Toxicity Bioassay and Cytotoxic Effects of the Benthic Marine Dinoflagellate Amphidinium operculatum



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Dinoflagellate Isolation and Maintenance
2.2. Dinoflagellate Identification
2.3. Preparation of Ethanolic Extracts of Cultured Amphidinium Cells
2.4. Acute Toxicity Artemia Assay
2.5. Dose-Response Curve
2.6. Cell Line Culture and Assays
2.7. Sulforhodamine B Assay
2.8. Statistics
3. Results and Discussion
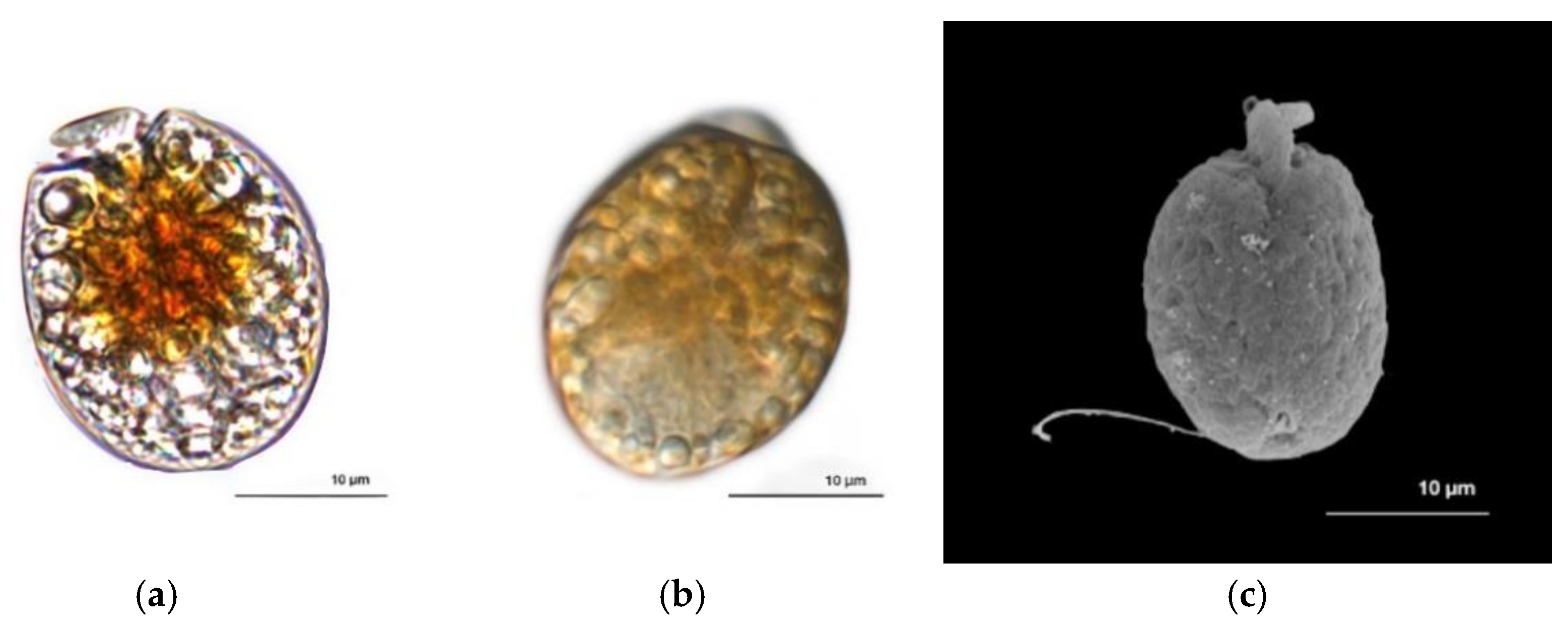
3.1. Dinoflagellate Description and Identification
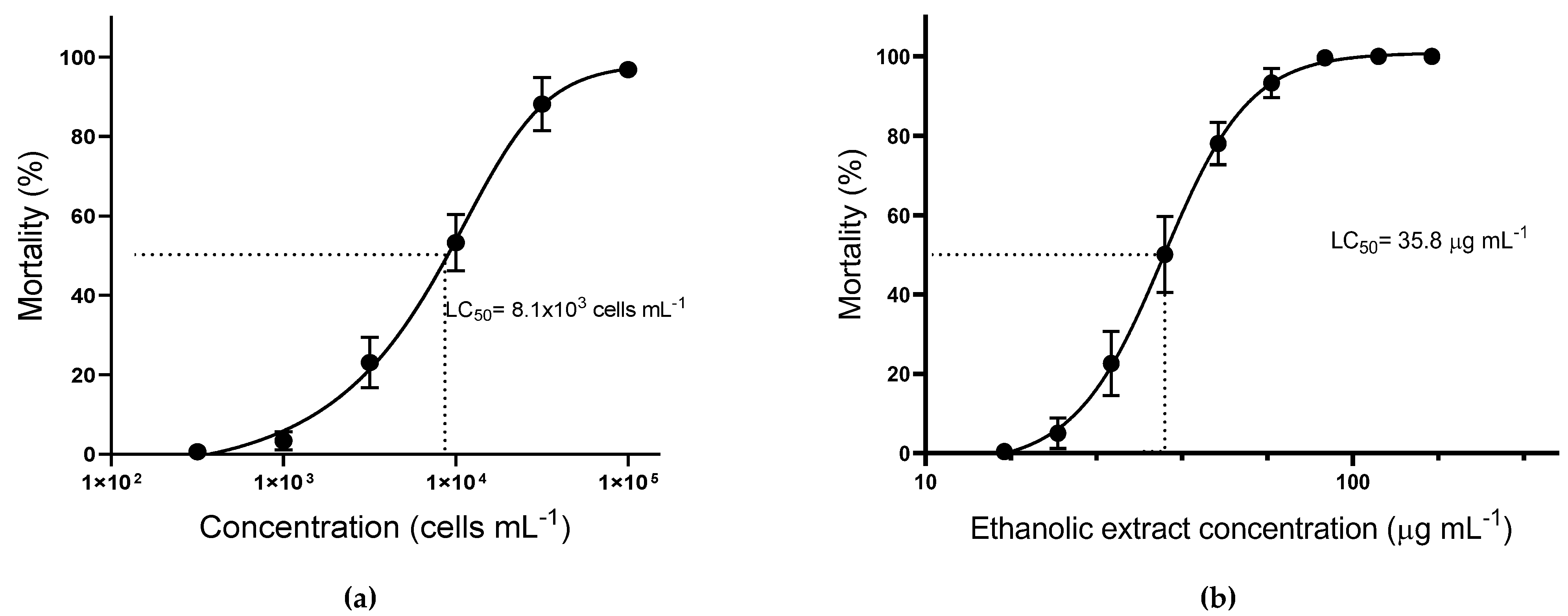
3.2. Acute Toxicity Assay
3.3. Ecotoxicological Assessment of A. operculatum
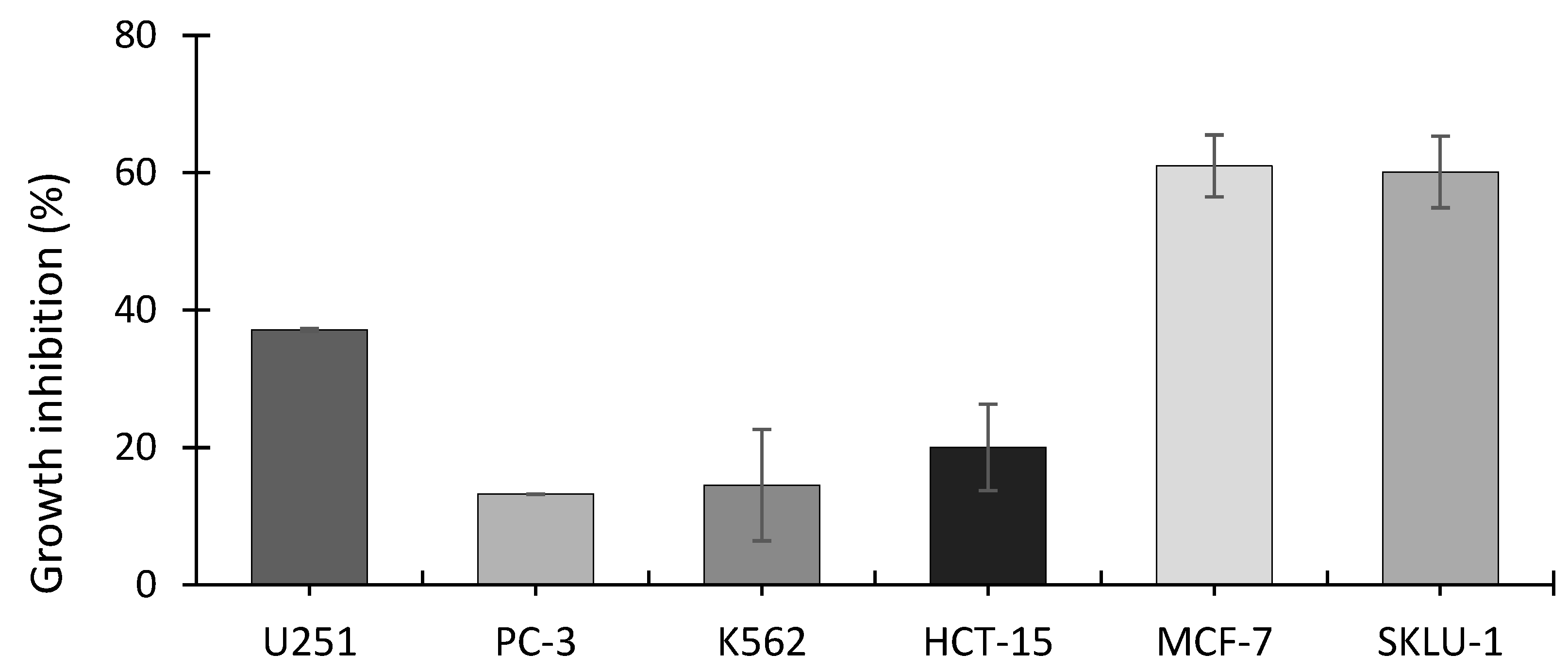
3.4. SRB Assay on Cell Lines
3.5. Cytotoxicity of A. operculatum on Cancer Cell Lines
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neves, R.A.; Fernandes, T.; Santos, L.N.D.; Nascimento, S.M. Toxicity of benthic dinoflagellates on grazing, behavior and survival of the brine shrimp Artemia salina. PLoS ONE 2017, 12, e0175168. [Google Scholar] [CrossRef]
- Durán-Riveroll, L.D.; Cembella, A.D.; Okolodkov, Y.B. A review on the biodiversity and biogeography of toxigenic benthic marine dinoflagellates of the coasts of Latin America. Front. Mar. Sci. 2019, 6, 148. [Google Scholar] [CrossRef]
- Cembella, A.D. Chemical ecology of eukaryotic microalgae in marine ecosystems. Phycologia 2003, 42, 420–447. [Google Scholar] [CrossRef]
- Martínez-Andrade, K.A.; Lauritano, C.; Romano, G.; Ianora, A. Marine microalgae with anti-cancer properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [PubMed]
- Ben-Gharbia, H.; Yahia, O.K.-D.; Amzil, Z.; Chomérat, N.; Abadie, E.; Masseret, E.; Sibat, M.; Zmerli Triki, H.; Nouri, H.; Laabir, M. Toxicity and growth assessments of three thermophilic benthic dinoflagellates (Ostreopsis cf. ovata, Prorocentrum lima and Coolia monotis) developing in the Southern Mediterranean basin. Toxins 2016, 8, 297. [Google Scholar] [CrossRef] [PubMed]
- Pagliara, P.; Caroppo, C. Toxicity assessment of Amphidinium carterae, Coolia cfr. monotis and Ostreopsis cfr. ovata (Dinophyta) isolated from the northern Ionian Sea (Mediterranean Sea). Toxicon 2012, 60, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Berdalet, E.; Tester, P.A.; Chinain, M.; Fraga, S.; Lemée, R.; Litaker, W.; Penna, A.; Usup, G.; Vila, M.; Zingone, A. Harmful algal blooms in benthic systems: Recent progress and future research. Oceanography 2017, 30, 36–45. [Google Scholar] [CrossRef]
- Karafas, S.; Teng, S.T.; Leaw, C.P.; Alves-de-Souza, C. An evaluation of the genus Amphidinium (Dinophyceae) combining evidence from morphology, phylogenetics, and toxin production, with the introduction of six novel species. Harmful Algae 2017, 68, 128–151. [Google Scholar] [CrossRef]
- Murray, S.A.; Garby, T.; Hoppenrath, M.; Neilan, B.A. Genetic diversity, morphological uniformity and polyketide production in dinoflagellates (Amphidinium, Dinoflagellata). PLoS ONE 2012, 7, e38253. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Seino, N.; Murakami, Y.; Murata, M. Toxins produced by benthic dinoflagellates. Biol. Bull. 1987, 172, 128–131. [Google Scholar] [CrossRef]
- Shah, M.M.R.; Samarakoon, K.W.; Ko, J.; Lakmal, H.C.; Lee, J.; An, S.; Jeon, Y. Potentiality of benthic dinoflagellate cultures and screening of their bioactivities in Jeju Island. Afr. J. Biotechnol. 2014, 13, 792–805. [Google Scholar]
- Lauritano, C.; De Luca, D.; Ferrarini, A.; Avanzato, C.; Minio, A.; Esposito, F.; Ianora, A. De novo transcriptome of the cosmopolitan dinoflagellate Amphidinium carterae to identify enzymes with biotechnological potential. Sci Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sampayo, M.A. Encystment and excystment of a Portugese isolate of Amphidinium carterae in cultures. In Toxic Dinoflagellates; Anderson, D.M., White, A.W., Baden, D.G., Eds.; Elsevier: Amsterdam, The, Netherlands, 1985; pp. 125–130. [Google Scholar]
- Murray, S.A.; Kohli, G.S.; Farrell, H.; Spiers, Z.B.; Place, A.R.; Dorantes-Aranda, J.J.; Ruszczyk, J. A fish kill associated with a bloom of Amphidinium carterae in a coastal lagoon in Sydney, Australia. Harmful Algae 2015, 49, 19–28. [Google Scholar] [CrossRef]
- Kobayashi, J.; Yamaguchi, N.; Ishibashi, H. Amphidinin-A, a novel Amphidinolide-related metabolite from the cultured marine dinoflagellate Amphidinium sp. Tetrahedron Lett. 1994, 35, 7049–7050. [Google Scholar] [CrossRef]
- Kubota, T.; Endo, T.; Takahashi, Y.; Tsuda, M.; Kobayashi, J. Amphidinin B, a new polyketide metabolite from marine dinoflagellate Amphidinium sp. J. Antibiot. 2006, 59, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Bauer, I.; Maranda, L.; Young, K.A.; Shimizu, Y.; Huang, S. The isolation and structures of unusual 1, 4-polyketides from the dinoflagellate, Amphidinium sp. Tetrahedron Lett. 1995, 36, 991–994. [Google Scholar] [CrossRef]
- Kobayashi, J.; Tsuda, M. Amphidinolides, bioactive macrolides from symbiotic marine dinoflagellates. Nat. Prod. Rep. 2004, 21, 77–93. [Google Scholar] [CrossRef]
- Kobayashi, J. Amphidinolides and its related macrolides from marine dinoflagellates. J. Antibiot. 2008, 61, 271–284. [Google Scholar] [CrossRef]
- Takahashi, Y.; Kubota, T.; Kobayashi, J. Amphidinolactone A, a new 13-memebered macrolide from dinoflagellate Amphidinium sp. Heterocycles 2007, 72, 567–572. [Google Scholar]
- Takahashi, Y.; Kubota, T.; Kobayashi, J. Amphidinolactone B, a new 26-membered macrolide from dinoflagellate Amphidinium sp. J. Antibiot. 2007, 60, 376–379. [Google Scholar] [CrossRef]
- Bauer, I.; Maranda, L.; Young, K.A.; Shimizu, Y.; Fairchild, C.; Cornell, L.; MacBeth, J.; Huang, S. Isolation and structure of caribenolide I, a highly potent antitumor macrolide from a cultured free-swimming caribbean dinoflagellate, Amphidinium sp. S1-36-5. J. Org. Chem. 1995, 60, 1084–1086. [Google Scholar] [CrossRef]
- Tsuda, M.; Oguchi, K.; Iwamoto, R.; Okamoto, Y.; Kobayashi, J.I.; Fukushi, E.; Kawabata, J.; Ozawa, T.; Masuda, A.; Kitaya, Y. Iriomoteolide-1a, a potent cytotoxic 20-membered macrolide from a benthic dinoflagellate Amphidinium species. J. Org. Chem. 2007, 72, 4469–4474. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.; Oguchi, K.; Iwamoto, R.; Okamoto, Y.; Fukushi, E.; Kawabata, J.; Ozawa, T.; Masuda, A. Iriomoteolides-1b and-1c, 20-membered macrolides from a marine dinoflagellate Amphidinium species. J. Nat. Prod. 2007, 70, 1661–1663. [Google Scholar] [CrossRef]
- Oguchi, K.; Fukushi, E.; Tsuda, M. Iriomoteolide-4a, a new 16-membered macrolide from dinoflagellate Amphidinium species. Planta Med. 2008, 74, 1041. [Google Scholar] [CrossRef]
- Espiritu, R.A.; Tan, M.C.S.; Oyong, G.G. Evaluation of the anti-cancer potential of amphidinol 2, a polyketide metabolite from the marine dinoflagellate Amphidinium klebsii. Jordan J. Biol. Sci. 2017, 10, 297–302. [Google Scholar]
- Wellkamp, M.; García-Camacho, F.; Durán-Riveroll, L.M.; Tebben, J.; Tillmann, U.; Krock, B. LC-MS/MS method development for the discovery and identification of amphidinols produced by Amphidinium. Mar. Drugs 2020, 18, 497. [Google Scholar] [CrossRef]
- Nunes, B.S.; Carvalho, F.D.; Guilhermino, L.M.; Van Stappen, G. Use of the genus Artemia in ecotoxicity testing. Environ. Pollut. 2006, 144, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Novais Oliveira, T.M.; Vaz, C. Marine toxicology: Assays and perspectives for developing countries. In Bioassays: Advanced Methods and Applications; Häder, D.P., Erzinger, G.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 387–401. [Google Scholar]
- Khalifa, S.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine natural products: A source of novel anticancer drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef]
- Giubergia, S.; Schleissner, C.; de la Calle, F.; Pretsch, A.; Pretsch, D.; Gram, L.; Schmidt Thogersen, M. Screening microorganisms for bioactive compounds. In The Marine Microbiome: An Untapped Source of Biodiversity and Biotechnological Potential; Stal, L.J., Cretoiu, M.S., Eds.; Springer: Cham, Switzerland, 2016; pp. 345–376. [Google Scholar]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Lee, K.H.; Jeong, H.J.; Park, K.; Kang, N.S.; Yoo, Y.D.; Lee, M.J.; Lee, J.; Lee, S.; Kim, T.; Kim, H.S.; et al. Morphology and molecular characterization of the epiphytic dinoflagellate Amphidinium massartii, isolated from the temperate waters off Jeju Island, Korea. Algae 2013, 28, 213–231. [Google Scholar] [CrossRef]
- Aquino-Cruz, A.; Okolodkov, Y. Impact of increasing water temperature on growth, photosynthetic efficiency, nutrient consumption, and potential toxicity of Amphidinium cf. carterae and Coolia monotis (Dinoflagellata). Rev. Biol. Mar. Oceanogr. 2016, 51, 565–580. [Google Scholar] [CrossRef]
- Blackburn, S.I.; Bolch, C.J.S.; Haskard, K.A.; Hallegraeff, G.M. Reproductive compatibility among four global populations of the toxic dinoflagellate Gymnodinium catenatum (Dinophyceae). Phycologia 2001, 40, 78–87. [Google Scholar] [CrossRef]
- Markham, J.; Hagmeier, E. Observations on the effects of germanium dioxide on the growth of macro-algae and diatoms. Phycologia 1982, 21, 125–130. [Google Scholar] [CrossRef]
- Pérez-López, H.; Durán-Riveroll, L.; Gómez-Lizárraga, L.E.; Mendoza-Garfias, M.B. Simple method for preparing delicate dinoflagellate of the genus Amphidinium for scanning electron microscopy. Microsc Microanal 2020, 26 (Suppl. S2), 1366–1369. [Google Scholar] [CrossRef]
- Murray, S.; Flø Jørgensen, M.; Daugbjerg, N.; Rhodes, L. Amphidinium revisited. II. Resolving species boundaries in the Amphidinium operculatum species complex (Dinophyceae), including the descriptions of Amphidinium trulla sp. nov. and Amphidinium gibbosum. comb. nov. J. Phycol. 2004, 40, 366–382. [Google Scholar] [CrossRef]
- Jørgensen, M.F.; Murray, S.; Daugbjerg, N. Amphidinium revisited. I. Redefinition of Amphidinium (Dinophyceae) based on cladistic and molecular phylogenetic analyses. J. Phycol. 2004, 40, 351–365. [Google Scholar] [CrossRef]
- Scholin, C.A.; Herzog, M.; Sogin, M.; Anderson, D.M. Identification of group and strain specific genetic markers of globally distributed Alexandrium (Dinophyceae). II. Sequence analysis of a fragment of the LSU rRNA gene 1. J. Phycol. 1994, 30, 999–1011. [Google Scholar] [CrossRef]
- Daugbjerg, N.; Hansen, G.; Larsen, J.; Moestrup, Ø. Phylogeny of some of the major genera of dinoflagellates based on ultrastructure and partial LSU rDNA sequence data, including the erection of three new genera of unarmoured dinoflagellates. Phycologia 2000, 39, 302–317. [Google Scholar] [CrossRef]
- European Chemicals Agency. Toxicity to Aquatic Algae and Cyanobacteria. Available online: https://echa.europa.eu/registration-dossier/-/registered-dossier/26915/6/2/6 (accessed on 1 March 2021).
- Arulvasu, C.; Jennifer, S.M.; Prabhu, D.; Chandhirasekar, D. Toxicity effect of silver nanoparticles in brine shrimp Artemia. Sci. World J. 2014, 2014, 256919. [Google Scholar] [CrossRef] [PubMed]
- Monks, A.; Scudiero, D.; Skehan, P.; Shoemaker, R.; Paul, K.; Vistica, D.; Hose, C.; Langley, J.; Cronise, P.; Vaigro-Wolff, A.; et al. Feasibility of a high-flux anticancer drug screen using a diverse panel of cultured human tumor cell lines. J. Natl Cancer I 1991, 38, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Hoppenrath, M.; Murray, S.A.; Chomérat, N.; Horiguchi, T. Marine Benthic Dinoflagellates-Unveiling Their Worldwide Biodiversity; Schweizerbart’sche Verlagsbuchhandlung: Stuttgart, Germany, 2014; p. 276. [Google Scholar]
- Moreira-González, A.R.; Fernandes, L.F.; Uchida, H.; Uesugi, A.; Suzuki, T.; Chomérat, N.; Bilien, G.; Pereira, T.A.; Mafra Jr, L.L. Morphology, growth, toxin production, and toxicity of cultured marine benthic dinoflagellates from Brazil and Cuba. J. Appl. Phycol. 2019, 31, 3699–3719. [Google Scholar] [CrossRef]
- Delattre, C.; Pierre, G.; Laroche, C.; Michaud, P. Production, extraction and characterization of microalgal and cyanobacterial exopolysaccharides. Biotechnol. Adv. 2016, 34, 1159–1179. [Google Scholar] [CrossRef]
- Privitera, D.; Giussani, V.; Isola, G.; Faimali, M.; Piazza, V.; Garaventa, F.; Asnaghi, V.; Cantamessa, E.; Cattaneo-Vietti, R.; Chiantore, M. Toxic effects of Ostreopsis ovata on larvae and juveniles of Paracentrotus lividus. Harmful Algae 2012, 18, 16–23. [Google Scholar] [CrossRef]
- Thornton, D. Diatom aggregation in the sea: Mechanisms and ecological implications. Eur. J. Phycol. 2002, 37, 149–161. [Google Scholar] [CrossRef]
- Moreira, A.; Rodríguez, F.; Riobó, P.; Franco, J.M.; Martínez, N.; Chamero, D.; Alonso, C. Notes on Ostreopsis sp. from southern-central coast of Cuba. Cryptogamie Algol 2012, 33, 217–225. [Google Scholar] [CrossRef]
- Vanhaecke, P.; Sorgeloos, P. International Study on Artemia. IV. The biometrics of Artemia strains from different geographical origin. In The Brine Shrimp Artemia. Ecology, Culturing, Use in Aquaculture; Persoone, G., Sorgeloos, P., Roels, O., Jaspers, E., Eds.; Universa Press: Wetteren, Belgian, 1980; Volume 3, pp. 393–405. [Google Scholar]
- Van Stappen, G. Introduction, biology and ecology of Artemia. In Manual on the Production and Use of Live Food for Aquaculture; Lavens, P., Sorgeloos, P., Eds.; FAO Fisheries Technical Paper: Rome, Italy, 1996; pp. 79–105. [Google Scholar]
- Reynolds, C.S. Variability in the provision and function of mucilage in phytoplankton: Facultative responses to the environment. Hydrobiologia 2007, 578, 37–45. [Google Scholar] [CrossRef]
- Giussani, V.; Sbrana, F.; Asnaghi, V.; Vassalli, M.; Faimali, M.; Casabianca, S.; Penna, A.; Ciminiello, P.; Dell’Aversano, C.; Tartaglione, L.; et al. Active role of the mucilage in the toxicity mechanism of the harmful benthic dinoflagellate Ostreopsis cf. ovata. Harmful Algae 2015, 44, 46–53. [Google Scholar] [CrossRef]
- Delgado, M.; Alcaraz, M. Interactions between red tide microalgae and herbivorous zooplankton: The noxious effects of Gyrodinium corsicum (Dinophyceae) on Acartia grani (Copepoda: Calanoida). J. Plankton Res. 1999, 21, 2361–2371. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. 2020. Available online: https://www.algaebase.org (accessed on 1 December 2020).
- Deeds, J.R.; Hoesch, R.E.; Place, A.R.; Kao, J.P. The cytotoxic mechanism of karlotoxin 2 (KmTx 2) from Karlodinium veneficum (Dinophyceae). Aquat. Toxicol. 2015, 159, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Martínez, K.A.; Lauritano, C.; Druka, D.; Romano, G.; Grohmann, T.; Jaspars, M.; Martín, J.; Díaz, C.; Cautain, B.; de la Cruz, M.; et al. Amphidinol 22, a new cytotoxic and antifungal amphidinol from the dinoflagellate Amphidinium carterae. Mar. Drugs 2019, 17, 385. [Google Scholar] [CrossRef]
- Samarakoon, K.W.; Ko, J.Y.; Shah, M.M.R.; Lee, J.H.; Kang, M.C.; O-Nam, K.; Lee, J.B.; Jeon, Y.J. In vitro studies of anti-inflammatory and anticancer activities of organic solvent extracts from cultured marine microalgae. Algae 2013, 28, 111–119. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mejía-Camacho, A.L.; Durán-Riveroll, L.M.; Cembella, A.D. Toxicity Bioassay and Cytotoxic Effects of the Benthic Marine Dinoflagellate Amphidinium operculatum. J. Xenobiot. 2021, 11, 33-45. https://doi.org/10.3390/jox11020003
Mejía-Camacho AL, Durán-Riveroll LM, Cembella AD. Toxicity Bioassay and Cytotoxic Effects of the Benthic Marine Dinoflagellate Amphidinium operculatum. Journal of Xenobiotics. 2021; 11(2):33-45. https://doi.org/10.3390/jox11020003
Chicago/Turabian StyleMejía-Camacho, Ana Luisa, Lorena María Durán-Riveroll, and Allan Douglas Cembella. 2021. "Toxicity Bioassay and Cytotoxic Effects of the Benthic Marine Dinoflagellate Amphidinium operculatum" Journal of Xenobiotics 11, no. 2: 33-45. https://doi.org/10.3390/jox11020003
APA StyleMejía-Camacho, A. L., Durán-Riveroll, L. M., & Cembella, A. D. (2021). Toxicity Bioassay and Cytotoxic Effects of the Benthic Marine Dinoflagellate Amphidinium operculatum. Journal of Xenobiotics, 11(2), 33-45. https://doi.org/10.3390/jox11020003