
Assessment of Bacteriocin-Antibiotic Synergy for the Inhibition and Disruption of Biofilms of Listeria monocytogenes and Vancomycin-Resistant Enterococcus

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteriocins Preparation
2.2. Determination of Minimum Inhibitory Concentrations (MIC) of Antibiotics against Planktonic Cells of L. monocytogenes ATCC15313 and E. faecium VRE19
2.3. Determination of Minimum Inhibitory Concentrations (MIC) of Bacteriocins against Planktonic Cells of Target Microorganisms
2.4. Molecular Detection of Vancomycin Resistance-Associated Genes of E. faecium VRE19
2.5. Biofilm Formation of L. monocytogenes ATCC15313 and E. faecium VRE19
2.6. Quantification of Biofilms by Crystal Violet Assay
2.7. Quantification of Viable Cells from Bacteriocin-Treated Biofilms of L. monocytogenes ATCC15313 and E. faecium VRE19 by Flow Cytometry
2.8. Determination of Metabolic Activity
2.9. Assessment of Synergistic Activities of Bacteriocins and Antibiotics against Biofilm Formation of L. monocytogenes ATCC15313 and E. faecium VRE19
2.10. Evaluation of Synergism of Bacteriocins and Antibiotics on the Biofilm Formed by L. monocytogenes ATCC15313 and E. faecium VRE19
3. Results
3.1. MIC of Antimicrobials Used
3.2. Molecular Detection of Vancomycin Resistance-Associated Genes in E. faecium VRE19
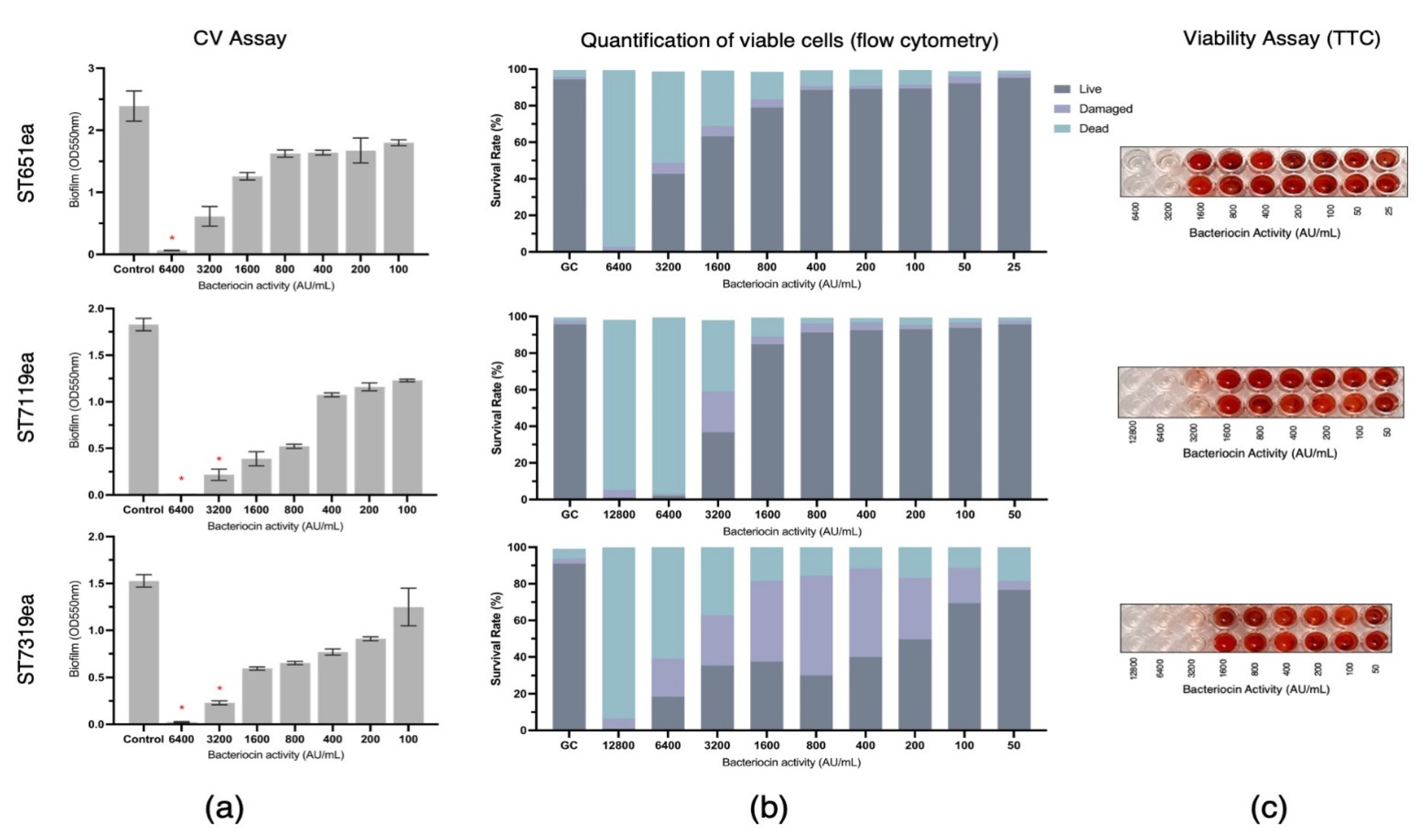
3.3. Biofilm Inhibition by Partially Purified Bacteriocins ST651ea, ST7119ea, and ST7319ea
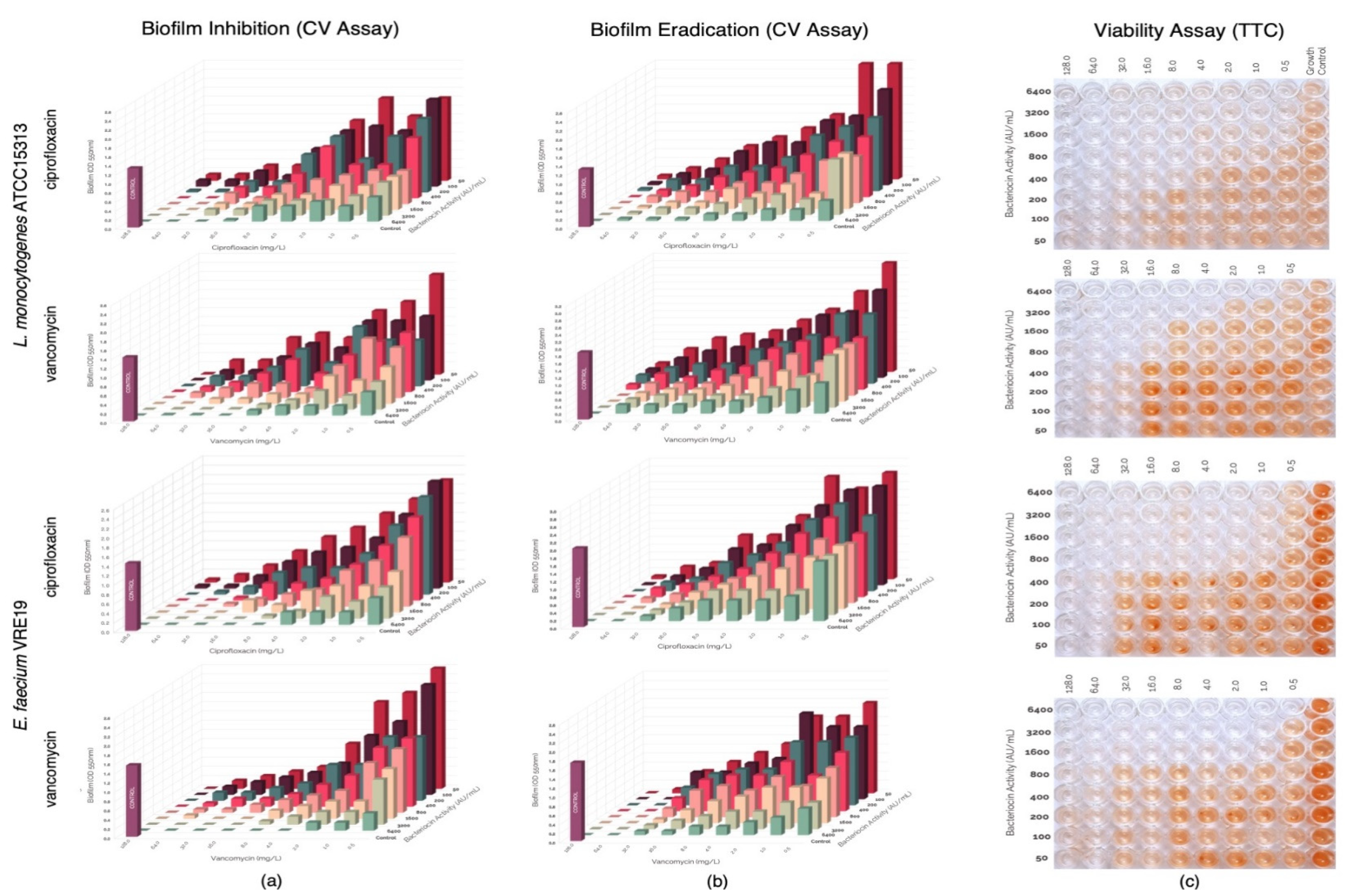
3.4. Assessment of Synergism of Bacteriocins and Antibiotics against Biofilm Formation of L. monocytogenes ATCC15313 and E. faecium VRE19
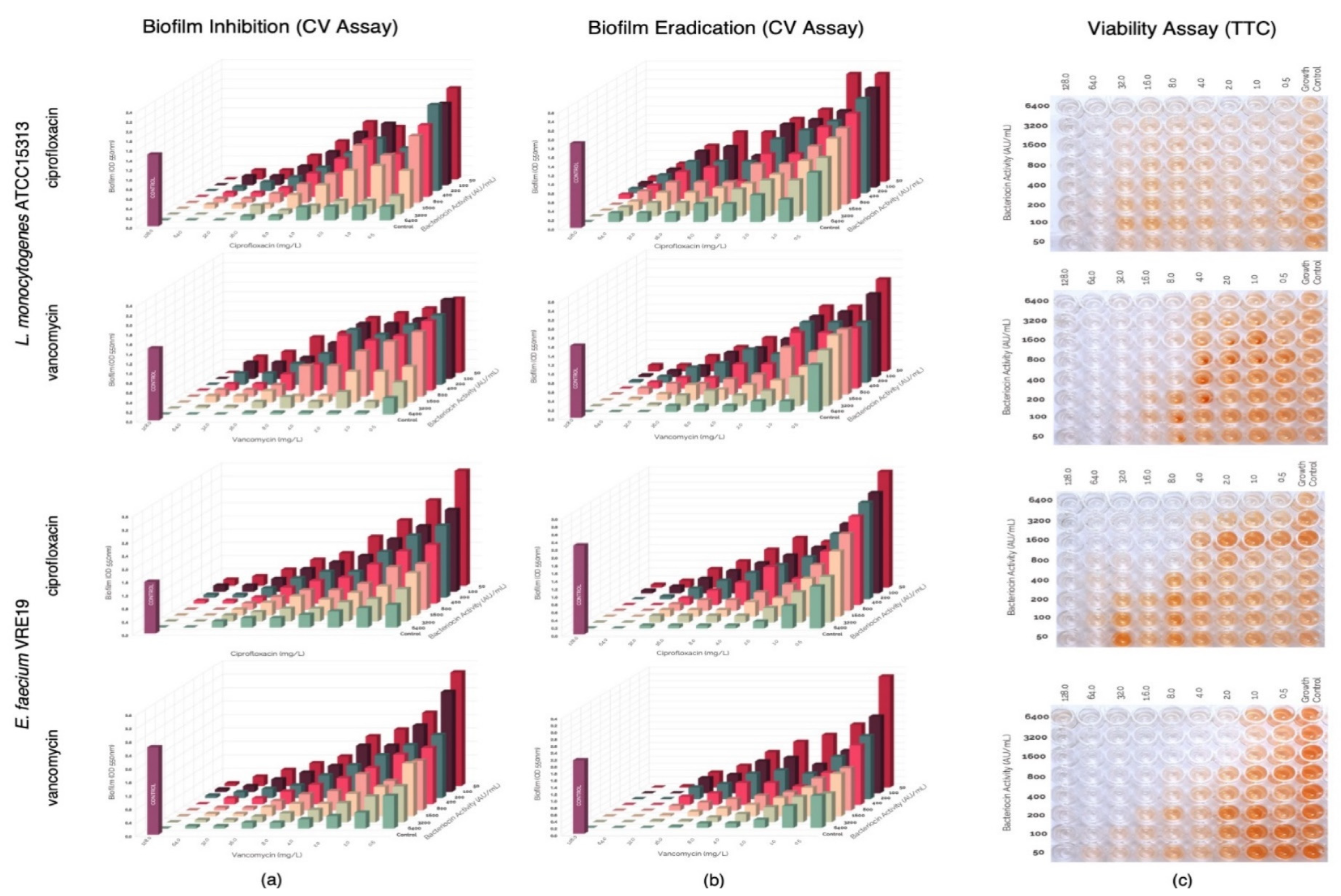
3.5. Assessment of Synergism of Bacteriocins and Antibiotics against Pre-Formed Biofilms of L. monocytogenes ATCC15313 and E. faecium VRE19
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balcázar, J.L.; Subirats, J.; Borrego, C.M. The role of biofilms as environmental reservoirs of antibiotic resistance. Front. Microbiol. 2015, 6, 1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chew, S.C.; Yang, L.; Biofilms, I. Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Boca Raton, FL, USA, 2016; pp. 407–415. [Google Scholar]
- Mah, T.F.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Al-Holy, M.A.; Shahbaz, H.M.; Al-Nabulsi, A.A.; Ghoush, M.H.A.; Osaili, T.M.; Ayyash, M.M.; Holley, R.A. Emergence of antibiotic resistance in Listeria monocytogenes isolated from food products: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1277–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diggle, S.P.; Williams, P. Quorum sensing. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Maloy, S., Hughes, K., Eds.; Academic Press: Boca Raton, FL, USA, 2013; pp. 25–27. [Google Scholar]
- Frederix, M.; Downie, J.A. Quorum Sensing: Regulating the Regulators; Poole, R.K., Ed.; Academic Press: Boca Raton, FL, USA, 2011; Volume 58, pp. 23–80. [Google Scholar]
- Houdt, R.V.; Michiels, C.W. Biofilm formation and the food industry, a focus on the bacterial outer surface. J. Appl. Microbiol. 2010, 109, 1117–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulsen, L.V. Microbial biofilm in food processing. LWT—Food Sci. Technol. 1999, 32, 321–326. [Google Scholar] [CrossRef]
- Fagerlund, A.; Langsrud, S.; Møretrø, T. Microbial diversity and ecology of biofilms in food industry environments associated with Listeria monocytogenes persistence. Curr. Opin. Food Sci. 2021, 37, 171–178. [Google Scholar] [CrossRef]
- Ranieri, M.R.; Whitchurch, C.B.; Burrows, L.L. Mechanisms of biofilm stimulation by subinhibitory concentrations of antimicrobials. Curr. Opin. Microbiol. 2018, 45, 164–169. [Google Scholar] [CrossRef]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial resistance in ESKAPE pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Lee Wong, A.C. Biofilms in food processing environments. J. Dairy Sci. 1998, 81, 2765–2770. [Google Scholar] [CrossRef]
- Patil, A.; Banerji, R.; Kanojiya, P.; Saroj, S.D. Foodborne ESKAPE biofilms and antimicrobial resistance: Lessons learned from clinical isolates. Pathol. Glob. Health 2021, 115, 339–356. [Google Scholar] [CrossRef]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti-Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- Thakur, M.; Asrani, R.K.; Patial, V. Listeria monocytogenes: A food-borne pathogen. In Foodborne Diseases; Holban, A.M., Grumezescu, A.M., Eds.; Academic Press: Boca Raton, FL, USA, 2018; pp. 157–192. [Google Scholar]
- Gandhi, M.; Chikindas, M.L. Listeria: A foodborne pathogen that knows how to survive. Int. J. Food Microbiol. 2007, 113, 1–15. [Google Scholar] [CrossRef]
- Pearson, L.J.; Marth, E.H. Listeria monocytogenes—Threat to a safe food supply: A Review. J. Dairy Sci. 1990, 73, 912–928. [Google Scholar] [CrossRef]
- Alzamora, S.M.; López-Malo, A.; Tapia, M.S.; Welti-Chanes, J. Minimally processed foods. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Boca Raton, FL, USA, 2016; pp. 767–771. [Google Scholar]
- Kumar, A.; Jha, A. Drug development strategies. In Anticandidal Agents; Kumar, A., Jha, A., Eds.; Academic Press: Boca Raton, FL, USA, 2017; pp. 63–71. [Google Scholar]
- Møretrø, T.; Langsrud, S. Listeria monocytogenes: Biofilm formation and persistence in food-processing environments. Biofilms 2004, 1, 107–121. [Google Scholar] [CrossRef]
- Oloketuyi, S.F.; Khan, F. Inhibition strategies of Listeria monocytogenes biofilms—Current knowledge and future outlooks. J. Basic Microbiol. 2017, 57, 728–743. [Google Scholar] [CrossRef]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti-Infect. Ther. 2014, 12, 1221–12236. [Google Scholar] [CrossRef]
- Miller, W.R.; Murray, B.E.; Rice, L.B.; Arias, C.A. Vancomycin-resistant enterococci: Therapeutic challenges in the 21st century. Infect. Dis. Clin. N. Am. 2016, 30, 415–439. [Google Scholar] [CrossRef]
- Jiang, X.; Yu, T.; Xu, P.; Xu, X.; Ji, S.; Gao, W.; Shi, L. Role of efflux pumps in the in vitro development of ciprofloxacin resistance in Listeria monocytogenes. Front. Microbiol. 2018, 9, 2350. [Google Scholar] [CrossRef]
- Said, M.S.; Tirthani, E.; Lesho, E. Enterococcus infections. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Chng, J.-H.; Chong, K.K.L.; Lam, L.N.; Wong, J.J.; Kline, K.A. Biofilm-associated infection by enterococci. Nat. Rev. Microbiol. 2019, 17, 82–94. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.d.L.B.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G. Guidance on the assessment of the safety of feed additives for the target species. EFSA J. 2017, 15, e05021. [Google Scholar]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Drider, D.; Fimland, G.; Héchard, Y.; McMullen, L.M.; Prévost, H. The continuing story of Class IIa bacteriocins. Microbiol. Mol. Biol. Rev. 2006, 70, 564–582. [Google Scholar] [CrossRef] [Green Version]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Sachadyn-Król, M.; Varzakas, T. Lactic acid bacteria as antibacterial agents to extend the shelf life of fresh and minimally processed fruits and vegetables: Quality and safety aspects. Microorganisms 2020, 8, 952. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C.; Ross, R.P. Bacterial lantibiotics: Strategies to improve therapeutic potential. Curr. Protein Pept. Sci. 2005, 6, 61–75. [Google Scholar] [CrossRef]
- Joerger, R.D. Alternatives to antibiotics: Bacteriocins, antimicrobial peptides, and bacteriophages. Poultry Sci. 2003, 82, 640–647. [Google Scholar] [CrossRef]
- Leite, E.L.; Oliveira, A.F.D., Jr.; Carmo, F.L.R.D.; Berkova, N.; Barh, D.; Ghosh, P.; Azevedo, V. Bacteriocins as an alternative in the treatment of infections by Staphylococcus aureus. An. Acad. Bras. Ciências 2020, 92, e20201216. [Google Scholar] [CrossRef]
- Melander, R.J.; Zurawski, D.V.; Melander, C. Narrow-spectrum antibacterial agents. MedChemComm 2018, 9, 12–21. [Google Scholar] [CrossRef]
- Fugaban, J.I.I.; Vazquez Bucheli, J.E.; Holzapfel, W.H.; Todorov, S.D. Characterization of partially purified bacteriocins produced by Enterococcus faecium strains isolated from soybean paste active against Listeria spp. and vancomycin-resistant enterococci. Microorganisms 2021, 9, 1085. [Google Scholar] [CrossRef]
- Todorov, S.D.; Dicks, L.M.T. Evaluation of lactic acid bacteria from kefir, molasses and olive brine as possible probiotics based on physiological properties. Ann. Microbiol. 2008, 58, 661–670. [Google Scholar] [CrossRef]
- Todorov, S.D.; de Paula, O.A.L.; Camargo, A.C.; Lopes, D.A.; Nero, L.A. Combined effect of bacteriocin produced by Lactobacillus plantarum ST8SH and vancomycin, propolis or EDTA for controlling biofilm development by Listeria monocytogenes. Rev. Arg. Microbiol. 2018, 50, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Doijad, S.P.; Barbuddhe, S.B.; Garg, S.; Poharkar, K.V.; Kalorey, D.R.; Kurkure, N.V.; Rawool, D.B.; Chakraborty, T. Biofilm-forming abilities of Listeria monocytogenes serotypes isolated from different sources. PLoS ONE 2015, 10, e0137046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Kranjec, C.; Ovchinnikov, K.V.; Grønseth, T.; Ebineshan, K.; Srikantam, A.; Diep, D.B. A bacteriocin-based antimicrobial formulation to effectively disrupt the cell viability of methicillin-resistant Staphylococcus aureus (MRSA) biofilms. NPJ Biofilms Microbiomes 2020, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Faleiro, M.L.; Miguel, M.G. Use of essential oils and their components against multidrug-resistant bacteria. In Fighting Multidrug Resistance with Herbal Extracts, Essential Oils and Their Components; Rai, M.K., Kon, K.V., Eds.; Academic Press: Boca Raton, FL, USA, 2013; pp. 65–94. [Google Scholar]
- Nes, I.F.; Diep, D.B.; Ike, Y. Enterococcal bacteriocins and antimicrobial proteins that contribute to niche control. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar., N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Nes, I.F.; Diep, D.B.; Hvarstein, L.S.; Brurberg, M.B.; Eijsink, V.; Holo, H. Biosynthesis of bacteriocins in lactic acid bacteria. Antonie Van Leeuwenhoek 1996, 70, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Diep, D.B.; Skaugen, M.; Salehian, Z.; Holo, H.; Nes, I.F. Common mechanisms of target cell recognition and immunity for class II bacteriocins. Proc. Nat. Acad. Sci. USA 2007, 104, 2384–2389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pag, U.; Sahl, H.-G. Multiple activities in lantibiotics—Models for the design of novel antibiotics? Curr. Pharm. Des. 2002, 8, 815–833. [Google Scholar] [CrossRef]
- Rea, M.C.; Ross, R.P.; Cotter, P.D.; Hill, C. Classification of bacteriocins from Gram-positive bacteria. In Prokaryotic Antimicrobial Peptides: From Genes to Applications; Drider, D., Rebuffat, S., Eds.; Springer: New York, NY, USA, 2011; pp. 29–53. [Google Scholar]
- LeBel, M. Ciprofloxacin: Chemistry, mechanism of action, resistance, antimicrobial spectrum, pharmacokinetics, clinical trials, and adverse reactions. J. Hum. Pharmacol. Drug Ther. 1988, 8, 3–30. [Google Scholar] [CrossRef]
- Brown, P.D. Ciprofloxacin for the management of urinary tract infection. Women’s Health 2006, 2, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Sanders, C.C. Ciprofloxacin: In vitro activity, mechanism of action, and resistance. Rev. Infect. Dis. 1988, 10, 516–527. [Google Scholar] [CrossRef]
- Lyon, S.A.; Berrang, M.E.; Fedorka-Cray, P.J.; Fletcher, D.L.; Meinersmann, R.J. Antimicrobial resistance of Listeria monocytogenes isolated from a poultry further processing plant. Foodborne Pathog. Dis. 2008, 5, 253–259. [Google Scholar] [CrossRef]
- Noll, M.; Kleta, S.; Al Dahouk, S. Antibiotic susceptibility of 259 Listeria monocytogenes strains isolated from food, food-processing plants and human samples in Germany. J. Infect. Public Health 2018, 11, 572–577. [Google Scholar] [CrossRef]
- Rakic-Martinez, M.; Drevets, D.A.; Dutta, V.; Katic, V.; Kathariou, S. Listeria monocytogenes strains selected on ciprofloxacin or the disinfectant benzalkonium chloride exhibit reduced susceptibility to ciprofloxacin, gentamicin, benzalkonium chloride, and other toxic compounds. Appl. Environ. Microbiol. 2011, 77, 8714–8721. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.D.; Ford, M.; Gould, F.K. Susceptibility of enterococci to ciprofloxacin. J. Antimicrob. Chemother. 1994, 34, 297–298. [Google Scholar] [CrossRef]
- Bruniera, F.R.; Ferreira, F.M.; Saviolli, L.R.M.; Bacci, M.R.; Feder, D.; Pedreira, M.D.L.G.; Peterlini, M.A.S.; Azzalis, L.A.; Junqueira, V.B.C.; Fonseca, F.L.A. The use of vancomycin with its therapeutic and adverse effects: A review. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 694–700. [Google Scholar]
- Scholar, E. Vancomycin. In XPharm: The Comprehensive Pharmacology Reference; Enna, S.J., Bylund, D.B., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 1–6. [Google Scholar]
- Boneca, I.G.; Chiosis, G. Vancomycin resistance: Occurrence, mechanisms, and strategies to combat it. Expert Opin. Ther. Targets 2003, 7, 311–328. [Google Scholar] [CrossRef]
- Mühlberg, E.; Umstätter, F.; Kleist, C.; Domhan, C.; Mier, W.; Uhl, P. Renaissance of vancomycin: Approaches for breaking antibiotic resistance in multidrug-resistant bacteria. Can. J. Microbiol. 2020, 66, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Cetinkaya, Y.; Falk, P.; Mayhall, C.G. Vancomycin-resistant enterococci. Clin. Microbiol. Rev. 2000, 13, 686–707. [Google Scholar] [CrossRef]
- Franz, C.M.A.P. Enterococci in foods—A conundrum for food safety. Int. J. Food Microbiol. 2003, 88, 105–122. [Google Scholar] [CrossRef]
- Franz, C.M.A.P.; Holzapfel, W.H.; Stiles, M.E. Enterococci at the crossroads of food safety? Int. J. Food Microbiol. 1999, 4, 1–24. [Google Scholar] [CrossRef]
- Coombs, G.W.; Pearson, J.C.; Daley, D.A.; Le, T.; Robinson, O.J.; Gottlieb, T.; Howden, B.P.; Johnson, P.D.R.; Bennett, C.M.; Stinear, T.P.; et al. Molecular epidemiology of enterococcal bacteremia in Australia. J. Clin. Microbiol. 2014, 52, 897–905. [Google Scholar] [CrossRef] [Green Version]
- Faron, M.L.; Ledeboer, N.A.; Buchan, B.W. Resistance mechanisms, epidemiology, and approaches to screening for vancomycin-resistant Enterococcus in the health care setting. J. Clin. Microbiol. 2016, 54, 2436–2447. [Google Scholar] [CrossRef] [Green Version]
- Courvalin, P. Vancomycin resistance in Gram-positive cocci. Clin. Infect. Dis. 2006, 42, S25–S34. [Google Scholar] [CrossRef]
- Leavis, H.L.; Willems, R.J.L.; Wamel, W.J.B.; Schuren, F.H.; Caspers, M.P.M.; Bonten, M.J.M. Insertion sequence–driven diversification creates a globally dispersed emerging multiresistant subspecies of E. faecium. PLoS Path 2007, 3, e7. [Google Scholar] [CrossRef] [PubMed]
- Tendolkar, P.M.; Baghdayan, A.S.; Gilmore, M.S.; Shankar, N. Enterococcal surface protein, esp, enhances biofilm formation by Enterococcus faecalis. Infect. Immun. 2004, 72, 6032–6039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, F.; Lv, X.; Duan, D.; Wang, L.; Huang, J. Characterization of a linezolid- and vancomycin-resistant Streptococcus suis isolate that harbors optrA and vanG operons. Front. Microbiol. 2019, 10, 2026. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ibarreche, M.; Field, D.; Ross, R.P.; Hill, C. A bioengineered nisin derivative to control Streptococcus uberis biofilms. Appl. Environ. Microbiol. 2021, 87, e0039121. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T.; Jensen, P.Ø.; Burmølle, M.; Hentzer, M.; Haagensen, J.A.J.; Hougen, H.P.; Calum, H.; Madsen, K.G.; Moser, C.; Molin, S.; et al. Pseudomonas aeruginosa tolerance to tobramycin, hydrogen peroxide and polymorphonuclear leukocytes is quorum-sensing dependent. Microbiology 2005, 151, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Kjos, M.; Nes, I.F.; Diep, D.B. Mechanisms of resistance to bacteriocins targeting the mannose phosphotransferase system. Appl. Environ. Microbiol. 2011, 77, 3335–3342. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, R.F. The interface between COVID-19 and bacterial healthcare-associated infections. Clin. Microbiol. Infect. 2021, 27, 1772–1776. [Google Scholar] [CrossRef]
- Pelfrene, E.; Botgros, R.; Cavaleri, M. Antimicrobial multidrug resistance in the era of COVID-19: A forgotten plight? Antimicrob. Resist. Infect. Control 2021, 10, 21. [Google Scholar] [CrossRef]
- Pingitore, E.V.; Todorov, S.D.; Sesma, F.; Franco, B.D.G.M. Application of bacteriocinogenic Enterococcus mundtii CRL35 and Enterococcus faecium ST88Ch in the control of Listeria monocytogenes in fresh Minas cheese. Food Microbiol. 2012, 32, 38–47. [Google Scholar] [CrossRef]
- Barraza, D.E.; Ríos Colombo, N.S.; Galván, A.E.; Acuña, L.; Minahk, C.J.; Bellomio, A.; Chalón, M.C. New insights into enterocin CRL35: Mechanism of action and immunity revealed by heterologous expression in Escherichia coli. Mol. Microbiol. 2017, 105, 922–933. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Prakash, P.; Achra, A.; Singh, G.P.; Das, A.; Singh, R.L. Standardization and classification of in vitro biofilm formation by clinical isolates of Staphylococcus aureus. J. Glob. Infect. Dis. 2017, 9, 93–101. [Google Scholar]
- Campoli-Richards, D.M.; Monk, J.P.; Price, A.; Benfield, P.; Todd, P.A.; Ward, A. Ciprofloxacin. A review of its antibacterial activity, pharmacokinetic properties and therapeutic use. Drugs 1988, 35, 373–447. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ciprofloxacin | Vancomycin | |||||||
|---|---|---|---|---|---|---|---|---|
| L. monocytogenes ATCC15313 | E. faecium VRE19 | L. monocytogenes ATCC15313 | E. faecium VRE19 | |||||
| Inhibition | Eradication | Inhibition | Eradication | Inhibition | Eradication | Inhibition | Eradication | |
| Individual component | ||||||||
| Antibiotic * | 512 | 64 | 128 | 128 | 64 | 64 | 128 | 128 |
| bacteriocin ST651ea ** | 1600 | 3200 | 1600 | 3200 | 1600 | 3200 | 1600 | 3200 |
| bacteriocin ST7119ea ** | 3200 | 6400 | 3200 | 6400 | 3200 | 6400 | 3200 | 6400 |
| bacteriocin ST7319ea ** | 3200 | 6400 | 6400 | 12,800 | 3200 | 6400 | 6400 | 12,800 |
| Combinations | ||||||||
| antibiotic * | 64 | 16 | 32 | 64 | 16 | 32 | 32 | 32 |
| bacteriocin ST651ea ** | 100 | 1600 | 200 | 400 | 400 | 1600 | 400 | 800 |
| FIC index | 0.1875 | 0.75 | 0.375 | 0.625 | 0.5 | 1 | 0.5 | 0.5 |
| antibiotic * | 16 | 64 | 32 | 64 | 32 | 32 | 16 | 32 |
| bacteriocin ST711ea ** | 800 | 1600 | 400 | 200 | 800 | 800 | 400 | 800 |
| FIC index | 0.28125 | 1.25 | 0.375 | 0.53125 | 0.75 | 0.625 | 0.25 | 0.135 |
| antibiotic * | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 |
| bacteriocin ST7319ea ** | 800 | 1600 | 800 | 400 | 400 | 400 | 1600 | 1600 |
| FIC index | 0.3125 | 0.75 | 0.375 | 0.28125 | 0.625 | 0.5625 | 0.5 | 0.135 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fugaban, J.I.I.; Vazquez Bucheli, J.E.; Holzapfel, W.H.; Todorov, S.D. Assessment of Bacteriocin-Antibiotic Synergy for the Inhibition and Disruption of Biofilms of Listeria monocytogenes and Vancomycin-Resistant Enterococcus. Microbiol. Res. 2022, 13, 480-499. https://doi.org/10.3390/microbiolres13030033
Fugaban JII, Vazquez Bucheli JE, Holzapfel WH, Todorov SD. Assessment of Bacteriocin-Antibiotic Synergy for the Inhibition and Disruption of Biofilms of Listeria monocytogenes and Vancomycin-Resistant Enterococcus. Microbiology Research. 2022; 13(3):480-499. https://doi.org/10.3390/microbiolres13030033
Chicago/Turabian StyleFugaban, Joanna Ivy Irorita, Jorge Enrique Vazquez Bucheli, Wilhelm Heinrich Holzapfel, and Svetoslav Dimitrov Todorov. 2022. "Assessment of Bacteriocin-Antibiotic Synergy for the Inhibition and Disruption of Biofilms of Listeria monocytogenes and Vancomycin-Resistant Enterococcus" Microbiology Research 13, no. 3: 480-499. https://doi.org/10.3390/microbiolres13030033
APA StyleFugaban, J. I. I., Vazquez Bucheli, J. E., Holzapfel, W. H., & Todorov, S. D. (2022). Assessment of Bacteriocin-Antibiotic Synergy for the Inhibition and Disruption of Biofilms of Listeria monocytogenes and Vancomycin-Resistant Enterococcus. Microbiology Research, 13(3), 480-499. https://doi.org/10.3390/microbiolres13030033