
The Effect of Heavy Metals on Microbial Communities in Industrial Soil in the Area of Piekary Śląskie and Bukowno (Poland)


Abstract
:1. Introduction
2. Material and Methods
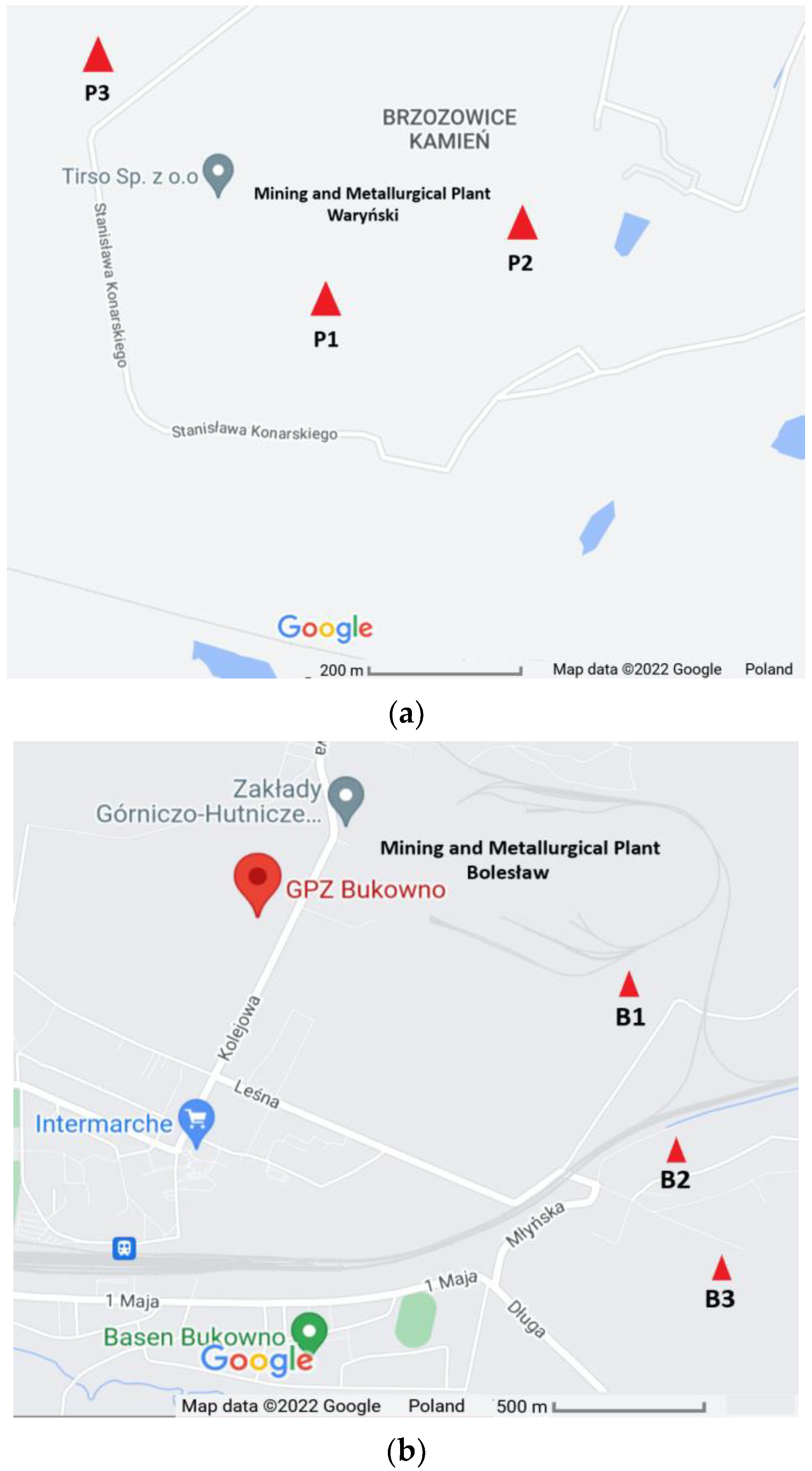
2.1. Soil Sample Collection
2.2. Physicochemical Analysis
2.3. Culturable Bacteria Enumeration
2.4. Total Bacteria Enumeration
2.5. Total Microbial Activity Measurement
2.6. Metallotolerant Bacteria Isolation and Identification
2.7. Minimal Inhibitory Concentration (MIC) of HMs
2.8. Microbial Community Structure
2.9. Microbial Community’s Physiological Profile
2.10. Genetic Biodiversity of Microbial Community Structure
2.11. CadA, CzcA, and NccA Genes Detection
2.12. Statistical Analysis
3. Results
3.1. Physicochemical Analysis of Soil Samples
3.2. Enumeration of Metal Tolerant Microorganisms in Soil Samples
3.3. Bacteria Isolated from Soil Samples
3.4. Microorganisms Structure and Metabolic Activity
3.5. Genes Encoding Mechanisms of HM Tolerance
3.6. Kendall Tau Correlation
4. Discussion
4.1. Physicochemical Characterization of Soils
4.2. The Influence of HMs on Bacterial Counts
4.3. Microbial Community Structure and Activity in HM-Contaminated Soils
4.4. Isolated Bacteria Characteristics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mitra, S.; Chakraborty, A.J.; Tareq, A.M.; Emran, T.B.; Nainu, F.; Khusro, A.; Idris, A.M.; Khandaker, M.U.; Osman, H.; Alhumaydhi, F.A.; et al. Impact of heavy metals on the environment and human health: Novel therapeutic insights to counter the toxicity. J. King Saud Univ. Sci. 2022, 34, 101865. [Google Scholar] [CrossRef]
- Li, C.; Zhou, K.; Qin, W.; Tian, C.; Qi, M.; Yan, X.; Han, W. A Review on Heavy Metals Contamination in Soil: Effects, Sources, and Remediation Techniques. Soil Sediment. Contam. 2019, 28, 380–394. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Ilahi, I. Environmental chemistry and ecotoxicology of hazardous heavy metals: Environmental persistence, toxicity, and bioaccumulation. J. Chem. 2019, 2019, 6730305. [Google Scholar] [CrossRef]
- Rai, P.K.; Lee, S.S.; Zhang, M.; Tsang, Y.F.; Kim, K.H. Heavy metals in food crops: Health risks, fate, mechanisms, and management. Environ. Int. 2019, 125, 365–385. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Habib, M.; Kakavand, S.N.; Zahid, Z.; Zahra, N.; Sharif, R.; Hasanuzzaman, M. Phytoremediation of cadmium: Physiological, biochemical, and molecular mechanisms. Biology 2020, 9, 177. [Google Scholar] [CrossRef] [PubMed]
- Ayangbenro, A.S.; Babalola, O.O. A new strategy for heavy metal polluted environments: A review of microbial biosorbents. Int. J. Environ. Res. Public Health 2017, 14, 94. [Google Scholar] [CrossRef] [PubMed]
- Bouchez, T.; Blieux, A.L.; Dequiedt, S.; Domaizon, I.; Dufresne, A.; Ferreira, S.; Godon, J.J.; Hellal, J.; Joulian, C.; Quaiser, A.; et al. Molecular microbiology methods for environmental diagnosis. Environ. Chem. Lett. 2016, 14, 423–441. [Google Scholar] [CrossRef]
- Giller, K.E.; Witter, E.; Mcgrath, S.P. Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: A review. Soil Biol. Biochem. 1998, 30, 1389–1414. [Google Scholar] [CrossRef]
- Khan, S.; Hesham, A.E.L.; Qiao, M.; Rehman, S.; He, J.Z. Effects of Cd and Pb on soil microbial community structure and activities. Environ. Sci. Pollut. Res. 2010, 17, 288–296. [Google Scholar] [CrossRef]
- Wang, Y.P.; Shi, J.Y.; Wang, H.; Lin, Q.; Chen, X.C.; Chen, Y.X. The influence of soil heavy metals pollution on soil microbial biomass, enzyme activity, and community composition near a copper smelter. Ecotoxicol. Environ. Saf. 2007, 67, 75–81. [Google Scholar] [CrossRef]
- Wang, X.; Gao, P.; Li, D.; Liu, J.; Yang, N.; Gu, W.; He, X.; Tang, W. Risk assessment for and microbial community changes in Farmland soil contaminated with heavy metals and metalloids. Ecotoxicol. Environ. Saf. 2019, 185, 109685. [Google Scholar] [CrossRef] [PubMed]
- Bååth, E. Effects of heavy metals in soil on microbial processes and populations (a review). Water Air Soil Pollut. 1989, 47, 335–379. [Google Scholar] [CrossRef]
- Chen, J.; He, F.; Zhang, X.; Sun, X.; Zheng, J.; Zheng, J. Heavy metal pollution decreases microbial abundance, diversity and activity within particle-size fractions of a paddy soil. FEMS Microbiol. Ecol. 2014, 87, 164–181. [Google Scholar] [CrossRef]
- Müller, A.K.; Westergaard, K.; Christensen, S.; Sørensen, S.J. The effect of long-term mercury pollution on the soil microbial community. FEMS Microbiol. Ecol. 2001, 36, 11–19. [Google Scholar] [CrossRef]
- Pennanen, T.; Frostegård, Å.; Fritze, H.; Bååth, E. Phospholipid fatty acid composition and heavy metal tolerance of soil microbial communities along two heavy metal-polluted gradients in coniferous forests. Appl. Environ. Microbiol. 1996, 62, 420–428. [Google Scholar] [CrossRef]
- Jin, Y.; Luan, Y.; Ning, Y.; Wang, L. Effects and mechanisms of microbial remediation of heavy metals in soil: A critical review. Appl. Sci. 2018, 8, 1336. [Google Scholar] [CrossRef]
- Guo, H.; Nasir, M.; Lv, J.; Dai, Y.; Gao, J. Understanding the variation of microbial community in heavy metals contaminated soil using high throughput sequencing. Ecotoxicol. Environ. Saf. 2017, 144, 300–306. [Google Scholar] [CrossRef]
- Zeng, X.Y.; Li, S.W.; Leng, Y.; Kang, X.H. Structural and functional responses of bacterial and fungal communities to multiple heavy metal exposure in arid loess. Sci. Total Environ. 2020, 723, 138081. [Google Scholar] [CrossRef]
- Tipayno, S.C.; Truu, J.; Samaddar, S.; Truu, M.; Preem, J.K.; Oopkaup, K.; Espenberg, M.; Chatterjee, P.; Kang, Y.; Kim, K.; et al. The bacterial community structure and functional profile in the heavy metal contaminated paddy soils, surrounding a nonferrous smelter in South Korea. Ecol. Evol. 2018, 8, 6157–6168. [Google Scholar] [CrossRef] [PubMed]
- Masum, S.A.; Islam, M.S. A case study on predicting the environmental impacts of untreated effluent generated from Tannery industrial estate in Dhaka, Bangladesh. Geogr. Environ. Sustain. 2020, 13, 22–31. [Google Scholar] [CrossRef]
- Masum, S.A.; Thomas, H.R. Effects of microbial spatial distribution on organic biodegradation and immobilization of trace metals in co-contaminated soils. Comput. Geotech. 2021, 133, 104063. [Google Scholar] [CrossRef]
- Wood, J.L.; Liu, W.; Tang, C.; Franks, A.E. Microorganisms in heavy metal bioremediation: Strategies for applying microbial-community engineering to remediate soils. AIMS Bioeng. 2016, 3, 211–229. [Google Scholar] [CrossRef]
- Szrek, D.; Bajda, T.; Manecki, M. A comparative study of the most effective amendment for Pb, Zn and Cd immobilization in contaminated soils. J. Environ. Sci. Health—Part A Toxic/Hazard. Subst. Environ. Eng. 2011, 46, 1491–1502. [Google Scholar] [CrossRef] [PubMed]
- Kucharski, R.; Sas-Nowosielska, A.; Małkowski, E.; Japenga, J.; Kuperberg, J.M.; Pogrzeba, M.; Krzyzak, J. The use of indigenous plant species and calcium phosphate for the stabilization of highly metal-polluted sites in southern Poland. Plant Soil 2005, 273, 291–305. [Google Scholar] [CrossRef]
- Oleksynowa, K.; Tokaj, J.; Jakubiec, J. Guide to Exercies in Soil Science and Geology; Wydawnictwo AR: Kraków, Poland, 1991. [Google Scholar]
- Edwards, U.; Rogall, T.; Blockerl, H.; Emde, M.; Bottger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [PubMed]
- Pacwa-Płociniczak, M.; Płaza, G.A.; Poliwoda, A.; Piotrowska-Seget, Z. Characterization of hydrocarbon-degrading and biosurfactant-producing Pseudomonas sp. P-1 strain as a potential tool for bioremediation of petroleum-contaminated soil. Environ. Sci. Pollut. Res. 2014, 21, 9385–9395. [Google Scholar] [CrossRef]
- Sułowicz, S.; Cycoń, M.; Piotrowska-Seget, Z. Non-target impact of fungicide tetraconazole on microbial communities in soils with different agricultural management. Ecotoxicology 2016, 25, 1047–1060. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Frostegard, A.; Tunlid, A.; Baath, E. Phospholipid fatty acid composition, biomass, and activity of microbial communities from two soil types experimentally exposed to different heavy metals. Appl. Environ. Microbiol. 1993, 59, 3605–3617. [Google Scholar] [CrossRef]
- Willers, C.; Jansen van Rensburg, P.J.; Claassens, S. Phospholipid fatty acid profiling of microbial communities-a review of interpretations and recent applications. J. Appl. Microbiol. 2015, 119, 1207–1218. [Google Scholar] [CrossRef]
- Kroppenstedt, R.M. Fatty Acid and Menaquinone Analysis of Actinomycetes and Related Organisms. In Chemical Methods in Bacterial Systematics; Goodfellow, M., Minnikin, D.E., Eds.; Academic Press: London, UK, 1985; pp. 173–199. [Google Scholar]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Use and misuse of PLFA measurements in soils. Soil Biol. Biochem. 2011, 43, 1621–1625. [Google Scholar] [CrossRef]
- Magdalena, F.; Stefania, J.T. Agricultural utilisation of dairy sewage sludge: Its effect on enzymatic activity and microorganisms of the soil environment. Afr. J. Microbiol. Res. 2011, 5, 1755–1762. [Google Scholar] [CrossRef]
- Muyzer, G.; De Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; Smalla, K. Application of denaturing gradient gel electrophoresis (DGGE) and temperature gradient gel electrophoresis (TGGE) in microbial ecology. Antonie Van Leeuwenhoek 1998, 73, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Ishii, K.; Fukui, M.; Takii, S. Microbial succession during a composting process as evaluated by denaturing gradient gel electrophoresis analysis. J. Appl. Microbiol. 2000, 89, 768–777. [Google Scholar] [CrossRef]
- Cycoń, M.; Markowicz, A.; Borymski, S.; Wójcik, M.; Piotrowska-Seget, Z. Imidacloprid induces changes in the structure, genetic diversity and catabolic activity of soil microbial communities. J. Environ. Manag. 2013, 131, 55–65. [Google Scholar] [CrossRef]
- Oger, C.; Mahillon, J.; Petit, F. Distribution and diversity of a cadmium resistance (cadA) determinant and occurrence of IS257 insertion sequences in Staphylococcal bacteria isolated from a contaminated estuary (Seine, France). FEMS Microbiol. Ecol. 2003, 43, 173–183. [Google Scholar] [CrossRef]
- Karelová, E.; Harichová, J.; Stojnev, T.; Pangallo, D.; Ferianc, P. The isolation of heavy-metal resistant culturable bacteria and resistance determinants from a heavy-metal-contaminated site. Biologia (Bratisl) 2011, 66, 18–26. [Google Scholar] [CrossRef]
- Nies, D.H. Efflux-mediated heavy metal resistance in prokaryotes. FEMS Microbiol. Rev. 2003, 27, 313–339. [Google Scholar] [CrossRef]
- Kim, R.Y.; Yoon, J.K.; Kim, T.S.; Yang, J.E.; Owens, G.; Kim, K.R. Bioavailability of heavy metals in soils: Definitions and practical implementation—a critical review. Environ. Geochem. Health 2015, 37, 1041–1061. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Z.; Wang, S.; Luo, S.; Ren, L.; Liang, Y.; Yang, R.; Li, Y.; Zhang, Y.; Deng, S.; Zou, L.; et al. Significant Impacts of Both Total Amount and Availability of Heavy Metals on the Functions and Assembly of Soil Microbial Communities in Different Land Use Patterns. Front. Microbiol. 2019, 10, 2293. [Google Scholar] [CrossRef]
- Guo, X.; Zhao, G.; Zhang, G.; He, Q.; Wei, Z.; Zheng, W.; Qian, T.; Wu, Q. Effect of mixed chelators of EDTA, GLDA, and citric acid on bioavailability of residual heavy metals in soils and soil properties. Chemosphere 2018, 209, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Nicol, G.W.; Leininger, S.; Schleper, C.; Prosser, J.I. The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environ. Microbiol. 2008, 10, 2966–2978. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Yu, M.; Tang, C.; Zhang, L.; Muhammad, N.; Zhao, H.; Feng, J.; Yu, L.; Xu, J. The negative impact of cadmium on nitrogen transformation processes in a paddy soil is greater under non-flooding than flooding conditions. Environ. Int. 2019, 129, 451–460. [Google Scholar] [CrossRef]
- Smolders, E.; Brans, K.; Coppens, F.; Merckx, R. Potential nitrification rate as a tool for screening toxicity in metal-contaminated soils. Environ. Toxicol. Chem. 2001, 20, 2469–2474. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Azarbad, H.; van Straalen, N.M.; Laskowski, R.; Nikiel, K.; Röling, W.F.M.; Niklińska, M. Susceptibility to additional stressors in metal-tolerant soil microbial communities from two pollution gradients. Appl. Soil Ecol. 2016, 98, 233–242. [Google Scholar] [CrossRef]
- Song, J.; Shen, Q.; Wang, L.; Qiu, G.; Shi, J.; Xu, J.; Brookes, P.C.; Liu, X. Effects of Cd, Cu, Zn and their combined action on microbial biomass and bacterial community structure. Environ. Pollut. 2018, 243, 510–518. [Google Scholar] [CrossRef]
- Díaz-Raviña, M.; Bååth, E. Development of metal tolerance in soil bacterial communities exposed to experimentally increased metal levels. Appl. Environ. Microbiol. 1996, 62, 2970–2977. [Google Scholar] [CrossRef]
- Borymski, S.; Cycon, M.; Beckmann, M.; Mur, L.A.J.; Piotrowska-Seget, Z. Plant species and heavy metals affect biodiversity of microbial communities associated with metal-tolerant plants in metalliferous soils. Front. Microbiol. 2018, 9, 1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajapaksha, R.M.C.P.; Tobor-Kapłon, M.A.; Bååth, E. Metal toxicity affects fungal and bacterial activities in soil differently. Appl. Environ. Microbiol. 2004, 70, 2966–2973. [Google Scholar] [CrossRef] [PubMed]
- Bååth, E.; Díaz-Raviña, M.; Bakken, L.R. Microbial biomass, community structure and metal tolerance of a naturally Pb-enriched forest soil. Microb. Ecol. 2005, 50, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Seshadri, B.; Bolan, N.; Sarkar, B.; Ok, Y.S.; Zhang, W.; Rumpel, C.; Sparks, D.; Farrell, M.; Hall, T.; et al. Microbial functional diversity and carbon use feedback in soils as affected by heavy metals. Environ. Int. 2019, 125, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yao, J.; Si, Y.; Chen, H.; Russel, M.; Chen, K.; Qian, Y.; Zaray, G.; Bramanti, E. Short-time effect of heavy metals upon microbial community activity. J. Hazard. Mater. 2010, 173, 510–516. [Google Scholar] [CrossRef]
- Azarbad, H.; Niklińska, M.; Van Gestel, C.A.M.; Van Straalen, N.M.; Röling, W.F.M.; Laskowski, R. Microbial community structure and functioning along metal pollution gradients. Environ. Toxicol. Chem. 2013, 32, 1992–2002. [Google Scholar] [CrossRef]
- Chodak, M.; Gołebiewski, M.; Morawska-Płoskonka, J.; Kuduk, K.; Niklińska, M. Diversity of microorganisms from forest soils differently polluted with heavy metals. Appl. Soil Ecol. 2013, 64, 7–14. [Google Scholar] [CrossRef]
- Klimek, B.; Sitarz, A.; Choczyński, M.; Niklińska, M. The Effects of Heavy Metals and Total Petroleum Hydrocarbons on Soil Bacterial Activity and Functional Diversity in the Upper Silesia Industrial Region (Poland). Water Air Soil Pollut. 2016, 227, 265. [Google Scholar] [CrossRef]
- Bodor, A.; Bounedjoum, N.; Vincze, G.E.; Erdeiné Kis, Á.; Laczi, K.; Bende, G.; Szilágyi, Á.; Kovács, T.; Perei, K.; Rákhely, G. Challenges of unculturable bacteria: Environmental perspectives. Rev. Environ. Sci. Biotechnol. 2020, 19, 1–22. [Google Scholar] [CrossRef]
- Pacwa-Płociniczak, M.; Płociniczak, T.; Yu, D.; Kurola, J.M.; Sinkkonen, A.; Piotrowska-Seget, Z.; Romantschuk, M. Effect of Silene vulgaris and Heavy Metal Pollution on Soil Microbial Diversity in Long-Term Contaminated Soil. Water Air Soil Pollut. 2018, 229, 13. [Google Scholar] [CrossRef]
- Navarro-Noya, Y.E.; Jan-Roblero, J.; González-Chávez, D.C.; Hernández-Gama, R.; Hernández-Rodríguez, C. Bacterial communities associated with the rhizosphere of pioneer plants (Bahia xylopoda and Viguiera linearis) growing on heavy metals-contaminated soils. Antonie Van Leeuwenhoek 2010, 97, 335–349. [Google Scholar] [CrossRef]
- Pereira, L.B.; Vicentini, R.; Ottoboni, L.M.M. Changes in the bacterial community of soil from a neutral mine drainage channel. PLoS ONE 2014, 9, e96605. [Google Scholar] [CrossRef] [PubMed]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [PubMed]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; El-Enshasy, H.A.; Dailin, D.J.; Suriani, N.L. Recent Understanding of Soil Acidobacteria and Their Ecological Significance: A Critical Review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef] [PubMed]
- Kielak, A.M.; Barreto, C.C.; Kowalchuk, G.A.; van Veen, J.A.; Kuramae, E.E. The ecology of Acidobacteria: Moving beyond genes and genomes. Front. Microbiol. 2016, 7, 744. [Google Scholar] [CrossRef] [PubMed]
- Hartman, W.H.; Richardson, C.J.; Vilgalys, R.; Bruland, G.L. Environmental and anthropogenic controls over bacterial communities in wetland soils. Proc. Natl. Acad. Sci. USA 2008, 105, 17842–17847. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef]
- Gołebiewski, M.; Deja-Sikora, E.; Cichosz, M.; Tretyn, A.; Wróbel, B. 16S rDNA pyrosequencing analysis of bacterial community in heavy metals polluted soils. Microb. Ecol. 2014, 67, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Tebo, B.M.; Davis, R.E.; Anitori, R.P.; Connell, L.B.; Schiffman, P.; Staudigel, H. Microbial communities in dark oligotrophic volcanic ice cave ecosystems of Mt. Erebus, Antarctica. Front. Microbiol. 2015, 6, 179. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Liao, R.; Zhang, X.X.; Wang, Y.; Wang, Z.; Shi, P.; Liu, B.; Li, A. Metagenomic insights into Cr(VI) effect on microbial communities and functional genes of an expanded granular sludge bed reactor treating high-nitrate wastewater. Water Res. 2015, 76, 43–52. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, T.; Wang, X.; Fu, J.; Zuo, M.; Yang, Y.; Yin, Z.; Wang, Z.; Tai, X.; Chang, G. Effects of heavy metals on bacterial community surrounding Bijiashan mining area located in northwest China. Open Life Sci. 2022, 17, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Tseng, S.C.; Liang, C.M.; Chia, T.; Ton, S.S. Changes in the composition of the soil bacterial community in heavy metal-contaminated farmland. Int. J. Environ. Res. Public Health 2021, 18, 8661. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Fan, J.; Zhu, W.; Amombo, E.; Lou, Y.; Chen, L.; Fu, J. Effect of heavy metals pollution on soil microbial diversity and bermudagrass genetic variation. Front. Plant Sci. 2016, 7, 755. [Google Scholar] [CrossRef] [PubMed]
- Mawang, C.I.; Azman, A.S.; Fuad, A.S.M.; Ahamad, M. Actinobacteria: An eco-friendly and promising technology for the bioaugmentation of contaminants. Biotechnol. Rep. 2021, 32, e00679. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.; Saez, J.M.; Davila Costa, J.S.; Colin, V.L.; Fuentes, M.S.; Cuozzo, S.A.; Benimeli, C.S.; Polti, M.A.; Amoroso, M.J. Actinobacteria: Current research and perspectives for bioremediation of pesticides and heavy metals. Chemosphere 2017, 166, 41–62. [Google Scholar] [CrossRef] [PubMed]
- Remenár, M.; Kamlárova, A.; Harichova, J.; Zámocky, M.; Ferianc, P. The heavy-metal resistance determinant of newly isolated bacterium from a nickel-contaminated soil in Southwest Slovakia. Pol. J. Microbiol. 2018, 67, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Harichová, J.; Karelová, E.; Pangallo, D.; Ferianc, P. Structure analysis of bacterial community and their heavy-metal resistance determinants in the heavy-metal-contaminated soil sample. Biology 2012, 67, 1038–1048. [Google Scholar] [CrossRef]
- Bruins, M.R.; Kapil, S.; Oehme, F.W. Microbial resistance to metals in the environment. Ecotoxicol. Environ. Saf. 2000, 45, 198–207. [Google Scholar] [CrossRef]
- Nies, D.H. The biological chemistry of the transition metal transportome of: Cupriavidus metallidurans. Metallomics 2016, 8, 481–507. [Google Scholar] [CrossRef]
- Guo, X.; Xie, C.; Wang, L.; Li, Q.; Wang, Y. Biodegradation of persistent environmental pollutants by Arthrobacter sp. Environ. Sci. Pollut. Res. 2019, 26, 8429–8443. [Google Scholar] [CrossRef]
- Han, J.I.; Choi, H.K.; Lee, S.W.; Orwin, P.M.; Kim, J.; LaRoe, S.L.; Kim, T.G.; O’Neil, J.; Leadbetter, J.R.; Lee, S.Y.; et al. Complete genome sequence of the metabolically versatile plant growth-promoting endophyte Variovorax paradoxus S110. J. Bacteriol. 2011, 193, 1183–1190. [Google Scholar] [CrossRef] [Green Version]
- Hu, N.; Zhao, B. Key genes involved in heavy-metal resistance in Pseudomonas putida CD2. FEMS Microbiol. Lett. 2007, 267, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Piekary Śląskie | Bukowno | |||||
|---|---|---|---|---|---|---|
| P1 | P2 | P3 | B1 | B2 | B3 | |
| Water content (%) | 26.21 ± 0.24 b | 29.48 ± 0.12 a | 8.54 ± 0.25 c | 5.01 ± 0.28 d | 5.34 ± 0.36 d | 1.56 ± 0.21 e |
| pH (H20) | 7.71 ± 0.03 b | 8.14 ± 0.02 a | 7.62 ± 0.01 c | 6.79 ± 0.01 e | 6.96 ± 0.01 d | 5.49 ± 0.01 f |
| pH (KCl) | 8 ± 0.03 b | 8.29 ± 0.01 a | 7.15 ± 0.01 c | 6.77 ± 0.01 e | 6.81 ± 0.01 d | 4.69 ± 0.01 f |
| pH (CaCl2) | 7.84 ± 0.01 b | 8.15 ± 0.01 a | 7.04 ± 0.01 c | 6.78 ± 0.01 e | 6.82 ± 0.01 d | 5.07 ± 0.01 f |
| Total N (mg/kg) | 829.9 ± 35.21 a | 780.8 ± 15.41 a | 997.8 ± 19.28 a | 747.97 ± 950.41 a | 215.63 ± 6.15 a | 151.2 ± 7.81 a |
| Organic matter (%) | 7.76 ± 0.02 b | 7.89 ± 0.06 b | 10.95 ± 0.55 a | 2.62 ± 0.04 c | 2.23 ± 0.07 c | 0.54 ± 0.04 d |
| Organic carbon (%) | 4.5 ± 0.01 b | 4.58 ± 0.03 b | 6.35 ± 0.32 a | 1.52 ± 0.02 c | 1.3 ± 0.04 c | 0.32 ± 0.02 d |
| P (%) | 0.83 ± 0.03 a | 1.14 ± 0.04 b | 0.38 ± 0.01 c | 0.2 ± 0.02 d | 0.29 ± 0.01 e | 0.24 ± 0.01de |
| Na (mg/kg) | 547.77 ± 23.24 a | 397.67 ± 63.28 b | 131.1 ± 5.11 c | 112.3 ± 10.87 cd | 38.6 ± 6.35 de | 15.17 ± 0.42 e |
| Mg (g/kg) | 37.486 ± 0.877 a | 14.965 ± 0.567 b | 5.865 ± 0.636 c | 19.257 ± 0.024 d | 3.420 ± 0.441 e | 0.277 ± 0.024 f |
| K (g/kg) | 0.708 ± 0.152 c | 1.093 ± 0.055 ab | 1.219 ± 0.059 a | 0.962 ± 0.111 b | 0.333 ± 0.076 d | 0.114 ± 0.056 d |
| Ca (g/kg) | 109.124 ± 2.746 a | 36.105 ± 3.384 b | 9.396 ± 0.425 d | 24.361 ± 1.402 c | 3.230 ± 0.285 e | 0.274 ± 0.049 e |
| Zn total (g/kg) | 1.208 ± 0.106 b | 1.282 ± 0.205 b | 1.035 ± 0.083 b | 1.8143 ± 0.00 a | 1.777 ± 0.00 b | 14.362 ± 0.00 b |
| Zn (H20) (mg/kg) | 0.49 ± 0.3 f | 1.14 ± 0.69 def | 3.47 ± 1.54 c | 19.38 ± 0.00 a | 1.98 ± 0.00 cd | 6.25 ± 0.00 b |
| Zn (CaCl2) (mg/kg) | 0.34 ± 0.12 e | 0.35 ± 0.11 e | 7.92 ± 0.88 d | 56.21 ± 0.00 a | 14.34 ± 0.00 b | 21.43 ± 0.00 c |
| Cd total (g/kg) | 0.293 ± 0.016 c | 1.215 ± 0.119 b | 0.0459 ± 0.006 d | 1.991 ± 0.00 a | 0.100 ± 0.00 d | 0.017 ± 0.00 d |
| Cd (H20) (mg/kg) | 0.06 ± 0.03 c | 0.14 ± 0.07 b | 0.03 ± 0.00 c | 0.51 ± 0.00 a | 0.02 ± 0.00 c | 0.01 ± 0.00 c |
| Cd (CaCl2) (mg/kg) | 0.05 ± 0.01 e | 0.11 ± 0.03 d | 0.24 ± 0.01 c | 3.32 ± 0.00 a | 0.49 ± 0.00 c | 0.21 ± 0.00 b |
| Ni total (mg/kg) | 3.94 ± 0.08 d | 3 ± 0.22 d | 0.62 ± 0.08 e | 54.16 ± 0.00 a | 44.63 ± 0.00 b | 15.25 ± 0.00 c |
| Ni (H20) (mg/kg) | 0.001 ± 0.001 e | 0.002 ± 0.001 de | 0.003 ± 0.001 d | 0.02 ± 0.00 c | 0.04 ± 0.00 a | 0.03 ± 0.00 b |
| Ni (CaCl2) (mg/kg) | 0.001 ± 0.002 a | 0.00 | 0.00 | 0.01 ± 0.00 c | 0.02 ± 0.00 b | 0.02 ± 0.00 b |
| Cu total (mg/kg) | 580.68 ± 59.12 b | 175.65 ± 13.7 c | 14.59 ± 0.69 d | 1116.87 ± 0.0 a | 75.81 ± 0.00 d | 0.00 ± 0.00 |
| Cu (H20) (mg/kg) | 0.04 ± 0.02 a | 0.05 ± 0.01 a | 0.03 ± 0.02 a | 0.00 ± 0.00 b | 0.02 ± 0.00 a | 0.01 ± 0.00 a |
| Cu (CaCl2) (mg/kg) | 0.02 ± 0.01 b | 0.05 ± 0.00 a | 0.01 ± 0.01b | 0.01 ± 0.00 b | 0.00 ± 0.00 b | 0.01 ± 0.00 b |
| Bacteria Count, | Piekary Śląskie | Bukowno | |||||
|---|---|---|---|---|---|---|---|
| (log10 CFU/g Dry Soil) | P1 | P2 | P3 | B1 | B2 | B3 | |
| OB | Total | 4.44 ± 0.09 ce | 4.79 ± 0.14 cd | 4.91 ± 0.03 c | 3.86 ± 0.04 f | 5.73 ± 0.43 a | 5.53 ± 0.16 ab |
| Cu | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Cd | 0.00 ± 0.00 | 0.00 ± 0.00 | 3.80 ± 0.14 a | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Ni | 0.00 ± 0.00 | 3.51 ± 0.17 a | 3.44 ± 0.35 ab | 2.48 ± 0.15 c | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Zn | 3.10 ±0.00 a | 3.11 ± 0.00 a | 3.67 ± 0.31 a | 3.59 ± 0.03 a | 4.58 ± 0.49 a | 2.83 ± 2.46 a | |
| HB (TSA) | Total | 4.35 ± 0.08 cde | 4.50 ± 0.04 ac | 4.64 ± 0.67 ab | 3.71 ± 0.09 f | 4.38 ± 0.10 acd | 4.73 ± 0.67 a |
| Cu | 3.98 ± 0.34 ce | 4.26 ± 0.06 c | 4.38 ± 0.65 ab | 3.28 ± 0.24 f | 4.56 ± 0.21 a | 4.19 ± 0.63 cd | |
| Cd | 3.10 ± 0.00 a | 3.59 ± 0.31a | 3.53 ± 0.52 a | 0.00 ± 0.00 | 0.00 ± 0.00 | 3.47 ± 0.54 a | |
| Ni | 3.99 ± 0.14 a | 4.33 ± 0.25 a | 4.42 ± 0.65 a | 3.87 ± 0.89 a | 5.12 ± 0.70 a | 4.66 ± 0.65 a | |
| Zn | 3.86 ± 0.28 a | 3.47 ± 0.39 a | 4.28 ± 0.63 a | 3.87 ± 2.06 a | 4.64 ± 0.83 a | 4.63 ± 0.65 a | |
| HB (Broth agar) | Total | 4.65 ± 0.10 ade | 4.77 ± 0.07 ac | 4.80 ± 0.24 ab | 4.65 ± 0.10 ef | 4.77 ± 0.07 ad | 4.80 ± 0.24 a |
| Cu | 4.14 ± 0.07 a | 3.99 ± 0.09 a | 4.03 ± 0.61 a | 4.14 ± 0.07 a | 3.99 ± 0.09 a | 4.03 ± 0.61 a | |
| Cd | 3.33 ± 0.40 a | 3.21 ± 0.17 a | 4.07 ± 0.22 a | 3.33 ± 0.40 a | 3.21 ± 0.17 a | 4.07 ± 0.22 a | |
| Ni | 3.99 ± 0.38 ade | 4.23 ± 0.26 ac | 4.16 ± 0.30 ad | 3.99 ± 0.38 ef | 4.23 ± 0.26 a | 4.16 ± 0.30 ab | |
| Zn | 3.77 ± 0.06 a | 3.65 ± 0.21 a | 4.27 ± 0.05 a | 3.77 ± 0.06 a | 3.65 ± 0.21 a | 4.27 ± 0.05 a | |
| RT-PCR | Total | 9.04 ± 0.08 bc | 9.11 ± 0.02 ab | 9.16 ± 0.03 a | 9.02 ± 0.03 bd | 9.06 ± 0.03 abd | 8.99 ± 0.03 cd |
| Method | Sampling Site | ||||||
|---|---|---|---|---|---|---|---|
| P1 | P2 | P3 | B1 | B2 | B3 | ||
| FDA (µg fluorescein/g dry soil) | 0.23 ± 0.08 b | 0.27 ± 0.06 b | 0.58 ± 0.1 a | 0.28 ± 0.1 b | 0.15 ± 0.04 b | 0.56 ± 0.07 a | |
| PLFA | Total, nmol PLFA/g dry soil | 2.83 ± 0.76 bc | 8.27 ± 0.95 ac | 12.64 ± 1.70 a | 5.88 ± 5.89 ac | 10.38 ± 1.26 ac | 77.62 ± 2.78 ac |
| Gram + (%) | 38.74 ± 3.75 a | 41.54 ± 0.46 a | 38.84 ± 0.25 a | 32.57 ± 19.47 a | 38.19 ± 2.36 a | 37.93 ± 3.01 a | |
| Gram − (%) | 46.04 ± 4.33 a | 50.49 ± 0.57 a | 47.14 ± 0.65 a | 59.84 ± 18.32 a | 47.75 ± 1.73 a | 47.33 ± 2.43 a | |
| Actinomycetes (%) | 15.21 ± 1.91 a | 4.65 ± 0.07 a | 9.23 ± 0.53 a | 6.71 ± 7.39 a | 8.23 ± 0.81 a | 8.43 ± 1.59 a | |
| Fungi (%) | 0.00 ± 0.00 | 3.31 ± 0.67 a | 4.80 ± 0.12 a | 0.88 ± 0.76 a | 5.83 ± 0.18 a | 6.31 ± 1.37 a | |
| BIOLOG | AWCD | 0.82 ± 0.05 ecd | 0.78 ± 0.04 cd | 1.15 ± 0.17 f | 0.09 ± 0.02 a | 0.82 ± 0.08 d | 1.38 ± 0.07 b |
| Carbohydrates (%) | 32.87 ± 7.14 b | 19.18 ± 1.53 a | 24.60 ± 0.10 b | 61.69 ± 3.17 b | 18.41 ± 2.74 b | 25.05 ± 1.99 b | |
| Carboxylic Acids (%) | 4.41 ± 3.81 cdf | 17.09 ± 0.46 c | 15.42 ± 1.13 cd | 7.67 ± 5.37 ce | 8.13 ± 3.66 b | 18.97 ± 2.07 a | |
| Amino acids (%) | 31.42 ± 3.52 cf | 35.48 ± 0.34 ce | 30.11 ± 1.85 c | 12.16 ± 1.82 cd | 41.28 ± 4.47 b | 26.94 ± 1.13 a | |
| Polymers (%) | 17.98 ± 0.66 c | 16.81 ± 3.78 cd | 16.45 ± 0.98 ce | 12.24 ± 6.99 cf | 18.90 ± 1.92 b | 13.52 ± 0.58 a | |
| Amines and amides (%) | 5.58 ± 3.97 bcd | 4.36 ± 0.76 bde | 5.99 ± 0.40 cb | 2.58 ± 3.00 bdf | 5.83 ± 0.66 b | 7.93 ± 3.40 a | |
| Miscellaneous (%) | 7.74 ± 5.31 c | 7.07 ± 3.19 ce | 7.44 ± 2.69 cd | 3.66 ± 2.56 cf | 7.45 ± 1.55 b | 7.60 ± 2.27 a | |
| DGGE | H’ | 1.02 ± 0.08 fde | 0.70±0.02 b | 0.95±0.04 d | 1.01±0.05 ed | 0.49±0.03 cb | 0.04 ± 0.00 a |
| Rs | 26 ± 0.00 a | 21 ±0.00 a | 21 ± 0.00 a | 18 ± 0.00 a | 24 ± 0.00 a | 24 ± 0.00 a | |
| EH | 0.31 ± 0.02 ed | 0.23 ± 0.01 cb | 0.31±0.01 dc | 0.35±0.02 fe | 0.23 ± 0.01 b | 0.01 ± 0.00 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jarosławiecka, A.K.; Piotrowska-Seget, Z. The Effect of Heavy Metals on Microbial Communities in Industrial Soil in the Area of Piekary Śląskie and Bukowno (Poland). Microbiol. Res. 2022, 13, 626-642. https://doi.org/10.3390/microbiolres13030045
Jarosławiecka AK, Piotrowska-Seget Z. The Effect of Heavy Metals on Microbial Communities in Industrial Soil in the Area of Piekary Śląskie and Bukowno (Poland). Microbiology Research. 2022; 13(3):626-642. https://doi.org/10.3390/microbiolres13030045
Chicago/Turabian StyleJarosławiecka, Anna Katarzyna, and Zofia Piotrowska-Seget. 2022. "The Effect of Heavy Metals on Microbial Communities in Industrial Soil in the Area of Piekary Śląskie and Bukowno (Poland)" Microbiology Research 13, no. 3: 626-642. https://doi.org/10.3390/microbiolres13030045
APA StyleJarosławiecka, A. K., & Piotrowska-Seget, Z. (2022). The Effect of Heavy Metals on Microbial Communities in Industrial Soil in the Area of Piekary Śląskie and Bukowno (Poland). Microbiology Research, 13(3), 626-642. https://doi.org/10.3390/microbiolres13030045