

Priming Mesenchymal Stem Cells with Lipopolysaccharide Boosts the Immunomodulatory and Regenerative Activity of Secreted Extracellular Vesicles

 , , , , , , and
, , , , , , and 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Background
2. Materials and Methods
2.1. Isolation, Culture and Preconditioning of MSCs
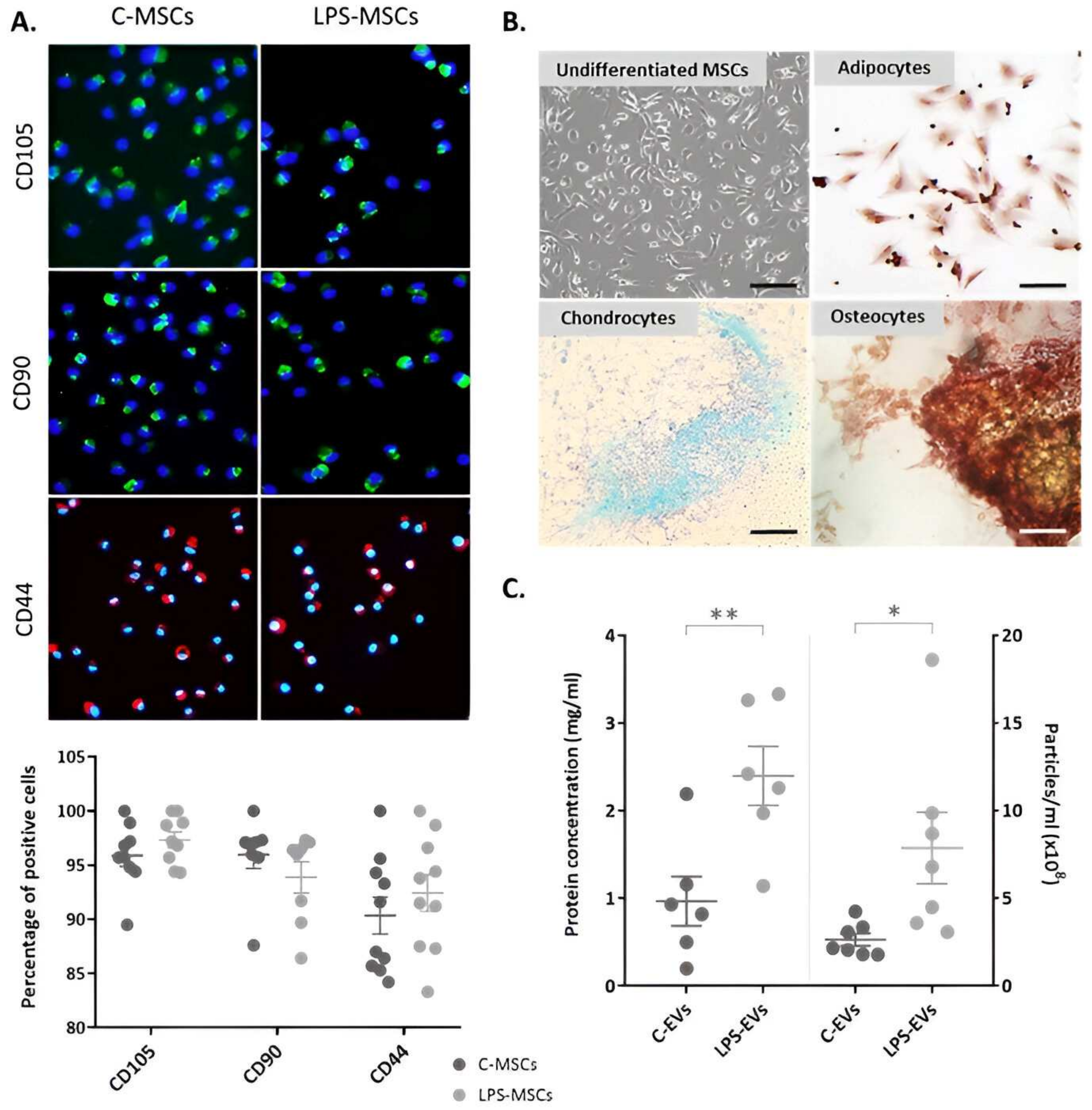
2.2. Characterization of C-MSCs and LPS-MSCs
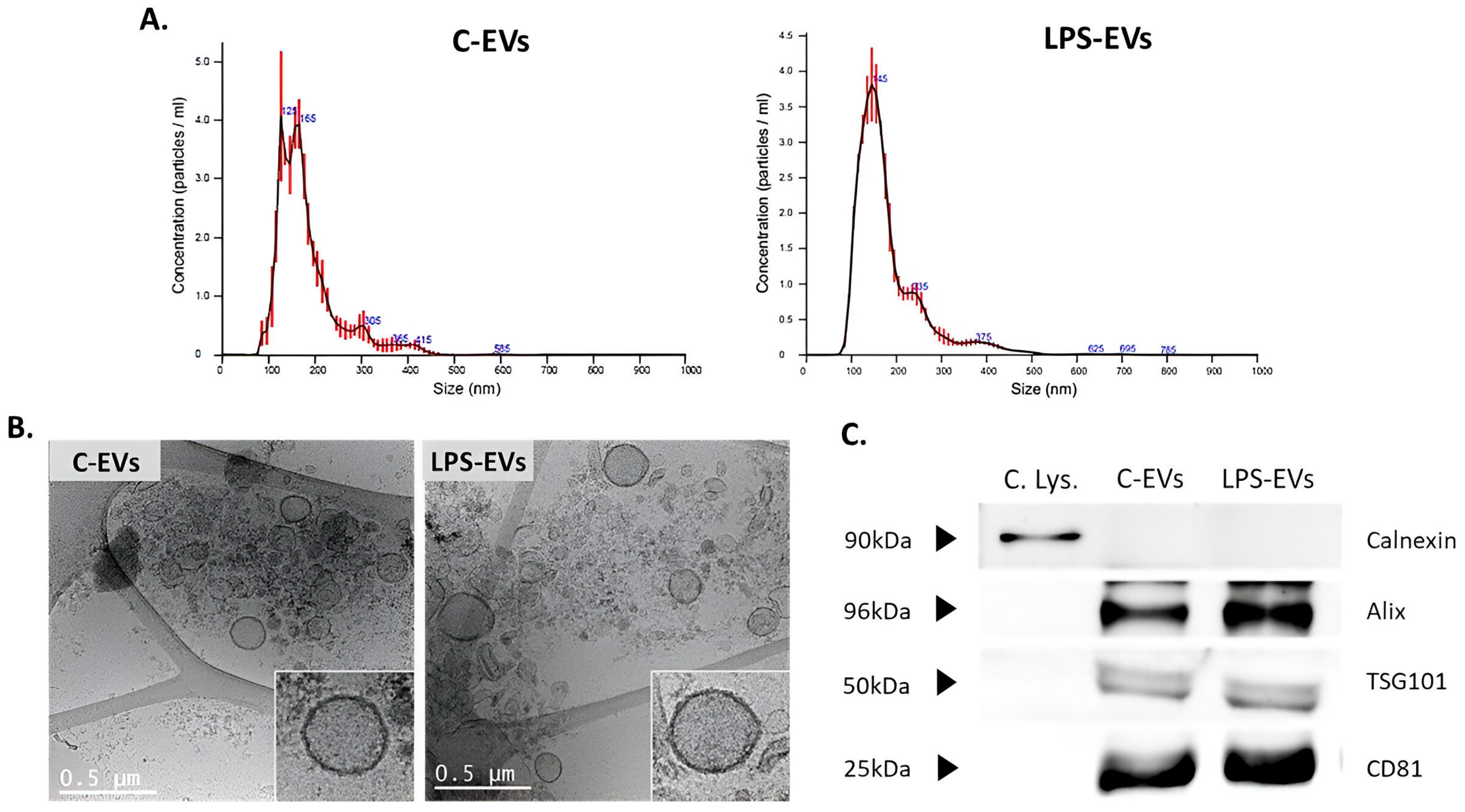
2.3. C-EVs and LPS-EVs Isolation and Characterization
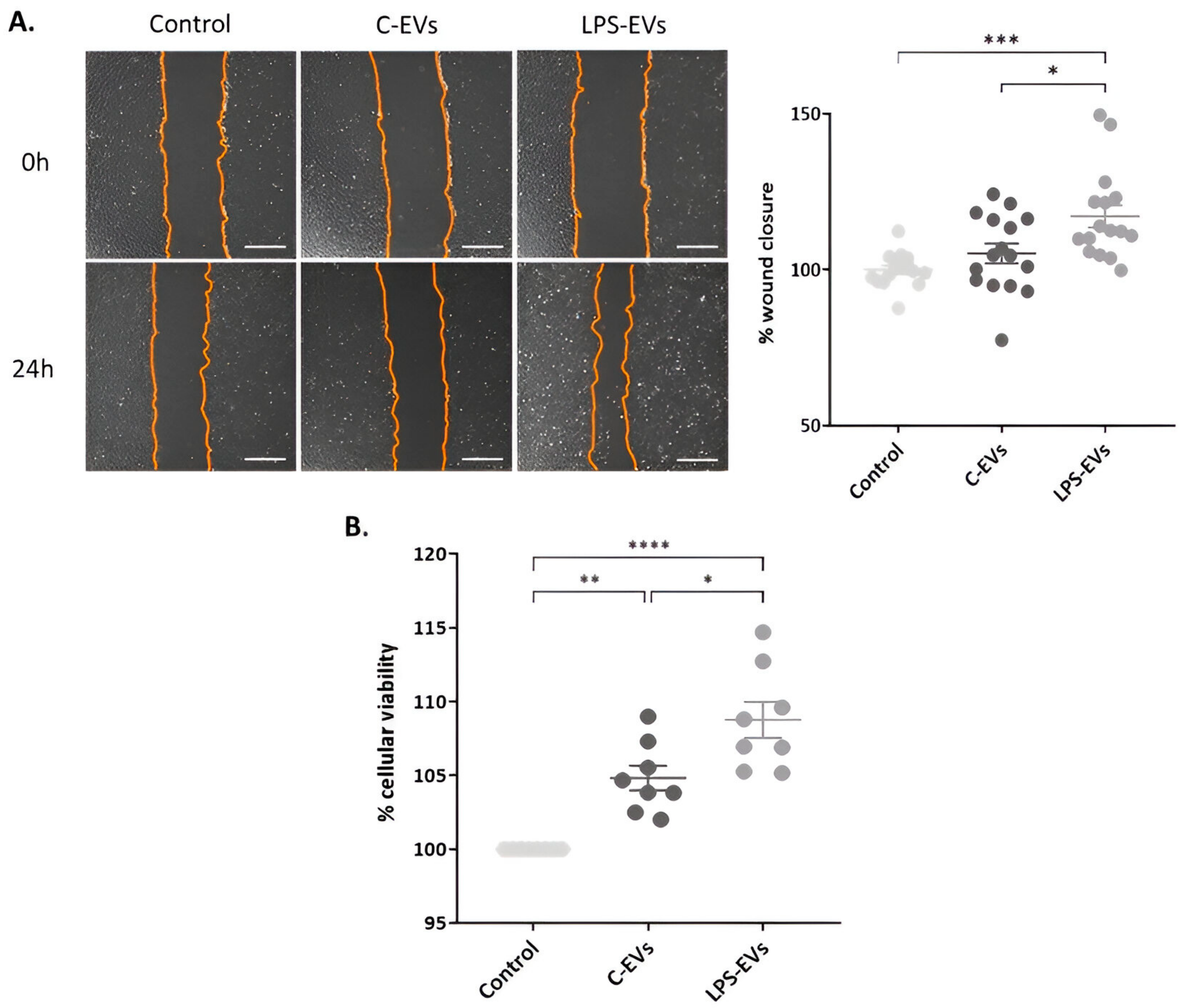
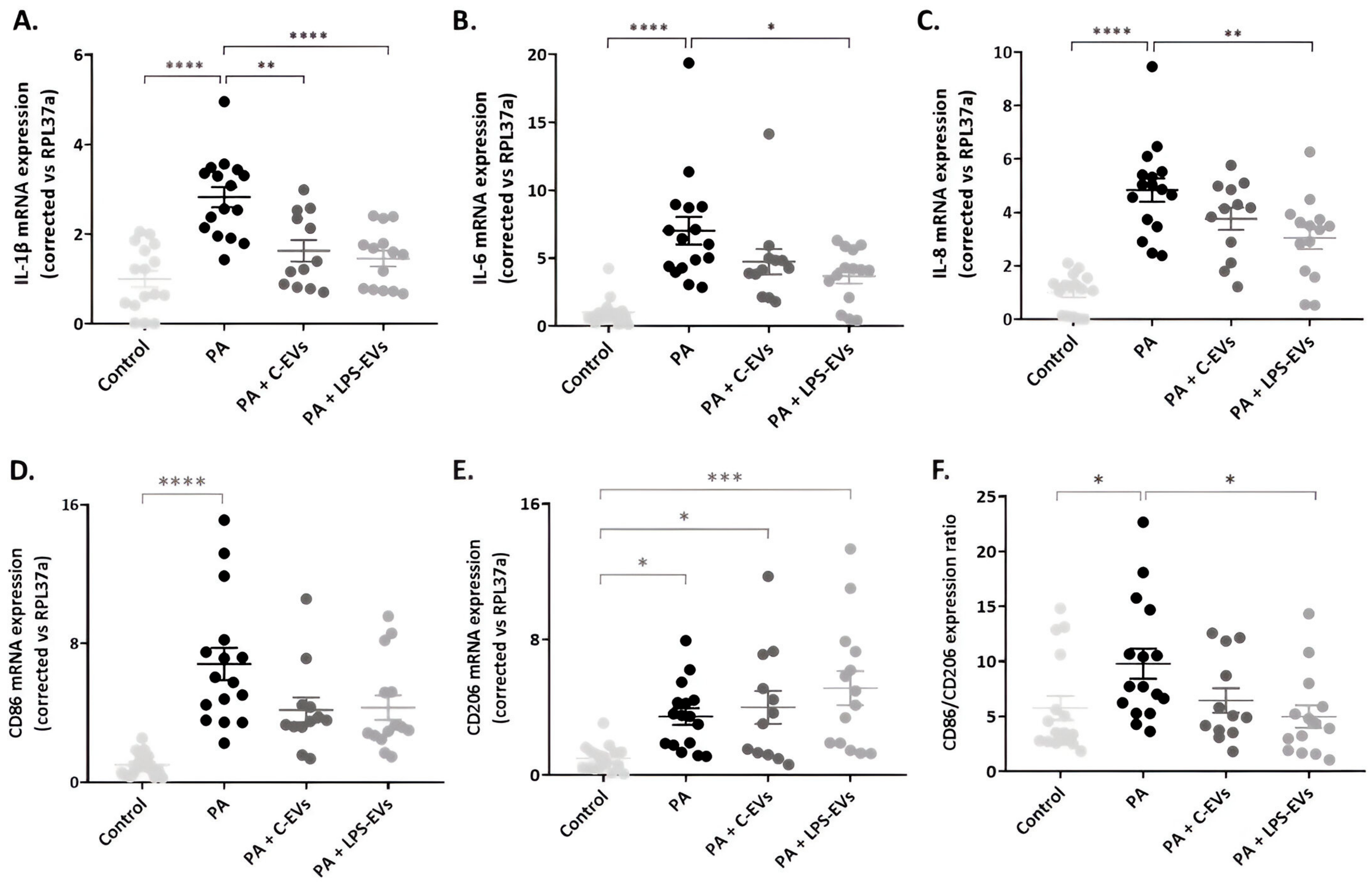
2.4. Assessment of MSCs-Derived EVs’ Therapeutic Effect In Vitro
2.5. In Vivo MSCs-Derived EVs’ Therapeutic Evaluation
2.6. Statistics for In Vitro and In Vivo Studies
3. Results
3.1. Characterization of Isolated MSCs
3.2. Characterization of MSCs-Derived EVs
3.3. LPS-EVs Accelerate Wound Healing and Cell Proliferation versus C-EVs In VItro
3.4. LPS-EVs Produce a Major Anti-Inflammatory Effect Compared to C-EVs in P. aeruginosa-Infected Macrophage-like THP-1 Cells In Vitro
3.5. C-EV and LPS-EV Therapy Ameliorates Lung/Body Weight Ratio In Vivo
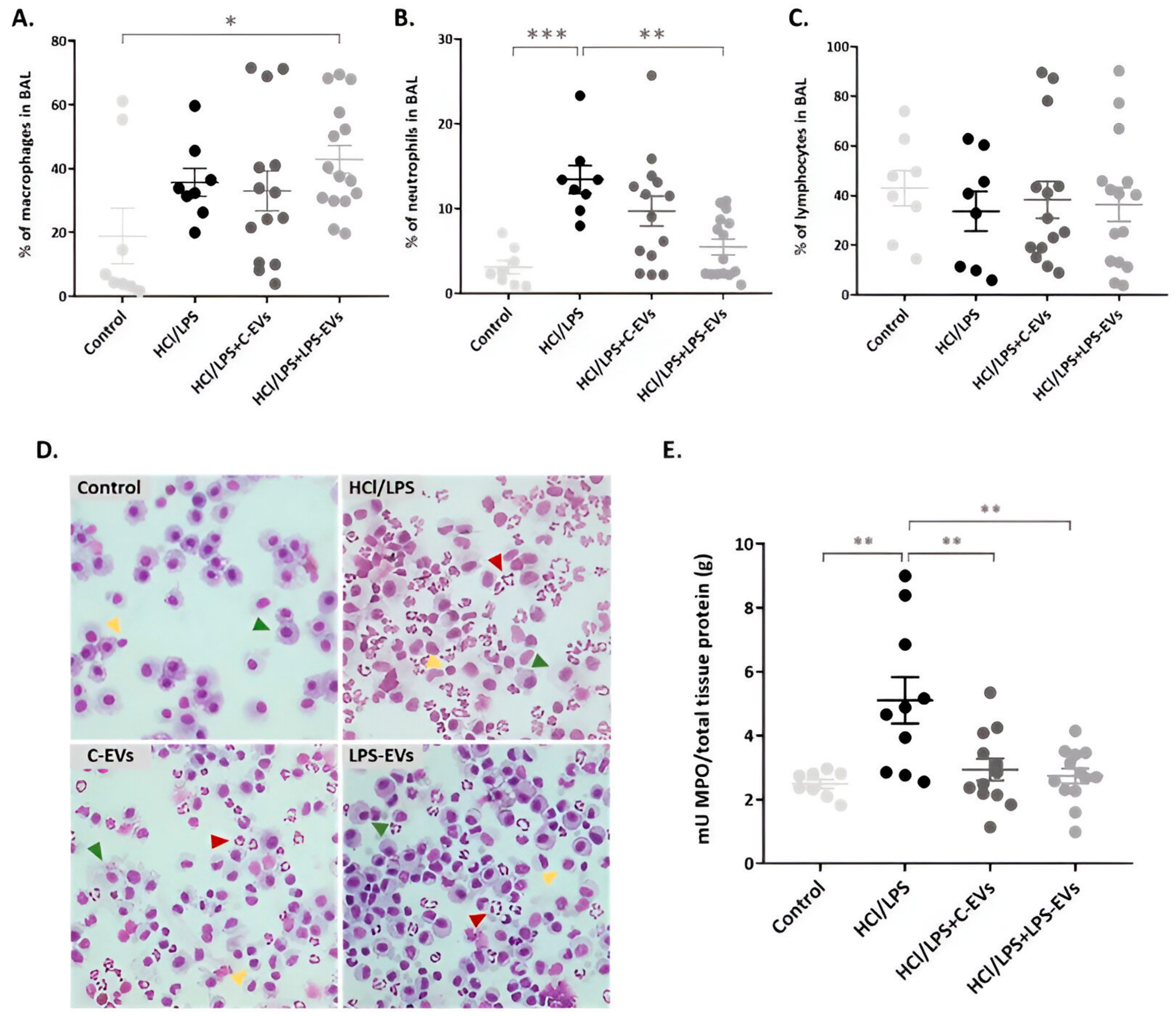
3.6. LPS-EVs and C-EVs Reduce Lung Permeability, but Only LPS-EVs Significantly Decrease Neutrophil Infiltration In Vivo
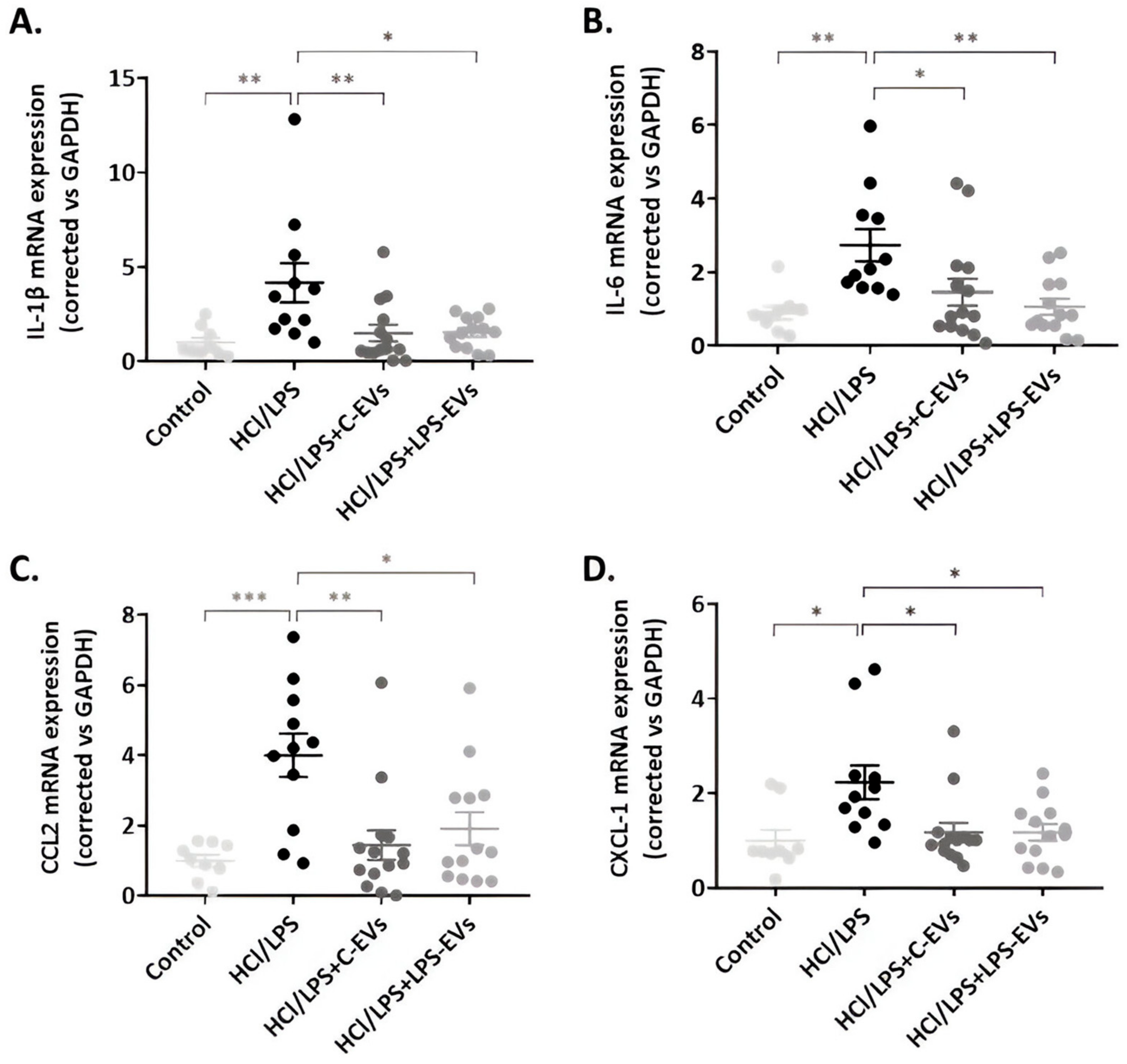
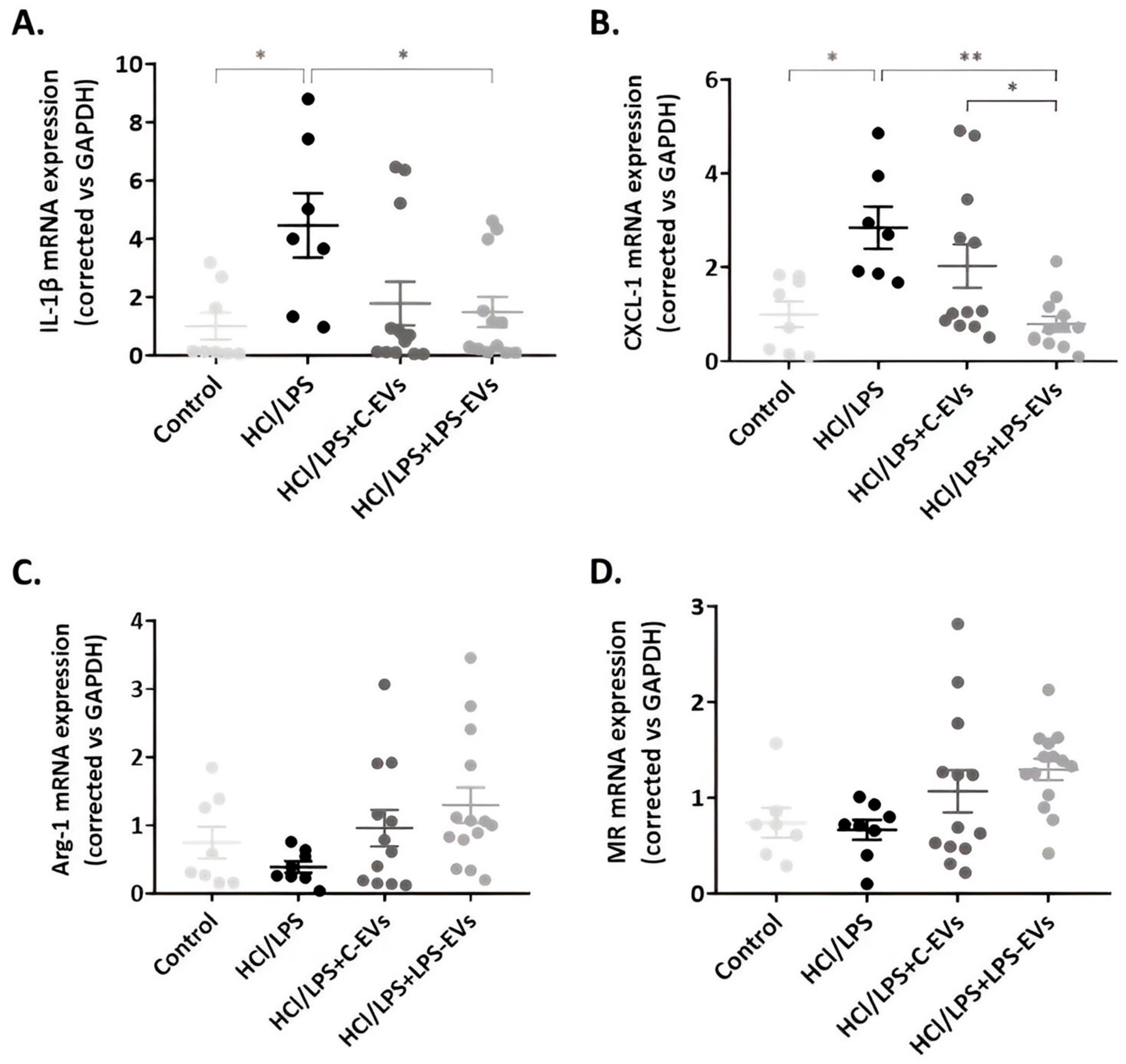
3.7. MSCs-EVs Modulate Lung Inflammation but LPS-EVs Further Decrease AM’s Chemotaxis Activity In Vivo
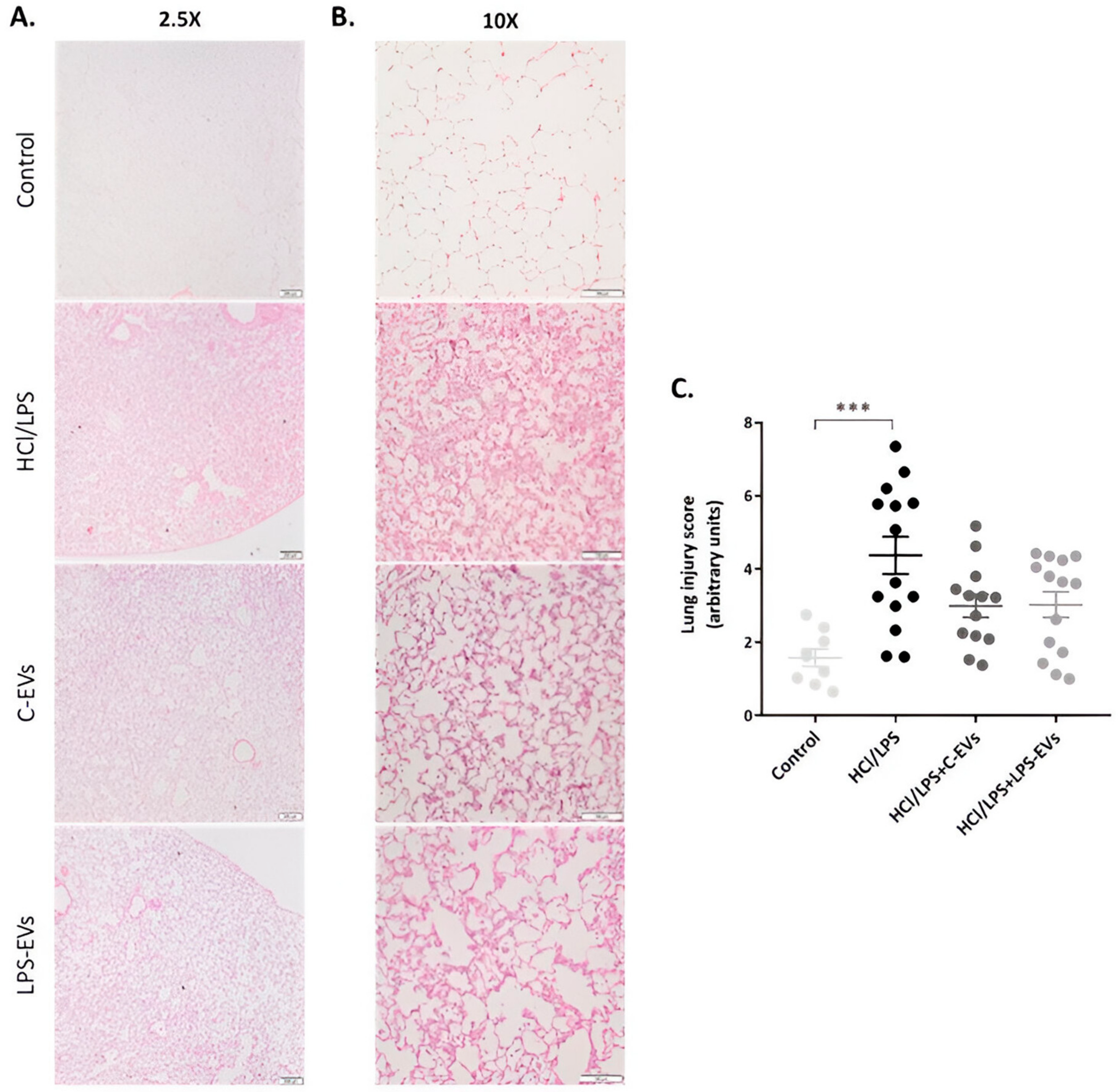
3.8. C-EVs and LPS-EVs Restored Lung Architecture In Vivo
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MSCs | Mesenchymal Stem Cells |
| EVs | Extracellular vesicles |
| ARDS | Acute Respiratory Distress Syndrome |
| LPS | Lipopolysaccharide |
| HCl | Hydrochloric acid |
| ALI | Acute Lung Injury |
| TLR4 | Toll-like receptor 4 |
| DMEM | Dulbecco’s Modified Eagle Medium |
| FBS | Fetal bovine serum |
| PBS | Phosphate-buffered saline |
| C-EVs | Control-extracellular vesicles |
| LPS-EVs | LPS-extracellular vesicles |
| Cryo-TEM | Cryo-transmission electron microscopy |
| HPAEpiC | Human Pulmonary Alveolar Epithelial cells |
| THP-1 | Human Leukemia Monocytic cell line |
| RPMI | Roswell Park Memorial Institute medium |
| PA | Pseudomonas aeruginosa |
| LB | Luria Broth |
| IL | Interleukin |
| RT-qPCR | Real-time quantitative reverse transcription polymerase chain reaction |
| NTA | Nanoparticle tracking analysis |
| BAL | Bronchoalveolar lavage |
| ACK | Ammonium chloride potassium |
| AM | Alveolar macrophages |
| H&E | Hematoxylin-eosin |
| LIS | Lung injury score |
| Arg-1 | Arginase-1 |
| MR | Mannose receptor |
| MPO | Myeloperoxidase |
| NETs | Neutrophil extracellular traps |
| HRP | Horseradish peroxidase |
| SEM | Standard error of the mean |
| p | p-value |
References
- Bos, L.D.J.; Ware, L.B. Acute respiratory distress syndrome: Causes, pathophysiology, and phenotypes. Lancet 2022, 400, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Bellani, G.; Laffey, J.G.; Pham, T.; Fan, E.; Brochard, L.; Esteban, A.; Gattinoni, L.; van Haren, F.; Larsson, A.; McAuley, D.F.; et al. Epidemiology, patterns of care, and mortality for patients with acute respiratory distress syndrome in intensive care units in 50 countries. JAMA—J. Am. Med. Assoc. 2016, 315, 788–800. [Google Scholar] [CrossRef] [PubMed]
- Laffey, J.G.; Matthay, M.A. Cell-based therapy for acute respiratory distress syndrome: Biology and potential therapeutic value. Am. J. Respir. Crit. Care Med. 2017, 196, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Heijnen, N.F.L.; Hagens, L.A.; Smit, M.R.; Schultz, M.J.; van der Poll, T.; Schnabel, R.M.; van der Horst, I.C.C.; Dickson, R.P.; Bergmans, D.C.J.J.; Bos, L.D.J.; et al. Biological subphenotypes of acute respiratory distress syndrome may not reflect differences in alveolar inflammation. Physiol. Rep. 2021, 9, e14693. [Google Scholar] [CrossRef] [PubMed]
- Guillamat-Prats, R.; Camprubí-Rimblas, M.; Bringué, J.; Tantinyà, N.; Artigas, A. Cell therapy for the treatment of sepsis and acute respiratory distress syndrome. Ann. Transl. Med. 2017, 5, 446. [Google Scholar] [CrossRef]
- McIntyre, L.A.; Moher, D.; Fergusson, D.A.; Sullivan, K.J.; Mei, S.H.; Lalu, M.; Marshall, J.; Mcleod, M.; Griffin, G.; Grimshaw, M.; et al. Efficacy of mesenchymal stromal cell therapy for acute lung injury in preclinical animal models: A systematic review. PLoS ONE 2016, 11, e0147170. [Google Scholar] [CrossRef]
- Guillamat-Prats, R.; Camprubí-Rimblas, M.; Puig, F.; Herrero, R.; Tantinyà, N.; Serrano-Mollar, A.; Artigas, A. Alveolar Type II Cells or Mesenchymal Stem Cells: Comparison of Two Different Cell Therapies for the Treatment of Acute Lung Injury in Rats. Cells 2020, 9, 1816. [Google Scholar] [CrossRef]
- Curley, G.F.; O’Kane, C.M.; McAuley, D.F.; Matthay, M.A.; Laffey, J.G. Cell-based Therapies for ARDS—Where Are We Now? Am. J. Respir. Crit. Care Med. 2024, 209, 789–797. [Google Scholar] [CrossRef]
- Liang, D.; Liu, C.; Yang, M. Mesenchymal stem cells and their derived exosomes for ALI/ARDS: A promising therapy. Heliyon 2023, 9, e20387. [Google Scholar] [CrossRef]
- Areny-Balagueró, A.; Solé-Porta, A.; Camprubí-Rimblas, M.; Campaña-Duel, E.; Ceccato, A.; Roig, A.; Closa, D.; Artigas, A. Bioengineered extracellular vesicles: Future of precision medicine for sepsis. Intensive Care Med. Exp. 2023, 11, 11. [Google Scholar] [CrossRef]
- Morrison, T.J.; Jackson, M.V.; Cunningham, E.K.; Kissenpfennig, A.; McAuley, D.F.; O’Kane, C.M.; Krasnodembskaya, A.D. Mesenchymal stromal cells modulate macrophages in clinically relevant lung injury models by extracellular vesicle mitochondrial transfer. Am. J. Respir. Crit. Care Med. 2017, 196, 1275–1286. [Google Scholar] [PubMed]
- Dutra Silva, J.; Su, Y.; Calfee, C.S.; Delucchi, K.L.; Weiss, D.; McAuley, D.F.; O’Kane, C.; Krasnodembskaya, A.D. Mesenchymal stromal cell extracellular vesicles rescue mitochondrial dysfunction improve barrier integrity in clinically relevant models of, A.R.D.S. Eur. Respir. J. 2021, 58, 2002978. [Google Scholar] [CrossRef]
- Alagesan, S.; Brady, J.; Byrnes, D.; Fandiño, J.; Masterson, C.; McCarthy, S.; Laffey, J.; O’Toole, D. Enhancement strategies for mesenchymal stem cells and related therapies. Stem Cell Res. Ther. 2022, 13, 75. [Google Scholar] [CrossRef] [PubMed]
- Kusuma, G.D.; Carthew, J.; Lim, R.; Frith, J.E. Effect of the Microenvironment on Mesenchymal Stem Cell Paracrine Signaling: Opportunities to Engineer the Therapeutic Effect. Stem Cells Dev. 2017, 26, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Crisostomo, P.R.; Wang, Y.; Markel, T.A.; Wang, M.; Lahm, T.; Meldrum, D.R. Human mesenchymal stem cells stimulated by TNF-α, LPS, or hypoxia produce growth factors by an NFκB- but not JNK-dependent mechanism. Am. J. Physiol.-Cell Physiol. 2008, 294, 675–682. [Google Scholar]
- de Jong, O.G.; Verhaar, M.C.; Chen, Y.; Vader, P.; Gremmels, H.; Posthuma, G.; Schiffelers, R.M.; Gucek, M.; van Balkom, B.W. Cellular stress conditions are reflected in the protein and RNA content of endothelial cell-derived exosomes. J. Extracell. Vesicles 2012, 1, 18396. [Google Scholar]
- Hwang, S.; Sung, D.K.; Kim, Y.E.; Yang, M.; Ahn, S.Y.; Sung, S.I.; Chang, Y.S. Mesenchymal Stromal Cells Primed by Toll-like Receptors 3 and 4 Enhanced Anti-Inflammatory Effects against LPS-Induced Macrophages via Extracellular Vesicles. Int. J. Mol. Sci. 2023, 24, 16264. [Google Scholar] [CrossRef]
- Zeuner, M.; Bieback, K.; Widera, D. Controversial role of toll-like receptor 4 in adult stem cells. Stem Cell Rev. Rep. 2015, 11, 621–634. [Google Scholar] [CrossRef]
- Akashi, S.; Saitoh, S.; Wakabayashi, Y.; Kikuchi, T.; Takamura, N.; Nagai, Y.; Kusumoto, Y.; Fukase, K.; Kusumoto, S.; Adachi, Y.; et al. Lipopolysaccharide interaction with cell surface toll-like receptor 4-MD-2: Higher affinity than that with MD-2 or CD14. J. Exp. Med. 2003, 198, 1035–1042. [Google Scholar]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3.22.1–3.22.29. [Google Scholar]
- Arraud, N.; Linares, R.; Tan, S.; Gounou, C.; Pasquet, J.M.; Mornet, S.; Brisson, A.R. Extracellular vesicles from blood plasma: Determination of their morphology, size, phenotype and concentration. J. Thromb. Haemost. 2014, 12, 614–627. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.A.; Goberdhan, D.C.I.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.P.; Erdbrügger, U.; et al. Minimal information for studies of extracellular vesicles (MISEV2023): From basic to advanced approaches. J. Extracell. Vesicles 2024, 13, e12404. [Google Scholar] [PubMed]
- Puig, F.; Herrero, R.; Guillamat-Prats, R.; Gómez, M.N.; Tijero, J.; Chimenti, L.; Stelmakh, O.; Blanch, L.; Serrano-Mollar, A.; Matthay, M.A.; et al. A new experimental model of acid-and endotoxin-induced acute lung injury in rats. Am. J. Physiol.-Lung Cell Mol. Physiol. 2016, 311, 229–237. [Google Scholar]
- Matute-Bello, G.; Downey, G.; Moore, B.B.; Groshong, S.D.; Matthay, M.A.; Slutsky, A.S.; Kuebler, W.M. An official american thoracic society workshop report: Features and measurements of experimental acute lung injury in animals. Am. J. Respir. Cell Mol. Biol. 2011, 44, 725–738. [Google Scholar]
- Zhai, R.; Bonda, W.L.M.; Matute-Bello, G.; Jabaudon, M. From preclinical to clinical models of acute respiratory distress syndrome. Signa Vitae 2022, 18, 3–14. [Google Scholar]
- Kulkarni, H.S.; Lee, J.S.; Bastarache, J.A.; Kuebler, W.M.; Downey, G.P.; Albaiceta, G.M.; Altemeier, W.A.; Artigas, A.; Bates, J.H.T.; Calfee, C.S.; et al. Update on the Features and Measurements of Experimental Acute Lung Injury in Animals an Official American Thoracic Society Workshop Report. Am. J. Respir. Cell Mol. Biol. 2022, 66, E1–E14. [Google Scholar]
- Zhang, J.; Lu, Y.; Mao, Y.; Yu, Y.; Wu, T.; Zhao, W.; Zhu, Y.; Zhao, P.; Zhang, F. IFN-γ enhances the efficacy of mesenchymal stromal cell-derived exosomes via miR-21 in myocardial infarction rats. Stem Cell Res. Ther. 2022, 13, 333. [Google Scholar] [CrossRef]
- Kim, M.; Shin Dil Choi, B.H.; Min, B.H. Exosomes from IL-1β-Primed Mesenchymal Stem Cells Inhibited IL-1β- and TNF-α-Mediated Inflammatory Responses in Osteoarthritic SW982 Cells. Tissue Eng. Regen. Med. 2021, 18, 525–536. [Google Scholar] [CrossRef]
- Ge, L.; Xun, C.; Li, W.; Jin, S.; Liu, Z.; Zhuo, Y.; Duan, D.; Hu, Z.; Chen, P.; Lu, M. Extracellular vesicles derived from hypoxia-preconditioned olfactory mucosa mesenchymal stem cells enhance angiogenesis via miR-612. J. Nanobiotechnol. 2021, 19, 380. [Google Scholar] [CrossRef]
- Miceli, V.; Zito, G.; Bulati, M.; Gallo, A.; Busà, R.; Iannolo, G.; Conaldi, P. Different priming strategies improve distinct therapeutic capabilities of mesenchymal stromal/stem cells: Potential implications for their clinical use. World J. Stem Cells 2023, 15, 400–420. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Lee, S.K.; Park, A.; Lee, J.; Jung, I.; Song, K.B.; Choi, E.J.; Kim, S.; Yu, J. Exosome from IFN-γ-Primed Induced Pluripotent Stem Cell-Derived Mesenchymal Stem Cells Improved Skin Inflammation and Barrier Function. Int. J. Mol. Sci. 2023, 24, 11635. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.; Maughon, T.; Marklein, R.; Stice, S. Priming of MSCs with inflammation-relevant signals affects extracellular vesicle biogenesis, surface markers, and modulation of T cell subsets. J. Immunol. Regen. Med. 2021, 13, 100036. [Google Scholar] [CrossRef]
- Huang, Q.; Le, Y.; Li, S.; Bian, Y. Signaling pathways and potential therapeutic targets in acute respiratory distress syndrome (ARDS). Respir. Res. 2024, 25, 30. [Google Scholar] [CrossRef]
- Ding, J.Y.; Chen, M.J.; Wu, L.F.; Shu, G.F.; Fang, S.J.; Li, Z.Y.; Chu, X.R.; Li, X.K.; Wang, Z.G.; Ji, J.S. Mesenchymal Stem Cells-Derived Extracellular Vesicles as Nanotherapeutics: An Application for Skin Wound Healing. Dermatol. Ther. 2023, 2023, 7916795. [Google Scholar]
- Ti, D.; Hao, H.; Tong, C.; Liu, J.; Dong, L.; Zheng, J.; Zhao, Y.; Liu, H.; Fu, X.; Han, W. LPS-preconditioned mesenchymal stromal cells modify macrophage polarization for resolution of chronic inflammation via exosome-shuttled let-7b. J. Transl. Med. 2015, 13, 308. [Google Scholar] [CrossRef]
- Soler-Botija, C.; Monguió-Tortajada, M.; Munizaga-Larroudé, M.; Gálvez-Montón, C.; Bayes-Genis, A.; Roura, S. Mechanisms governing the therapeutic effect of mesenchymal stromal cell-derived extracellular vesicles: A scoping review of preclinical evidence. Biomed. Pharmacother. 2022, 147, 112683. [Google Scholar] [CrossRef]
- Forsberg, M.H.; Kink, J.A.; Thickens, A.S.; Lewis, B.M.; Childs, C.J.; Hematti, P.; Capitini, C.M. Exosomes from primed MSCs can educate monocytes as a cellular therapy for hematopoietic acute radiation syndrome. Stem Cell Res. Ther. 2021, 12, 459. [Google Scholar] [CrossRef]
- Smith, T.D.; Tse, M.J.; Read, E.L.; Liu, W.F. Regulation of macrophage polarization and plasticity by complex activation signals. Integr. Biol. 2016, 8, 946–955. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5148158/pdf/nihms-808608.pdf (accessed on 6 May 2024).
- Huang, X.; Xiu, H.; Zhang, S.; Zhang, G. The role of macrophages in the pathogenesis of ALI/ARDS. Mediat. Inflamm. 2018, 2018, 1264913. [Google Scholar] [CrossRef]
- Nordlohne, J.; Hulsmann, I.; Schwafertz, S.; Zgrajek, J.; Grundmann, M.; von Vietinghoff, S.; Eitner, F.; Becker, M.S. A flow cytometry approach reveals heterogeneity in conventional subsets of murine renal mononuclear phagocytes. Sci. Rep. 2021, 11, 13251. [Google Scholar] [CrossRef]
- Potter, D.R.; Miyazawa, B.Y.; Gibb, S.L.; Deng, X.; Togaratti, P.P.; Croze, R.H.; Srivastava, A.K.; Trivedi, A.; Matthay, M.; Holcomb, J.B.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles Attenuate Pulmonary Vascular Permeability and Lung Injury Induced by Hemorrhagic Shock and Trauma. Physiol. Behav. 2016, 176, 139–148. [Google Scholar]
- Yi, X.; Wei, X.; Lv, H.; An, Y.; Li, L.; Lu, P.; Yang, Y.; Zhang, Q.; Yi, H.; Chen, G. Exosomes derived from microRNA-30b-3p-overexpressing mesenchymal stem cells protect against lipopolysaccharide-induced acute lung injury by inhibiting SAA3. Exp. Cell Res. 2019, 383, 111454. [Google Scholar] [CrossRef] [PubMed]
- Varkouhi, A.K.; Jerkic, M.; Ormesher, L.; Gagnon, S.; Goyal, S.; Rabani, R.; Masterson, C.; Spring, C.; Chen, P.Z.; Gu, F.X.; et al. Extracellular Vesicles from Interferon-γ–primed Human Umbilical Cord Mesenchymal Stromal Cells Reduce Escherichia coli–induced Acute Lung Injury in Rats. Anesthesiology 2019, 130, 778–790. [Google Scholar] [PubMed]
- Munir, S.; Basu, A.; Maity, P.; Krug, L.; Haas, P.; Jiang, D.; Strauss, G.; Wlaschek, M.; Geiger, H.; Singh, K.; et al. TLR4-dependent shaping of the wound site by MSCs accelerates wound healing. EMBO Rep. 2020, 21, e48777. [Google Scholar] [PubMed]
- Metzler, K.D.; Fuchs, T.A.; Nauseef, W.M.; Reumaux, D.; Roesler, J.; Schulze, I.; Wahn, V.; Papayannopoulos, V.; Zychlinsky, A. Myeloperoxidase is required for neutrophil extracellular trap formation: Implications for innate immunity. Blood 2011, 117, 953–959. [Google Scholar] [PubMed]
- Potey, P.M.D.; Rossi, A.G.; Lucas, C.D.; Dorward, D.A. Neutrophils in the initiation and resolution of acute pulmonary inflammation: Understanding biological function and therapeutic potential. J. Pathol. 2019, 247, 672–685. [Google Scholar] [CrossRef]
- Castanheira, F.V.S.; Kubes, P. Neutrophils and NETs in modulating acute and chronic inflammation. Blood 2019, 133, 2178–2185. [Google Scholar]
- Wang, X.; Adler, K.B.; Erjefalt, J.; Bai, C. Airway epithelial dysfunction in the development of acute lung injury and acute respiratory distress syndrome. Expert. Rev. Respir. Med. 2007, 1, 149–155. [Google Scholar]
- Barilli, A.; Visigalli, R.; Ferrari, F.; Bianchi, M.G.; Dall’Asta, V.; Rotoli, B.M. Immune-Mediated Inflammatory Responses of Alveolar Epithelial Cells: Implications for COVID-19 Lung Pathology. Biomedicines 2022, 10, 618. [Google Scholar] [CrossRef]
- Camprubí-Rimblas, M.; Tantinyà, N.; Guillamat-Prats, R.; Bringué, J.; Puig, F.; Gómez, M.N.; Blanch, L.; Artigas, A. Effects of nebulized antithrombin and heparin on inflammatory and coagulation alterations in an acute lung injury model in rats. J. Thromb. Haemost. 2020, 18, 571–583. [Google Scholar] [CrossRef]
- Chimenti, L.; Camprubí-Rimblas, M.; Guillamat-Prats, R.; Gomez, M.N.; Tijero, J.; Blanch, L.; Artigas, A. Nebulized heparin attenuates pulmonary coagulopathy and inflammation through alveolar macrophages in a rat model of acute lung injury. Thromb. Haemost. 2017, 117, 2125–2134. [Google Scholar] [PubMed]
- Schouten, L.R.A.; Bos, L.D.J.; Serpa Neto, A.; van Vught, L.A.; Wiewel, M.A.; Hoogendijk, A.J.; Bonten, M.J.M.; Cremer, O.L.; Horn, J.; van der Poll, T.; et al. Increased mortality in elderly patients with acute respiratory distress syndrome is not explained by host response. Intensive Care Med. Exp. 2019, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Heffernan, D.S.; Dossett, L.A.; Lightfoot, M.A.; Fremont, R.D.; Ware, L.B.; Sawyer, R.G.; May, A.K. Gender and ARDS in Critically Injured Adults: A Prospective Study. J. Trauma 2011, 71, 878–885. [Google Scholar] [PubMed]
- Horie, S.; Masterson, C.; Devaney, J.; Laffey, J.G. Stem cell therapy for acute respiratory distress syndrome: A promising future? Curr. Opin. Crit. Care 2016, 22, 14–20. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Areny-Balagueró, A.; Camprubí-Rimblas, M.; Campaña-Duel, E.; Solé-Porta, A.; Ceccato, A.; Roig, A.; Laffey, J.G.; Closa, D.; Artigas, A. Priming Mesenchymal Stem Cells with Lipopolysaccharide Boosts the Immunomodulatory and Regenerative Activity of Secreted Extracellular Vesicles. Pharmaceutics 2024, 16, 1316. https://doi.org/10.3390/pharmaceutics16101316
Areny-Balagueró A, Camprubí-Rimblas M, Campaña-Duel E, Solé-Porta A, Ceccato A, Roig A, Laffey JG, Closa D, Artigas A. Priming Mesenchymal Stem Cells with Lipopolysaccharide Boosts the Immunomodulatory and Regenerative Activity of Secreted Extracellular Vesicles. Pharmaceutics. 2024; 16(10):1316. https://doi.org/10.3390/pharmaceutics16101316
Chicago/Turabian StyleAreny-Balagueró, Aina, Marta Camprubí-Rimblas, Elena Campaña-Duel, Anna Solé-Porta, Adrián Ceccato, Anna Roig, John G. Laffey, Daniel Closa, and Antonio Artigas. 2024. "Priming Mesenchymal Stem Cells with Lipopolysaccharide Boosts the Immunomodulatory and Regenerative Activity of Secreted Extracellular Vesicles" Pharmaceutics 16, no. 10: 1316. https://doi.org/10.3390/pharmaceutics16101316
APA StyleAreny-Balagueró, A., Camprubí-Rimblas, M., Campaña-Duel, E., Solé-Porta, A., Ceccato, A., Roig, A., Laffey, J. G., Closa, D., & Artigas, A. (2024). Priming Mesenchymal Stem Cells with Lipopolysaccharide Boosts the Immunomodulatory and Regenerative Activity of Secreted Extracellular Vesicles. Pharmaceutics, 16(10), 1316. https://doi.org/10.3390/pharmaceutics16101316