
Bioactive Diepoxy Metabolites and Highly Oxygenated Triterpenoids from Marine and Plant-Derived Bacteria and Fungi


Abstract
1. Introduction
2. α,β-Diepoxy Metabolites Derived from Bacterial Species and Fungal Endophytes
3. Highly Oxygenated Triterpenoids Derived from Terrestrial Microbial Symbionts
4. Highly Oxygenated Triterpenoids Derived from Marine Bacterial Species and Fungal Endophytes
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koide, R.T. On holobionts, holospecies, and holoniches: The role of microbial symbioses in ecology and evolution. Microb. Ecol. 2023, 85, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Bright, M.; Bulgheresi, S. A complex journey: Transmission of microbial symbionts. Nat. Rev. Microbiol. 2010, 8, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Haygood, M.G.; Schmidt, E.W.; Davidson, S.K.; Faulkner, D.J. Microbial symbionts of marine invertebrates: Opportunities for microbial biotechnology. J. Mol. Microbiol. Biotechnol. 1999, 1, 33–43. [Google Scholar] [PubMed]
- Hays, G.; Patrick, W.G.; Ziesack, M.; Oxman, N.; Silver, P.A. Better together: Engineering and application of microbial symbioses. Curr. Opin. Biotechnol. 2015, 36, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, E.; Michel, L. Boundary lines in symbiosis forms. Symbiosis 2013, 60, 1–5. [Google Scholar] [CrossRef]
- Álvarez, C.; Jiménez-Ríos, L.; Iniesta-Pallarés, M.; Jurado-Flores, A.; Molina-Heredia, F.P.; Ng, C.K.Y.; Mariscal, V. Symbiosis between cyanobacteria and plants: From molecular studies to agronomic applications. J. Exp. Bot. 2023, 74, 6145–6157. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.; Godina, R.; Azevedo, S.G.; Matias, J.C.O. A comprehensive review of industrial symbiosis. J. Clean. Prod. 2020, 247, 119113. [Google Scholar] [CrossRef]
- Hooper, L. Do symbiotic bacteria subvert host immunity? Nat. Rev. Microbiol. 2009, 7, 367–374. [Google Scholar] [CrossRef]
- Piel, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep. 2009, 26, 338–362. [Google Scholar] [CrossRef]
- Grube, M.; Berg, G. Microbial consortia of bacteria and fungi with focus on the lichen symbiosis. Fungal Biol. Rev. 2009, 23, 72–85. [Google Scholar] [CrossRef]
- Vorburger, C.; Perlman, S.J. The role of defensive symbionts in host–parasite coevolution. Biol. Rev. 2018, 93, 1747–1764. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.P.; Hawrelak, J. The causes of intestinal dysbiosis: A Review. Altern. Med. Rev. 2004, 9, 180–197. [Google Scholar]
- Ducatelle, R.; Eeckhaut, V.; Haesebrouck, F.; Van Immerseel, F. A review on prebiotics and probiotics for the control of dysbiosis: Present status and future perspectives. Animal 2015, 9, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Carballo-Sánchez, M.P.; Alarcón, A.; Pérez-Moreno, J.; Ferrera-Cerrato, R. Agricultural and forestry importance of microorganism-plant symbioses: A microbial source for biotechnological innovations. Rev. Agric. Sci. 2022, 10, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Vil, V.; Gloriozova, T.A.; Poroikov, V.V.; Dembitsky, V.M. Naturally occurring of α,β-diepoxy-containing compounds: Origin, structures, and biological activities. Appl. Microbiol. Biotechnol. 2019, 103, 3249–3264. [Google Scholar] [CrossRef] [PubMed]
- Fretland, A.J.; Omiecinski, C.J. Epoxide hydrolases: Biochemistry and molecular biology. Chem. Biol. Interact. 2000, 129, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.W.; Morisseau, C.; Hammock, B.D. Epoxide hydrolases: Their roles and interactions with lipid metabolism. Prog. Lipid Res. 2005, 44, 1–51. [Google Scholar] [CrossRef]
- Dembitsky, V.M. Bioactive steroids bearing oxirane ring. Biomedicines 2023, 11, 2237. [Google Scholar] [CrossRef]
- Kuklev, D.V.; Dembitsky, V.M. Epoxy acetylenic lipids: Their analogues and derivatives. Prog. Lipid Res. 2014, 56, 67–91. [Google Scholar] [CrossRef]
- Arbuzov, Y.A. The diels–alder reaction with molecular oxygen as dienophile. Russ. Chem. Rev. 1965, 34, 558. [Google Scholar] [CrossRef]
- Jacobsen, E.N. Asymmetric catalysis of epoxide ring-opening reactions. Acc. Chem. Res. 2000, 33, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Bučko, M.; Kaniaková, K.; Hronská, H.; Gemeiner, P.; Rosenberg, M. Epoxide Hydrolases: Multipotential Biocatalysts. Int. J. Mol. Sci. 2023, 24, 7334. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Xia, J.; Zhou, W.; Zhou, L.; Hussain, G.; Li, Q.; Ostrikov, K. Adhesion and cohesion of epoxy-based industrial composite coatings. Compos. Part B Eng. 2020, 193, 108035. [Google Scholar] [CrossRef]
- Scotti, M.T.; Fernandes, M.B.; Ferreira, M.J.P.; Emerenciano, V.P. Quantitative structure–activity relationship of sesquiterpene lactones with cytotoxic activity. Bioorg. Med. Chem. 2007, 15, 2927–2934. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.R.; Varela, C.L.; Tavares-da-Silva, E.J.; Roleira, F.M.F. Epoxide containing molecules: A good or a bad drug design approach. Eur. J. Med. Chem. 2020, 201, 112327. [Google Scholar] [CrossRef] [PubMed]
- Vil, V.; Terent’ev, A.O.; Al Quntar, A.A.A.; Gloriozova, T.A.; Savidov, N.; Dembitsky, V.M. Oxetane-containing metabolites: Origin, structures, and biological activities. Appl. Microbiol. Biotechnol. 2019, 103, 2449–2467. [Google Scholar] [CrossRef] [PubMed]
- Dembitsky, V.M.; Gloriozova, T.A.; Poroikov, V.V. Naturally occurring marine α, β-epoxy steroids: Origin and biological activities. Vietnam J. Chem. 2018, 56, 409–433. [Google Scholar] [CrossRef]
- Lugtenberg, B.J.; Caradus, J.R.; Johnson, L.J. Fungal endophytes for sustainable crop production. FEMS Microbiol. Ecol. 2016, 92, fiw194. [Google Scholar] [CrossRef]
- Xu, K.; Li, X.-Q.; Zhao, D.-L.; Zhang, P. Antifungal secondary metabolites produced by the fungal endophytes: Chemical diversity and potential use in the development of biopesticides. Front. Microbiol. 2021, 12, 689527. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C.; Draeger, S.; Römmert, A.K.; Krohn, K. Endophytic fungi: A source of novel biologically active secondary metabolites. Mycol. Res. 2002, 106, 996–1004. [Google Scholar] [CrossRef]
- Borders, D.B.; Barbatschi, F.; Shay, A.J.; Shu, P. Fermentation, isolation, and characterization of LL-Z1220, a new antibiotic. Antimicrob. Agents Chemother. (Bethesda) 1969, 9, 233–235. [Google Scholar] [PubMed]
- Borders, D.B.; Shu, P.; Lancaster, J.E. Structure of LL-Z1220. New antibiotic containing a cyclohexene diepoxide ring system. J. Am. Chem. Soc. 1972, 94, 2540–2541. [Google Scholar] [CrossRef] [PubMed]
- Parker, W.L.; Rathnum, M.L.; Seiner, V.; Trejo, W.H.; Principe, P.A.; Sykes, R.B. Cepacin A and cepacin B, two new antibiotics produced by Pseudomonas cepacia. J. Antibiot. 1984, 37, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Sivakumari, V.; Dhinakaran, J.; Rajendran, A. Screening and productivity of penicillin antibiotic from Penicillium sp. J. Environ. Sci. Eng. 2009, 51, 247–248. [Google Scholar] [PubMed]
- Fujiwara, A.; Okuda, T.; Masuda, S.; Shiomi, Y.; Miyamoto, C.; Sekine, Y.; Tazoe, M.; Fujiwara, M. Fermentation, isolation and characterization of isonitrile antibiotics. Agric. Biol. Chem. 1982, 46, 1803–1809. [Google Scholar] [CrossRef]
- Tamura, A.; Kotani, H.; Naruto, S. Trichoviridin and dermadin from Trichoderma sp. TK-1. J. Antibiot. 1975, 28, 161–162. [Google Scholar] [CrossRef]
- Che, Y.; Gloer, J.B.; Koster, B.; Malloch, D. Decipienin A and decipienolides A-B: New bioactive metabolites from the coprophilous fungus Podopsora decipiens. J. Nat. Prod. 2002, 65, 916–919. [Google Scholar] [CrossRef]
- Holub, M.; Buděšínský, M.; Smítalová, Z.; Šaman, D.; Rychłewska, U. Structure of isosilerolide, relative and absolute configuration of silerolide and lasolide—Sesquiterpenic lactones of new stereoisomeric type of eudesmanolides. Collect. Czechoslov. Chem. Commun. 1985, 51, 903–929. [Google Scholar] [CrossRef]
- Massanet, G.M.; Guerra, F.M.; Jorge, Z.D.; Astorga, C. Sesquiterpenolides from Melanoselinum decipiens. Phytochemistry 1997, 45, 1645–1651. [Google Scholar] [CrossRef]
- Sundin, A.; Anke, H.; Bergquist, K.E.; Mayer, A.; Sheldrick, W.S.; Stadler, M.; Sterner, O. The structure determination of panellon and panellol, two 14-noreudesmanes isolated from Resupinatus leightonii. Tetrahedron 1993, 49, 7519–7524. [Google Scholar] [CrossRef]
- Tomoda, H.; Tabata, N.; Yang, D.J.; Takayanagi, H.; Omura, S. Terpendoles, novel ACAT inhibitors produced by Albophoma yamanashiensis. III. Production, isolation and structure elucidation of new components. J. Antibiot. 1995, 48, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Sassa, T.; Kinoshita, H.; Nukina, M. Acremolactone A, a novel herbicidal epoxy-hydropyranyl γ-lactone from Acremonium roseum I4267. J. Antibiot. 1998, 51, 967–969. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wu, H.C.; Ge, H.M.; Zang, L.Y.; Bei, Y.C.; Niu, Z.Y.; Wei, W.; Feng, X.J.; Ding, S.; Ng, S.W.; Shen, P.P.; et al. Diaporine, a novel endophyte-derived regulator of macrophage differentiation. Org. Biomol. Chem. 2014, 12, 6545–6548. [Google Scholar] [CrossRef] [PubMed]
- Itoh, J.; Shomura, T.; Tsuyuki, T.; Yoshida, J.; Ito, M.; Sezaki, M.; Kojima, M. Studies on a new antibiotic SF-2330. I. Taxonomy, isolation and characterization. J. Antibiot. 1986, 39, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Itoh, J.; Tsuyuki, T.; Fujita, K.; Sezaki, M. Studies on a new antibiotic SF-2330. II. The structural elucidation. J. Antibiot. 1986, 39, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Fujii, N.; Katsuyama, T.; Kobayashi, E.; Hara, M.; Nakano, H. The clecarmycins, new antitumor antibiotics produced by Streptomyces: Fermentation, isolation and biological properties. J. Antibiot. 1995, 48, 768–772. [Google Scholar] [CrossRef] [PubMed]
- Iwami, M.; Kawai, Y.; Kiyoto, S.; Terano, H.; Kohsaka, M.; Aoki, H.; Imanaka, H. A new antitumor antibiotic, chromoxymycin. I. Taxonomic studies on the producing strain: A new subspecies of the genus Streptomyces. J. Antibiot. 1986, 39, 6–11. [Google Scholar] [CrossRef]
- Hori, Y.; Hino, M.; Kawai, Y.; Kiyoto, S.; Terano, H.; Kohsaka, M.; Aoki, H.; Hashimoto, M.; Imanaka, H. A new antitumor antibiotic, chromoxymycin. II. Production, isolation, characterization and antitumor activity. J. Antibiot. 1986, 39, 12–16. [Google Scholar] [CrossRef]
- Sato, Y.; Watabe, H.; Nakazawa, T.; Shomura, T.; Yamamoto, H.; Sezaki, M.; Kondo, S. Ankinomycin, a potent antitumor antibiotic. J. Antibiot. 1989, 42, 149–152. [Google Scholar] [CrossRef][Green Version]
- Igarashi, Y.; Kuwamori, Y.; Takagi, K.; Ando, T.; Fudou, R.; Furumai, T.; Oki, T. Xanthoepocin, a new antibiotic from Penicillium simplicissimum IFO5762. J. Antibiot. 2000, 53, 928–933. [Google Scholar] [CrossRef]
- Dhar, T.K.; Siddiqui, K.A.I.; Ali, E. Structure of phaseolinone, a novel phytotoxin from Macrophomina phaseolina. Tetrahedron Lett. 1982, 23, 5459–5462. [Google Scholar]
- Smetanina, O.F.; Yurchenko, A.N.; Afiyatullov, S.S.; Kalinovsky, A.I.; Pushilin, M.A.; Khudyakova, Y.A.; Slinkina, N.N.; Ermakova, S.P.; Yurchenko, E.A. Oxirapentyns B–D produced by a marine sediment-derived fungus Isaria felina (DC.) Fr. Phytochem. Lett. 2012, 5, 165–169. [Google Scholar] [CrossRef]
- Rukachaisirikul, V.; Kannai, S.; Klaiklay, S.; Phongpaichit, S.; Sakayaroj, J. Rare 2-phenylpyran-4-ones from the seagrass-derived fungi Polyporales PSU-ES44 and PSU-ES83. Tetrahedron 2013, 69, 6981–6986. [Google Scholar] [CrossRef]
- Vicente, J.; Stewart, A.K.; van Wagoner, R.M.; Elliott, E.; Bourdelais, A.J.; Wright, J.L.C. Monacyclinones, new angucyclinone metabolites isolated from Streptomyces sp. M7_15 associated with the Puerto Rican sponge Scopalina ruetzleri. Mar. Drugs 2015, 13, 4682–4700. [Google Scholar] [CrossRef] [PubMed]
- Daferner, M.; Mensch, S.; Anke, T.; Sterner, O. Hypoxysordarin, a new sordarin derivative from Hypoxylon croceum. Z. Naturforsch. C 1999, 54, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Debbab, A.; Aly, A.H.; Proksch, P. Endophytes and associated marine derived fungi—Ecological and chemical perspectives. Fungal Divers. 2012, 57, 45–83. [Google Scholar] [CrossRef]
- Govinda Rajulu, M.B.; Thirunavukkarasu, N.; Suryanarayanan, T.S.; Venkatachalam, A. Endophytic fungi of marine algae and seagrasses: A novel source of chitin modifying enzymes. Mycosphere 2015, 6, 345–355. [Google Scholar]
- Elkhateeb, W.; Daba, G.M. Marine endophytes a natural novel source for a treasure of bioactive compounds. J. Adv. Microbiol. 2022, 5, 018. [Google Scholar]
- Gareth, J.E.B.; Stanley, S.J.; Pinruan, U. Marine endophyte sources of new chemical natural products: A review. Bot. Mar. 2008, 51, 163–170. [Google Scholar]
- Liu, Y.; Palaniveloo, K.; Alias, S.A.; Sathiya Seelan, J.S. Species diversity and secondary metabolites of Sarcophyton-associated marine fungi. Molecules 2021, 26, 3227. [Google Scholar] [CrossRef]
- Fernandes, L.; Kamat, S.Y.; Paknikar, S.K. New diterpenoids of the brown seaweed Stoechospermum marginatum: Structure of stoechospermol. Tetrahedron Lett. 1980, 21, 2249–2252. [Google Scholar]
- Gerwick, W.H.; Fenical, W.; Ven Engen, D.; Clardy, J. Isolation and structure of spatol, a potent inhibitor of cell reolication from the brown seaweed Spatoglossum schmittii. J. Am. Chem. Soc. 1980, 102, 7991–7993. [Google Scholar] [CrossRef]
- Erickson, K.L.; Beutler, J.A.; Gray, G.N.; Cardellina, J.H., II; Boyd, M.R. Majapolene A, a cytotoxic peroxide, and related sesquiterpenes from the red alga Laurencia majuscula. J. Nat. Prod. 1995, 58, 1848–1860. [Google Scholar] [CrossRef]
- Kimura, J.; Kamada, N.; Tsujimoto, Y. Fourteen chamigrane derivatives from a red alga, Laurencia nidifica. Bull. Chem. Soc. Jpn. 1999, 72, 289–292. [Google Scholar] [CrossRef]
- Cikoš, A.M.; Jurin, M.; Čož-Rakovac, R.; Gašo-Sokač, D.; Jokić, S.; Jerković, I. Update on sesquiterpenes from red macroalgae of the Laurencia genus and their biological activities (2015–2020). Algal Res. 2021, 56, 102330. [Google Scholar] [CrossRef]
- Nakamura, H.; Asari, T.; Murai, A.; Kan, Y.; Kondo, T.; Yoshida, K.; Ohizumi, Y. Zooxanthellatoxin-A, a potent vasoconstrictive 62-membered lactone from a symbiotic Dinoflagellate. J. Am. Chem. Soc. 1995, 117, 550–551. [Google Scholar] [CrossRef]
- Ermolenko, E.V.; Imbs, A.B.; Gloriozova, T.A.; Poroikov, V.V.; Sikorskaya, T.V.; Dembitsky, V.M. Chemical diversity of soft coral steroids and their pharmacological activities. Mar. Drugs 2020, 18, 613. [Google Scholar] [CrossRef]
- Damare, S.; Singh, P.; Raghukumar, S. Biotechnology of Marine Fungi. In Biology of Marine Fungi. Progress in Molecular and Subcellular Biology; Raghukumar, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 53. [Google Scholar] [CrossRef]
- Imbs, A.B.; Dembitsky, V.M. Coral Lipids. Mar. Drugs 2023, 21, 539. [Google Scholar] [CrossRef]
- Dembitsky, V.M. Fascinating furanosteroids and their pharmacological profile. Molecules 2023, 28, 5669. [Google Scholar] [CrossRef]
- Dembitsky, V.M. Biological activity and structural diversity of steroids containing aromatic rings, phosphate groups, or halogen atoms. Molecules 2023, 28, 5549. [Google Scholar] [CrossRef]
- Zhang, G.W.; Ma, X.Q.; Su, J.Y.; Zhang, K.; Kurihara, H.; Yao, X.S. Two new bioactive sesquiterpenes from the soft coral Sinularia sp. Nat. Prod. Res. 2006, 20, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Lelong, H.; Ahond, A.; Chiaroni, A.; Poupat, C.; Riche, C.; Potier, P.; Pusset, J.; Pusset, M.; Laboute, P.; Menou, J.L. Invertébrés marins du lagon Néo-Calédonien, VIII. métabolites terpéniques de Xenia membranacea. J. Nat. Prod. 1987, 50, 203–210. [Google Scholar] [CrossRef]
- Bishara, A.; Rudi, A.; Goldberg, I.; Benayahu, Y.; Kashman, Y. Novaxenicins A–D and xeniolides I–K, seven new diterpenes from the soft coral Xenia novaebrittanniae. Tetrahedron 2006, 62, 12092–12097. [Google Scholar] [CrossRef]
- Rodríguez, A.D.; Boulanger, A.; Martínez, J.R.; Huang, S.D. Sesquiterpene lactones from the Caribbean Sea plume Pseudopterogorgia americana. J. Nat. Prod. 1998, 61, 451–455. [Google Scholar] [CrossRef]
- Rodríguez, A.D.; Soto, J.J. Pseudopterane and norcembrane diterpenoids from the Caribbean Sea plume Pseudopterogorgia acerosa. J. Nat. Prod. 1998, 61, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.D.; Boulanger, A. Americanolides A−C, new guaianolide sesquiterpenes from the Caribbean Sea plume Pseudopterogorgia americana. J. Nat. Prod. 1996, 59, 653–657. [Google Scholar] [CrossRef]
- Berrueab, F.; Kerr, R.G. Diterpenes from gorgonian corals. Nat. Prod. Rep. 2009, 26, 681–710. [Google Scholar] [CrossRef] [PubMed]
- Iwagawa, T.; Amano, Y.; Nakatani, M.; Hase, T. New xenia diterpenoids from a soft coral, Xenia species containing fatty acyl side chains. Bull. Chem. Soc. Jpn. 1996, 69, 1309–1312. [Google Scholar] [CrossRef]
- König, G.M.; Coll, J.C.; Bowden, B.F.; Gulbis, J.M.; MacKay, M.F.; Labarre, S.C.; Laurent, D. The structure determination of a xenicane diterpene from Xenia garciae. J. Nat. Prod. 1989, 52, 294–299. [Google Scholar] [CrossRef]
- Ng, S.Y.; Phan, C.S.; Ishii, T.; Kamada, T.; Hamada, T.; Santhanaraju Vairappan, C. Terpenoids from marine soft coral of the genus Xenia in 1977 to 2019. Molecules 2020, 25, 5386. [Google Scholar] [CrossRef]
- Almourabit, A.; Ahond, A.; Chiaroni, A.; Poupat, C.; Riche, C.; Potier, P.; Laboute, P.; Menou, J.L. Invertébrés marins du lagon Néo-Calédonien, IX. Havannachlorhydrines, nouveaux métabolites de Xenia membranacea: Étude structurale et configuration absolue. J. Nat. Prod. 1988, 51, 282–292. [Google Scholar] [CrossRef]
- Schmitt, S.; Tsai, P.; Bell, J.; Fromont, J.; Ilan, M.; Lindquist, N. Assessing the complex sponge microbiota: Core, variable and species-specific bacterial communities in marine sponges. ISME J. 2012, 6, 564–576. [Google Scholar] [CrossRef] [PubMed]
- Dat, T.T.H.; Steinert, G.; Cuc, N.T.K.; Smidt, H.; Sipkema, D. Archaeal and bacterial diversity and community composition from 18 phylogenetically divergent sponge species in Vietnam. Peer J. 2018, 6, e4970. [Google Scholar] [CrossRef]
- Steinert, G.; Rohde, S.; Janussen, D.; Blaurock, C.; Schupp, P.J. Host-specific assembly of sponge-associated prokaryotes at high taxonomic ranks. Sci. Rep. 2017, 7, 2542. [Google Scholar] [CrossRef]
- Taylor, J.A.; Palladino, G.; Wemheuer, B.; Steinert, G.; Sipkema, D.; Williams, T.J. Phylogeny resolved, metabolism revealed: Functional radiation within a widespread and divergent clade of sponge symbionts. ISME J. 2021, 15, 503–519. [Google Scholar] [CrossRef] [PubMed]
- Moitinho-Silva, L.; Nielsen, S.; Amir, A.; Gonzalez, A.; Ackermann, G.L.; Cerrano, C. The sponge microbiome project. Giga Science 2017, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Moitinho-Silva, L.; Lurgi, M.; Bjork, J.R.; Easson, C.; Astudillo-Garcia, C. Diversity, structure and convergent evolution of the global sponge microbiome. Nat. Commun. 2016, 7, 11870. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tietjen, I.; Chen, M.; Williams, D.E.; Daoust, J.; Brockman, M.A.; Andersen, R.J. Sesterterpenoids isolated from the sponge Phorbas sp. activate latent HIV-1 provirus expression. J. Org. Chem. 2016, 81, 11324–11334. [Google Scholar] [CrossRef]
- Chianese, G.; Yu, H.B.; Yang, F.; Sirignano, C.; Luciano, P.; Han, B.N.; Khan, S.; Lin, H.W.; Taglialatela-Scafati, O. PPAR Modulating polyketides from a Chinese Plakortis simplex and clues on the origin of their chemodiversity. J. Org. Chem. 2016, 81, 5135–5143. [Google Scholar] [CrossRef]
- De Marino, S.; Ummarino, R.; D’Auria, M.V.; Chini, M.G.; Bifulco, G.; Renga, B.; D’Amore, C.; Fiorucci, S.; Debitus, C.; Zampella, A. Theonellasterols and conicasterols from Theonella swinhoei. Novel marine natural ligands for human nuclear receptors. J. Med. Chem. 2011, 54, 3065–3075. [Google Scholar] [CrossRef]
- Cueto, M.; D’Croz, L.; Mate, J.L.; San-Martın, A.; Darias, J. Elysiapyrones from Elysia diomedea. Do such metabolites evidence an enzymatically assisted electrocyclization cascade for the biosynthesis of their bicyclo [4.2.0]octane core? Org. Lett. 2005, 7, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Olatunji, T.L.; Odebunmi, C.A.; Adetunji, A.E. Biological activities of limonoids in the Genus Khaya (Meliaceae): A review. Future J. Pharm. Sci. 2021, 7, 74. [Google Scholar] [CrossRef]
- Joshi, R.K. Bioactive usual and unusual triterpenoids derived from natural sources used in traditional medicine. Chem. Biodiver. 2023, 20, e202200853. [Google Scholar] [CrossRef] [PubMed]
- Dembitsky, V.M.; Dzhemileva, L.; Gloriozova, T.; D’yakonov, V.D. Natural and synthetic drugs used for the treatment of the dementia. Biochem. Biophys. Res. Commun. 2020, 524, 772–783. [Google Scholar] [CrossRef] [PubMed]
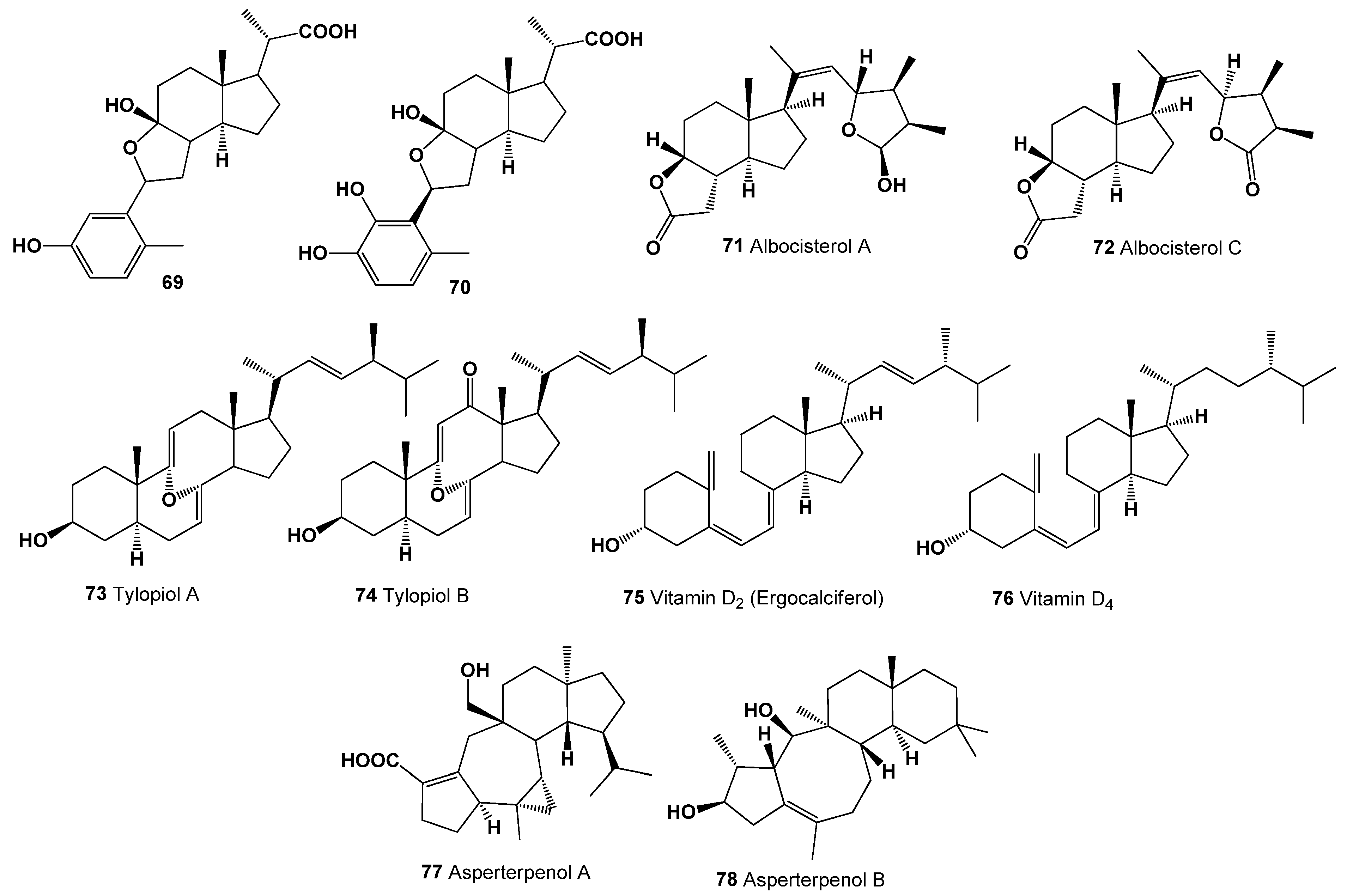
- Dembitsky, V.M. In Silico Prediction of steroids and triterpenoids as potential regulators of lipid metabolism. Mar. Drugs 2021, 19, 650. [Google Scholar] [CrossRef] [PubMed]
- Gezen-Ak, D.; Dursun, E. Vitamin D, a secosteroid hormone and its multifunctional receptor, vitamin D receptor, in Alzheimer’s type neurodegeneration. J. Alzheimer’s Disease 2023, 95, 1273–1299. [Google Scholar] [CrossRef] [PubMed]
- Karampela, I.; Sakelliou, A.; Vallianou, N. Vitamin D and obesity: Current evidence and controversies. Curr. Obes. Rep. 2021, 10, 162–180. [Google Scholar] [CrossRef]
- Amrein, K.; Scherkl, M.; Hoffmann, M. Vitamin D deficiency 2.0: An update on the current status worldwide. Eur. J. Clin. Nutr. 2020, 74, 1498–1513. [Google Scholar] [CrossRef]
- Duecker, F.L.; Reuß, F.; Heretsch, F. Rearranged ergostane-type natural products: Chemistry, biology, and medicinal aspects. Org. Biomol. Chem. 2019, 17, 1624–1633. [Google Scholar] [CrossRef]
- Wu, J.; Zhao, J.; Zhang, T.; Gu, Y.; Khan, I.A.; Zou, Z.; Xu, Q. Naturally occurring physalins from the genus Physalis: A review. Phytochemistry 2021, 191, 112925. [Google Scholar] [CrossRef]
- Grishko, V.V.; Galaiko, N.V. Structural diversity, natural sources and pharmacological potential of naturally occurring A-seco-triterpenoids studies. Nat. Prod. Chem. 2016, 51, 51–149. [Google Scholar]
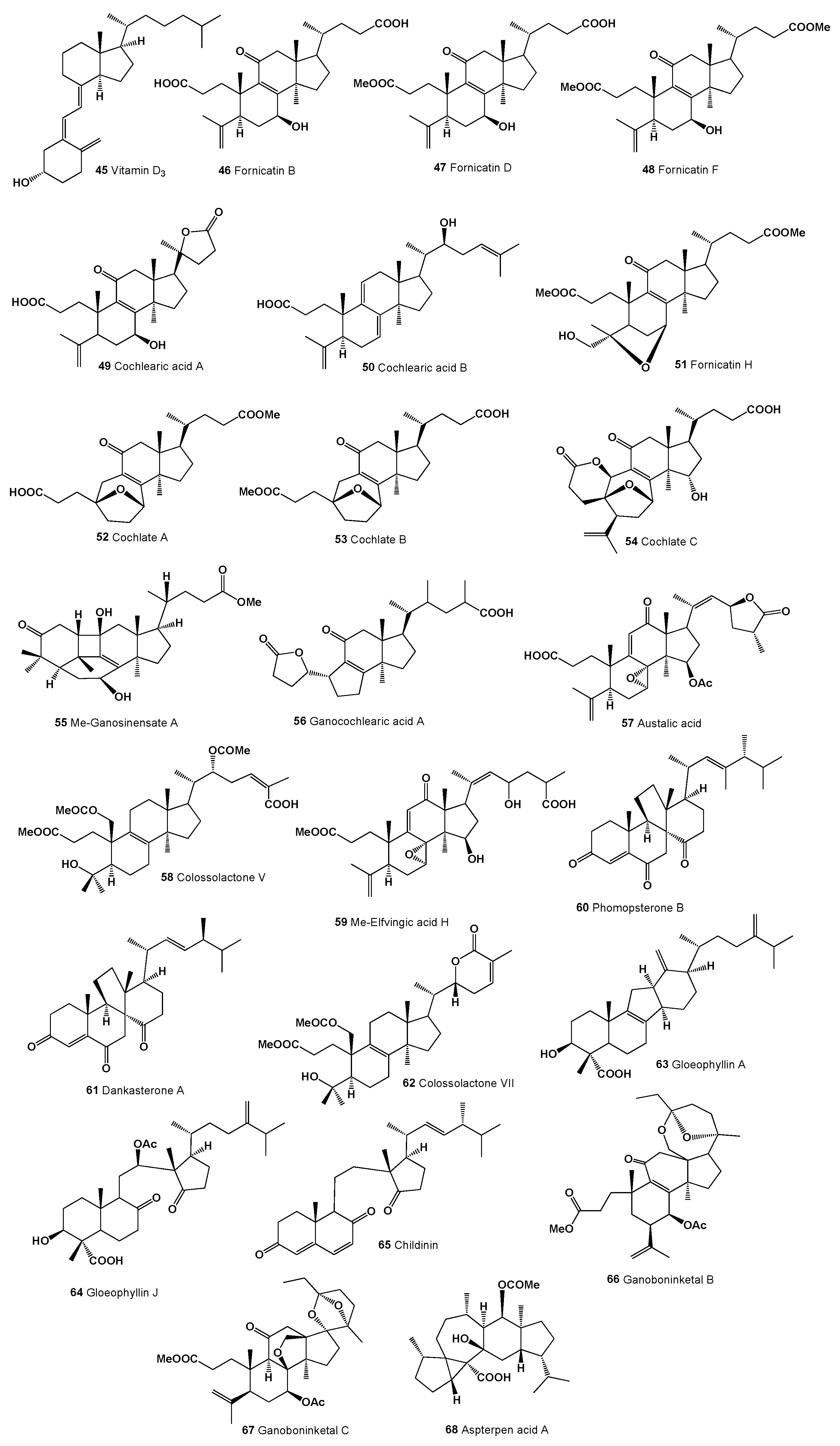
- Peng, X.R.; Wang, X.; Zhou, L.; Hou, B.; Zuo, Z.L.; Qiu, M.H. Ganocochlearic acid A, a rearranged hexanorlanostane triterpenoid, and cytotoxic triterpenoids from the fruiting bodies of Ganoderma cochlear. RSC Adv. 2015, 5, 95212–95222. [Google Scholar] [CrossRef]
- Peng, X.R.; Liu, J.Q.; Wang, C.F.; Li, X.Y.; Shu, Y.; Zhou, L.; Qiu, M.H. Hepatoprotective effects of triterpenoids from Ganoderma cochlear. J. Nat. Prod. 2014, 77, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Galappaththi, M.C.A.; Patabendige, N.M.; Premarathne, B.M.; Hapuarachchi, K.K.; Tibpromma, S.; Dai, D.-Q.; Suwannarach, N.; Rapior, S.; Karunarathna, S.C. A review of Ganoderma triterpenoids and their bioactivities. Biomolecules 2023, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A.; Connolly, V.M. Triterpenoids. Nat. Prod. Rep. 2020, 37, 962–998. [Google Scholar] [CrossRef]
- Leon, F.; Valencia, M.; Rivera, A.; Nieto, I.; Quintana, J.; Estevez, F.; Bermejo, J. Novel cytostatic lanostanoid triterpenes from Ganoderma australe. Helv. Chim. Acta 2003, 86, 3088–3095. [Google Scholar] [CrossRef]
- Yoshikawa, K.; Nishimura, N.; Bando, S.; Arihara, S.; Matsumura, E.; Katayama, S. New lanostanoids, elfvingic acids A−H, from the fruit body of Elfvingia applanata. J. Nat. Prod. 2002, 65, 548–552. [Google Scholar] [CrossRef]
- Kleinwächter, P.; Anh, N.; Kiet, T.T.; Schlegel, B.; Dahse, H.M.; Härtl, A.; Gräfe, U. Colossolactones, new triterpenoid metabolites from a Vietnamese mushroom Ganoderma colossum. J. Nat. Prod. 2001, 64, 236–239. [Google Scholar] [CrossRef]
- Hu, Z.; Wu, Y.; Xie, S.; Sun, W.; Guo, Y.; Li, X.N.; Liu, J.; Li, H.; Wang, J.; Luo, Z.; et al. Phomopsterones A and B, two functionalized ergostane-type steroids from the endophytic fungus Phomopsis sp. TJ507A. Org. Lett. 2017, 19, 258–261. [Google Scholar] [CrossRef]
- Han, J.J.; Bao, L.; Tao, Q.Q.; Yao, Y.J.; Liu, X.Z.; Yin, W.B.; Liu, H.W. Gloeophyllins A–J, cytotoxic ergosteroids with various skeletons from a Chinese Tibet fungus Gloeophyllum abietinum. Org. Lett. 2015, 17, 2538–2541. [Google Scholar] [CrossRef]
- Zhao, Z.Z.; Chen, H.P.; Huang, Y.; Zhang, S.B.; Li, Z.H.; Feng, T.; Liu, J.K. Bioactive polyketides and 8,14-seco-ergosterol from fruiting bodies of the ascomycete Daldinia childiae. Phytochemistry 2017, 142, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Ren, J.; Han, J.; Bao, L.; Li, L.; Yao, Y.; Sun, C.; Zhou, B.; Liu, H. Ganoboninketals A–C, antiplasmodial 3,4-seco-27-norlanostane triterpenes from Ganoderma boninense Pat. J. Nat. Prod. 2014, 77, 1847–1852. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, Y.; Chen, S.; Liu, Y.; Lu, Y.; Chen, D.; Lin, Y.; Huang, X. Aspterpenacids A and B, two sesterterpenoids from a Mangrove endophytic fungus Aspergillus terreus H010. Org. Lett. 2016, 18, 1406–1409. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Giovannini, P.P.; Fantin, G.; Medici, A.; Pedrini, P. New 9,10-secosteroids from biotransformations of hyodeoxycholic acid with Rhodococcus spp. Helv. Chim. Acta 2013, 96, 1062–1071. [Google Scholar] [CrossRef]
- Chen, Z.M.; Yang, X.Y.; Fan, Q.Y.; Lia, Z.H.; Weia, K.; Chena, H.P.; Fenga, T.; Liu, J.K. Three novel degraded steroids from cultures of the Basidiomycete Antrodiella albocinnamomea. Steroids 2014, 87, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Luo, X.D.; Ma, Y.B.; Liu, J.K.; Wu, D.G.; Zhao, B.; Lu, Y.; Zheng, Q.T. Two novel secoergosterols from the fungus Tylopilus plumbeoviolaceus. J. Nat. Prod. 2000, 63, 534–536. [Google Scholar] [CrossRef] [PubMed]
- Phillips, K.M.; Horst, R.L.; Koszewski, N.J.; Simon, R.R. Vitamin D4 in mushrooms. PLoS ONE 2012, 7, e40702. [Google Scholar] [CrossRef]
- Xiao, Z.; Huang, H.; Shao, C.; Xia, X.; Ma, L.; Huang, X.; Lu, Y.; Lin, Y.; Long, Y.; She, Z. Asperterpenols A and B, new sesterterpenoids isolated from a mangrove endophytic fungus Aspergillus sp. 085242. Org. Lett. 2013, 15, 2522–2525. [Google Scholar] [CrossRef]
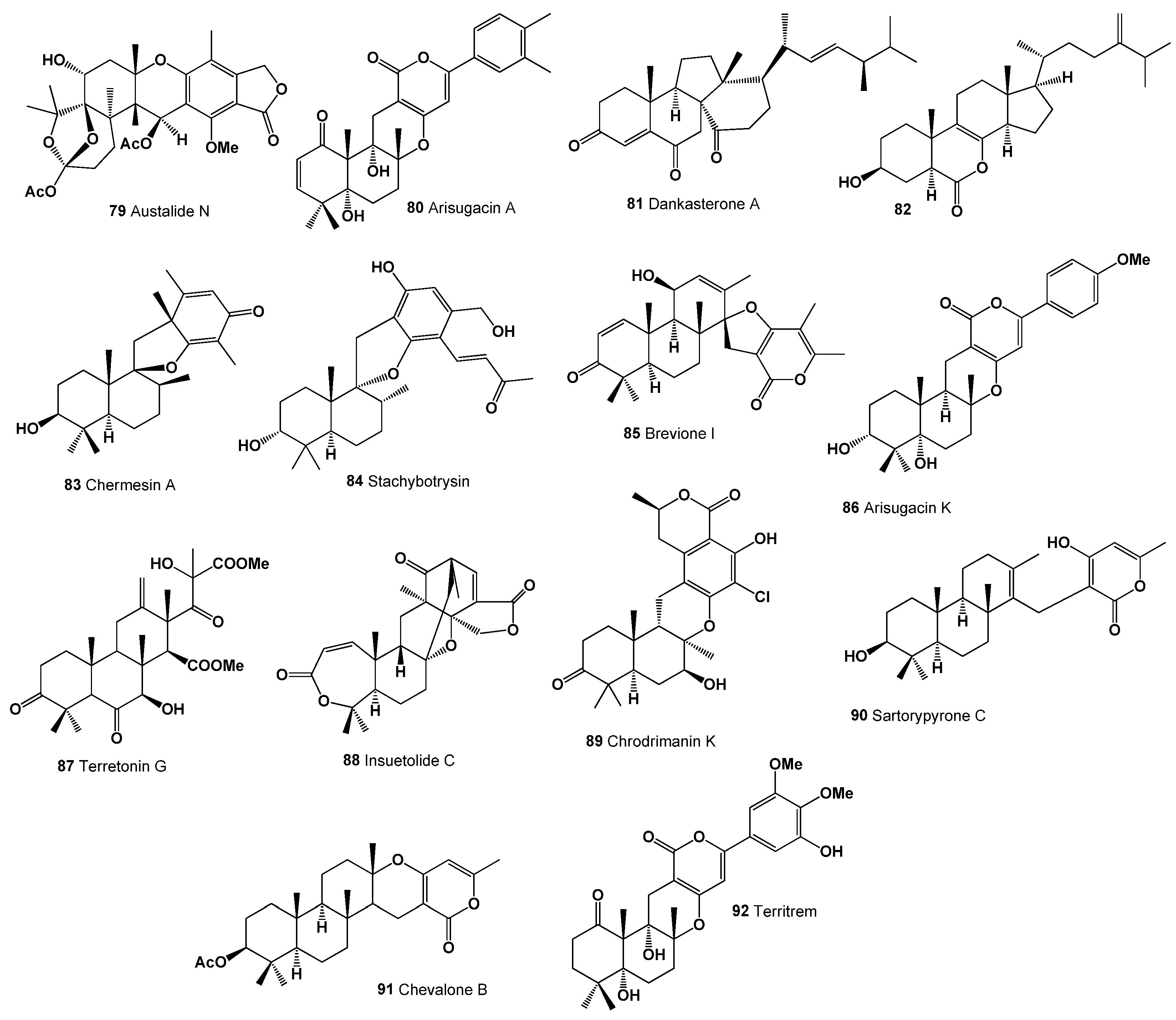
- Zhou, Y.M.; Mandi, A.; Debbab, A.; Wray, V.; Schulz, B.; Muller, W.E.G.; Lin, W.H.; Proksch, P.; Kurtan, T.; Aly, A.H. New austalides from the sponge-associated fungus Aspergillus sp. Eur. J. Org. Chem. 2011, 30, 6009–6019. [Google Scholar] [CrossRef]
- Zhou, Y.M.; Debbab, A.; Wray, V.; Lin, W.H.; Schulz, B.; Trepos, R.; Pile, C.; Hellio, C.; Proksch, P.; Aly, A.H. Marine bacterial inhibitors from the sponge-derived fungus Aspergillus sp. Tetrahedron Lett. 2014, 55, 2789–2792. [Google Scholar] [CrossRef]
- Nong, X.H.; Wang, Y.F.; Zhang, X.Y.; Zhou, M.P.; Xu, X.Y.; Qi, S.H. Territrem and butyrolactone derivatives from a marine-derived fungus Aspergillus terreus. Mar. Drugs 2014, 12, 6113–6124. [Google Scholar] [CrossRef] [PubMed]
- Kumla, D.; Aung, T.S.; Buttachon, S.; Dethoup, T.; Gales, L.; Pereira, J.A.; Inácio, Â.; Costa, P.M. A new dihydrochromone dimer and other secondary metabolites from cultures of the marine sponge-associated fungi Neosartorya fennelliae KUFA 0811 and Neosartorya tsunodae KUFC 9213. Mar. Drugs 2017, 15, 375. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.M.; Li, X.M.; Meng, L.; Li, C.S.; Huang, C.G.; Wang, B.G. 7-Nor-ergosterolide, a pentalactone-containing norsteroid and related steroids from the marine-derived endophytic Aspergillus ochraceus EN-31. J. Nat. Prod. 2010, 73, 1780–1784. [Google Scholar] [CrossRef] [PubMed]
- Onodera, H.; Ichimura, M.; Baba, K.; Agatsuma, T.; Sasho, S.; Suzuki, M.; Iwamoto, S.; Kakita, S. Nerve Trunk Cell Propagation Accelerator. International Patent Application No. PCT/JP2009/051417; WO 2009096445A1, 6 August 2009. [Google Scholar]
- Liu, H.; Li, X.M.; Liu, Y.; Zhang, P.; Wang, J.N.; Wang, B.G. Chermesins A–D: Meroterpenoids with a drimane-type spirosesquiterpene skeleton from the marine algal-derived endophytic fungus Penicillium chermesinum EN-480. J. Nat. Prod. 2016, 79, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Ko, S.K.; Kim, H.M.; Kim, G.H.; Son, S.; Kim, G.S.; Hwang, G.J. Stachybotrysin, an osteoclast differentiation inhibitor from the marine-derived fungus Stachybotrys sp. KCB13F013. J. Nat. Prod. 2016, 79, 2703–2708. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ye, D.; Shao, Z.; Cui, C.; Che, Y. A sterol and spiroditerpenoids from a Penicillium sp. isolated from a deep sea sediment sample. Mar. Drugs 2012, 10, 497–508. [Google Scholar] [CrossRef]
- Li, X.; Miao, F.; Liang, X.; Ji, N. Meroterpenes from an algicolous strain of Penicillium echinulatum. Magn. Reson. Chem. 2014, 52, 247–250. [Google Scholar] [CrossRef]
- Fukuda, T.; Tomoda, H.; Kurihara, Y.; Kanamoto, A. Terretonin G, a new sesterterpenoid antibiotic from marine-derived Aspergillus sp. OPMF00272. J. Antibiot. 2014, 67, 593–595. [Google Scholar] [CrossRef]
- Trisuwan, K.; Rukachaisirikul, V.; Kaewpet, M.; Phongpaichit, S.; Hutadilok-Towatana, N.; Preedanon, S.; Sakayaroj, J. Sesquiterpene and xanthone derivatives from the sea fan-derived fungus Aspergillus sydowii PSU-F154. J. Nat. Prod. 2012, 74, 1663–1667. [Google Scholar] [CrossRef]
- Kong, F.D.; Ma, Q.Y.; Huang, S.Z.; Wang, P.; Wang, J.F.; Zhou, L.M.; Yuan, J.Z.; Dai, H.F.; Zhao, Y.X. Chrodrimanins K–N and related meroterpenoids from the fungus Penicillium sp. SCS-KFD09 isolated from a marine worm, Sipunculus nudus. J. Nat. Prod. 2017, 80, 1039–1047. [Google Scholar]
- Gomes, N.M.; Bessa, L.J.; Buttachon, S.; Costa, P.M.; Buaruang, J.; Dethoup, T.; Silva, A.M.; Kijjoa, A. Antibacterial and antibiofilm activities of tryptoquivalines and meroditerpenes isolated from the marine-derived fungi Neosartorya paulistensis, N. laciniosa, N. tsunodae, and the soil fungi N. fischeri and N. siamensis. Mar. Drugs 2014, 12, 822–839. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Dominated Biological Activity, Pa * | Additional Activity, Pa * |
|---|---|---|
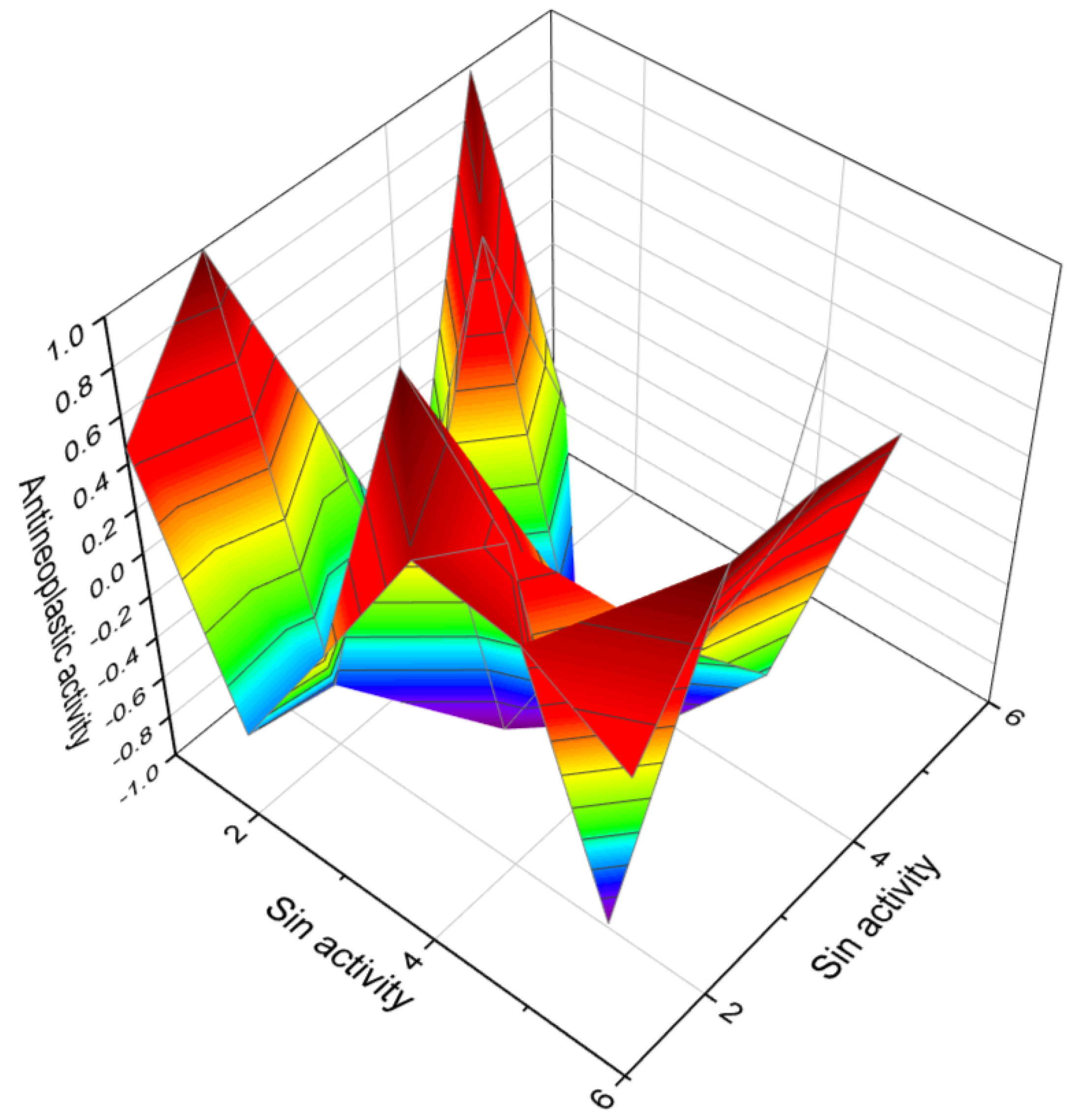
| 1 | Antineoplastic (0.920) Cytostatic (0.699) Apoptosis agonist (0.688) | Anti-leukemic (0.652) Antimitotic (0.598) |
| 2 | Antineoplastic (0.923) Cytostatic (0.827) Apoptosis agonist (0.827) | Anti-leukemic (0.706) Antimitotic (0.682) |
| 3 | Antineoplastic (0.930) Apoptosis agonist (0.900) | Anti-leukemic (0.872) Antimitotic (0.784) |
| 4 | Antineoplastic (0.878) | Antiprotozoal (Plasmodium) (0.742) |
| 5 | Anti-inflammatory (0.884) Antiviral (Arbovirus) (0.704) Antiviral (Picornavirus) (0.691) | Antineoplastic (0.865) Leukotriene synthesis inhibitor (0.605) DNA synthesis inhibitor (0.547) |
| 6 | Apoptosis agonist (0.827) Antineoplastic (0.806) | Antimitotic (0.704) |
| 7 | Antineoplastic (0.837) | Anti-eczematic (0.853) |
| 8 | Antineoplastic (0.888) Apoptosis agonist (0.606) | Antiprotozoal (Plasmodium) (0.764) Antifungal (0.601) |
| 9 | Antineoplastic (0.837) | Anti-eczematic (0.853) |
| 10 | Antineoplastic (0.951) Apoptosis agonist (0.647) Anti-metastatic (0.581) | Anti-psoriatic (0.698) Anti-eczematic (0.639) Anti-allergic (0.658) |
| 11 | Antineoplastic (0.937) Apoptosis agonist (0.710) | Angiogenesis inhibitor (0.903) Antiprotozoal (0.609) |
| 12 | Antineoplastic (0.920) Cytostatic (0.699) | Angiogenesis inhibitor (0.636) Antimitotic (0.598) |
| 13 | Genital warts treatment (0.923) Immunosuppressant (0.692) Atherosclerosis treatment (0.656) | Antineoplastic (0.918) Apoptosis agonist (0.839) Prostate cancer treatment (0.579) |
| 14 | Antineoplastic (0.876) Apoptosis agonist (0.819) | Genital warts treatment (0.843) Immunosuppressant (0.720) |
| 15 | Antineoplastic (0.810) | Antifungal (0.746) |
| 16 | Antineoplastic (0.926) Apoptosis agonist (0.896) | Anti-inflammatory (0.870) |
| 17 | Antineoplastic (0.893) | Antiprotozoal (Plasmodium) (0.750) |
| 18 | Antiprotozoal (Leishmania) (0.896) | Antineoplastic (0.829) |
| 19 | Antineoplastic (0.902) Apoptosis agonist (0.871) | Cardiotonic (0.727) Immunosuppressant (0.690) |
| 20 | Apoptosis agonist (0.836) Antineoplastic (0.783) | Autoimmune disorders treatment (0.692) Anti-leukemic (0.680) |
| 21 | Antineoplastic (0.834) | Anti-eczematic (0.736) |
| 22 | Antifungal (0.940) Antibacterial (0.760) | Antineoplastic (0.892) Apoptosis agonist (0.579) |
| 23 | Antineoplastic (0.974) | Anti-eczematic (0.821) |
| 24 | Antineoplastic (0.889) Apoptosis agonist (0.753) | Anti-leukemic (0.714) Anti-metastatic (0.563) |
| No. | Dominated Biological Activity, Pa * | Additional Activity, Pa * |
|---|---|---|
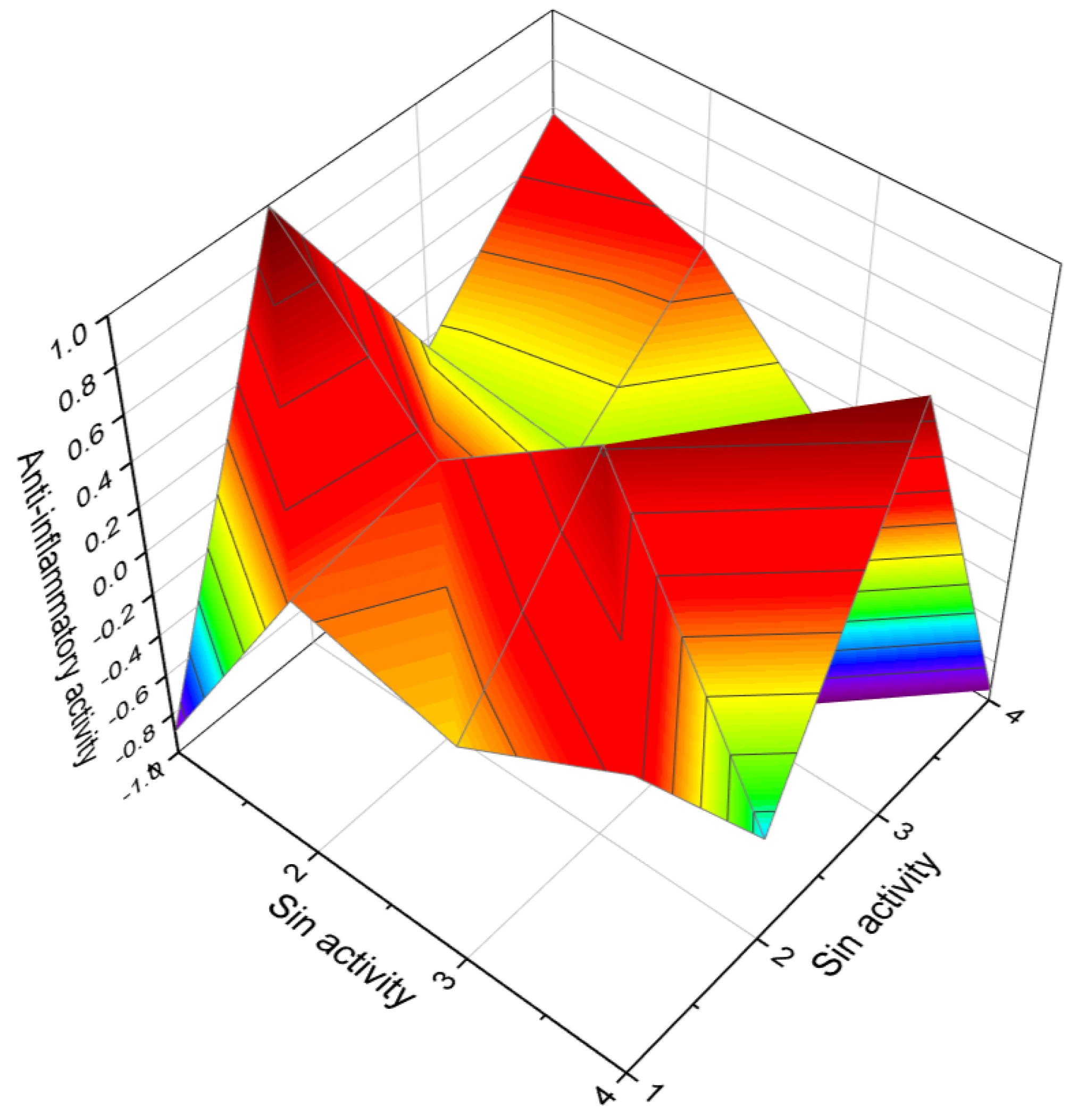
| 25 | Antineoplastic (0.902) Apoptosis agonist (0.622) | Anti-leukemic (0.831) Anti-emphysemic (0.745) |
| 26 | Antineoplastic (0.867) Apoptosis agonist (0.581) | Anti-leukemic (0.634) Anti-emphysemic (0.615) |
| 27 | Antineoplastic (0.863) Apoptosis agonist (0.789) | Antifungal (0.812) Antibacterial (0.763) |
| 28 | Antineoplastic (0.953) Anti-metastatic (0.612) | Anti-inflammatory (0.856) Autoimmune disorders treatment (0.820) |
| 29 | Antineoplastic (0.937) Apoptosis agonist (0.710) | Angiogenesis inhibitor (0.903) Antiarthritic (0.720) |
| 30 | Antineoplastic (0.924) Antimitotic (0.842) Apoptosis agonist (0.805) | Antieczematic (0.791) Antipsoriatic (0.647) |
| 31 | Apoptosis agonist (0.859) Antineoplastic (0.800) | Anti-parasitic (0.732) Antifungal (0.649) |
| 32 | Antineoplastic (0.949) | Anti-inflammatory (0.836) |
| 33 | Antineoplastic (0.911) Apoptosis agonist (0.821) | Antimitotic (0.856) |
| 34 | Antineoplastic (0.878) | Antiprotozoal (Plasmodium) (0.742) |
| 35 | Antineoplastic (0.923) Apoptosis agonist (0.885) | Anti-glaucomic (0.796) Cognition disorders treatment (0.778) |
| 36 | Antineoplastic (0.931) Apoptosis agonist (0.889) | Anti-psoriatic (0.812) Anti-eczematic (0.738) |
| 37 | Antineoplastic (0.935) Apoptosis agonist (0.805) | Anti-allergic (0.727) Anti-asthmatic (0.693) |
| 38 | Antineoplastic (0.937) Apoptosis agonist (0.710) | Angiogenesis inhibitor (0.903) Antimetastatic (0.599) |
| 39 | Antineoplastic (0.935) Apoptosis agonist (0.805) | Antiallergic (0.727) Antiasthmatic (0.693) |
| 40 | Anti-inflammatory (0.889) Immunosuppressant (0.672) | Antineoplastic (0.819) Apoptosis agonist (0.771) |
| 41 | Anti-inflammatory (0.920) Anti-leukemic (0.595) | Antineoplastic (0.898) Apoptosis agonist (0.869) |
| 42 | Antineoplastic (solid tumors) (0.967) Antineoplastic (0.918) | Antidiabetic symptomatic (0.687) Hypolipemic (0.661) |
| 43 | Antineoplastic (0.948) Apoptosis agonist (0.914) | Antieczematic (0.942) Antipsoriatic (0.788) |
| 44 | Antineoplastic (0.916) | Immunosuppressant (0.599) |
| No. | Dominated Biological Activity, Pa * | Additional Activity, Pa * |
|---|---|---|
| 45 | Calcium absorption stimulator (0.914); Immunosuppressant (0.857) | Antifungal (0.699); Angiogenesis inhibitor (0.690) |
| 46 | Anti-eczematic (0.907); Hepatoprotectant (0.829); Anti-psoriatic (0.660) | Chemopreventive (0.832); Antineoplastic (0.795); Apoptosis agonist (0.652) |
| 47 | Anti-eczematic (0.891); Hepatoprotectant (0.786); Anti-psoriatic (0.643) | Chemopreventive (0.849); Antineoplastic (0.776); Apoptosis agonist (0.644) |
| 48 | Anti-eczematic (0.883); Hepatoprotectant (0.768); Anti-psoriatic (0.639) | Chemopreventive (0.832); Antineoplastic (0.780); Apoptosis agonist (0.661) |
| 49 | Antineoplastic (0.844); Chemopreventive (0.821) | Anti-eczematic (0.829); Antipsoriatic (0.618) |
| 50 | Apoptosis agonist (0.867); Antineoplastic (0.859) Chemopreventive (0.798) | Anti-eczematic (0.853); Antipsoriatic (0.684); Lipoprotein disorders treatment (0.656) |
| 51 | Hepatoprotectant (0.846); Immunosuppressant (0.733) | Antineoplastic (0.734); Apoptosis agonist (0.649) |
| 52 | Antineoplastic (0.730); Chemopreventive (0.718) | Antifungal (0.693) |
| 53 | Antineoplastic (0.730); Chemopreventive (0.718) | Antifungal (0.693) |
| 54 | Chemopreventive (0.861); Antineoplastic (0.839) | Antifungal (0.825); Antibacterial (0.676) |
| 55 | Antineoplastic (0.783); Apoptosis agonist (0.731) | Antifungal (0.785) |
| 56 | Antineoplastic (0.793); Apoptosis agonist (0.664) | Antifungal (0.791); Anti-inflammatory (0.740) |
| 57 | Antineoplastic (0.896); Apoptosis agonist (0.809) | Antifungal (0.822); Antibacterial (0.717) |
| 58 | Anti-inflammatory (0.856); Antifungal (0.732) | Antineoplastic (0.801); Apoptosis agonist (0.770) |
| 59 | Antineoplastic (0.906); Apoptosis agonist (0.737) | Antifungal (0.789); Anti-inflammatory (0.617) |
| 60 | Antineoplastic (0.919); Apoptosis agonist (0.762) | Anti-inflammatory (0.824) |
| 61 | Antineoplastic (0.935); Apoptosis agonist (0.892) | Anti-inflammatory (0.827) |
| 62 | Antineoplastic (0.789); Apoptosis agonist (0.674) | Antifungal (0.764); Anti-inflammatory (0.646) |
| 63 | Antineoplastic (0.880); Apoptosis agonist (0.831) | Anti-inflammatory (0.869); Antifungal (0.804) |
| 64 | Anti-hypercholesterolemic (0.908) Cholesterol synthesis inhibitor (0.744) | Antifungal (0.843); Antiinflammatory (0.807) |
| 65 | Antineoplastic (0.916); Apoptosis agonist (0.794) Prostate cancer treatment (0.532) | Anti-hypercholesterolemic (0.629); Atherosclerosis treatment (0.550) |
| 66 | Antineoplastic (0.774); Antiprotozoal (0.663) | |
| 67 | Antineoplastic (0.866); Antiprotozoal (Plasmodium) (0.651); Antineoplastic (renal cancer) (0.518) | Anti-inflammatory (0.673); Antifungal (0.663) |
| 68 | Antineoplastic (0.849); Apoptosis agonist (0.678) |
| No. | Dominated Biological Activity, Pa * | Additional Activity, Pa * |
|---|---|---|
| 69 | Antineoplastic (0.958) Apoptosis agonist (0.946) Prostate cancer treatment (0.769) | Respiratory analeptic (0.899) |
| 70 | Proliferative diseases treatment (0.932) Antineoplastic (0.926) Apoptosis agonist (0.864) | Respiratory analeptic (0.903) |
| 71 | Antineoplastic (0.923) Proliferative diseases treatment (0.890) Apoptosis agonist (0.885) | Respiratory analeptic (0.887) |
| 72 | Antineoplastic (0.915) Apoptosis agonist (0.893) Proliferative diseases treatment (0.748) | Anti-inflammatory (0.882); Respiratory analeptic (0.866) |
| 73 | Antineoplastic (0.928) Apoptosis agonist (0.911) Prostate cancer treatment (0.628) | Anti-hypercholesterolemic (0.815); Atherosclerosis treatment (0.660) |
| 74 | Antineoplastic (0.912); Apoptosis agonist (0.890) | Anti-inflammatory (0.869) |
| 75 | Antineoplastic (0.886); Apoptosis agonist (0.846) | Antifungal (0.751); Antibacterial (0.683) |
| 76 | Anti-inflammatory (0.879); Antifungal (0.803) | Antineoplastic (0.801); Apoptosis agonist (0.670) |
| 77 | Antineoplastic (0.869); Apoptosis agonist (0.769) | |
| 78 | Antineoplastic (0.886); Apoptosis agonist (0.864) |
| No. | Dominated Biological Activity, Pa * | Additional Activity, Pa * |
|---|---|---|
| 79 | Apoptosis agonist (0.860); Antineoplastic (0.853) | Hepatic disorders treatment (0.817) |
| 80 | Antineoplastic (0.814); Antimitotic (0.566) | Angiogenesis inhibitor (0.690) |
| 81 | Anti-hypercholesterolemic (0.916) Cholesterol synthesis inhibitor (0.685) Atherosclerosis treatment (0.633) | Antifungal (0.835); Antibacterial (0.693) |
| 82 | Antineoplastic (0.957); Apoptosis agonist (0.788) | Antifungal (0.679); Antibacterial (0.658) |
| 83 | Antineoplastic (0.925); Apoptosis agonist (0.609) | Signal transduction pathways inhibitor (0.870) |
| 84 | Antineoplastic (0.802); Apoptosis agonist (0.719) | Spasmolytic. urinary (0.609) |
| 85 | Anti-hypercholesterolemic (0.916); Hypolipemic (0.827); Cholesterol synthesis inhibitor (0.685); Atherosclerosis treatment (0.633) | Antifungal (0.835); Antibacterial (0.693) |
| 86 | Antineoplastic (0.957); Apoptosis agonist (0.788) | Antifungal (0.679); Antibacterial (0.658) |
| 87 | Antineoplastic (0.925); Apoptosis agonist (0.609) | Signal transduction pathways inhibitor (0.870) |
| 88 | Genital warts treatment (0.826); Antieczematic (0.743) | |
| 89 | Anti-hypercholesterolemic (0.916); Cholesterol synthesis inhibitor (0.685); Atherosclerosis treatment (0.633) | Antifungal (0.835); Antibacterial (0.693) |
| 90 | Anti-eczematic (0.922); Antipsoriatic (0.639) | Antineoplastic (0.775); Apoptosis agonist (0.742) |
| 91 | Anti-eczematic (0.922); Cell adhesion molecule inhibitor (0.864); Antipsoriatic (0.645) | Apoptosis agonist (0.776); Antineoplastic (0.765) |
| 92 | Antineoplastic (0.903); Apoptosis agonist (0.693) | Respiratory analeptic (0.817) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dembitsky, V.M. Bioactive Diepoxy Metabolites and Highly Oxygenated Triterpenoids from Marine and Plant-Derived Bacteria and Fungi. Microbiol. Res. 2024, 15, 66-90. https://doi.org/10.3390/microbiolres15010005
Dembitsky VM. Bioactive Diepoxy Metabolites and Highly Oxygenated Triterpenoids from Marine and Plant-Derived Bacteria and Fungi. Microbiology Research. 2024; 15(1):66-90. https://doi.org/10.3390/microbiolres15010005
Chicago/Turabian StyleDembitsky, Valery M. 2024. "Bioactive Diepoxy Metabolites and Highly Oxygenated Triterpenoids from Marine and Plant-Derived Bacteria and Fungi" Microbiology Research 15, no. 1: 66-90. https://doi.org/10.3390/microbiolres15010005
APA StyleDembitsky, V. M. (2024). Bioactive Diepoxy Metabolites and Highly Oxygenated Triterpenoids from Marine and Plant-Derived Bacteria and Fungi. Microbiology Research, 15(1), 66-90. https://doi.org/10.3390/microbiolres15010005