

The Influence of Nitrogen and Phosphorus Addition on Growth of the Invasive C4 Grass Saccharum spontaneum



Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Nutrient Concentration Analysis
2.4. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moran, E.F.; Brondizio, E.S.; Tucker, J.M.; da Silva-Forsberg, M.C.; McCracken, S.; Falesi, I. Effects of soil fertility and land-use on forest succession in Amazonia. For. Ecol. Manag. 2000, 139, 93–108. [Google Scholar] [CrossRef]
- Nepstad, D.C.; Uhl, C.; Serrao, E.A. Recuperation of a degraded Amazonian landscape: Forest recovery and agricultural restoration. Ambio 1991, 20, 248–255. [Google Scholar]
- D’Antonio, C.M.; Vitousek, P.M. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annu. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar] [CrossRef]
- Holl, K.D. Factors limiting tropical rain forest regeneration in abandoned pasture: Seed rain, seed germination, microclimate, and soil. Biotropica 1999, 31, 229–242. [Google Scholar] [CrossRef]
- Aide, T.M.; Cavelier, J. Barriers to lowland tropical forest restoration in the Sierra Nevada de Santa Marta, Colombia. Restor. Ecol. 1994, 2, 219–229. [Google Scholar] [CrossRef]
- Vitousek, P.M. Litterfall, nutrient cycling, and nutrient limitation in tropical forests. Ecology 1984, 65, 285–298. [Google Scholar] [CrossRef]
- Sollins, P. Factors influencing species composition in tropical lowland rain forest: Does soil matter? Ecology 1998, 79, 23–30. [Google Scholar] [CrossRef]
- Townsend, A.R.; Cleveland, C.C.; Asner, G.P.; Bustamante, M.M. Controls over foliar N: P ratios in tropical rain forests. Ecology 2007, 88, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Moran, E.; Mausel, P. Linking Amazonian secondary succession forest growth to soil properties. Land Degrad. Devel. 2002, 13, 331–343. [Google Scholar] [CrossRef]
- Newman, E. Phosphorus balance of contrasting farming systems, past and present. Can food production be sustainable? J. Appl. Ecol. 1997, 34, 1334–1347. [Google Scholar] [CrossRef]
- Lawrence, D.; D’Odorico, P.; Diekmann, L.; DeLonge, M.; Das, R.; Eaton, J. Ecological feedbacks following deforestation create the potential for a catastrophic ecosystem shift in tropical dry forest. Proc. Natl. Acad. Sci. USA 2007, 104, 20696–20701. [Google Scholar] [CrossRef] [PubMed]
- Baribault, T.W.; Kobe, R.K.; Finley, A.O. Tropical tree growth is correlated with soil phosphorus, potassium, and calcium, though not for legumes. Ecol. Monogr. 2012, 82, 189–203. [Google Scholar] [CrossRef]
- Mercado, L.M.; Patino, S.; Domingues, T.F.; Fyllas, N.M.; Weedon, G.P.; Sitch, S.; Quesada, C.A.; Phillips, O.L.; Aragão, L.E.; Malhi, Y. Variations in Amazon forest productivity correlated with foliar nutrients and modelled rates of photosynthetic carbon supply. Philos. Trans. R. Soc. Lond. Ser. B 2011, 366, 3316–3329. [Google Scholar] [CrossRef] [PubMed]
- Chaer, G.M.; Resende, A.S.; Campello, E.F.C.; de Faria, S.M.; Boddey, R.M. Nitrogen-fixing legume tree species for the reclamation of severely degraded lands in Brazil. Tree Physiol. 2011, 31, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.G.; Baruch, Z. African grass invasion in the Americas: Ecosystem consequences and the role of ecophysiology. Biol. Invasions 2000, 2, 123–140. [Google Scholar] [CrossRef]
- Parsons, J.J. Spread of African pasture grasses to the American tropics. Rangel. Ecol. Manag. 1972, 25, 12–17. [Google Scholar] [CrossRef]
- Hooper, E.; Legendre, P.; Condit, R. Barriers to forest regeneration of deforested and abandoned land in Panama. J. Appl. Ecol. 2005, 42, 1165–1174. [Google Scholar] [CrossRef]
- Hooper, E.R.; Legendre, P.; Condit, R. Factors affecting community composition of forest regeneration in deforested, abandoned land in Panama. Ecology 2004, 85, 3313–3326. [Google Scholar] [CrossRef]
- D’Antonio, C.M.; Hughes, R.; Tunison, J. Long-term impacts of invasive grasses and subsequent fire in seasonally dry Hawaiian woodlands. Ecol. Appl. 2011, 21, 1617–1628. [Google Scholar] [CrossRef]
- Edwards, E.J.; Smith, S.A. Phylogenetic analyses reveal the shady history of C4 grasses. Proc. Natl. Acad. Sci. USA 2010, 107, 2532–2537. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology; Springer: Berlin, Germany, 2008; Volume 2. [Google Scholar]
- D’Antonio, C.M.; Hughes, R.F.; Vitousek, P.M. Factors influencing dynamics of two invasive C4 grasses in seasonally dry Hawaiian woodlands. Ecology 2001, 82, 89–104. [Google Scholar]
- Hammond, B.W. Saccharum spontaneum (Gramineae) in Panama: The physiology and ecology of invasion. J. Sustain. Forest. 1998, 8, 23–38. [Google Scholar] [CrossRef]
- Saltonstall, K.; Bonnett, G.D.; Aitken, K.S. A perfect storm: Ploidy and preadaptation facilitate Saccharum spontaneum escape and invasion in the Republic of Panama. Biol. Invasions 2021, 23, 1101–1115. [Google Scholar] [CrossRef]
- Craven, D.; Hall, J.; Verjans, J.M. Impacts of herbicide application and mechanical cleanings on growth and mortality of two timber species in Saccharum spontaneum grasslands of the Panama Canal Watershed. Restor. Ecol. 2009, 17, 751–761. [Google Scholar] [CrossRef]
- Boeschoten, L.E.; van Breugel, M.; Bailon, M.; Balbuena, J.; Nuñez, M.; Cerezo, A.; Hall, J.S. Framework species approach proves robust in restoring forest on fire prone invasive grass: A case study from Panama. J. Sustain. For. 2021, 40, 197–215. [Google Scholar] [CrossRef]
- Saltonstall, K.; Bonnett, G.D. Fire promotes growth and reproduction of Saccharum spontaneum (L.) in Panama. Biol. Invasions 2012, 14, 2479–2488. [Google Scholar] [CrossRef]
- Wishnie, M.; Dent, D.; Mariscal, E.; Deago, J.; Cedeno, N.; Ibarra, D.; Condit, R.; Ashton, P. Initial performance and reforestation potential of 24 tropical tree species planted across a precipitation gradient in the Republic of Panama. For. Ecol. Manag. 2007, 243, 39–49. [Google Scholar] [CrossRef]
- Park, A.; van Breugel, M.; Ashton, M.S.; Wishnie, M.; Mariscal, E.; Deago, J.; Ibarra, D.; Cedeño, N.; Hall, J.S. Local and regional environmental variation influences the growth of tropical trees in selection trials in the Republic of Panama. For. Ecol. Manag. 2010, 260, 12–21. [Google Scholar] [CrossRef]
- Wright, S.J.; Yavitt, J.B.; Wurzburger, N.; Turner, B.L.; Tanner, E.V.; Sayer, E.J.; Santiago, L.S.; Kaspari, M.; Hedin, L.O.; Harms, K.E. Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest. Ecology 2011, 92, 1616–1625. [Google Scholar] [CrossRef] [PubMed]
- Maynard, D.; Kalra, Y.; Crumbaugh, J. Nitrate and exchangeable ammonium nitrogen. Soil Sampl. Methods Anal. 1993, 1, 25–38. [Google Scholar]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Baldeck, C.A.; Harms, K.E.; Yavitt, J.B.; John, R.; Turner, B.L.; Valencia, R.; Navarrete, H.; Davies, S.J.; Chuyong, G.B.; Kenfack, D. Soil resources and topography shape local tree community structure in tropical forests. Proc. R. Soc. B 2013, 280, 20122532. [Google Scholar] [CrossRef] [PubMed]
- Novozamsky, I.; Houba, V.; Van Eck, R.; Van Vark, W. A novel digestion technique for multi-element plant analysis. Commun. Soil Sci. Plant Anal. 1983, 14, 239–248. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Zhu, J. Nitrogen stress reduces the efficiency of the C4CO2 concentrating system, and therefore quantum yield, in Saccharum (sugarcane) species. J. Exp. Bot. 1998, 49, 1227–1234. [Google Scholar] [CrossRef]
- Rubio, G.; Gutiérrez Boem, F.H.; Lavado, R.S. Responses of C3 and C4 grasses to application of nitrogen and phosphorus fertilizer at two dates in the spring. Grass Forage Sci. 2010, 65, 102–109. [Google Scholar] [CrossRef]
- Barger, N.N.; D’Antonio, C.M.; Ghneim, T.; Brink, K.; Cuevas, E. Nutrient limitation to productivity in a secondary savanna in Venezuela. Biotropica 2002, 34, 493–501. [Google Scholar] [CrossRef]
- Lafarge, M. Reproductive tillers in cut tall fescue swards: Differences according to sward age and fertilizer nitrogen application, and relationships with the local dynamics of the sward. Grass Forage Sci. 2006, 61, 182–191. [Google Scholar] [CrossRef]
- Jaramillo, V.J.; Detling, J.K. Small-scale heterogeneity in a semi-arid North American grassland. I. Tillering, N uptake and retranslocation in simulated urine patches. J. Appl. Ecol. 1992, 29, 1–8. [Google Scholar] [CrossRef]
- Moyer, J.; Sweeney, D. Long-term responses in the yield of Eastern Gamagrass [Tripsacum dactyloides (L.) L.] to nitrogen fertilizer under two harvest regimes in the United States. Grass Forage Sci. 2008, 63, 390–397. [Google Scholar] [CrossRef]
- Simoes, M.; Baruch, Z. Responses to simulated herbivory and water stress in two tropical C4 grasses. Oecologia 1991, 88, 173–180. [Google Scholar] [CrossRef]
- Baruch, Z.; Gómez, J.A. Dynamics of energy and nutrient concentration and construction cost in a native and two alien C4 grasses from two neotropical savannas. Plant Soil 1996, 181, 175–184. [Google Scholar] [CrossRef]
- Klimeš, L.; Klimešová, J. The effects of mowing and fertilization on carbohydrate reserves and regrowth of grasses: Do they promote plant coexistence in species-rich meadows? In Proceedings of the Ecology and Evolutionary Biology of Clonal Plants: Proceedings of Clone-2000, Obergurgl, Austria, 20–25 August 2000; pp. 141–160. [Google Scholar]
- Hooper, E.; Condit, R.; Legendre, P. Responses of 20 native tree species to reforestation strategies for abandoned farmland in Panama. Ecol. Appl. 2002, 12, 1626–1641. [Google Scholar] [CrossRef]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. Saccharum spontaneum L. In World Weeds: Natural Histories and Distribution; University of Hawaii Press: Honolulu, HI, USA, 1977; pp. 693–698. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
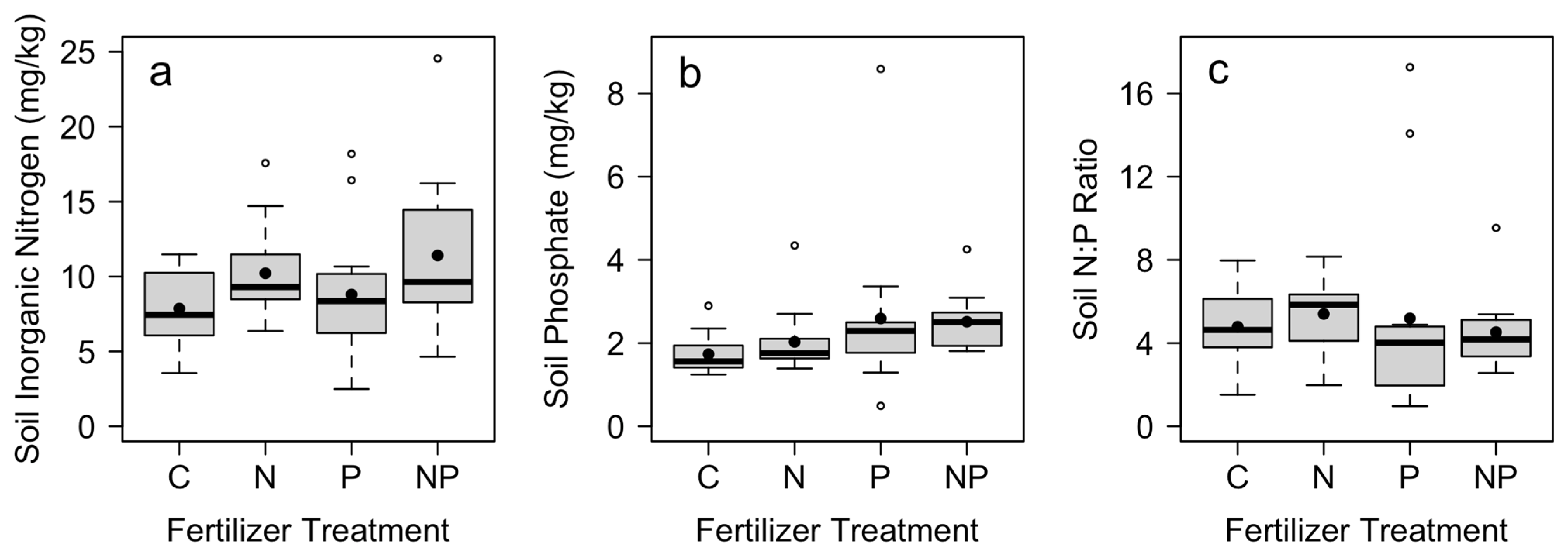
| Ammonium (mg/kg) | Nitrate (mg/kg) | Total Inorganic N (mg/kg) | Phosphate (mg/kg) | N:P | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | df | F | p | F | p | F | p | F | p | F | p |
| N addition | 1.33 | 9.20 | 0.005 | 0.10 | 0.10 | 9.73 | 0.004 | 0.11 | 0.75 | 0.001 | 0.98 |
| P addition | 1.33 | 1.77 | 0.19 | 0.59 | 0.59 | 1.78 | 0.19 | 4.14 | 0.05 | 0.08 | 0.78 |
| N × P interaction | 1.33 | 0.16 | 0.70 | 1.49 | 0.23 | 0.03 | 0.87 | 0.32 | 0.35 | 0.57 | 0.46 |
| Block | 11.33 | 5.91 | <0.001 | 2.51 | 0.02 | 5.95 | <0.001 | 1.17 | 0.58 | 1.12 | 0.38 |
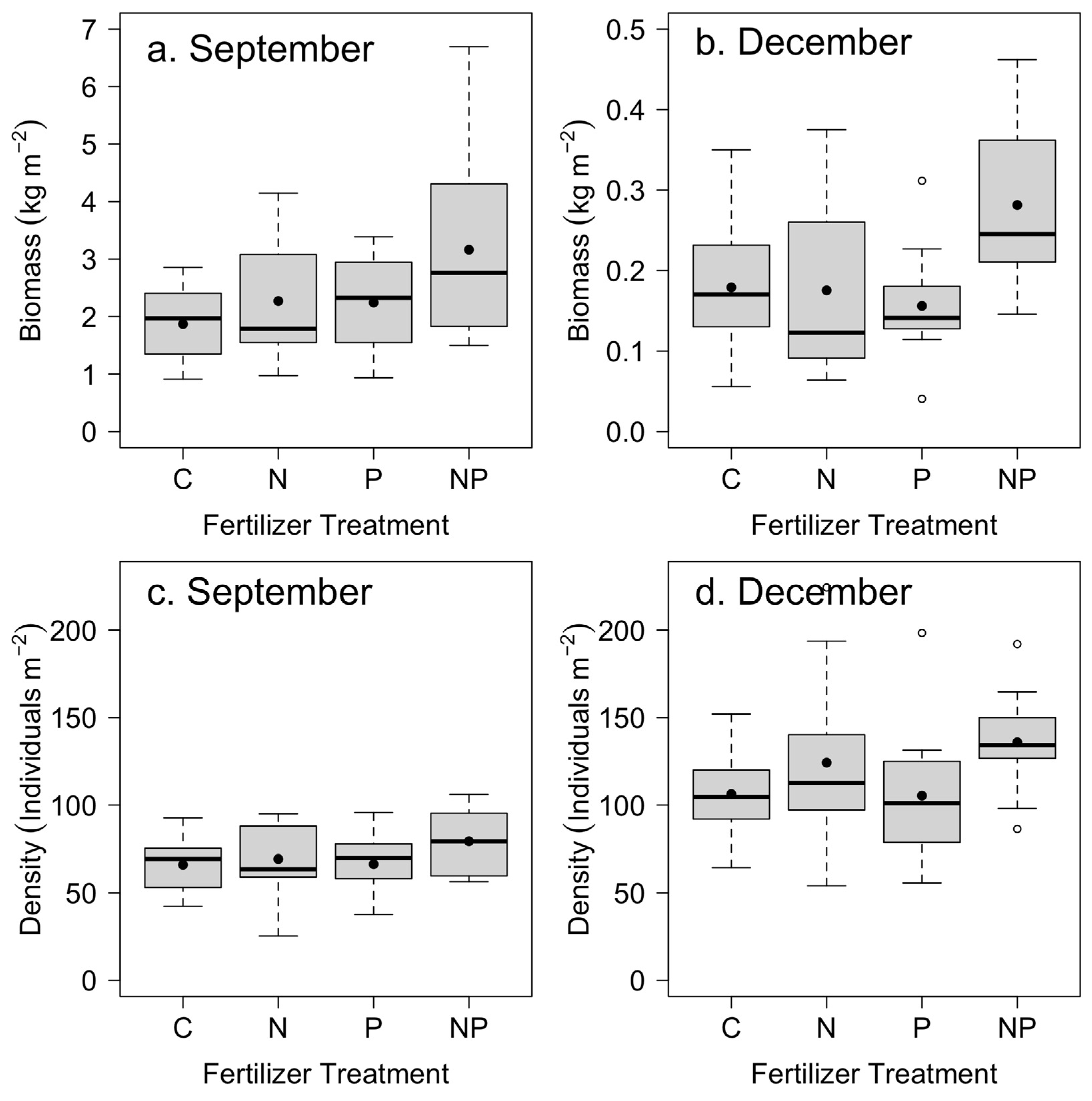
| Density | Biomass | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| September | December | September | December | ||||||
| Treatment | df | F | p | F | p | F | p | F | p |
| N addition | 1.33 | 2.87 | 0.10 | 5.84 | 0.02 | 6.78 | 0.01 | 6.88 | 0.01 |
| P addition | 1.33 | 1.17 | 0.29 | 0.29 | 0.59 | 6.23 | 0.02 | 3.20 | 0.08 |
| N × P interaction | 1.33 | 1.02 | 0.17 | 0.39 | 0.31 | 0.81 | 0.31 | 7.74 | 0.008 |
| Block | 11.33 | 1.53 | 0.32 | 1.22 | 0.53 | 3.36 | 0.003 | 2.20 | 0.04 |
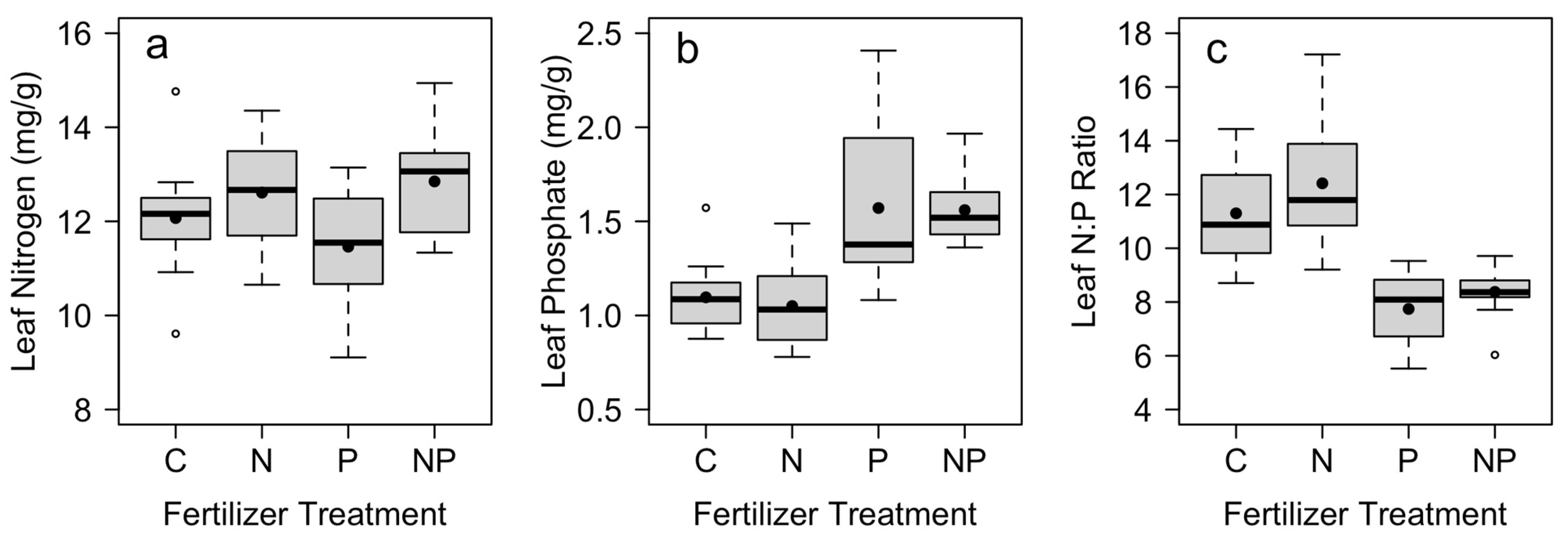
| Leaf N (mg/g) | Leaf P (mg/g) | Leaf N:P | |||||
|---|---|---|---|---|---|---|---|
| Treatment | df | F | p | F | p | F | p |
| N addition | 1.33 | 10.82 | 0.002 | 0.22 | 0.64 | 6.71 | 0.01 |
| P addition | 1.33 | 0.40 | 0.53 | 65.20 | <0.001 | 126.63 | <0.001 |
| N × P interaction | 1.33 | 2.10 | 0.16 | 0.08 | 0.77 | 0.51 | 0.48 |
| Block | 11.33 | 2.19 | 0.04 | 3.94 | 0.001 | 5.14 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cummings, J.A.; Parker, I.M.; Gilbert, G.S. The Influence of Nitrogen and Phosphorus Addition on Growth of the Invasive C4 Grass Saccharum spontaneum. Int. J. Plant Biol. 2023, 14, 474-482. https://doi.org/10.3390/ijpb14020036
Cummings JA, Parker IM, Gilbert GS. The Influence of Nitrogen and Phosphorus Addition on Growth of the Invasive C4 Grass Saccharum spontaneum. International Journal of Plant Biology. 2023; 14(2):474-482. https://doi.org/10.3390/ijpb14020036
Chicago/Turabian StyleCummings, Justin A., Ingrid M. Parker, and Gregory S. Gilbert. 2023. "The Influence of Nitrogen and Phosphorus Addition on Growth of the Invasive C4 Grass Saccharum spontaneum" International Journal of Plant Biology 14, no. 2: 474-482. https://doi.org/10.3390/ijpb14020036
APA StyleCummings, J. A., Parker, I. M., & Gilbert, G. S. (2023). The Influence of Nitrogen and Phosphorus Addition on Growth of the Invasive C4 Grass Saccharum spontaneum. International Journal of Plant Biology, 14(2), 474-482. https://doi.org/10.3390/ijpb14020036