6.1. Cytotoxicity of SPP, BSP, and PPP
2.6.2. IR-HepG2 Cell Model
2.6.3. Effect of Cordyceps Cicadae Polysaccharides on Glucose Consumption in the IR-HepG2 Cell Model
2.7. In Vivo Hypoglycemic Activity
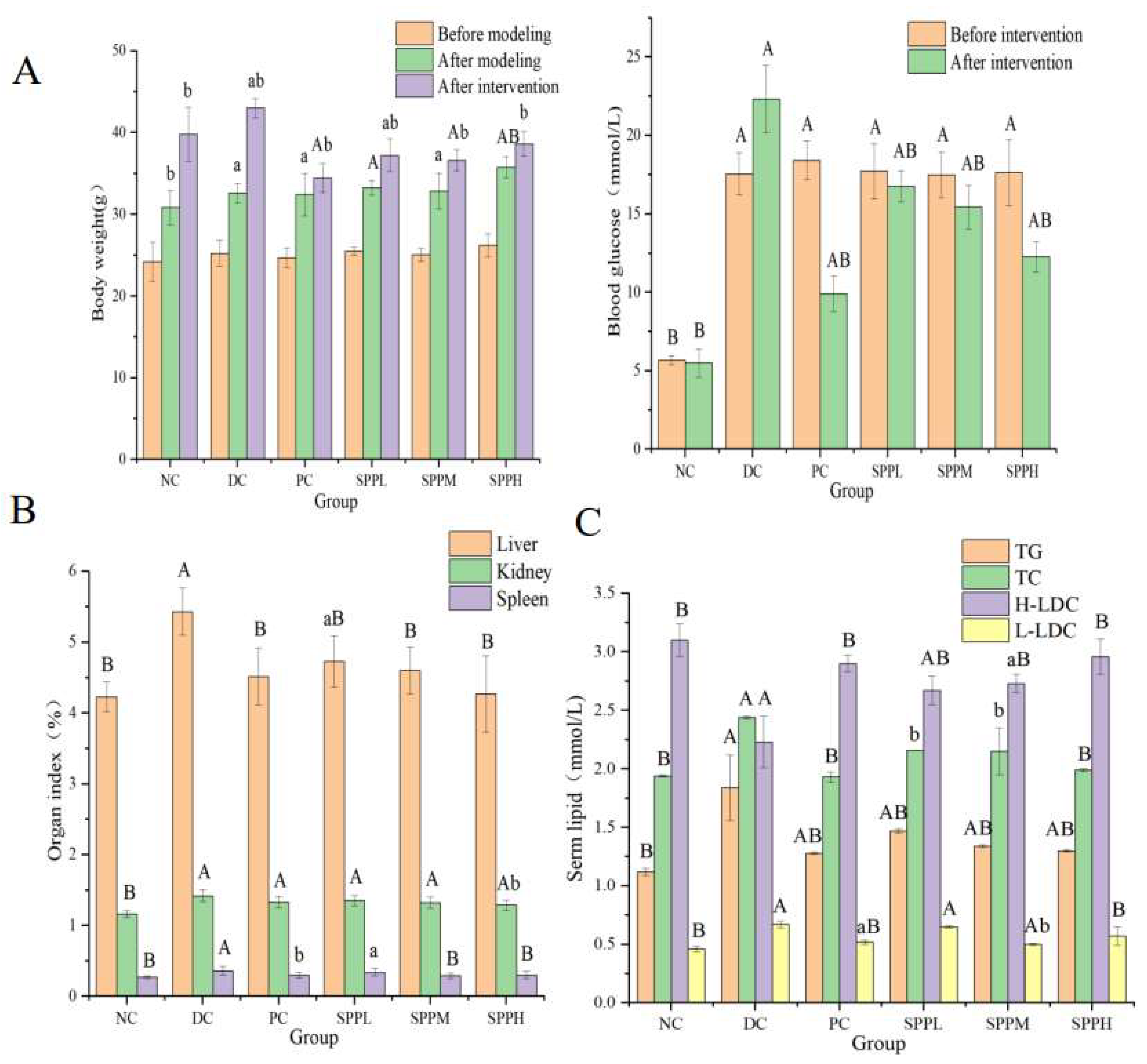
2.7.1. Effects of SPP on Body Weight and Blood Glucose in Mice
2.7.2. Effect of SPP on the Organ Index in T2DM Mice
2.7.3. Effect of SPP on Blood Lipid Levels in T2DM Mice
2.7.4. Effects of SPP on the Morphology of Pancreatic and Liver Tissue
2.7.5. Effect of SPP on the Liver Expression Levels of IRS-1, IRS-2, PI3K, Akt, and GLUT4 in T2DM Mice
2.7.6. Effect of SPP on the Liver Expression of IRS-1, PI3K, Akt, and GLUT4 Proteins in T2DM Mice
3. Materials and Methods
3.1. Materials and Reagents
3.2. Extraction and Purification of Polysaccharides
3.3. Chemical Composition Analysis
3.4. Monosaccharide Composition Analysis
3.5. Molecular Weight Analysis of SPP, BSP, and PPP
3.6. FT-IR and UV-Vis Spectroscopy
3.7. Methylation Analysis
3.8. Nuclear Magnetic Resonance (NMR) Analysis
3.9. In Vitro Hypoglycemic Activity Analysis
3.9.1. Cytotoxicity Assays of SPP, BSP, and PPP
3.9.2. Establishment of the IR-HepG2 Cell Model
Effect of Insulin Concentration on the Survival Rate of HepG2 Cells
Effect of Insulin Concentration on Glucose Consumption by HepG2 Cells
Effects of SPP, BSP, and PPP on Glucose Consumption by IR-HepG2 Cells
3.10. In Vivo Hypoglycemic Activity Analysis
3.10.1. Animals and Experimental Design
3.10.2. Fasting Blood Glucose (FBG) and Serum Lipid Level Analysis
3.10.3. Histopathological Analysis and Organ Index Calculation
3.10.4. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
3.10.5. Determination of Protein Expression Levels
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Tang, Y.; Li, Y.; Zhang, M.; Chen, Y.; Sun, Q.S. 3/4 genotype of the apolipoprotein E is associated with a higher risk of Alzheimer’s disease in patients with type 2 diabetes mellitus. Gene 2019, 703, 65–70. [Google Scholar] [CrossRef]
- Wang, X.; Qin, A.; Xiao, F.; Olatunji, O.J.; Zhang, S.; Pan, D.; Han, W.; Wang, D.; Ni, Y. N6-(2-hydroxyethyl)-adenosine from Cordyceps cicadae protects against diabetic kidney disease via alleviation of oxidative stress and inflammation. J. Food Biochem. 2019, 43, 12727. [Google Scholar] [CrossRef]
- Herman, W.H.; Zimmet, P. Type 2 diabetes: An epidemic requiring global attention and urgent action. Diabetes Care 2012, 35, 943–944. [Google Scholar] [CrossRef]
- Mazzone, T.; Chait, A.; Plutzky, J. Cardiovascular disease risk in type 2 diabetes mellitus: Insights from mechanistic studies. Lancet 2008, 371, 1800–1809. [Google Scholar] [CrossRef]
- Thorens, B. Of Fat, β Cells, and Diabetes. Cell Metab. 2011, 14, 439–440. [Google Scholar] [CrossRef]
- Ruoshi, L.; Aien, T.; Runmei, Y.; Min, F.; Zhang, X.C.; Du, Z.F.; Shang, F.; Xia, C.; Duan, B. Structural characterization, hypoglycemic effects and antidiabetic mechanism of a novel polysaccharides from Polygonatum kingianum Coll. et Hemsl. Biomed. Pharmacother. 2020, 131, 110687. [Google Scholar] [CrossRef]
- Carbone, S.; Dixon, D.L.; Buckley, L.F.; Abbate, A. Glucose-lowering therapies for cardiovascular risk reduction in type 2 diabetes mellitus: State-of-the-Art Review. Mayo Clin. Proc. 2018, 93, 1629–1647. [Google Scholar] [CrossRef]
- Hsieh, Y.T.; Tsai, M.J.; Tu, S.T.; Hsieh, M.C. Association of abnormal renal profiles and proliferative diabetic retinopathy and diabetic macular edema in an Asian population with type 2 diabetes. JAMA Ophthalmol. 2018, 136, 68–74. [Google Scholar] [CrossRef]
- Strack, T. Metformin: A review. Drugs Today 2008, 44, 303–314. [Google Scholar] [CrossRef]
- Richard, K.R.; Shelburne, J.S.; Kirk, J.K. Tolerability of Dipeptidyl Peptidase-4Inhibitors: A Review. Clin. Ther. 2011, 33, 1609–1629. [Google Scholar] [CrossRef]
- Kumar, R.; Kerinlis, D.M.; Walther, T. Cardiovascular safety of anti-diabetic drugs. Eur. Heart J. Cardiovasc. Pharmacother. 2016, 1, 32–43. [Google Scholar] [CrossRef]
- Khan, S.; Khan, H.U.; Khan, F.A.; Shah, A.; Wadood, A.; Ahmad, S.; Almehmadi, M.; Alsaiari, A.A.; Shah, F.U.; Kamran, N. Anti-Alzheimer and Antioxidant Effects of Nelumbo nucifera L. Alkaloids, Nuciferine and Norcoclaurine in Alloxan-Induced Diabetic Albino Rats. Pharmaceuticals 2022, 15, 1205. [Google Scholar] [CrossRef]
- Alam, A.; Ali, M.; Rehman, N.U.; Ullah, S.; Halim, S.A.; Latif, A.; Zainab; Khan, A.; Ullah, O.; Ahmad, S.; et al. Bio-Oriented Synthesis of Novel (S)-Flurbiprofen Clubbed Hydrazone Schiff’s Bases for Diabetic Management: In Vitro and In Silico Studies. Pharmaceuticals 2022, 15, 672. [Google Scholar] [CrossRef]
- Artunc, F.; Schleicher, E.; Weigert, C.; Fritsche, A.; Stefan, N.; Haring, H. The impact of insulin resistance on the kidney and vasculature. Nat. Rev. Nephrol. 2016, 12, 721–737. [Google Scholar] [CrossRef]
- Onyango, A. Cellular stresses and stress responses in the pathogenesis of insulin resistance. Oxidative Med. Cell. Longev. 2018, 2018, 4321714. [Google Scholar] [CrossRef]
- Ganesan, X.; Xu, B.J. Anti-Diabetic Effects and Mechanisms of Dietary Polysaccharides. Molecules 2019, 24, 2556. [Google Scholar] [CrossRef]
- Petersen, M.C.; Shulman, G.I. Mechanisms of Insulin Action and Insulin Resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef]
- Fukushima, T.; Arai, T.; Ariganedachi, M.; Okajima, H.; Ooi, Y.; Iijima, Y.; Sone, M.; Cho, Y.; Ando, Y.; Kasahara, K.; et al. Insulin receptor substrates form high-molecular-mass complexes that modulate their availability to insulin/insulin-like growth factor-I receptor tyrosine kinases. Biochem. Biophys. Res. Commun. 2011, 404, 767–773. [Google Scholar] [CrossRef]
- Zhu, Y.; Pereira, R.O.; O’Neill, B.T.; Riehle, C.; Ilkun, O.; Wende, A.R.; Rawlings, T.A.; Zhang, Y.C.; Zhang, Q.; Klip, A.; et al. Cardiac PI3K-Akt impairs insulin-stimulated glucose uptake independent of mTORC1 and GLUT4 translocation. Mol. Endocrinol. 2013, 27, 172–184. [Google Scholar] [CrossRef]
- Arden, K.C.; Biggs, W. Regulation of the FoxO family of transcription factors by phosphatidylinositol-3 kinase-activated signaling. Arch. Biochem. Biophys. 2012, 403, 292–298. [Google Scholar] [CrossRef]
- Zhang, Y.; Wen, X.; Huang, X.; Zhao, Y.; Ren, Q.; Hong, Z.; Huang, M.; Xing, X. Fucoxanthin ameliorates hyperglycemia, hyperlipidemia and insulin resistance in diabetic mice partially through IRS-1/PI3K/Akt and AMPK pathways. J. Funct. Foods 2018, 48, 515–524. [Google Scholar] [CrossRef]
- Santoleri, D.; Titchenell, P. Resolving the paradox of hepatic insulin resistance. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 447–456. [Google Scholar] [CrossRef]
- Titchenell, P.; Lazar, M.; Birnbaum, M. Unraveling the regulation of hepatic metabolism by insulin. Trends Endocrinol. Metab. 2017, 28, 497–505. [Google Scholar] [CrossRef] [PubMed]
- King, G.; Park, K.; Li, Q. Selective insulin resistance and the development of cardiovascular diseases in diabetes: The 2015 Edwin Bierman award lecture. Diabetes 2016, 65, 1462–1471. [Google Scholar] [CrossRef] [PubMed]
- Favaretto, F.; Milan, G.; Collin, G.B.; Marshall, J.D.; Stasi, F.; Maffei, P.; Vettor, R.; Naggert, J.K. GLUT4 defects in adipose tissue are early signs of metabolic alterations in Alms1GT/GT, a mouse model for obesity and insulin resistance. PLoS ONE 2014, 9, e109540. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Wong, T.C.; Huang, C.L.; Jhou, B.Y.; Hsu, J.H.; Yeh, S.H.; Tsai, Y.T.; Yang, S.H. Clinical evaluation of blood glucose regulation and safety of Cordyceps cicadae mycelium. J. Food Nutr. Res. 2017, 5, 137–143. [Google Scholar]
- Ng, T.B.; Wang, H.X. Pharmacological actions of Cordyceps, a prized folk medicine. J. Pharm. Pharmacol. 2005, 57, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.J.; Yu, T.; Yang, L.; Liu, T.; Song, Z.Y.; Liu, S.H.; Zhang, D.Y.; Tang, C.Z. Polysaccharide from Cordyceps cicadae inhibit mitochondrial apoptosis to ameliorate drug-induced kidney injury via Bax/Bcl-2/Caspase-3 pathway. J. Funct. Foods 2022, 97, 105244. [Google Scholar] [CrossRef]
- Hsu, J.H.; Jhou, B.Y.; Yeh, S.H.; Chen, Y.L.; Chen, C.C. Healthcare Functions of Cordyceps cicadae. J. Nutr. Food Sci. 2015, 5, 432. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, D.; Wang, D.; Lai, S.; Zhong, R.; Liu, Y.; Yang, C.; Liu, B.; Sarker, M.R.; Zhao, C. Hypoglycemic activity and gut microbiota regulation of a novel polysaccharide from Grifola frondosa in type 2 diabetic mice. Food Chem. Toxicol. 2019, 126, 295–302. [Google Scholar] [CrossRef]
- Shao, T.; Yuan, P.; Zhu, L.; Xu, H.; Li, X.; He, S.; Wang, G.; Chen, K. Caebon nanoparticles inhibit A-glucosidase activity and induce a hypoglycemic effect in diabetic mice. Molecules 2019, 24, 3257. [Google Scholar] [CrossRef]
- Cheng, J.W.; Wang, Y.B.; Liang, H.; Qian, H.; Fu, L.Z.; Li, H.B. Optimization of the fermentation process for the production of intracellular polysaccharides from paecilomyces cicadae and the immuno-stimulating activity of intracellular polysaccharides. World J. Microbiol. Biotechnol. 2012, 28, 3293–3299. [Google Scholar] [CrossRef] [PubMed]
- Gericke, M.; Schulze, P.; Heinze, T. Nanoparticles based on hydrophobic polysaccharide Derivatives-Formation principles, characterization techniques, and biomedical applications. Macromol. Biosci. 2020, 20, 1900415. [Google Scholar] [CrossRef] [PubMed]
- Olatunji, O.J.; Yan, F.; Olatunji, O.O.; Jian, T.; Zhen, O.; Su, Z.; Wang, D.; Yu, X. Neuroprotective effects of adenosine isolated from Cordyceps cicadae against oxidative and er stress damages induced by glutamate in PC12 cells. Environ. Toxicol. Pharmacol. 2016, 44, 53–61. [Google Scholar] [CrossRef]
- Das, S.K.; Masuda, M.; Sakurai, A.; Sakakibara, M. Medicinal uses of the mushroom Cordyceps militaris: Current state and prospects. Fitoterapia 2010, 81, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Dedhia, N.; Marathe, S.J.; Singhal, R.S. Food polysaccharides: A review on emerging microbial sources, bioactivities, nanoformulations and safety considerations. Carbohydr. Polym. 2022, 287, 119355. [Google Scholar] [CrossRef]
- Yang, X.Y.; Ren, Y.M.; Zhang, L.N.; Wang, Z.W.; Li, L. Structural characteristics and antioxidant properties of exopolysaccharides isolated from soybean protein gel induced by lactic acid bacteria. LWT 2021, 150, 111811. [Google Scholar] [CrossRef]
- Zhu, H.J.; Tian, L.; Zhang, L.; Bi, J.X.; Song, Q.Q.; Yang, H.; Qiao, J. Preparation, characterization and antioxidant activity of polysaccharide from spent Lentinus edodes substrate. Int. J. Biol. Macromol. 2018, 112, 976–984. [Google Scholar] [CrossRef]
- He, S.D.; Wang, X.; Zhang, Y.; Wang, J.; Sun, H.J.; Wang, J.H.; Cao, X.D.; Ye, Y.K. Isolation and prebiotic activity of water-soluble polysaccharides fractions from the bamboo shoots (Phyllostachys praecox). Carbohydr. Polym. 2016, 151, 295–304. [Google Scholar] [CrossRef]
- Kowalczyk, A.; Szpakowska, N.; Babinska, W.; Motyka-Pomagruk, A.; Sledz, W.; Lojkowska, E.; Kaczyński, Z. The structure of an abequose—Containing O-polysaccharide isolated from Pectobacterium aquaticum IFB5637. Carbohydr. Res. 2022, 522, 108696. [Google Scholar] [CrossRef]
- Zhai, Z.Y.; Chen, A.; Zhou, H.M.; Zhang, D.Y.; Du, X.H.; Liu, Q.; Wu, X.Y.; Cheng, J.; Chen, L.J.; Hu, F.; et al. Structural characterization and functional activity of an exopolysaccharide secreted by Rhodopseudomonas palustris GJ-22. Int. J. Biol. Macromol. 2021, 167, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Lakra, A.K.; Domdi, L.; Tilwani, Y.M.; Arul, V. Physicochemical and functional characterization of mannan exopolysaccharide from Weissella confusa MD1 with bioactivities. Int. J. Biol. Macromol. 2020, 143, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Liu, F.; Peng, Q.; Wang, M. Purification, structural characterization, and hypolipidemic effects of a neutral polysaccharide from Ziziphus Jujuba cv. Muzao. Food Chem. 2018, 245, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.F.; Zhou, Y.B.; Zhang, Q.; Zhang, K.; Pai, P.; Chen, L.C.; Xiao, B. Hydrothermal extraction, structural characterization and inhibition HeLa cells proliferation of functional polysaccharides from Chinese tea Zhongcha 108. J. Funct. Foods 2017, 39, 1–8. [Google Scholar] [CrossRef]
- Hong, P.; Na, W.; Hu, Z.; Yu, Z.; Liu, Y.; Zhang, J.; Ruan, R. Physicochemical characterization of hemicelluloses from bamboo (Phyllostachys pubescens Mazel) stem. Ind. Crops Prod. 2012, 37, 41–50. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, L.; Chen, F. Production and characterization of exopolysaccharides from Chlorella zofingiensis and Chlorella vulgaris with anticolorectal cancer activity. Int. J. Biol. Macromol. 2019, 134, 976–983. [Google Scholar] [CrossRef]
- Ren, Q.; Chen, J.; Ding, Y.; Cheng, S.H.; Yang, S.; Ding, Z.H.; Dai, Q.Y.; Ding, Z. In vitro antioxidant and immunostimulating activities of polysaccharides from Ginkgo biloba leaves. Int. J. Biol. Macromol. 2019, 124, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.H.; Zhou, S.; Olatunji, O.J.; Wu, Y. Nucleosides rich extract from Cordyceps cicadae alleviated cisplatin-induced neurotoxicity in rats: A behavioral, biochemical and histopathological study. Arab. J. Chem. 2022, 15, 103476. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bitter, T.; Muir, H.M. A modified uronic acid carbazole reaction. Anal. Biochem. 1962, 4, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Nep, E.I.; Carnachan, S.M.; Ngwuluka, N.C.; Kontogiorgos, V.; Morris, G.A.; Sims, I.M.; Smith, A. Structural characterization and rheological properties of a polysaccharide from sesame leaves (Sesamum radiatum Schumach. Thonn.). Carbohydr. Polym. 2016, 152, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Drouillard, S.; Poulet, L.; Marechal, E.; Amato, A.; Buon, L.; Loiodice, M.; Helbert, W. Structure and enzymatic degradation of the polysaccharide secreted by Nostoc commune. Carbohydr. Res. 2022, 515, 108544. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | RT | Methylated Sugar | The Molar Rat(%) | Type of Linkage |
|---|---|---|---|---|
| SPP | 17.567 | 2,3,5-Me3-Araf | 0.194 | Araf-(1⟶ |
| 19.151 | 2,3,4-Me3-Xylp | 0.005 | Xylp-(1⟶ | |
| 23.73 | 2,3-Me2-Araf | 0.372 | ⟶5)-Araf-(1⟶ | |
| 25.77 | 2,3,4,6-Me4-Manp | 0.029 | Manp-(1⟶ | |
| 26.899 | 2,3,4,6-Me4-Galp | 0.006 | Galp-(1⟶ | |
| 28.309 | 2-Me1-Araf | 0.095 | ⟶3,5)-Araf-(1⟶ | |
| 30.974 | 3,4,6-Me3-Manp | 0.097 | ⟶2)-Manp-(1⟶ | |
| 31.849 | 2,3,6-Me3-Glcp | 0.107 | ⟶4)-Glcp-(1⟶ | |
| 33.141 | 2,3,4-Me3-Manp | 0.011 | ⟶6)-Manp-(1⟶ | |
| 38.611 | 2,3-Me2-Glcp | 0.009 | ⟶4,6)-Glcp-(1⟶ | |
| 39.054 | 2,3-Me2-Galp | 0.018 | ⟶4,6)-Galp-(1⟶ | |
| 39.441 | 3,4-Me2-Manp | 0.056 | ⟶2,6)-Manp-(1⟶ | |
| BSP | 17.556 | 2,3,5-Me3-Araf | 0.058 | Araf-(1⟶ |
| 23.676 | 2,3-Me2-Araf | 0.050 | ⟶5)-Araf-(1⟶ | |
| 25.729 | 2,3,4,6-Me4-Glcp | 0.060 | Glcp-(1⟶ | |
| 28.276 | 2-Me1-Araf | 0.017 | ⟶3,5)-Araf-(1⟶ | |
| 31.164 | 2,4,6-Me3-Glcp | 0.017 | ⟶3)-Glcp-(1⟶ | |
| 31.837 | 2,3,6-Me3-Glcp | 0.737 | ⟶4)-Glcp-(1⟶ | |
| 35.936 | 2,6-Me2-Glcp | 0.014 | ⟶3,4)-Glcp-(1⟶ | |
| 38.985 | 2,3-Me2-Glcp | 0.046 | ⟶4,6)-Glcp-(1⟶ | |
| PPP | 17.632 | 2,3,5-Me3-Araf | 0.112 | Araf-(1⟶ |
| 19.213 | 2,3,4-Me3-Xylp | 0.007 | Xylp-(1⟶ | |
| 23.802 | 2,3-Me2-Araf | 0.159 | ⟶5)-Araf-(1⟶ | |
| 25.852 | 2,3,4,6-Me4-Manp | 0.049 | Manp-(1⟶ | |
| 26.974 | 2,3,4,6-Me4-Galp | 0.006 | Galp-(1⟶ | |
| 28.393 | 2-Me1-Araf | 0.036 | ⟶3,5)-Araf-(1⟶ | |
| 31.09 | 3,4,6-Me3-Manp | 0.140 | ⟶2)-Manp-(1⟶ | |
| 31.296 | 2,4,6-Me3-Glcp | 0.005 | ⟶3)-Glcp-(1⟶ | |
| 31.46 | 2,3,6-Me3-Galp | 0.014 | ⟶4)-Galp-(1⟶ | |
| 32.086 | 2,3,6-Me3-Glcp | 0.258 | ⟶4)-Glcp-(1⟶ | |
| 32.48 | 2,3,4-Me3-Glcp | 0.006 | ⟶6-Glcp-(1⟶ | |
| 33.23 | 2,3,4-Me3-Manp | 0.021 | ⟶6)-Manp-(1⟶ | |
| 35.968 | 2,6-Me2-Glcp | 0.023 | ⟶3,4)-Glcp-(1⟶ | |
| 39.172 | 2,3-Me2-Glcp | 0.043 | ⟶4,6)-Glcp-(1⟶ | |
| 39.594 | 3,4-Me2-Manp | 0.123 | ⟶2,6)-Manp-(1⟶ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zeng, T.; Li, H.; Wang, Y.; Wang, J.; Yuan, H. Structural Characterization and Hypoglycemic Function of Polysaccharides from Cordyceps cicadae. Molecules 2023, 28, 526. https://doi.org/10.3390/molecules28020526
Wang Y, Zeng T, Li H, Wang Y, Wang J, Yuan H. Structural Characterization and Hypoglycemic Function of Polysaccharides from Cordyceps cicadae. Molecules. 2023; 28(2):526. https://doi.org/10.3390/molecules28020526
Chicago/Turabian StyleWang, Yani, Tingting Zeng, Hang Li, Yidi Wang, Junhui Wang, and Huaibo Yuan. 2023. "Structural Characterization and Hypoglycemic Function of Polysaccharides from Cordyceps cicadae" Molecules 28, no. 2: 526. https://doi.org/10.3390/molecules28020526
APA StyleWang, Y., Zeng, T., Li, H., Wang, Y., Wang, J., & Yuan, H. (2023). Structural Characterization and Hypoglycemic Function of Polysaccharides from Cordyceps cicadae. Molecules, 28(2), 526. https://doi.org/10.3390/molecules28020526