

Butylated Hydroxyanisole (BHA) Disrupts Brain Signalling in Embryo–Larval Stage of Zebrafish Leading to Attention Deficit Hyperactivity Disorder (ADHD)

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Maintenance of Zebrafish, Embryo Collection, and BHA Exposure
2.3. Teratological Analysis of BHA-Exposed Zebrafish Larvae
2.4. Evaluation of Apoptotic Activity in BHA-Exposed Zebrafish Larvae Using Acridine Orange Staining
2.5. Neurobehavioural Assay of BHA-Exposed Zebrafish Larvae
2.6. Acetylcholinesterase (AChE) Activity
2.7. Quantification of Serotonin
2.8. Antioxidant Enzyme Assay of BHA-Exposed Zebrafish Larvae
2.9. Gene Expression
2.10. Statistical Analysis
3. Result
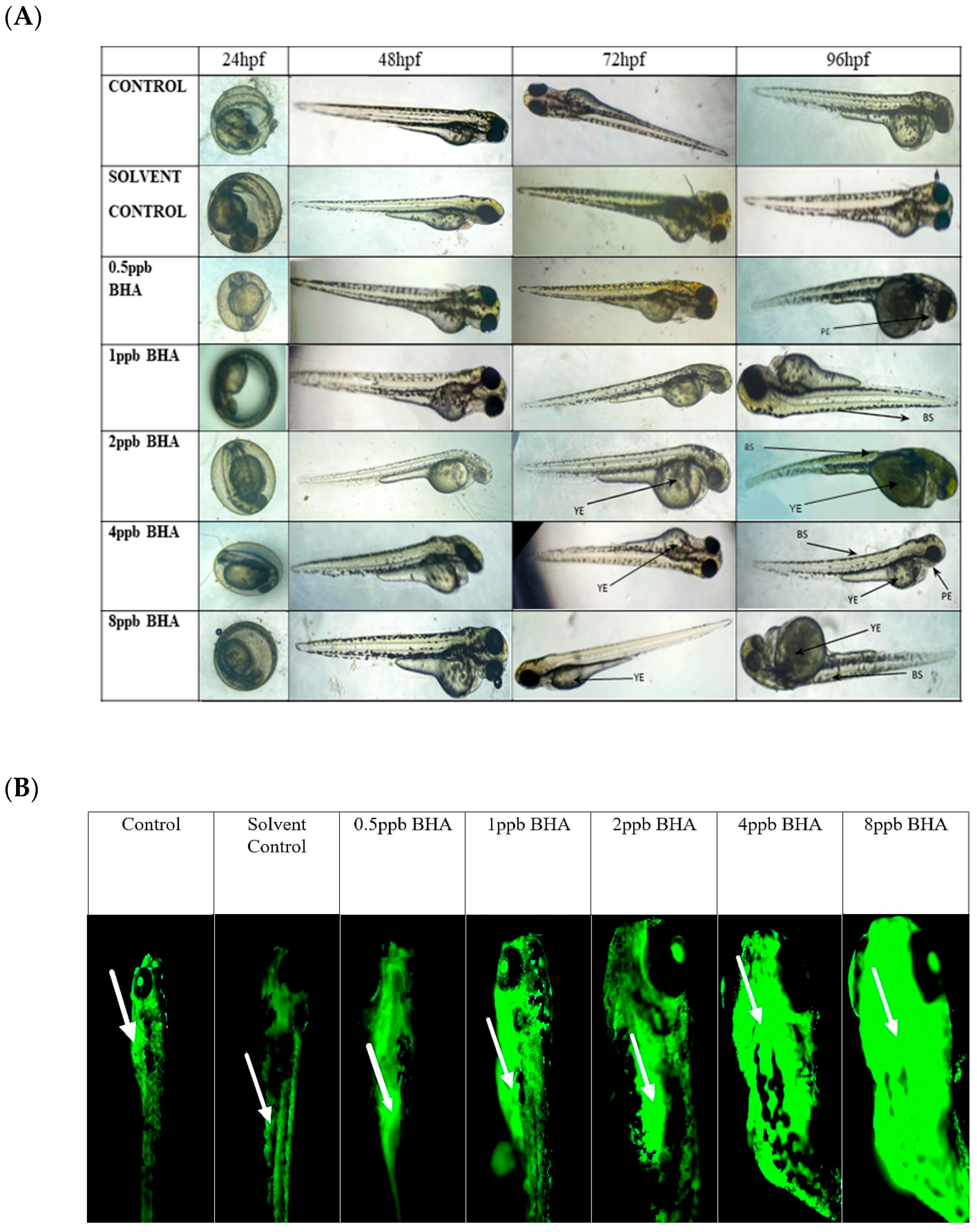
3.1. Teratological Effects of BHA-Exposed Zebrafish Larvae
3.2. Evaluation of Apoptotic Activity in BHA-Exposed Zebrafish Larvae Using Acridine Orange Staining
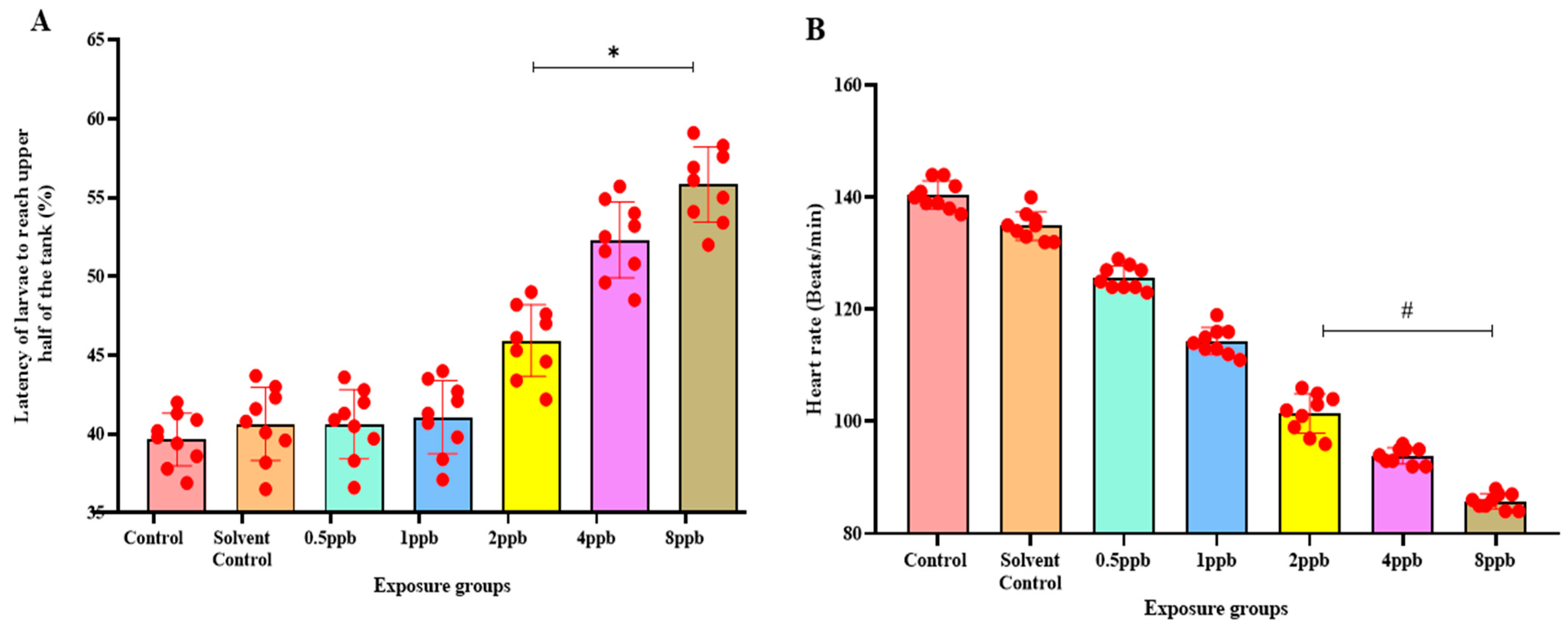
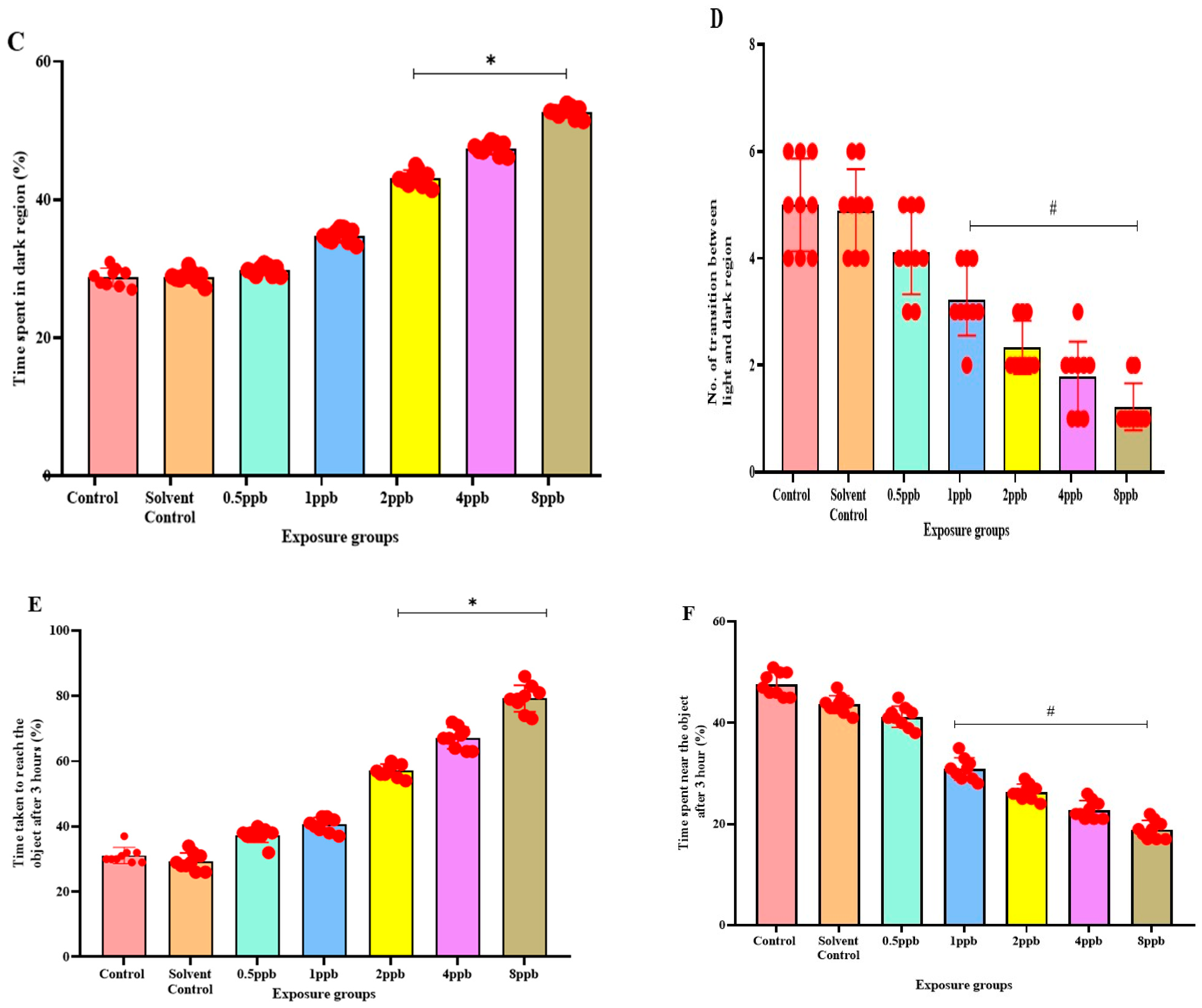
3.3. Effect of BHA on Neurobehaviour in Zebrafish Larvae
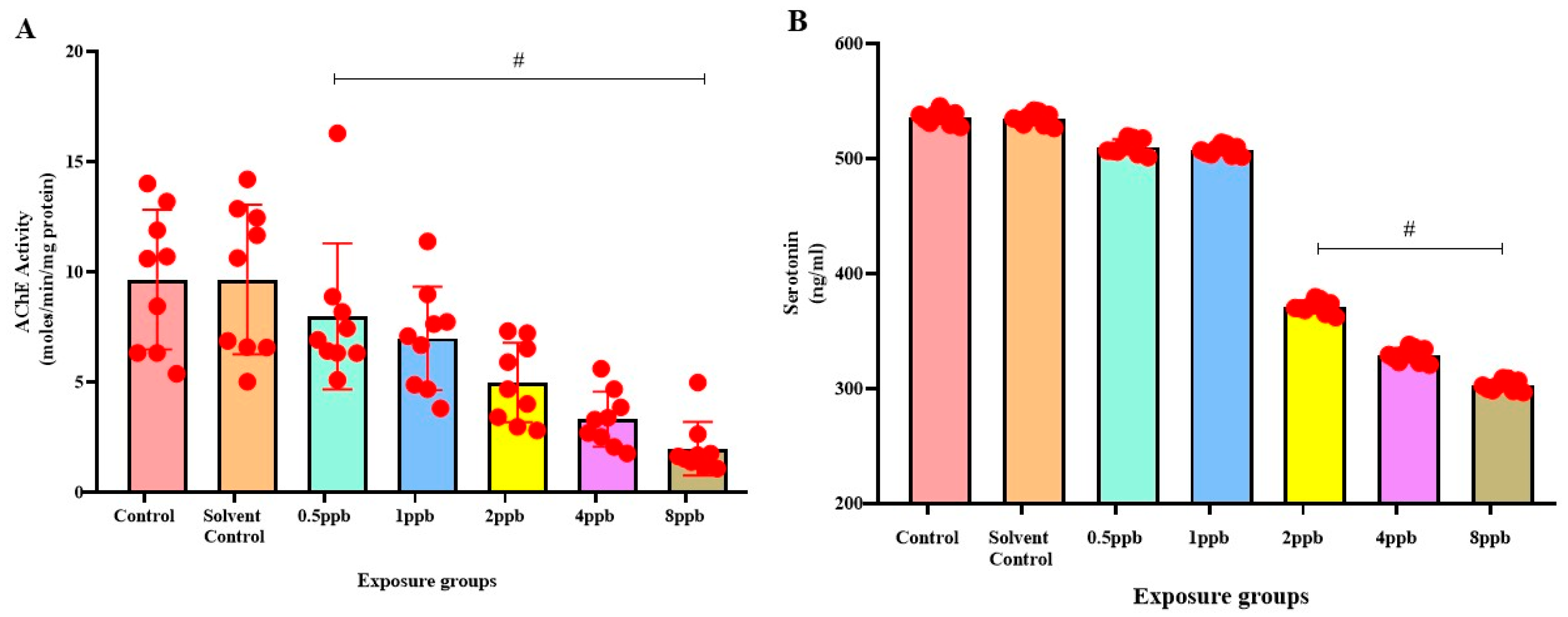
3.4. Effect of BHA on Acetylcholinesterase Activity in Zebrafish Larvae
3.5. Effect of BHA on Serotonin Levels in Zebrafish Larvae
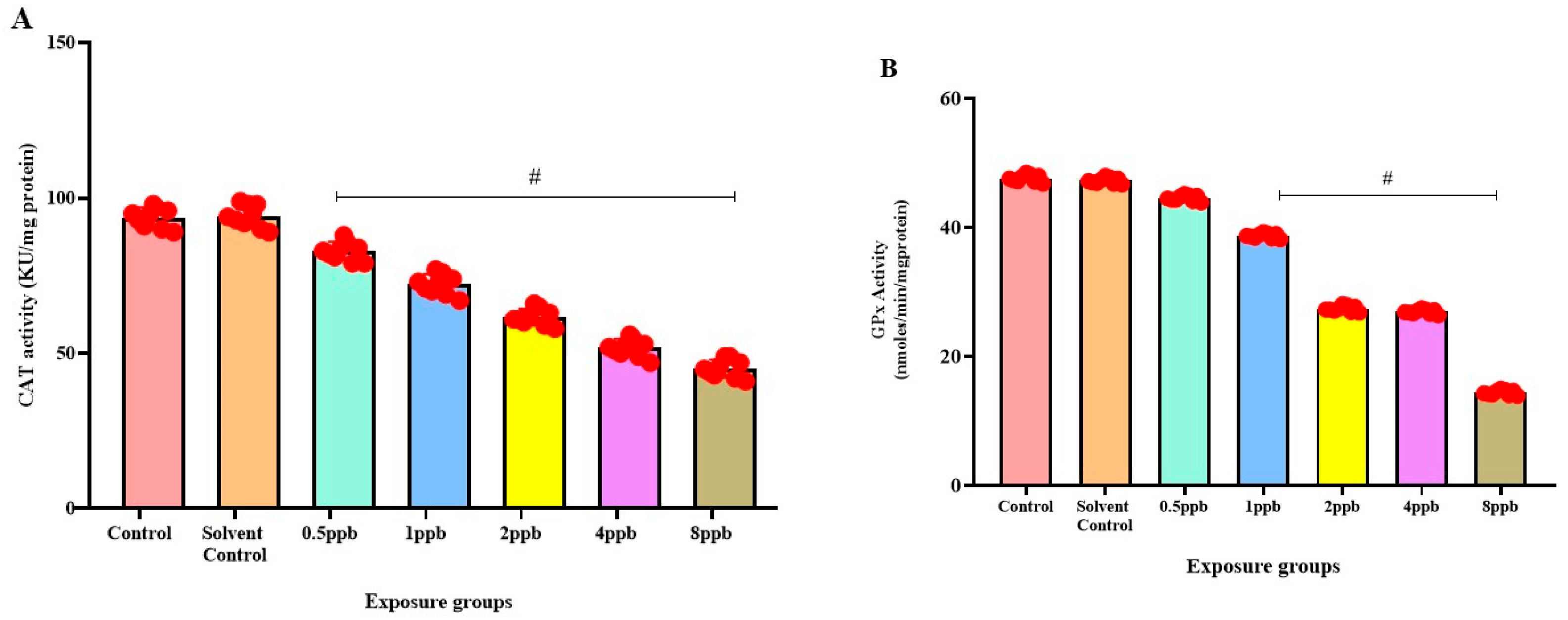
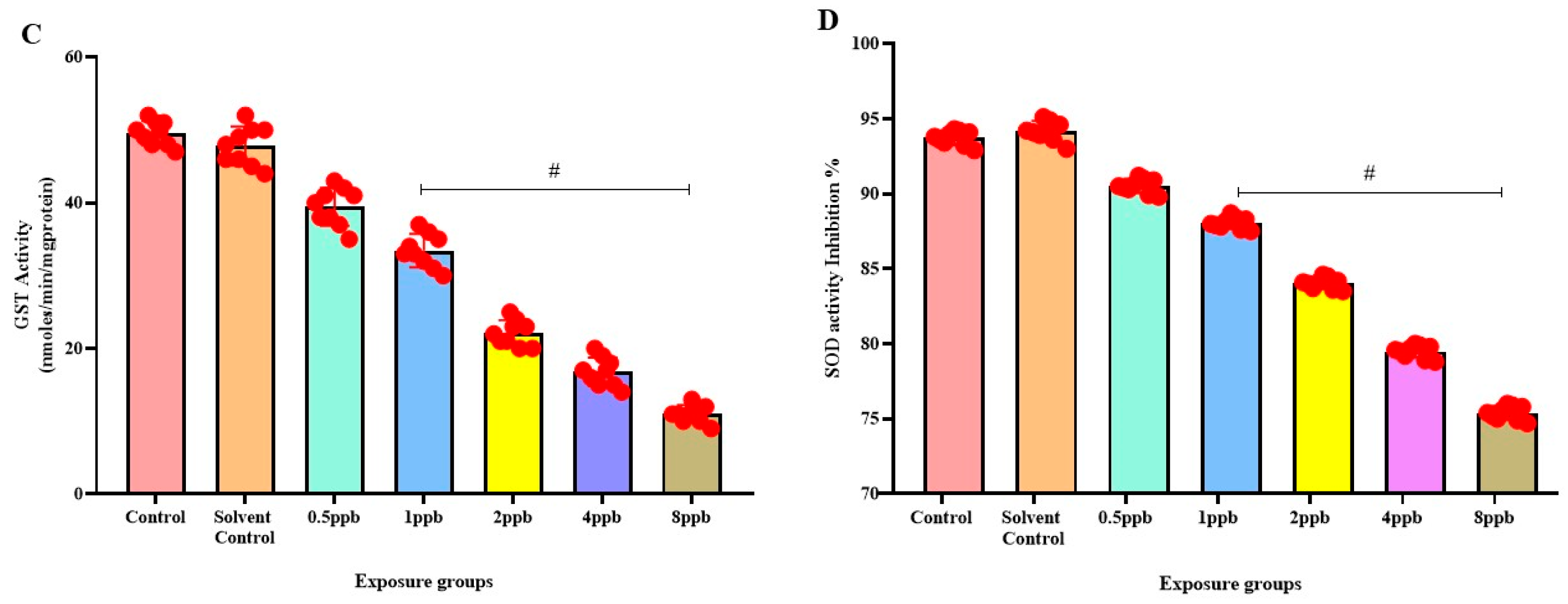
3.6. Effect of BHA Exposure on Antioxidant Enzymes in Zebrafish Larvae
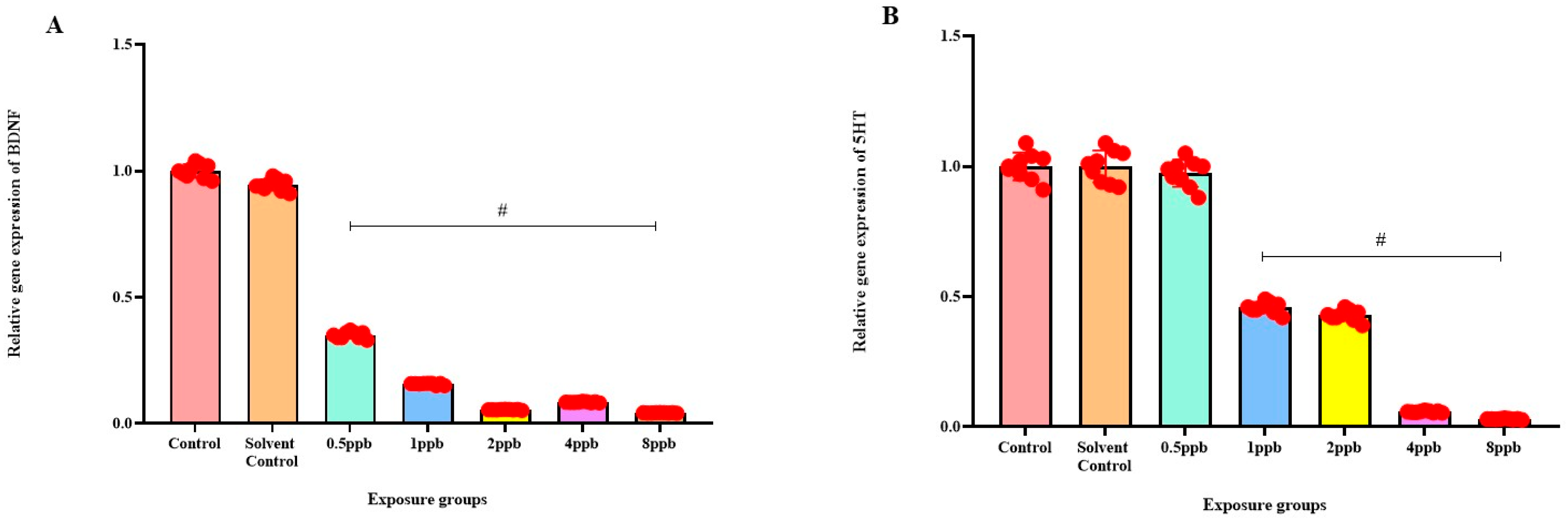
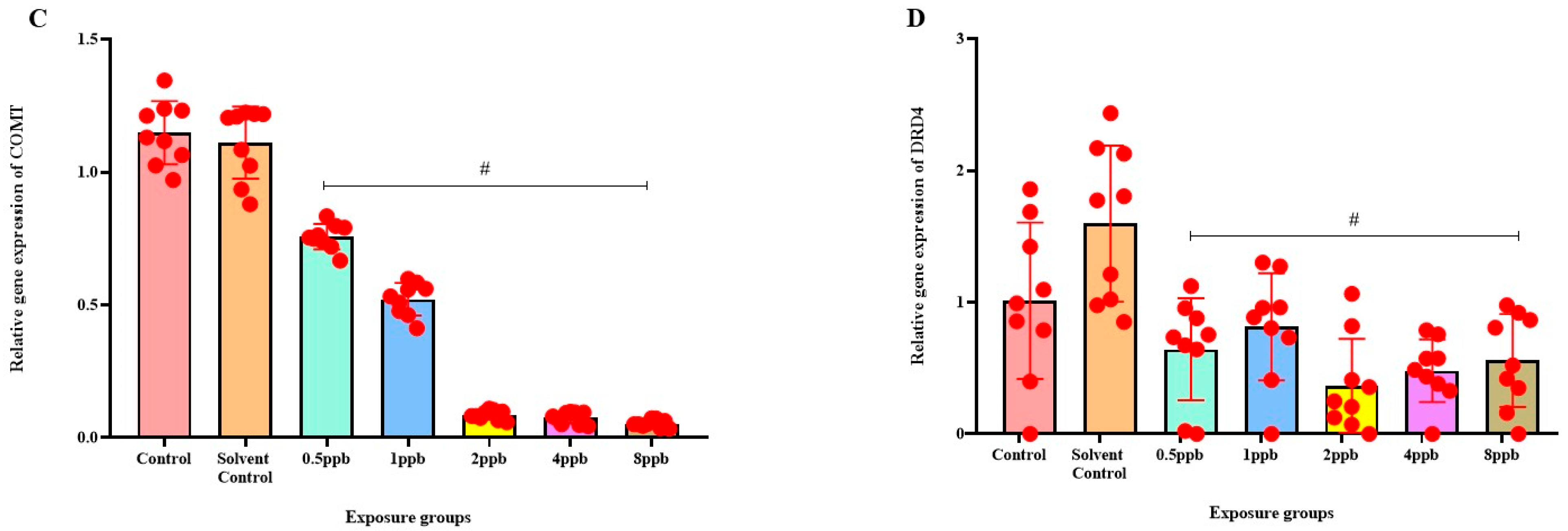
3.7. Effect of BHA on the Expression of ADHD-Related Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ham, J.; Lim, W.; You, S.; Song, G. Butylated hydroxyanisole induces testicular dysfunction in mouse testis cells by dysregulating calcium homeostasis and stimulating endoplasmic reticulum stress. Sci. Total Environ. 2020, 702, 134775. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, A.; Hu, S.; Ares, I.; Martínez-Larrañaga, M.R.; Wang, X.; Anadón, A.; Martínez, M.A. Synthetic phenolic antioxidants: Metabolism, hazards and mechanism of action. Food Chem. 2021, 353, 129488. [Google Scholar] [CrossRef] [PubMed]
- Felter, S.P.; Zhang, X.; Thompson, C. Butylated hydroxyanisole: Carcinogenic food additive to be avoided or harmless antioxidant important to protect food supply? Regul. Toxicol. Pharmacol. 2021, 121, 104887. [Google Scholar] [PubMed]
- Zhang, X.J.; Diao, M.N.; Zhang, Y.F. A review of the occurrence, metabolites and health risks of butylated hydroxyanisole (BHA). J. Sci. Food Agric. 2023, 103, 6150–6166. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages, and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Sciuto, A.M.; Moran, T.S. BHA diet enhances the survival of mice exposed to phosgene: The effect of BHA on glutathione levels in the lung. Inhal. Toxicol. 1999, 11, 855–871. [Google Scholar] [CrossRef]
- Pop, A.; Kiss, B.; Loghin, F. Endocrine disrupting effects of butylated hydroxyanisole (BHA-E320). Clujul Med. 2013, 86, 1. [Google Scholar]
- Adefegha, S.A.; Okeke, B.M.; Oboh, G. Antioxidant properties of eugenol, butylated hydroxylanisole, and butylated hydroxyl toluene with key biomolecules relevant to Alzheimer’s diseases—In vitro. J. Food Biochem. 2021, 45, e13276. [Google Scholar] [CrossRef]
- Yang, X.; Sun, Z.; Wang, W.; Zhou, Q.; Shi, G.; Wei, F.; Jiang, G. Developmental toxicity of synthetic phenolic antioxidants to the early life stage of zebrafish. Sci. Total Environ. 2018, 643, 559–568. [Google Scholar] [CrossRef]
- Wang, W.; Xiong, P.; Zhang, H.; Zhu, Q.; Liao, C.; Jiang, G. Analysis, occurrence, toxicity and environmental health risks of synthetic phenolic antioxidants: A review. Environ. Res. 2021, 201, 111531. [Google Scholar] [CrossRef]
- Park, M.W.; Cha, H.W.; Kim, J.; Kim, J.H.; Yang, H.; Yoon, S.; Boonpraman, N.; Yi, S.S.; Yoo, I.D.; Moon, J.S. NOX4 promotes ferroptosis of astrocytes by oxidative stress-induced lipid peroxidation via the impairment of mitochondrial metabolism in Alzheimer’s diseases. Redox Biol. 2021, 41, 101947. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, W.; Li, F.; Wang, X.; Pan, X.; Liu, Y.; Wang, L.; Sun, W.; Li, F.; Jiang, S. Dissecting the long-term neurobehavioral impact of embryonic benz[a]anthracene exposure on zebrafish: Social dysfunction and molecular pathway activation. Sci. Total Environ. 2024, 930, 172615. [Google Scholar] [CrossRef] [PubMed]
- Oades, R.D. Role of the Serotonin System in ADHD: Treatment Implications. Expert Rev. Neurother. 2007, 7, 1357–1374. [Google Scholar] [CrossRef]
- Baran, A.; Kokturk, M.; Atamanalp, M.; Ceyhun, S.B. Determination of developmental toxicity of zebrafish exposed to propyl gallate dosed lower than ADI (acceptable daily intake). Regul. Toxicol. Pharmacol. 2018, 94, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Reuter, M.; Kirsch, P.; Hennig, J. Inferring candidate genes for Attention Deficit Hyperactivity Disorder (ADHD) assessed by the World Health Organization Adult ADHD Self-Report Scale (ASRS). J. Neural Transm. 2006, 113, 929–938. [Google Scholar] [CrossRef]
- Dambal, V.Y.; Selvan, K.P.; Lite, C.; Barathi, S.; Santosh, W. Developmental toxicity and induction of vitellogenin in embryo-larval stages of zebrafish (Danio rerio) exposed to methyl Paraben. Ecotoxicol. Environ. Saf. 2017, 141, 113–118. [Google Scholar]
- Raja, G.L.; Subhashree, K.D.; Lite, C.; Santosh, W.; Barathi, S. Transient exposure of methylparaben to zebrafish (Danio rerio) embryos altered cortisol level, acetylcholinesterase activity and induced anxiety-like behaviour. Gen. Comp. Endocrinol. 2019, 279, 53–59. [Google Scholar] [CrossRef]
- Tshering, G.; Plengsuriyakarn, T.; Na-Bangchang, K.; Pimtong, W. Embryotoxicity evaluation of atractylodin and β-eudesmol using the zebrafish model. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 239, 108869. [Google Scholar] [CrossRef]
- Zhu, X.; Zhu, L.; Li, Y.; Duan, Z.; Chen, W.; Alvarez, P.J. Developmental toxicity in zebrafish (Danio rerio) embryos after exposure to manufactured nanomaterials: Buckminsterfullerene aggregates (nC60) and fullerol. Environ. Toxicol. Chem. 2007, 26, 976–979. [Google Scholar] [CrossRef]
- Parng, C.; Seng, W.L.; Semino, C.; McGrath, P. Zebrafish: A preclinical model for drug screening. Assay Drug Dev. Technol. 2002, 1, 41–48. [Google Scholar] [CrossRef]
- Maximino, C.; Benzecry, R.; Oliveira KR, M.; Batista ED, J.O.; Herculano, A.M.; Rosemberg, D.B.; de Oliveira, D.L.; Blaser, R. A comparison of the light/dark and novel tank tests in zebrafish. Behaviour 2012, 149, 1099–1123. [Google Scholar] [CrossRef]
- Bruzzone, M.; Gatto, E.; Xiccato, T.L.; Dalla Valle, L.; Fontana, C.M.; Meneghetti, G.; Bisazza, A. Measuring recognition memory in zebrafish larvae: Issues and limitations. PeerJ 2020, 8, e8890. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, I.; Vasantharekha, R.; Seetharaman, B. Gut dysbiosis, anxiety and depression and the neurotoxic effects of formulated favipiravir and the restorative impact of vitamin D supplementation in adult zebrafish. J. Food Sci. Technol. 2025. [Google Scholar] [CrossRef]
- Kuppuswamy, J.M.; Seetharaman, B. Monocrotophos Based Pesticide Alters the Behavior Response Associated with Oxidative Indices and Transcription of Genes Related to Apoptosis in Adult Zebrafish (Danio rerio) Brain. Biomed. Pharmacol. J. 2020, 13, 1291–1304. [Google Scholar] [CrossRef]
- Peterson, S.M.; Freeman, J.L. RNA isolation from embryonic zebrafish and cDNA synthesis for gene expression analysis. J. Vis. Exp. 2009, 30, 1470. [Google Scholar] [CrossRef]
- Sourbron, J.; Schneider, H.; Kecskés, A.; Liu, Y.; Buening, E.M.; Lagae, L.; Smolders, I.; de Witte, P. Serotonergic modulation as effective treatment for Dravet syndrome in a zebrafish mutant model. ACS Chem. Chem. Neurosci. 2016, 7, 588–598. [Google Scholar] [CrossRef]
- Pavlidis, M.; Sundvik, M.; Chen, Y.C.; Panula, P. Adaptive changes in zebrafish brain in dominant–subordinate behavioral context. Behav. Brain Brain Res. 2011, 225, 529–537. [Google Scholar] [CrossRef]
- Boehmler, W.; Carr, T.; Thisse, C.; Thisse, B.; Canfield, V.A.; Levenson, R. D4 Dopamine receptor genes of zebrafish and effects of the antipsychotic clozapine on larval swimming behaviour. Genes Brain Behav. 2007, 6, 155–166. [Google Scholar] [CrossRef]
- De Felice, E.; Porreca, I.; Alleva, E.; De Girolamo, P.; Ambrosino, C.; Ciriaco, E.; Germanà, A.; Sordino, P. Localization of BDNF expression in the developing brain of zebrafish. J. Anat. 2014, 224, 564–574. [Google Scholar] [CrossRef]
- Verhagen, H.; Schilderman, P.A.; Kleinjans, J.C. Butylated hydroxyanisole in perspective. Chem. Biol. Interact. 1991, 80, 109–134. [Google Scholar] [CrossRef]
- Bailey, J.; Oliveri, A.; Levin, E.D. Zebrafish model systems for developmental neurobehavioral toxicology. Birth Defects Res. Part C Embryo Today Rev. 2013, 99, 14–23. [Google Scholar] [CrossRef]
- Lin, C.C.; Hui, M.N.; Cheng, S.H. Toxicity and cardiac effects of carbaryl in early developing zebrafish (Danio rerio) embryos. Toxicol. Appl. Pharmacol. 2007, 222, 159–168. [Google Scholar] [CrossRef]
- Steenbergen, P.J.; Richardson, M.K.; Champagne, D.L. The use of the zebrafish model in stress research. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 1432–1451. [Google Scholar] [CrossRef]
- Magyary, I. Floating novel object recognition in adult zebrafish: A pilot study. Cogn Process 2019, 20, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Win-Shwe, T.T.; Fujimaki, H.; Fujitani, Y.; Hirano, S. Novel object recognition ability in female mice following exposure to nanoparticle-rich diesel exhaust. Toxicol. Appl. Pharmacol. 2012, 262, 355–362. [Google Scholar] [CrossRef]
- Kienle, C.; Köhler, H.R.; Gerhardt, A. Behavioural and developmental toxicity of chlorpyrifos and nickel chloride to zebrafish (Danio rerio) embryos and larvae. Ecotoxicol. Environ. Saf. 2009, 72, 1740–1747. [Google Scholar] [CrossRef]
- Tsim, K.; Soreq, H. Acetylcholinesterase: Old questions and new developments. Front. Mol. Neurosci. 2013, 5, 101. [Google Scholar] [CrossRef]
- Vázquez, D.M.; Neal, C.R., Jr.; Patel, P.D.; Kaciroti, N.; López, J.F. Regulation of corticoid and serotonin receptor brain system following early life exposure of glucocorticoids: Long term implications for the neurobiology of mood. Psychoneuroendocrinology 2012, 37, 421–437. [Google Scholar] [CrossRef]
- Krain, A.L.; Castellanos, F.X. Brain development and ADHD. Clin. Psychol. Rev. 2006, 26, 433–444. [Google Scholar] [CrossRef]
- Kaźmierczak-Barańska, J.; Boguszewska, K.; Adamus-Grabicka, A.; Karwowski, B.T. Two faces of vitamin C—Antioxidative and pro-oxidative agent. Nutrients 2020, 12, 1501. [Google Scholar] [CrossRef]
- Kathore, V.; Shete, A.N.; Zingade, U.S.; Bansode, D.G. The effect of vitamin C on serum superoxide dismutase and blood sugar levels in the patients of type 2 diabetes mellitus. Int. J. Health Sci. 2014, 4, 94–100. [Google Scholar]
- Massarsky, A.; Kozal, J.S.; Di Giulio, R.T. Glutathione and zebrafish: Old assays to address a current issue. Chemosphere 2017, 168, 707–715. [Google Scholar] [CrossRef]
- Brennan, A.R.; Arnsten, A.F.T. Neuronal mechanisms underlying attention deficit hyperactivity disorder: The influence of arousal on prefrontal cortical function. Ann. N. Y. Acad. Sci. 2008, 1129, 236–245. [Google Scholar] [CrossRef]
- Banerjee, E.; Nandagopal, K. Does serotonin deficit mediate susceptibility to ADHD? Neurochem. Int. 2015, 82, 52–68. [Google Scholar] [CrossRef] [PubMed]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Levy, F. What do dopamine transporter and catechol-O-methyltransferase tell us about Attention Deficit–Hyperactivity Disorder? Pharmacogenomic implications. Aust. N. Z. J. Psychiatry 2007, 41, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Yuan, F.; Shen, X.; Xiong, G.; Wu, J. Role of COMT in ADHD: A Systematic Meta-Analysis. Mol. Neurobiol. 2014, 49, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Bhat, A.A.; Hashem, S.; Nisar, S.; Kamal, M.; Syed, N.; Temanni, M.-R.; Gupta, R.K.; Kamran, S.; Azeem, M.W.; et al. Genetic variations influence brain changes in patients with attention-deficit hyperactivity disorder. Transl Psychiatry 2021, 11, 349. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer (5′–3′) | Gene ID | Reference |
|---|---|---|---|
| 5-HTR1aa | Forward Primer: ATCTCTCTAGACGTGCTGTGCTGC Reverse Primer: GTTTTCCTTATGCGAAACCTCGCC | 100001828 | [26] |
| COMT | Forward Primer: ACTCGACCACAGCGTCTGCT Reverse Primer: AGCCCATTCGCGGTGTCTGC | 565370 | [27] |
| DRD4a | Forward Primer: ATGGTAGAGGCAGACATGCCA Reverse Primer: TTAGCATGCTCAGGCTAGCAG | 503564 | [28] |
| BDNF | Forward Primer: ATAGTAACGAACAGGATGG Reverse Primer: GCTCAGTCATGGGAGTCC | 58118 | [29] |
| Exposure Groups | 24 hpf | 48 hpf | 72 hpf | 96 hpf | |||
|---|---|---|---|---|---|---|---|
| BS | YE | BS | PE | YE | |||
| Control | N/O | N/O | N/O | N/O | N/O | N/O | N/O |
| Solvent Control | N/O | N/O | N/O | N/O | N/O | N/O | N/O |
| 0.5 ppb BHA | N/O | N/O | N/O | N/O | N/O | 15% | N/O |
| 1 ppb BHA | N/O | N/O | N/O | N/O | 20% | N/O | N/O |
| 2 ppb BHA | N/O | N/O | N/O | 25% | 15% | N/O | 25% |
| 4 ppb BHA | N/O | N/O | N/O | 30% | 45% | 15% | 30% |
| 8 ppb BHA | N/O | N/O | N/O | 35% | 50% | N/O | 40% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veshaal, K.; Vasantharekha, R.; Balu, U.R.; Aayush, M.V.; Pillai, S.S.B.; Santosh, W.; Seetharaman, B. Butylated Hydroxyanisole (BHA) Disrupts Brain Signalling in Embryo–Larval Stage of Zebrafish Leading to Attention Deficit Hyperactivity Disorder (ADHD). J. Xenobiot. 2025, 15, 116. https://doi.org/10.3390/jox15040116
Veshaal K, Vasantharekha R, Balu UR, Aayush MV, Pillai SSB, Santosh W, Seetharaman B. Butylated Hydroxyanisole (BHA) Disrupts Brain Signalling in Embryo–Larval Stage of Zebrafish Leading to Attention Deficit Hyperactivity Disorder (ADHD). Journal of Xenobiotics. 2025; 15(4):116. https://doi.org/10.3390/jox15040116
Chicago/Turabian StyleVeshaal, Kandhasamy, Ramasamy Vasantharekha, Usha Rani Balu, Mahesh Vallabi Aayush, Saheshnu Sai Balaji Pillai, Winkins Santosh, and Barathi Seetharaman. 2025. "Butylated Hydroxyanisole (BHA) Disrupts Brain Signalling in Embryo–Larval Stage of Zebrafish Leading to Attention Deficit Hyperactivity Disorder (ADHD)" Journal of Xenobiotics 15, no. 4: 116. https://doi.org/10.3390/jox15040116
APA StyleVeshaal, K., Vasantharekha, R., Balu, U. R., Aayush, M. V., Pillai, S. S. B., Santosh, W., & Seetharaman, B. (2025). Butylated Hydroxyanisole (BHA) Disrupts Brain Signalling in Embryo–Larval Stage of Zebrafish Leading to Attention Deficit Hyperactivity Disorder (ADHD). Journal of Xenobiotics, 15(4), 116. https://doi.org/10.3390/jox15040116