
The Role of Estrogen Signaling and Exercise in Drug Abuse: A Review




Abstract
1. Introduction
2. Drugs of Abuse: Mechanisms of Action
2.1. General Mechanisms
2.2. Alcohol
2.3. Cocaine
2.4. Cannabinoids
2.5. Heroin/Opioids
3. Sex Differences in Drug Abuse
3.1. Human Studies
3.2. Animal Studies
4. Effect of Estrogen on Drug Abuse
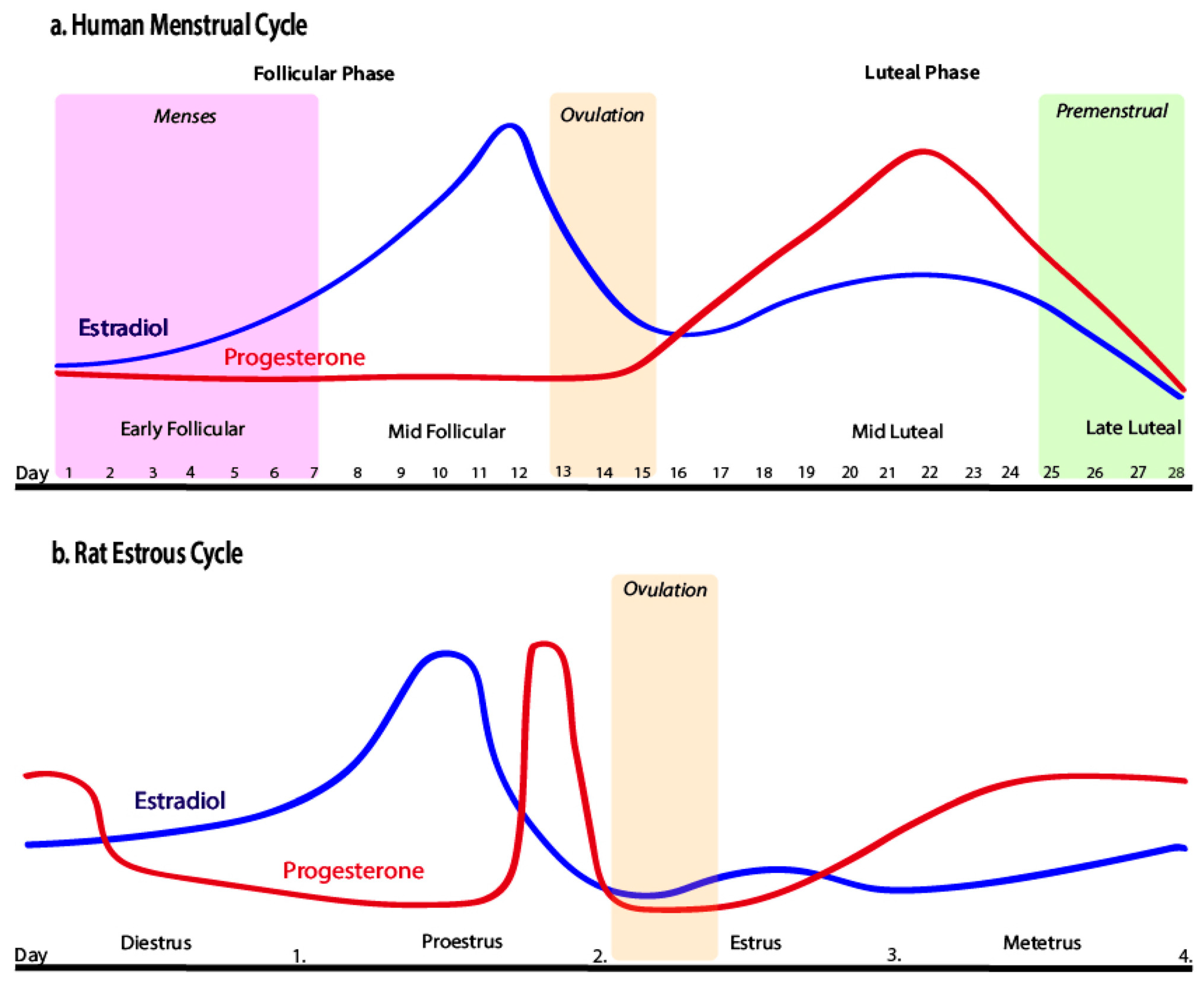
4.1. Ovarian Cycles
4.2. Ovariectomy
4.3. Estrogen: Mechanisms of Action Males vs. Females
5. Treatments for Drug Abuse
5.1. Pharmacotherapy
5.2. Exercise
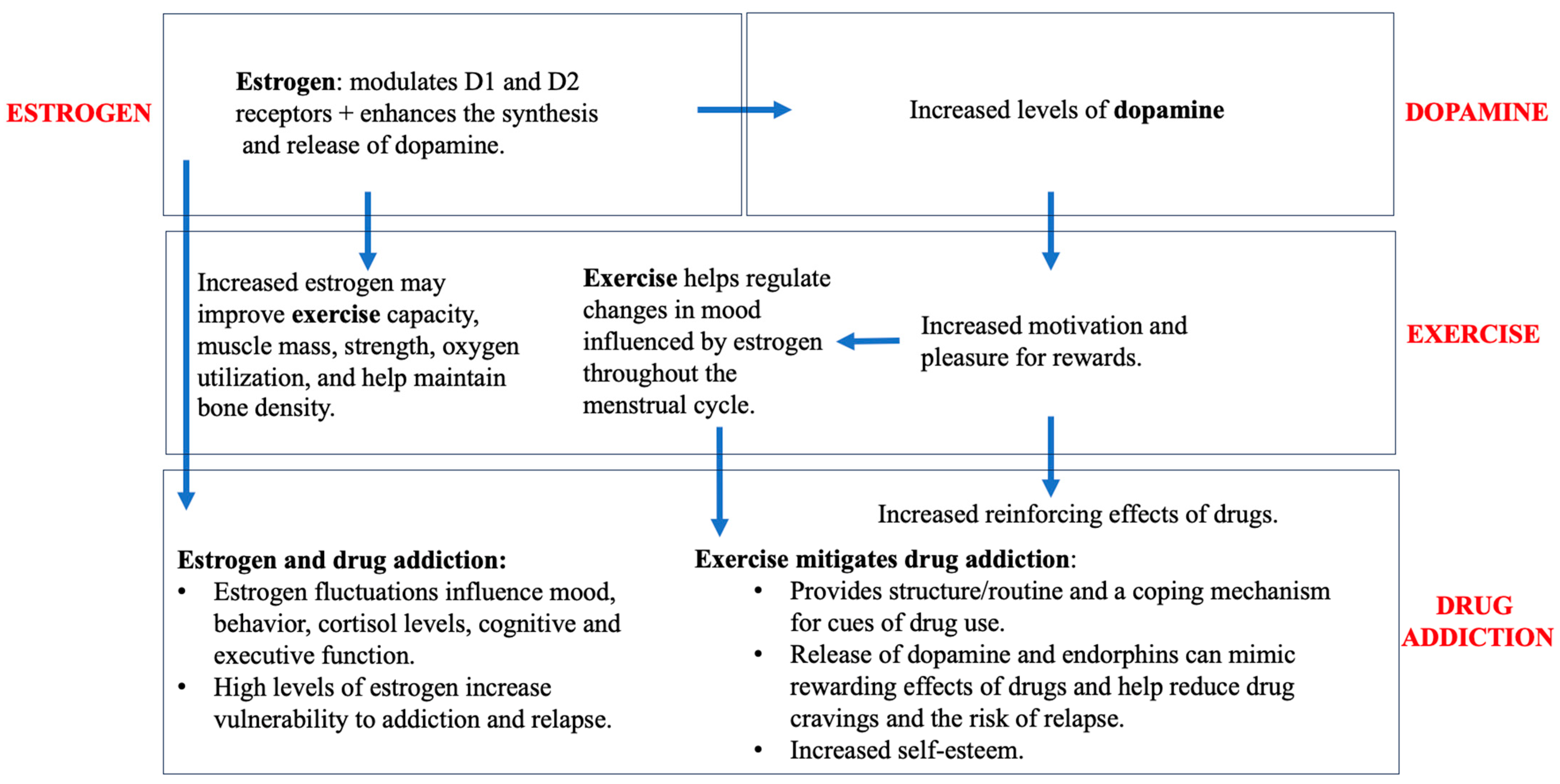
6. Exercise and Estrogen
6.1. Wheel Running
6.2. Treadmill
7. Limitations and Future Directions
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- U.S. Department of Health and Human Services; Substance Abuse and Mental Health Services Administration; Center for Behavioral Health Statistics and Quality. National Survey on Drug Use and Health 2021; Substance Abuse and Mental Health Services Administration (SAMHSA): Washington, DC, USA, 2023.
- Zhou, Y.; Zhao, M.; Zhou, C.; Li, R. Sex differences in drug addiction and response to exercise intervention: From human to animal studies. Front. Neuroendocrinol. 2016, 40, 24–41. [Google Scholar] [CrossRef] [PubMed]
- Jong, I.E.M.; Kloet, E.R. Glucocorticoids and Vulnerability to Psychostimulant Drugs: Toward Substrate and Mechanism. Ann. N. Y. Acad. Sci. 2004, 1018, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Lüscher, C. The Emergence of a Circuit Model for Addiction. Annu. Rev. Neurosci. 2016, 39, 257–276. [Google Scholar] [CrossRef]
- Pierce, R.C.; Wolf, M.E. Psychostimulant-Induced Neuroadaptations in Nucleus Accumbens AMPA Receptor Transmission. Cold Spring Harb. Perspect. Med. 2013, 3, a012021. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-Y.; Qi, J.; Han, W.-Y.; Wang, F.; Wu, C.-F. Inhibitory role of oxytocin in psychostimulant-induced psychological dependence and its effects on dopaminergic and glutaminergic transmission. Acta Pharmacol. Sin. 2010, 31, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.-Y.; Long, J.-D.; Liu, Y.; Liu, J.-G. Formation of aversive memories associated with conditioned drug withdrawal requires BDNF expression in the amygdala in acute morphine-dependent rats. Acta Pharmacol. Sin. 2015, 36, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Buck, C.L.; Cohen, A.; Edwards, S.; Park, P.E.; Schlosburg, J.E.; Schmeichel, B.; Vendruscolo, L.F.; Wade, C.L.; Whitfield, T.W.; et al. Addiction as a stress surfeit disorder. Neuropharmacology 2014, 76, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.-H.; Hou, Y.-Y.; Xi, T.; Liu, Y.; Liu, J.-G. The small GTPase RhoA, but not Rac1, is essential for conditioned aversive memory formation through regulation of actin rearrangements in rat dorsal hippocampus. Acta Pharmacol. Sin. 2013, 34, 811–818. [Google Scholar] [CrossRef][Green Version]
- Hamilton, K.L.; Harris, A.C.; Gewirtz, J.C. Affective and neuroendocrine effects of withdrawal from chronic, long-acting opiate administration. Brain Res. 2013, 1538, 73–82. [Google Scholar] [CrossRef][Green Version]
- Ahmed, R.; Blum, K.; Thanos, P.K. Epigenetic Effects of Psychoactive Drugs. Curr. Pharm. Des. 2023, 29, 2124–2139. [Google Scholar] [CrossRef]
- Koob, G.F.; Volkow, N.D. Neurobiology of addiction: A neurocircuitry analysis. Lancet Psychiatry 2016, 3, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Gilpin, N.W.; Koob, G.F. Neurobiology of alcohol dependence: Focus on motivational mechanisms. Alcohol Res. Health J. Natl. Inst. Alcohol Abus. Alcohol. 2008, 31, 185–195. [Google Scholar]
- Herbison, A.E. Estrogen regulation of GABA transmission in rat preoptic area. Brain Res. Bull. 1997, 44, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Morena, M.; Patel, S.; Bains, J.S.; Hill, M.N. Neurobiological Interactions Between Stress and the Endocannabinoid System. Neuropsychopharmacology 2016, 41, 80–102. [Google Scholar] [CrossRef] [PubMed]
- Henricks, A.M.; Berger, A.L.; Lugo, J.M.; Baxter-Potter, L.N.; Bieniasz, K.V.; Petrie, G.; Sticht, M.A.; Hill, M.N.; McLaughlin, R.J. Sex- and hormone-dependent alterations in alcohol withdrawal-induced anxiety and corticolimbic endocannabinoid signaling. Neuropharmacology 2017, 124, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.M.; Richards, Q.; Keith, D.R. The long-term effects of cocaine use on cognitive functioning: A systematic critical review. Behav. Brain Res. 2018, 348, 241–262. [Google Scholar] [CrossRef] [PubMed]
- Hudson, A.; Stamp, J.A. Ovarian hormones and propensity to drug relapse: A review. Neurosci. Biobehav. Rev. 2011, 35, 427–436. [Google Scholar] [CrossRef]
- Calipari, E.S.; Juarez, B.; Morel, C.; Walker, D.M.; Cahill, M.E.; Ribeiro, E.; Roman-Ortiz, C.; Ramakrishnan, C.; Deisseroth, K.; Han, M.H.; et al. Dopaminergic dynamics underlying sex-specific cocaine reward. Nat. Commun. 2017, 8, 13877. [Google Scholar] [CrossRef]
- Silveira, M.M.; Arnold, J.C.; Laviolette, S.R.; Hillard, C.J.; Celorrio, M.; Aymerich, M.S.; Adams, W.K. Seeing through the smoke: Human and animal studies of cannabis use and endocannabinoid signalling in corticolimbic networks. Neurosci. Biobehav. Rev. 2017, 76, 380–395. [Google Scholar] [CrossRef]
- Parsons, L.H.; Hurd, Y.L. Endocannabinoid signalling in reward and addiction. Nat. Rev. Neurosci. 2015, 16, 579–594. [Google Scholar] [CrossRef]
- Walker, O.S.; Holloway, A.C.; Raha, S. The role of the endocannabinoid system in female reproductive tissues. J. Ovarian Res. 2019, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.J. The effect of opioid therapy on endocrine function. Am. J. Med. 2013, 126 (Suppl. 1), S12–S18. [Google Scholar] [CrossRef] [PubMed]
- Al-Hasani, R.; Bruchas, M.R. Molecular Mechanisms of Opioid Receptor-dependent Signaling and Behavior. Anesthesiology 2011, 115, 1363–1381. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.B.; Koob, G.F. Sex Differences in Animal Models: Focus on Addiction. Pharmacol. Rev. 2016, 68, 242–263. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.B.; McClellan, M.L.; Reed, B.G. Sex differences, gender and addiction. J. Neurosci. Res. 2017, 95, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Wetherill, R.R.; Franklin, T.R.; Allen, S.S. Ovarian Hormones, Menstrual Cycle Phase, and Smoking: A Review with Recommendations for Future Studies. Curr. Addict. Rep. 2016, 3, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kerstetter, K.A.; Ballis, M.A.; Duffin-Lutgen, S.; Carr, A.E.; Behrens, A.M.; Kippin, T.E. Sex Differences in Selecting Between Food and Cocaine Reinforcement are Mediated by Estrogen. Neuropsychopharmacology 2012, 37, 2605–2614. [Google Scholar] [CrossRef]
- Lynch, W.J. Modeling the development of drug addiction in male and female animals. Pharmacol. Biochem. Behav. 2018, 164, 50–61. [Google Scholar] [CrossRef]
- Moyer, A.M.; Matey, E.T.; Miller, V.M. Individualized medicine: Sex, hormones, genetics, and adverse drug reactions. Pharmacol. Res. Perspect. 2019, 7, e00541. [Google Scholar] [CrossRef]
- Joyce, K.M.; Good, K.P.; Tibbo, P.; Brown, J.; Stewart, S.H. Addictive behaviors across the menstrual cycle: A systematic review. Arch. Womens Ment. Health 2021, 24, 529–542. [Google Scholar] [CrossRef]
- Terner, J.M.; de Wit, H. Menstrual cycle phase and responses to drugs of abuse in humans. Drug Alcohol Depend. 2006, 84, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.G.; Fallon, V.M.; Goodwin, L.; Gage, S.H.; Rose, A.K. Menstrual Cycle Phase, Hormonal Contraception, and Alcohol Consumption in Premenopausal Females: A Systematic Review. Front. Glob. Womens Health 2021, 2, 745263. [Google Scholar] [CrossRef]
- Roche, D.J.; King, A.C. Sex differences in acute hormonal and subjective response to naltrexone: The impact of menstrual cycle phase. Psychoneuroendocrinology 2015, 52, 59–71. [Google Scholar] [CrossRef] [PubMed]
- File, D.; Bőthe, B.; File, B.; Demetrovics, Z. The Role of Impulsivity and Reward Deficiency in “Liking” and “Wanting” of Potentially Problematic Behaviors and Substance Uses. Front. Psychiatry 2022, 13, 820836. [Google Scholar] [CrossRef] [PubMed]
- Evans, S.M.; Haney, M.; Foltin, R.W. The effects of smoked cocaine during the follicular and luteal phases of the menstrual cycle in women. Psychopharmacology 2002, 159, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.J.; Collins, A.L.; Sanford, C.A.; Phillips, P.E.M. Dopamine Encoding of Pavlovian Incentive Stimuli Diminishes with Extended Training. J. Neurosci. 2013, 33, 3526–3532. [Google Scholar] [CrossRef]
- Difeliceantonio, A.G.; Berridge, K.C. Dorsolateral neostriatum contribution to incentive salience: Opioid or dopamine stimulation makes one reward cue more motivationally attractive than another. Eur. J. Neurosci. 2016, 43, 1203–1218. [Google Scholar] [CrossRef]
- Berridge, K.C. The debate over dopamine’s role in reward: The case for incentive salience. Psychopharmacology 2007, 191, 391–431. [Google Scholar] [CrossRef]
- Reichel, C.M.; Chan, C.H.; Ghee, S.M.; See, R.E. Sex differences in escalation of methamphetamine self-administration: Cognitive and motivational consequences in rats. Psychopharmacology 2012, 223, 371–380. [Google Scholar] [CrossRef]
- Fattore, L.; Fadda, P.; Fratta, W. Sex differences in the self-administration of cannabinoids and other drugs of abuse. Psychoneuroendocrinology 2009, 34, S227–S236. [Google Scholar] [CrossRef]
- Kerstetter, K.A.; Aguilar, V.R.; Parrish, A.B.; Kippin, T.E. Protracted time-dependent increases in cocaine-seeking behavior during cocaine withdrawal in female relative to male rats. Psychopharmacology 2008, 198, 63–75. [Google Scholar] [CrossRef]
- Fattore, L.; Spano, M.S.; Altea, S.; Angius, F.; Fadda, P.; Fratta, W. Cannabinoid self-administration in rats: Sex differences and the influence of ovarian function. Br. J. Pharmacol. 2007, 152, 795–804. [Google Scholar] [CrossRef]
- Jackson, L.R.; Robinson, T.E.; Becker, J.B. Sex Differences and Hormonal Influences on Acquisition of Cocaine Self-Administration in Rats. Neuropsychopharmacology 2006, 31, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Parada, M.; Vargas, E.B.; Kyres, M.; Burnside, K.; Pfaus, J.G. The role of ovarian hormones in sexual reward states of the female rat. Horm. Behav. 2012, 62, 442–447. [Google Scholar] [CrossRef]
- Torres, O.V.; Natividad, L.A.; Tejeda, H.A.; Van Weelden, S.A.; O’Dell, L.E. Female rats display dose-dependent differences to the rewarding and aversive effects of nicotine in an age-, hormone-, and sex-dependent manner. Psychopharmacology 2009, 206, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Feltenstein, M.W.; Byrd, E.A.; Henderson, A.R.; See, R.E. Attenuation of cocaine-seeking by progesterone treatment in female rats. Psychoneuroendocrinology 2009, 34, 343–352. [Google Scholar] [CrossRef]
- Feltenstein, M.W.; Henderson, A.R.; See, R.E. Enhancement of cue-induced reinstatement of cocaine-seeking in rats by yohimbine: Sex differences and the role of the estrous cycle. Psychopharmacology 2011, 216, 53–62. [Google Scholar] [CrossRef]
- Ramôa, C.P.; Doyle, S.E.; Naim, D.W.; Lynch, W.J. Estradiol as a Mechanism for Sex Differences in the Development of an Addicted Phenotype following Extended Access Cocaine Self-Administration. Neuropsychopharmacology 2013, 38, 1698–1705. [Google Scholar] [CrossRef]
- Larson, E.B.; Anker, J.J.; Gliddon, L.A.; Fons, K.S.; Carroll, M.E. Effects of estrogen and progesterone on the escalation of cocaine self-administration in female rats during extended access. Exp. Clin. Psychopharmacol. 2007, 15, 461–471. [Google Scholar] [CrossRef]
- Luoni, A.; Berry, A.; Raggi, C.; Bellisario, V.; Cirulli, F.; Riva, M.A. Sex-Specific Effects of Prenatal Stress on Bdnf Expression in Response to an Acute Challenge in Rats: A Role for Gadd45β. Mol. Neurobiol. 2016, 53, 7037–7047. [Google Scholar] [CrossRef]
- Becker, J.B.; Perry, A.N.; Westenbroek, C. Sex differences in the neural mechanisms mediating addiction: A new synthesis and hypothesis. Biol. Sex Differ. 2012, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Handa, R.J.; Weiser, M.J. Gonadal steroid hormones and the hypothalamo–pituitary–adrenal axis. Front. Neuroendocrinol. 2014, 35, 197–220. [Google Scholar] [CrossRef] [PubMed]
- Kippin, T.E.; Fuchs, R.A.; Mehta, R.H.; Case, J.M.; Parker, M.P.; Bimonte-Nelson, H.A.; See, R.E. Potentiation of cocaine-primed reinstatement of drug seeking in female rats during estrus. Psychopharmacology 2005, 182, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Pestana, J.E.; Islam, N.; Van der Eyk, N.L.; Graham, B.M. What Pre-clinical Rat Models Can Tell Us About Anxiety Across the Menstrual Cycle in Healthy and Clinically Anxious Humans. Curr. Psychiatry 2022, 24, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, R.A.; Evans, K.A.; Mehta, R.H.; Case, J.M.; See, R.E. Influence of sex and estrous cyclicity on conditioned cue-induced reinstatement of cocaine-seeking behavior in rats. Psychopharmacology 2005, 179, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Dreher, J.C.; Schmidt, P.J.; Kohn, P.; Furman, D.; Rubinow, D.; Berman, K.F. Menstrual cycle phase modulates reward-related neural function in women. Proc. Natl. Acad. Sci. USA 2007, 104, 2465–2470. [Google Scholar] [CrossRef]
- Li, L.H.; Wang, Z.C.; Yu, J.; Zhang, Y.Q. Ovariectomy results in variable changes in nociception, mood and depression in adult female rats. PLoS ONE 2014, 9, e94312. [Google Scholar] [CrossRef]
- Stoffel, E.C.; Craft, R.M. Ovarian hormone withdrawal-induced “depression” in female rats. Physiol. Behav. 2004, 83, 505–513. [Google Scholar] [CrossRef]
- Perreault, B.; Hammond, N.; Thanos, P.K. Effects of Exercise on Testosterone and Implications of Drug Abuse: A Review. Clin. Neuropharmacol. 2023, 46, 112–122. [Google Scholar] [CrossRef]
- Phan, A.; Lancaster, K.E.; Armstrong, J.N.; Maclusky, N.J.; Choleris, E. Rapid Effects of Estrogen Receptor α and β Selective Agonists on Learning and Dendritic Spines in Female Mice. Endocrinology 2011, 152, 1492–1502. [Google Scholar] [CrossRef]
- Newman, A.H.; Blaylock, B.L.; Nader, M.A.; Bergman, J.; Sibley, D.R.; Skolnick, P. Medication discovery for addiction: Translating the dopamine D3 receptor hypothesis. Biochem. Pharmacol. 2012, 84, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Boyle, M. Neuroscience of Addiction: Relevance to Prevention and Treatment. Am. J. Psychiatry 2018, 175, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Gandía, M.C.; Rodríguez-Arias, M. Pharmacological treatments for opiate and alcohol addiction: A historical perspective of the last 50 years. Eur. J. Pharmacol. 2018, 836, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.B.; Mariani, J.J.; Levin, F.R. New directions in the treatment of opioid withdrawal. Lancet 2020, 395, 1938–1948. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Blanco, C. The changing opioid crisis: Development, challenges, and opportunities. Mol. Psychiatry 2021, 26, 218–233. [Google Scholar] [CrossRef]
- Burnette, E.M.; Nieto, S.J.; Grodin, E.N.; Meredith, L.R.; Hurley, B.; Miotto, K.; Gillis, A.J.; Ray, L.A. Novel Agents for the Pharmacological Treatment of Alcohol Use Disorder. Drugs 2022, 82, 251–274. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Khan, H.; Kaur, A.; Singh, T.G. Novel Targets Explored in the Treatment of Alcohol Withdrawal Syndrome. CNS Neurol. Disord. Drug Targets 2021, 20, 158–173. [Google Scholar] [CrossRef]
- More, A.; Jackson, B.; Dimmock, J.A.; Thornton, A.L.; Colthart, A.; Furzer, B.J. Exercise in the Treatment of Youth Substance Use Disorders: Review and Recommendations. Front. Psychol. 2017, 8, 1839. [Google Scholar] [CrossRef]
- Rosenthal, A.; Ebrahimi, C.; Wedemeyer, F.; Romanczuk-Seiferth, N.; Beck, A. The Treatment of Substance Use Disorders: Recent Developments and New Perspectives. Neuropsychobiology 2022, 81, 451–472. [Google Scholar] [CrossRef]
- Venniro, M.; Shaham, Y. An operant social self-administration and choice model in rats. Nat. Protoc. 2020, 15, 1542–1559. [Google Scholar] [CrossRef]
- Gardner, E.L. Addiction and Brain Reward and Antireward Pathways. Adv. Psychosom. Med. 2011, 30, 22–60. [Google Scholar] [CrossRef] [PubMed]
- Bauer, J.; Pedersen, A.; Scherbaum, N.; Bening, J.; Patschke, J.; Kugel, H.; Heindel, W.; Arolt, V.; Ohrmann, P. Craving in Alcohol-Dependent Patients After Detoxification Is Related to Glutamatergic Dysfunction in the Nucleus Accumbens and the Anterior Cingulate Cortex. Neuropsychopharmacology 2013, 38, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Wagner, L.; Howe, K.; Philbrick, K.A.; Maddalozzo, G.F.; Kuah, A.F.; Wong, C.P.; Olson, D.A.; Branscum, A.J.; Iwaniec, U.T.; Turner, R.T. Effects of Alcohol and Estrogen Receptor Blockade Using ICI 182,780 on Bone in Ovariectomized Rats. Alcohol Clin. Exp. Res. 2019, 43, 2301–2311. [Google Scholar] [CrossRef]
- Lynch, W.J.; Peterson, A.B.; Sanchez, V.; Abel, J.; Smith, M.A. Exercise as a novel treatment for drug addiction: A neurobiological and stage-dependent hypothesis. Neurosci. Biobehav. Rev. 2013, 37, 1622–1644. [Google Scholar] [CrossRef] [PubMed]
- Robison, L.; Swenson, S.; Hamilton, J.; Thanos, P.K. Exercise Reduces Dopamine D1R and Increases D2R in Rats: Implications for Addiction. Med. Sci. Sports Exerc. 2018, 50, 1596–1602. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, X. A Systematic Review of Exercise Intervention Program for People With Substance Use Disorder. Front. Psychiatry 2022, 13, 817927. [Google Scholar] [CrossRef] [PubMed]
- Alessi, S.M.; Rash, C.J.; Pescatello, L.S. Reinforcing exercise to improve drug abuse treatment outcomes: A randomized controlled study in a substance use disorder outpatient treatment setting. Psychol. Addict. Behav. 2020, 34, 52–64. [Google Scholar] [CrossRef]
- Konhilas, J.P.; Maass, A.H.; Luckey, S.W.; Stauffer, B.L.; Olson, E.N.; Leinwand, L.A. Sex modifies exercise and cardiac adaptation in mice. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H2768–H2776. [Google Scholar] [CrossRef]
- Niyomchai, T.; Jenab, S.; Festa, E.D.; Akhavan, A.; Quiñones-Jenab, V. Effects of short- and long-term estrogen and progesterone replacement on female rats after acute cocaine administration. Brain Res. 2006, 1126, 193–199. [Google Scholar] [CrossRef]
- Hertrampf, T.; Seibel, J.; Laudenbach, U.; Fritzemeier, K.H.; Diel, P. Analysis of the effects of oestrogen receptor α (ERα)- and ERβ-selective ligands given in combination to ovariectomized rats. Br. J. Pharmacol. 2008, 153, 1432–1437. [Google Scholar] [CrossRef]
- Cotman, C.W.; Engesser-Cesar, C. Exercise Enhances and Protects Brain Function. Exerc. Sport Sci. Rev. 2002, 30, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, E.; Curtis, K.S. Increased locomotor activity in estrogen-treated ovariectomized rats is associated with nucleus accumbens dopamine and is not reduced by dietary sodium deprivation. Integr. Zool. 2018, 13, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Grigsby, K.B.; Kovarik, C.M.; Mao, X.; Booth, F.W. Medial preoptic estrogen receptor-beta blunts the estrogen receptor-alpha mediated increases in wheel-running behavior of female rats. Behav. Brain Res. 2020, 379, 112341. [Google Scholar] [CrossRef] [PubMed]
- Ehringer, M.A.; Hoft, N.R.; Zunhammer, M. Reduced alcohol consumption in mice with access to a running wheel. Alcohol 2009, 43, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Walker, K.L.; Cole, K.T.; Lang, K.C. The effects of aerobic exercise on cocaine self-administration in male and female rats. Psychopharmacology 2011, 218, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Pichard, C.; Gorwood, P.A.P.M.; Hamon, M.; Cohen-Salmon, C. Differential effects of free versus imposed motor activity on alcohol consumption in C57BL/6J versus DBA/2J mice. Alcohol 2009, 43, 593–601. [Google Scholar] [CrossRef]
- Rauf, S.; Soejono, S.K.; Partadiredja, G. Effects of treadmill exercise training on cerebellar estrogen and estrogen receptors, serum estrogen, and motor coordination performance of ovariectomized rats. Iran. J. Basic Med. Sci. 2015, 18, 587–592. [Google Scholar]
- Kaidah, S.; Soejono, S.K.; Partadiredja, G. Exercise improves hippocampal estrogen and spatial memory of ovariectomized rats. Bratisl. Med. J. 2016, 117, 94–99. [Google Scholar] [CrossRef]
- Goldsmith, E.; Glaister, M. The effect of the menstrual cycle on running economy. J. Sports Med. Phys. Fit. 2020, 60, 610–617. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Drug | Menstrual Phase | Impact on Drug Abuse: Effects on Behavior and Mood | References |
|---|---|---|---|
| Cocaine | Follicular | ↑ mood altering effects: wanting more, energy feelings of euphoria. | [32,35] |
| Luteal | ↑ symptoms of dysphoria | [32,36] | |
| Alcohol | Follicular | ↑ anxiety and depression ↑ consumption overall ↑ consumption in those with premenstrual syndrome compared to control. ↑ consumption in those on hormonal contraceptives compared to control | [32,33] |
| Luteal | ↑ anxiety ↑ consumption | [32] | |
| No cycle differences. No significant effect of cycle phase on consumption/relationship inconclusive | [32] [33] | ||
| Opioids | Follicular | ↑ sensitivity to adverse effects | [34] |
| Luteal | ↑ cortisol and prolactin responses Significantly ↑ sensitivity to adverse effects compared to FP and men | [34] | |
| No cycle differences | [32] | ||
| Nicotine | Follicular | ↑ depressive symptoms, relapse tendency, physiological reactivity | [26,27] |
| Luteal | ↑ stimulation and cognitive task performance ↓ urge to smoke and reactivity | [27,32] | |
| Estrogen Level | Strain/Age | Cocaine Dose | Cocaine Self-Administration | References |
|---|---|---|---|---|
| Intact during estrus phase (high levels of estradiol) | Sprague-Dawley rats | 0.5 mg/kg per infusion | Increases | [47] |
| 0.5 mg/kg per infusion | Increases | [48] | ||
| Intact during non-estrus phases (low levels of estradiol) | Sprague-Dawley rats | 0.5 mg/kg per infusion | Decreases | [47] |
| 0.5 mg/kg per infusion | Decreases | [48] | ||
| Ovariectomized | Sprague-Dawley rats about 3 months of age | 1.5 mg/kg per infusion | Decreases | [49] |
| 0.4 mg/kg per infusion | Decreases | [50] | ||
| Ovariectomized with estradiol treatment | Sprague-Dawley rats about 3 months of age | 1.5 mg/kg per infusion | Increases | [49] |
| 0.4 mg/kg per infusion | Increases | [50] |
| Estrogen Receptors | Voluntary Wheel Running | References |
|---|---|---|
| ERα signaling in non-OVX | Increases | [81,82,83,84] |
| ERβ signaling in non-OVX | Decreases slightly | [81,84] |
| ERα agonist in OVX | Increases | [81] |
| ERβ agonist in OVX | No effect | [81] |
| ERα and ERβ agonists in OVX | No effect | [84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, R.; Zyla, S.; Hammond, N.; Blum, K.; Thanos, P.K. The Role of Estrogen Signaling and Exercise in Drug Abuse: A Review. Clin. Pract. 2024, 14, 148-163. https://doi.org/10.3390/clinpract14010012
Ahmed R, Zyla S, Hammond N, Blum K, Thanos PK. The Role of Estrogen Signaling and Exercise in Drug Abuse: A Review. Clinics and Practice. 2024; 14(1):148-163. https://doi.org/10.3390/clinpract14010012
Chicago/Turabian StyleAhmed, Rania, Samuel Zyla, Nikki Hammond, Kenneth Blum, and Panayotis K. Thanos. 2024. "The Role of Estrogen Signaling and Exercise in Drug Abuse: A Review" Clinics and Practice 14, no. 1: 148-163. https://doi.org/10.3390/clinpract14010012
APA StyleAhmed, R., Zyla, S., Hammond, N., Blum, K., & Thanos, P. K. (2024). The Role of Estrogen Signaling and Exercise in Drug Abuse: A Review. Clinics and Practice, 14(1), 148-163. https://doi.org/10.3390/clinpract14010012