
Fundamental Cause of Bio-Chirality: Space-Time Symmetry—Concept Review

Abstract
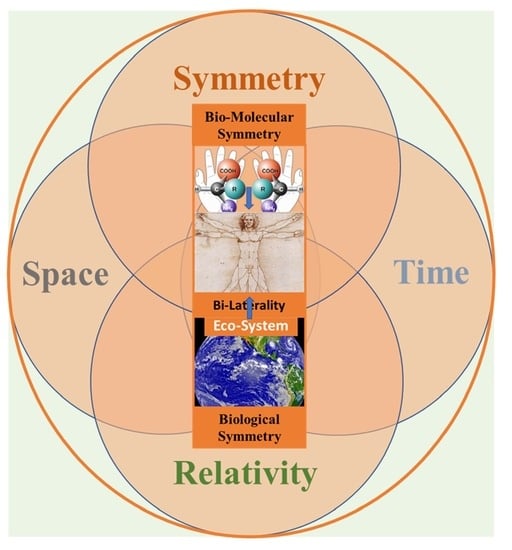
1. Introduction
Space-Time Symmetry and Relativity
“Geometrical properties of space are not independent, but they are determined by matter. We can draw conclusions about the geometrical structure of the universe only if we base our considerations on the state of the matter”[22].
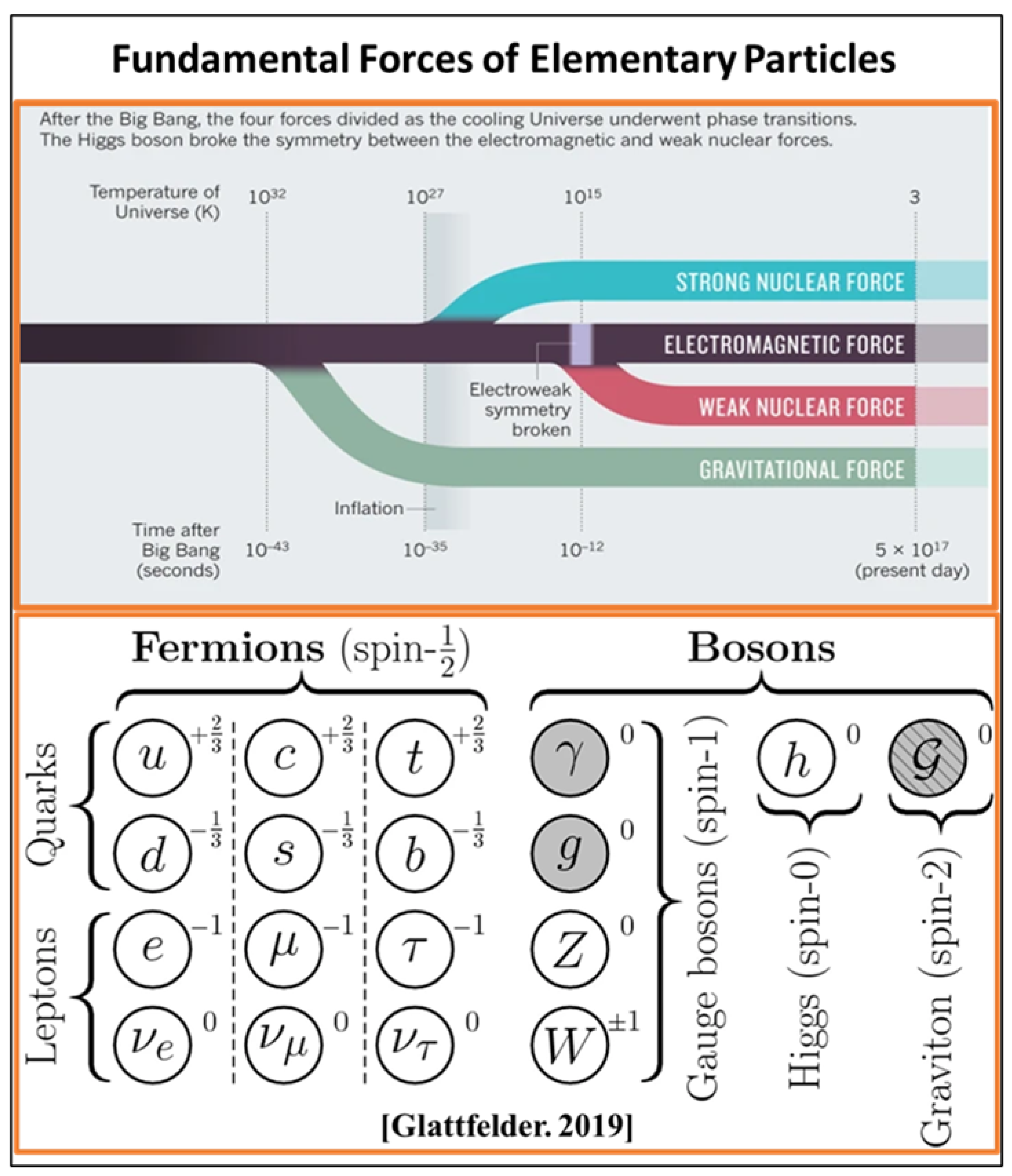
2. Standard Model and String Theory
2.1. Fundamental Forces of Nature
2.2. Symmetry and Relativity
2.3. Fragile Space-Time Symmetry: CPT Theorem
3. Bio-Chirality
3.1. Hierarchical Levels
“All forms of life, from single-cell eukaryotes to complex, highly differentiated multicellular“organisms, exhibit a property referred to as symmetry”[141].
3.2. Bilateral Symmetry
3.3. Molecular Level
4. After Words
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
Appendix A
Appendix B
Appendix C
Appendix D
References
- Ellis, G.F.R.; Kopel, J. The Dynamical Emergence of Biology From Physics: Branching Causation via Biomolecules. Front. Physiol. 2019, 9, 1966. [Google Scholar] [CrossRef] [PubMed]
- Gross, D.J. The role of symmetry in fundamental physics. Proc. Natl. Acad. Sci. USA 1996, 93, 14256–14259. [Google Scholar] [CrossRef] [PubMed]
- North, J. Two Views on Time Reversal. Philos. Sci. 2008, 75, 201–223. [Google Scholar] [CrossRef]
- Grodner, E. The mechanism of nuclear spontaneous time reversal symmetry breaking transfered to digital logic system. J. Physics: Conf. Ser. 2014, 533, 012040. [Google Scholar] [CrossRef]
- Cheong, S.A.-W.; Talbayev, D.; Kiryukhin, V.; Saxena, V.A. Broken symmetries, non-reciprocity, and multiferroicity. NPJ Quantum Mater. 2018, 3, 19. [Google Scholar] [CrossRef]
- Kai, S.; Kojima, T.; Thorp-Greenwood, F.L.; Hardie, M.J.; Hiraoka, S. How does chiral self-sorting take place in the formation of homochiral Pd6L8 capsules consisting of cyclotriveratrylene-based chiral tritopic ligands? Chem. Sci. 2018, 9, 4104–4108. [Google Scholar] [CrossRef]
- Sakata, Y.; Chiba, S.; Akine, S. Transient chirality inversion during racemization of a helical cobalt(III) complex. Proc. Natl. Acad. Sci. USA 2022, 119, e2113237119. [Google Scholar] [CrossRef]
- Li, M.; Chen, L.-J.; Cai, Y.; Luo, Q.; Li, W.; Yang, H.-B.; Tian, H.; Zhu, W.-H. Light-Driven Chiral Switching of Supramolecular Metallacycles with Photoreversibility. Chem 2019, 5, 634–648. [Google Scholar] [CrossRef]
- Naaman, N.; Paltiel, Y.; Waldeck, D.H. Chiral Induced Spin Selectivity and Its Implications for Biological Functions. Annu. Rev. Biophys. 2022, 51, 99–114. [Google Scholar] [CrossRef]
- Evers, F.; Aharony, A.; Bar-Gill, N.; Entin-Wohlman, O.; Hedegård, P.; Hod, O.; Jelinek, P.; Kamieniarz, G.; Lemeshko, M.; Michaeli, K.; et al. Theory of Chirality Induced Spin Selectivity: Progress and Challenges. Adv. Mater. 2022, 34, 2106629. [Google Scholar] [CrossRef]
- Wolf, Y.I.; Katsnelson, M.I.; Koonin, E.V. Physical foundations of biological complexity. Proc. Natl. Acad. Sci. USA 2018, 115, E8678–E8687. [Google Scholar] [CrossRef]
- Devínsky, F. Chirality and the Origin of Life. Symmetry 2021, 13, 2277. [Google Scholar] [CrossRef]
- Barron, L. True and false chirality and parity violation. Chem. Phys. Lett. 1986, 123, 423–427. [Google Scholar] [CrossRef]
- Manev, V. The Unity in the Universe: Theory of Theories Theory of Reality; Xlibris Corporation: Bloomington, IN, USA, 2014. [Google Scholar]
- The Philosophy of Right and Left: Incongruent Counterparts and the Nature of Space; van Cleve, J., Frederic, R.E., Eds.; Springer Science+Business Media, B.V.: Berlin/Heidelberg, Germany, 1991. [Google Scholar]
- Hoefer, C. Kant’s hands and Earman’s pions: Chirality arguments for substantival space. Int. Stud. Philos. Sci. 2010, 14, 237–256. [Google Scholar] [CrossRef]
- Weizsäcker, C.F.V. Kant’s first analogy of experience and conservation principles of physics. Synthese 1971, 23, 75–95. [Google Scholar] [CrossRef]
- Ezio, V. Leibniz and Clarke: A Study of Their Correspondence; Oxford University Press: Oxford, NY, USA, 1997; ISBN 9780195113990. [Google Scholar]
- Minkowski, H. Space and Time: Minkowski’s Papers on Relativity; Petkov, V., Ed.; Minkowski Institute Press: Montreal, QC, Canada, 1912. [Google Scholar]
- Einstein, A. The Meaning of Relativity: Four Lectures Delivered at Princeton University; Amazon: Seattle, DC, USA, 1921. [Google Scholar]
- Lorentz, H.A.; Einstein, A.; Minkowski, H.; Weyl, H. The Principle of Relativity; Methuen and Company, Ltd.: Methuen, MA, USA, 1923; reprinted in Dover Publications Inc.: Dover, UK, 1952. [Google Scholar]
- Einstein, A. Relativity. The Special and General Theory. In The Structure of Space According to the General Theory of Relativity; Einstein, A., Ed.; Henty Holt and Company: New York, NY, USA, 2020. [Google Scholar]
- Loveridge, L.; Miyadera, T.; Busch, P. Symmetry, Reference Frames, and Relational Quantities in Quantum Mechanics. Found. Phys. 2018, 48, 135–198. [Google Scholar] [CrossRef]
- De La Hamette, A.-C.; Galley, T.D. Quantum reference frames for general symmetry groups. Quantum 2020, 4, 367. [Google Scholar] [CrossRef]
- Penrose, R. Relativistic Symmetry Groups. In Group Theory in Non-Linear Problems; Barut, A.O., Ed.; Nato Advanced Study Institutes Series; Springer: Berlin/Heidelberg, Germany, 1974; Volume 7. [Google Scholar] [CrossRef]
- Harrison, B.K. Applications of Symmetry to General Relativity. Proc. Inst. Math. NAS Ukraine. 2004, 50, 131–141. [Google Scholar]
- Chao, S.D. Lorentz Transformations from Intrinsic Symmetries. Symmetry 2016, 8, 94. [Google Scholar] [CrossRef]
- Friedman, Y.; Scarr, T. Symmetry and Special Relativity. Symmetry 2019, 11, 1235. [Google Scholar] [CrossRef]
- Mišković, B. Relativity and/or Symmetry. Int. J. Theor. Math. Phys. 2014, 4, 165–172. [Google Scholar] [CrossRef]
- Fields, C. Symmetry in Quantum Theory of Gravity. Symmetry 2022, 14, 775. [Google Scholar] [CrossRef]
- Strocchi, F. Symmetry Breaking; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar] [CrossRef]
- Kostelecky, A. The Search for Relativity Violations. Sci. Am. Spec. Ed. 2006, 15, 12–21. [Google Scholar] [CrossRef]
- Bailey, Q.G. Catching relativity violations with atoms. Physics 2009, 2, 1–3. [Google Scholar] [CrossRef]
- Amelino-Camelia, G. Doubly-Special Relativity: Facts, Myths and Some Key Open Issues. Symmetry 2010, 2, 230–271. [Google Scholar] [CrossRef]
- Ajaltouni, Z.J. Symmetry and relativity: From classical mechanics to modern particle physics. Nat. Sci. 2014, 6, 43343. [Google Scholar] [CrossRef]
- Barceló, C.; Carballo-Rubio, R.; Garay, L.J. Weyl relativity: A novel approach to Weyl’s ideas. J. Cosmol. Astropart. Phys. 2017, 1706, 014. [Google Scholar] [CrossRef]
- Douglas, M.R.; Zhou, C.-G. Chirality Change in String Theory. J. High Energy Phys. 2004, 2004, 014. [Google Scholar] [CrossRef]
- Bernabeu, J. Symmetries and Their Breaking in the Fundamental Laws of Physics. Symmetry 2020, 12, 1316. [Google Scholar] [CrossRef]
- Qadir, A.; Camci, U. Physical Significance of Noether Symmetries. Symmetry 2022, 14, 476. [Google Scholar] [CrossRef]
- Glattfelder, J.B. The Unification Power of Symmetry. In Information—Consciousness—Reality; The Frontiers Collection; Springer: Cham, Switzerland, 2019; pp. 93–138. [Google Scholar] [CrossRef]
- Galloway, G.J.; Khuri, M.A.; Woolgar, E. The topology of general cosmological models. Class. Quantum Gravity 2022, 39, 195004. [Google Scholar] [CrossRef]
- Wang, Q.; William, G. Unruh. Vacuum fluctuation, microcyclic universes, and the cosmological constant problem Phys. Rev. D 2020, 102, 023537. [Google Scholar] [CrossRef]
- Dyakin, V.V.; Dyakina-Fagnano, N.V.; Mcintire, L.B.; Uversky, V.N. Fundamental Clock of Biological Aging: Convergence of Molecular, Neurodegenerative, Cognitive and Psychiatric Pathways: Non-Equilibrium Thermodynamics Meet Psychology. Int. J. Mol. Sci. 2022, 23, 285. [Google Scholar] [CrossRef]
- Huntley, H.E. The Divine Proportion; Courier Corporation: North Chelmsford, MA, USA, 2012. [Google Scholar]
- Wikimedia Commons. File: Chirality with hands. jpg from NASA. 2022. Available online: https://commons.wikimedia.org/wiki/File:Chirality_with_hands.jpg (accessed on 21 December 2022).
- Oerter, R. The Theory of Almost Everything: The Standard Model, the Unsung Triumph of Modern Physics, Kindle ed.; Penguin Group: New York, NY, USA, 2006; ISBN 978-0-13-236678-6. [Google Scholar]
- Bhadra, N.K. Revised Standard Model of Physics and Origin of Biology. J. Appl. Phys. 2019, 11, 12–40. [Google Scholar] [CrossRef]
- Bernabeu, J. Symmetries in the Standard Model. In Fundamental Physics and Physics Education Research; Sidharth, B.G., Murillo, J.C., Michelini, M., Perea, C., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Vaccaro, J.A. Quantum asymmetry between time and space. Proc. R. Soc. A Math. Phys. Eng. Sci. 2016, 472, 20150670. [Google Scholar] [CrossRef]
- Livio, M. Why symmetry matters. Nature 2012, 490, 472–473. [Google Scholar] [CrossRef]
- Longo, M.J. Detection of a dipole in the handedness of spiral galaxies with redshifts z∼0.04. Phys. Lett. B 2011, 699, 224–229. [Google Scholar] [CrossRef]
- Salam, S.; Taylor, J.C. Unification of Fundamental Forces; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Ellis, G.; Silk, J. Scientific method: Defend the integrity of physics. Nature 2014, 516, 321–323. [Google Scholar] [CrossRef]
- Letokhov, V. On difference of energy levels of left and right molecules due to weak interactions. Phys. Lett. A 1975, 53, 275–276. [Google Scholar] [CrossRef]
- Darquié, B.; Stoeffler, C.; Shelkovnikov, A.; Daussy, C.; Amy-Klein, A.; Chardonnet, C.; Zrig, S.; Guy, L.; Crassous, J.; Soulard, P.; et al. Progress toward the first observation of parity violation in chiral molecules by high-resolution laser spectroscopy. Chirality 2010, 22, 870–884. [Google Scholar] [CrossRef]
- Lin, H.; Jiang, Y.; Zhang, Q.; Hu, K.; Li, Z. An in-tether sulfilimine chiral center induces helicity in short peptides. Chem. Commun. 2016, 52, 10389–10391m. [Google Scholar] [CrossRef]
- Fuchs, J.-N.; Tissier, M. An Introduction to Symmetries and Quantum Field Theory. {cource_tqc4(4). pdf} Pg. 1–86. 2019. Available online: https://jnfuchs.monsite-orange.fr/file/d8233ff3c3420c3461be6b551d8ace47.pdf (accessed on 21 December 2022).
- Gu, Y.-Q. Theory of Spinors in Curved Space-Time. Symmetry 2021, 13, 1931. [Google Scholar] [CrossRef]
- Yamada, K.; Tanaka, T. Parametrized test of parity-violating gravity using GWTC-1 events. Prog. Theor. Exp. Phys. 2022, 2020, 093E01. [Google Scholar] [CrossRef]
- Cai, R.-G.; Fu, C.; Yu, W.-W. Parity violation in stochastic gravitational wave background from inflation in Nieh-Yan modified teleparallel gravity. Phys. Rev. D 2022, 105, 103520. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Jiang, X.-D.; Zhou, J.-G. Chiral extension of the standard model. Nuovo Cim. A 1995, 108, 167–174. [Google Scholar] [CrossRef]
- Volovik, G.E. Momentum-Space Topology of Standard Model. J. Low Temp. Phys. 1999, 119, 241–247. [Google Scholar] [CrossRef]
- Goldfain, E. Fractal Geometry and the Dynamics of the Standard Model. J. Phys. Astron. 2014, 3, 92–96. Available online: WWW.Mehtapress.Com (accessed on 21 December 2022).
- Miyazawa, H. Superalgebra and fermion-boson symmetry. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 158–164. [Google Scholar] [CrossRef][Green Version]
- Ibáñez, L.E.; Uranga, A.M. String Theory and Particle Physics—An Introduction to String Phenomenology; Cambridge Univ. Press: Cambridge, UK, 2012. [Google Scholar] [CrossRef]
- Fuentealba, O.; González, H.A.; Pino, M.; Troncoso, R. The anisotropic chiral boson. J. High Energy Phys. 2019, 2019, 123. [Google Scholar] [CrossRef]
- Wang, J. CT or P Problem and Symmetric Gapped Fermion Solution. arXiv 2022, arXiv:2207.14813. [Google Scholar] [CrossRef]
- Jacobs, C. Invariance or equivalence: A tale of two principles. Synthese 2021, 199, 9337–9357. [Google Scholar] [CrossRef]
- Petitjean, M. About Chirality in Minkowski Spacetime. Symmetry 2019, 11, 1320. [Google Scholar] [CrossRef]
- Liu, Y.-C.; Gao, L.-L.; Mameda, K.; Huang, X.-G. Chiral kinetic theory in curved spacetime. Phys. Rev. D 2019, 99, 085014m. [Google Scholar] [CrossRef]
- Petitjean, M. Chirality in Affine Spaces and in Spacetime. Cornel Univ. 2022, arXiv:2203.04066. [Google Scholar]
- Lyre, H. Gauge Symmetry. In Compendium of Quantum Physics; Greenberger, D., Hentschel, K., Weinert, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; p. 248. [Google Scholar] [CrossRef]
- Huang, K.A. Superfluid Universe; World Scientific: Singapore, 2016. [Google Scholar]
- Gibney, E. Force of nature gave life its asymmetry. Nature 2014. [Google Scholar] [CrossRef]
- Senami, M.; Shimizu, T. Electron chirality in amino acid molecules. Phys. Lett. A 2020, 384, 126796. [Google Scholar] [CrossRef]
- Greensite, J.; Matsuyama, K. Symmetry, Confinement, and the Higgs Phase (This article belongs to the Special Issue New Applications of Symmetry in Lattice Field Theory). Symmetry 2022, 14, 177. [Google Scholar] [CrossRef]
- Kondepudi, D.K.; Durand, D.J. Chiral asymmetry in spiral galaxies? Chirality 2001, 13, 351–356. [Google Scholar] [CrossRef]
- Ananthaswamy, A. Symmetric universe may be written in fabric of space-time. New Sci. 2009, 201, 13. [Google Scholar] [CrossRef]
- Palle, D. On Chirality of the Vorticity of the Universe. Entropy 2012, 14, 958–965. [Google Scholar] [CrossRef]
- Brauner, T. Spontaneous Symmetry Breaking and Nambu–Goldstone Bosons in Quantum Many-Body Systems. Symmetry 2010, 2, 609–657. [Google Scholar] [CrossRef]
- Kharuk, I.; Shkerin, A. Solving puzzles of spontaneously broken spacetime symmetries. Phys. Rev. D 2018, 98, 125016. [Google Scholar] [CrossRef]
- Li, S.; Du, W.; Wang, Z. Properties of the physical space, the information space and out-structure space of information. J. Phys. Conf. Ser. 2019, 1168, 032054. [Google Scholar] [CrossRef]
- Mashhoon, B. Conformal Symmetry, Accelerated Observers, and Nonlocality. Symmetry 2019, 11, 978. [Google Scholar] [CrossRef]
- Samokhvalov, S.; House, M.D.A.P. About the Symmetry of General Relativity. J. Geom. Symmetry Phys. 2020, 55, 75–103. [Google Scholar] [CrossRef]
- Fields, C.; Glazebrook, J.; Marcianò, A. Reference Frame Induced Symmetry Breaking on Holographic Screens. Symmetry 2021, 13, 408. [Google Scholar] [CrossRef]
- Dittrich, B. Diffeomorphism Symmetry in Quantum Gravity Models. Adv. Sci. Lett. 2008, 2, 151–163. [Google Scholar] [CrossRef]
- Fichet, S.; Saraswat, P. Approximate symmetries and gravity. J. High Energy Phys. 2020, 2020, 88. [Google Scholar] [CrossRef]
- Penrose, R.; Feynman, R.P.; Leighton, R.B.; Sands, M. Einstein’s Relativity, Symmetry, and Space-Time. In Six Not-So-Easy Pieces; Basic Book: New York, NY, USA, 2011. [Google Scholar]
- Stedman, G.E. Space-time symmetries and photon selection rules. Am. J. Phys. 1983, 51, 750–755. [Google Scholar] [CrossRef]
- Cameron, R.P.; Götte, J.B.; Barnett, S.M.; Yao, A.M. Chirality and the angular momentum of light. Philos. Trans. R. Soc. A 2017, 375, 20150433. [Google Scholar] [CrossRef]
- Newton, I. The Principia: Mathematical Principles of Natural Philosophy; Cohen, B., Whitman, A., Translator; University of California Press: Berkeley, CA, USA, 1999. [Google Scholar]
- Arthur, R.T.W. Time, Inertia and the Relativity Principle. In: Unspecified. Phil Sci Archive. 2007. Available online: http://philsci-archive.pitt.edu/id/eprint/4543 (accessed on 21 December 2022).
- Field, J.H. Space-time exchange invariance: Special relativity as a symmetry principle. Am. J. Phys. 2001, 69, 569–575. [Google Scholar] [CrossRef]
- Hon, G.; Goldstein, B.R. How Einstein Made Asymmetry Disappear: Symmetry and Relativity in 1905. Arch. Hist. Exact Sci. 2005, 59, 437–544. [Google Scholar] [CrossRef]
- Pfister, H.; King, M. Inertia and Gravitation: The Fundamental Nature and Structure of Space-Time; Springer: Heidelberg, Germany, 2015. [Google Scholar]
- Valente, M.B. Time in the Theory of Relativity: Inertial Time, Light Clocks, and Proper Time. arXiv 2016, arXiv:1610.08131. [Google Scholar]
- Tehrani, F.T. On Lorentz Transformations and the Theory of Relativity. J. Mod. Phys. 2022, 13, 1341–1347. [Google Scholar] [CrossRef]
- Byer, N.E. Noether’s Discovery of the Deep Connection Between Symmetries and Conservation Laws. Isreal Math. Conf. Proc. 1999, 12, 1999. Available online: http://cwp.library.ucla.edu/articles/noether.asg/noether.html (accessed on 21 December 2022).
- Vilasi, G.; Vitale, P. On SO(2,1) symmetry in general relativity. Class. Quantum Gravity 2002, 19, 3333–3340. [Google Scholar] [CrossRef][Green Version]
- Torretti, R. Relativity and Geometry; Pergamon Press: Oxford, UK, 1983. [Google Scholar]
- Auffray, C.; Nottale, L. Scale relativity theory and integrative systems biology: 1: Founding principles and scale laws. Prog. Biophys. Mol. Biol. 2008, 97, 79–114. [Google Scholar] [CrossRef]
- Noble, D. A theory of biological relativity: No privileged level of causation. Interface Focus 2012, 2, 55–64. [Google Scholar] [CrossRef]
- Ross, L.N. Causal Concepts in Biology: How Pathways Differ from Mechanisms and Why It Matters. Br. J. Philos. Sci. 2021, 72, 131–158. [Google Scholar] [CrossRef]
- Noether, E. Nachrichten von der Gesellschaft der Wissenschaften zu Göttingen. Mathematisch-Physikalische Klasse. Invariante Var. Probl. 1918, 1918, 235–257. [Google Scholar]
- Keller, J. General Relativity as a Symmetry of a Unified Space–Time–Action Geometrical Space. Proc. Inst. Math. NAS Ukr. 2002, 43, 557–568. [Google Scholar]
- Dirac, P.A.M. The quantum theory of the electron. Proc. R. Soc. Lond. 1928, 117, 610–624. [Google Scholar]
- Gronwald, F.; Muench, U.; Macías, A.; Hehl, F.W. Volume elements of spacetime and a quartet of scalar fields. Phys. Rev. D 1998, 58, 084021. [Google Scholar] [CrossRef]
- Singh, D.; Mobed, N.; Papini, G. Helicity precession of spin-1/2 particles in weak inertial and gravitational fields. J. Phys. A Math. Gen. 2004, 37, 8329–8347. [Google Scholar] [CrossRef]
- Guijarro, A.; Yus, M. The Origin of Chirality in the Molecules of Life.A Revision from Awareness to the Current Theories and Perspectives of this Unsolved Problem. Angew. Chem. Int. Ed. 2009, 48, 2079–2080. [Google Scholar] [CrossRef]
- Nakamura, S.; Sato, S. Fragile Symmetry in the Strong Interactions. Prog. Theor. Phys. 1962, 28, 752–754. [Google Scholar] [CrossRef][Green Version]
- Znojil, M. Fragile PT-symmetry in a solvable model. J. Math. Phys. 2004, 45, 4418–4430. [Google Scholar] [CrossRef][Green Version]
- Li, R.; Bowerman, B. Symmetry Breaking in Biology; Cold Spring Harbor Perspectives in Biology; Bungtown Road, Cold Spring Harbor: New York, NY, USA, 2022; Volume 14, p. 11724. [Google Scholar]
- Ji, X.; Axford, D.; Owen, R.; Evans, G.; Ginn, H.M.; Sutton, G.; Stuart, D.I. Polyhedra structures and the evolution of the insect viruses. J. Struct. Biol. 2015, 192, 88–99. [Google Scholar] [CrossRef]
- Mezey, P.G. Generalized chirality and symmetry deficiency. J. Math. Chem. 1998, 23, 5–84. [Google Scholar] [CrossRef]
- Wang, J.-S.; Wang, G.; Feng, X.-Q.; Kitamura, T.; Kang, Y.-L.; Yu, S.-W.; Qin, Q. Hierarchical chirality transfer in the growth of Towel Gourd tendrils. Sci. Rep. 2013, 3, 3102. [Google Scholar] [CrossRef] [PubMed]
- Quack, M. Time and Time Reversal Symmetry in Quantum Chemical Kinetics. In Fundamental World of Quantum Chemistry; Brändas, E.J., Kryachko, E.S., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 423–474. [Google Scholar] [CrossRef]
- Quack, M. On Biomolecular Homochirality as a Quasi-Fossil of the Evolution of Life. In Proceedings of the 240 Conference: Science's Great Challenges; Wiley: Hoboken, NJ, USA, 2014; Volume 157. [Google Scholar] [CrossRef]
- Barron, L.D. Symmetry and Chirality: Where Physics Shakes Hands with Chemistry and Biology. Isr. J. Chem. 2021, 61, 517–529. [Google Scholar] [CrossRef]
- Quack, M. Molecular chirality and the fundamental symmetries of physics: Influence of parity violation on rovibrational frequencies and thermodynamic properties. Chirality 2001, 13, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.A. Fundamental Forces of Nature: The Story of Gauge Fields; World Scientific: Singapore, 2007; p. 284. [Google Scholar] [CrossRef]
- Kumar, A.; Capua, E.; Kesharwani, M.K.; Martin, J.M.L.; Sitbon, E.; Waldeck, D.H.; Naaman, R. Chirality-induced spin polarization places symmetry constraints on biomolecular interactions. Proc. Natl. Acad. Sci. USA 2017, 114, 2474–2478. [Google Scholar] [CrossRef] [PubMed]
- Kosso, P. Symmetry arguments in physics. Stud. Hist. Philos. Sci. Part A 1999, 30, 479–492. [Google Scholar] [CrossRef]
- Dunitz, J.D. Symmetry arguments in chemistry. Proc. Natl. Acad. Sci. USA 1996, 93, 14260–14266. [Google Scholar] [CrossRef]
- Dyakin, V.V.; Lucas, J.L.; Dyakina-Fagnano, N.V.; Posner, E.V.; Vadasz, C. The Chain of Chirality Transfer as Determinant of Brain Functional Laterality. Breaking the Chirality Silence: Search for New Generation of Biomarkers; Relevance to Neurodegenerative Diseases, Cognitive Psychology, and Nutrition Science. Neurol. Neurosci. Res. 2017, 1, 2. [Google Scholar] [CrossRef]
- Cline, D.B. Possible Physical Mechanisms in the Galaxy to Cause Homochiral Biomaterials for Life. Symmetry 2010, 2, 1450–1460. [Google Scholar] [CrossRef]
- Takahashi, J.-I.; Kobayashi, K. Origin of Terrestrial Bioorganic Homochirality and Symmetry Breaking in the Universe. Symmetry 2019, 11, 919. [Google Scholar] [CrossRef]
- Hestenes, D. Space-Time Algebra; Gordon and Breach Science Publishers: New York, NY, USA, 1966; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Petitjean, M. Chirality in Geometric Algebra. Mathematics 2021, 9, 1521. [Google Scholar] [CrossRef]
- Goodsell, D.S. Biomolecules and Nanotechnology. Am. Sci. 2000, 88, 230–237. [Google Scholar] [CrossRef]
- Billig, Y.; Futorny, V. Representations of the Lie algebra of vector fields on a torus and the chiral de Rham complex. Trans. Am. Math. Soc. 2014, 366, 4697–4731. Available online: https://www.jstor.org/stable/23813963 (accessed on 21 December 2022). [CrossRef]
- Edmonds, A.R. Symmetry and Conservation Laws Symmetry in Physical Laws. Lecture 17. Chapter 52, Vol. I. In Angular Momentum in Quantum Mechanics; Princeton University Press: Princeton, NJ, USA, 1957; Available online: https://www.feynmanlectures.caltech.edu/III_17.html (accessed on 21 December 2022).
- Viazminsky, C.P. Framing Noether’s Theorem. Appl. Math. 2014, 5, 274–286. [Google Scholar] [CrossRef][Green Version]
- Penchev, V. The Symmetries of Quantum and Classical Information. Quantum Informatiom. HAL ID hal-03251242. Available online: https://hal.archives-ouvertes.fr/hal-03251242 (accessed on 21 December 2022).
- Higgins, I.; Racanière, S.; Rezende, D. Symmetry-Based Representations for Artificial and Biological General Intelligence. Front. Comput. Neurosci. 2022, 14, 836498. [Google Scholar] [CrossRef]
- Hermann, S.; Schmidt, M. Why Noether’s theorem applies to statistical mechanics. J. Physics: Condens. Matter 2022, 34, 213001. [Google Scholar] [CrossRef] [PubMed]
- Levin, M. Left–right asymmetry in embryonic development: A comprehensive review. Mech. Dev. 2005, 122, 3–25. [Google Scholar] [CrossRef]
- Chiccoli, C.; Pasini, P.; Skačej, G.; Zannoni, C.; Žumer, S. Chirality Transfer from Helical Nanostructures to Nematics: A Monte Carlo Study. Mol. Cryst. Liq. Cryst. 2013, 576, 151–156. [Google Scholar] [CrossRef]
- Huang, S.; Yu, H.; Li, Q. Supramolecular Chirality Transfer toward Chiral Aggregation: Asymmetric Hierarchical Self-Assembly. Adv. Sci. 2021, 8, 2002132. [Google Scholar] [CrossRef]
- Cheng, L.; Liu, K.; Duan, Y.; Duan, H.; Li, Y.; Gao, M.; Cao, L. Adaptive Chirality of an Achiral Cage: Chirality Transfer, Induction, and Circularly Polarized Luminescence through Aqueous Host–Guest Complexation. CCS Chem. 2021, 3, 2749–2763. [Google Scholar] [CrossRef]
- Zhang, Q.; Crespi, S.; Toyoda, R.; Costil, R.; Browne, W.R.; Qu, D.-H.; Tian, H.; Feringa, B.L. Stereodivergent Chirality Transfer by Noncovalent Control of Disulfide Bonds. J. Am. Chem. Soc. 2022, 144, 4376–4382. [Google Scholar] [CrossRef]
- Sevim, S.; Sorrenti, A.; Vale, J.P.; El-Hachemi, Z.; Pané, S.; Flouris, A.D.; Mayor, T.S.; Puigmartí-Luis, J. Chirality transfer from a 3D macro shape to the molecular level by controlling asymmetric secondary flows. Nat. Commun. 2022, 13, 1766. [Google Scholar] [CrossRef]
- Gandhi, P.; Ciocanel, M.-V.; Niklas, K.; Dawes, A.T. Identification of approximate symmetries in biological development. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2021, 379, 20200273. [Google Scholar] [CrossRef]
- Keszthelyi, L. Origin of the Homochirality of Biomolecules; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Lee, C.; Weber, J.M.; Rodriguez, L.E.; Sheppard, R.Y.; Barge, L.M.; Berger, E.L.; Burton, A.S. Chirality in Organic and Mineral Systems: A Review of Reactivity and Alteration Processes Relevant to Prebiotic Chemistry and Life Detection Missions. Symmetry 2022, 14, 460. [Google Scholar] [CrossRef]
- Gal, J.; Cintas, P. Early History of the Recognition of Molecular Biochirality. In Biochirality; Cintas, P., Ed.; Topics in Current Chemistry; Springer: Berlin/Heidelberg, Germany, 2012; Volume 333, pp. 1–40. [Google Scholar]
- Tamada, A. Chiral Neuronal Motility: The Missing Link between Molecular Chirality and Brain Asymmetry. Symmetry 2019, 11, 102. [Google Scholar] [CrossRef]
- Mao, J. Cosmic Background. Eur. J. Appl. Phys. 2021, 3, 67–70. [Google Scholar] [CrossRef]
- Ribó, J.M.; Hochberg, D.; Crusats, J.; El-Hachemi, Z.; Moyano, A. Spontaneous mirror symmetry breaking and origin of biological homochirality. J. R. Soc. Interface 2017, 14, 20170699. [Google Scholar] [CrossRef]
- Benítez, H.; Lemic, D.; Villalobos-Leiva, A.; Bažok, R.; Órdenes-Claveria, R.; Živković, I.P.; Mikac, K. Breaking Symmetry: Fluctuating Asymmetry and Geometric Morphometrics as Tools for Evaluating Developmental Instability under Diverse Agroecosystems. Symmetry 2020, 12, 1789. [Google Scholar] [CrossRef]
- Satir, P. Chirality of the cytoskeleton in the origins of cellular asymmetry. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150408. [Google Scholar] [CrossRef]
- Tasker, E. Why is Life Left-Handed? The Answer Is in the Stars. The Conversation 2015. Available online: http://theconversation.com/why-is-life-left-handed-the-answer-isin-the-stars-44862 (accessed on 20 July 2015).
- Jordi, C. The Unity of Science. In The Stanford Encyclopedia of Philosophy, Spring 2022 ed.; Zalta, E.N., Ed.; Available online: https://plato.stanford.edu/archives/spr2022/entries/scientific-unity/ (accessed on 21 December 2022).
- Blackmond, D.G. The Origin of Biological Homochirality. Cold Spring Harb. Perspect. Biol. 2022, 14, a032540. [Google Scholar] [CrossRef]
- Bordács, S.; Kézsmárki, I.; Szaller, D.; Demkó, L.; Kida, N.; Murakawa, H.; Onose, Y.; Shimano, R.; Rõõm, T.; Nagel, U.; et al. Chirality of matter shows up via spin excitations. Nat. Phys. 2012, 8, 734–738. [Google Scholar] [CrossRef]
- Forbes, K.A.; Andrews, D.L. Orbital angular momentum of twisted light: Chirality and optical activity. J. Phys. Photon. 2021, 2, 022007. [Google Scholar] [CrossRef]
- Crimin, F.; MacKinnon, N.; Götte, J.; Stephen, M.; Barnett, S.M. Continuous symmetries and conservation laws in chiral media. In Proceedings of the SPIE OPTO, San Francisco, CA, USA, 1–6 February 2020. [Google Scholar] [CrossRef]
- Cottet, A.; Kontos, T.; Douçot, B. Electron-photon coupling in mesoscopic quantum electrodynamics. Phys. Rev. B 2015, 91, 205417. [Google Scholar] [CrossRef]
- Mun, J.; Kim, M.; Yang, Y.; Badloe, T.; Ni, J.; Chen, Y.; Qiu, C.-W.; Rho, J. Electromagnetic chirality: From fundamentals to nontraditional chiroptical phenomena. Light Sci. Appl. 2020, 9, 139. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, J.P.; Kashian, D.M.; Bradford, J.B.; Freeman, D.C. Variation in Fractal Symmetry of Annual Growth in Aspen as an Indicator of Developmental Stability in Trees. Symmetry 2015, 7, 354–364. [Google Scholar] [CrossRef]
- Toga, A.W.; Thompson, P.M. Mapping brain asymmetry. Nat. Rev. Neurosci. 2003, 4, 37–48. [Google Scholar] [CrossRef]
- Li, P.; Ensink, E.; Lang, S.; Marshall, L.; Schilthuis, M.; Lamp, J.; Vega, I.; Labrie, V. Hemispheric asymmetry in the human brain and in Parkinson’s disease is linked to divergent epigenetic patterns in neurons. Genome Biol. 2020, 21, 61. [Google Scholar] [CrossRef]
- Davankov, V.A. Biological Homochirality on the Earth, or in the Universe? A Selective Review. Symmetry 2018, 10, 749. [Google Scholar] [CrossRef]
- Gong, C.; Zhu, T.; Niu, R.; Wu, Q.; Cui, J.-L.; Zhang, X.; Zhao, W.; Wang, A. Gravitational wave constraints on Lorentz and parity violations in gravity: High-order spatial derivative cases. Phys. Rev. D 2022, 105, 044034. [Google Scholar] [CrossRef]
- Avalos, M.; Babiano, R.; Cintas, P.; Jiménez, J.L.; Palacios, J.C.; Barron, L.D. Absolute Asymmetric Synthesis under Physical Fields: Facts and Fictions. Chem. Rev. 1998, 98, 2391–2404. [Google Scholar] [CrossRef]
- Mineo, P.; Villari, V.; Scamporrino, E.; Micali, N. Supramolecular chirality induced by a weak thermal force. Soft Matter 2014, 10, 44–47. [Google Scholar] [CrossRef]
- Mondal, P.C.; Fontanesi, C.; Waldeck, D.H.; Naaman, R. Spin-Dependent Transport through Chiral Molecules Studied by Spin-Dependent Electrochemistry. Acc. Chem. Res. 2016, 49, 2560–2568. [Google Scholar] [CrossRef]
- Rosenberg, R.A. Electrochirogenesis: The Possible Role of Low-Energy Spin-Polarized Electrons in Creating Homochirality. Symmetry 2019, 11, 528. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Klabunovskii, E.I. The origin of homochirality in nature: A possible version. Russ. Chem. Rev. 2015, 18, 93–114. [Google Scholar] [CrossRef]
- Dyakin, V.V.; Lucas, J. Non-equilibrium phase transition in biochemical Systems. Chain of chirality transfer as Determinant of Brain Functional Laterality. Relevance to Alzheimer disease and cognitive psychology. In Proceedings of the Alzheimer’s Association International Conference (AAIC-2017), London, UK, 16–20 July 2017. [Google Scholar]
- Dyakin, V.V.; Wisniewski, T.M.; Lajtha, A. Racemization in Post-Translational Modifications Relevance to Protein Aging, Aggregation and Neurodegeneration: Tip of the Iceberg. Symmetry 2021, 13, 455. [Google Scholar] [CrossRef]
- Chotera, A.; Sadihov, H.; Cohen-Luria, R.; Monnard, P.; Ashkenasy, G. Functional Assemblies Emerging in Complex Mixtures of Peptides and Nucleic Acid–Peptide Chimeras. Chem. A Eur. J. 2018, 24, 10128–10135. [Google Scholar] [CrossRef]
- Kohut, P. Fundamental Discovery of the Dialectical Unity Principle and its Consequences for Theoretical Physics. J. Phys. Math. 2016, 7, 1–15. [Google Scholar] [CrossRef]
- Korenić, A.; Perović, S.; Ćirković, M.M.; Miquel, P.-A. Symmetry breaking and functional incompleteness in biological systems. Prog. Biophys. Mol. Biol. 2019, 150, 1–12. [Google Scholar] [CrossRef]
- Dyakin, V.V.; Uversky, V.N. Arrow of Time, Entropy, and Protein Folding: Holistic View on Biochirality. Int. J. Mol. Sci. 2022, 23, 3687. [Google Scholar] [CrossRef]
- Voit, G.M. Tracing cosmic evolution with clusters of galaxies. Rev. Mod. Phys. 2005, 77, 207–258. [Google Scholar] [CrossRef]
- Iqbal, N.; Khan, M.S.; Masood, T. Entropy changes in the clustering of galaxies in an expanding universe. Nat. Sci. 2011, 3, 65–68. [Google Scholar] [CrossRef]
- Aoki, I. Entropy Principle for the Development of Complex Biotic Systems: Organisms, Ecosystems, the Earth; Elsevier: London, UK, 2012. [Google Scholar] [CrossRef]
- Book by Szabados, L. Symmetries in Stellar, Galactic, and Extragalactic Astronomy. Complex Symmetries 2022, 191–206. [Google Scholar] [CrossRef]
- Gal, J. Molecular Chirality: Language, History, and Significance. Top. Curr. Chem. 2013, 340, 1–20. [Google Scholar] [CrossRef]
- Boyd, R.N.; Famiano, M.A. Amino Acids, Chirality, and Neutrinos. In Creating the Molecules of Life; IOP Publishing Ltd.: Bristol, UK, 2018; pp. 5-1–5-25. ISBN 978-0-7503-1993-5. [Google Scholar]
- Chen, Y.; Ma, W. The origin of biological homochirality along with the origin of life. PLoS Comput. Biol. 2020, 16, e1007592. [Google Scholar] [CrossRef]
- Stocker, K. The Theory of Cognitive Spacetime. Metaphor Symb. 2014, 29, 71–93. [Google Scholar] [CrossRef]
- Moser, M.-B.; Rowland, D.C.; Moser, E.I. Place Cells, Grid Cells, and Memory. Cold Spring Harb. Perspect. Biol. 2015, 14, a021808. [Google Scholar] [CrossRef]
- Hameroff, S. Consciousness, the brain, and spacetime geometry. Ann. New York Acad. Sci. 2001, 929, 74–104. [Google Scholar] [CrossRef]
- Pillai, A.S.; Jirsa, V.K. Symmetry Breaking in Space-Time Hierarchies Shapes Brain Dynamics and Behavior. Neuron 2017, 94, 1010–1026. [Google Scholar] [CrossRef]
- Signorelli, C.M.; Dündar-Coecke, S.; Wang, V.; Coecke, B. Cognitive Structures of Space-Time. Front. Psychol. 2020, 11, 527114. [Google Scholar] [CrossRef]
- O’Keefe, J.; Nadel, L. The Hippocampus as a Cognitive Map; Oxford University Press: Oxford, UK, 1978; ISBN 978-0-19-857206-0. [Google Scholar]
- Peer, M.; Salomon, R.; Goldberg, I.; Blanke, O.; Arzy, S. Brain system for mental orientation in space, time, and person. Proc. Natl. Acad. Sci. USA 2015, 112, 11072–11077. [Google Scholar] [CrossRef]
- Buzsáki, G.; Llinás, R. Space and time in the brain. Science 2017, 358, 482–485. [Google Scholar] [CrossRef]
- Le Bihan, D. On time and space in the brain: A relativistic pseudo-diffusion framework. Brain Multiphysics 2020, 1, 100016. [Google Scholar] [CrossRef]
- Angelaki, D.E.; Ng, J.; Abrego, A.M.; Cham, H.X.; Asprodini, E.K.; Dickman, J.D.; Laurens, J. A gravity-based three-dimensional compass in the mouse brain. Nat. Commun. 2020, 11, 1855. [Google Scholar] [CrossRef] [PubMed]
- Valeriy, P. Physical Space and the World, 1st ed.; Kindle, Valeriy, P., Eds.; Available online: https://www.amazon.com.br/Physical-Space-English-Valeriy-Polulyakh-ebook/dp/B008NBGTOO (accessed on 19 July 2012).
- Mullins, S.-M.; Weissker, H.-C.; Sinha-Roy, R.; Pelayo, J.J.; Garzón, I.L.; Whetten, R.L.; López-Lozano, X. Chiral symmetry breaking yields the I-Au60 perfect golden shell of singular rigidity. Nat. Commun. 2018, 9, 3352. [Google Scholar] [CrossRef] [PubMed]
- Laurent, G.; Lacoste, D.; Gaspard, P. Emergence of homochirality in large molecular systems. Proc. Natl. Acad. Sci. USA 2021, 118, e2012741118. [Google Scholar] [CrossRef] [PubMed]
- Kurakin, A. The self-organizing fractal theory as a universal discovery method: The phenomenon of life. Theor. Biol. Med. Model. 2011, 8, 4. [Google Scholar] [CrossRef]
- Gaite, J. Scale Symmetry in the Universe. Symmetry 2020, 12, 597. [Google Scholar] [CrossRef]
- Krishnamurthy, V.M.; Kaufman, G.K.; Urbach, A.R.; Gitlin, I.; Gudiksen, K.L.; Weibel, D.B.; Whitesides, G.M. Carbonic Anhydrase as a Model for Biophysical and Physical-Organic Studies of Proteins and Protein−Ligand Binding. Chem. Rev. 2008, 108, 946–1051. [Google Scholar] [CrossRef]
- Pereyaslavets, L.B.; Glyakina, A.B.; Dovidchenko, N.V.; Sokolovskiy, I.V.; Galzitskaya, O.V. What handedness and angles between helices has the studied three-helical protein domain? Bioinformatics 2015, 31, 963–965. [Google Scholar] [CrossRef][Green Version]
- Chasova, V. Direct and Indirect Empirical Statuses Compared to the Newtonian and Leibnizian Interpretations of Theoretical Symmetries in Physics. Phil. Sci Atchive. pp. 1–9. Available online: http://philsci-archive.pitt.edu/16245/ (accessed on 21 December 2022).
- Brading, K.; Brown, H.R. Are Gauge Symmetry Transformations Observable? Br. J. Philos. Sci. 2004, 55, 645–665. [Google Scholar] [CrossRef]
- Palmer, M.W. Fractal geometry: A tool for describing spatial patterns of plant communities. Vegetatio 1988, 75, 91–102. [Google Scholar] [CrossRef]
- Buldyrev, S.V. Fractals in Biology. In Encyclopedia of Complexity and Systems Science; Meyers, R., Ed.; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Losa, G.A. The fractal geometry of life. Riv. Biol. 2009, 102, 29–59. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dyakin, V.V. Fundamental Cause of Bio-Chirality: Space-Time Symmetry—Concept Review. Symmetry 2023, 15, 79. https://doi.org/10.3390/sym15010079
Dyakin VV. Fundamental Cause of Bio-Chirality: Space-Time Symmetry—Concept Review. Symmetry. 2023; 15(1):79. https://doi.org/10.3390/sym15010079
Chicago/Turabian StyleDyakin, Victor Vasilyevich. 2023. "Fundamental Cause of Bio-Chirality: Space-Time Symmetry—Concept Review" Symmetry 15, no. 1: 79. https://doi.org/10.3390/sym15010079
APA StyleDyakin, V. V. (2023). Fundamental Cause of Bio-Chirality: Space-Time Symmetry—Concept Review. Symmetry, 15(1), 79. https://doi.org/10.3390/sym15010079