

Cis-2-Decenoic Acid and Bupivacaine Delivered from Electrospun Chitosan Membranes Increase Cytokine Production in Dermal and Inflammatory Cell Lines
 , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Membrane Fabrication
2.3. Cytocompatibility
2.4. NHDF Collagen Production
2.5. Cytokine Production
2.6. Statistical Analysis
3. Results
3.1. Normal Adult Human Dermal Fibroblasts (NHDFs)
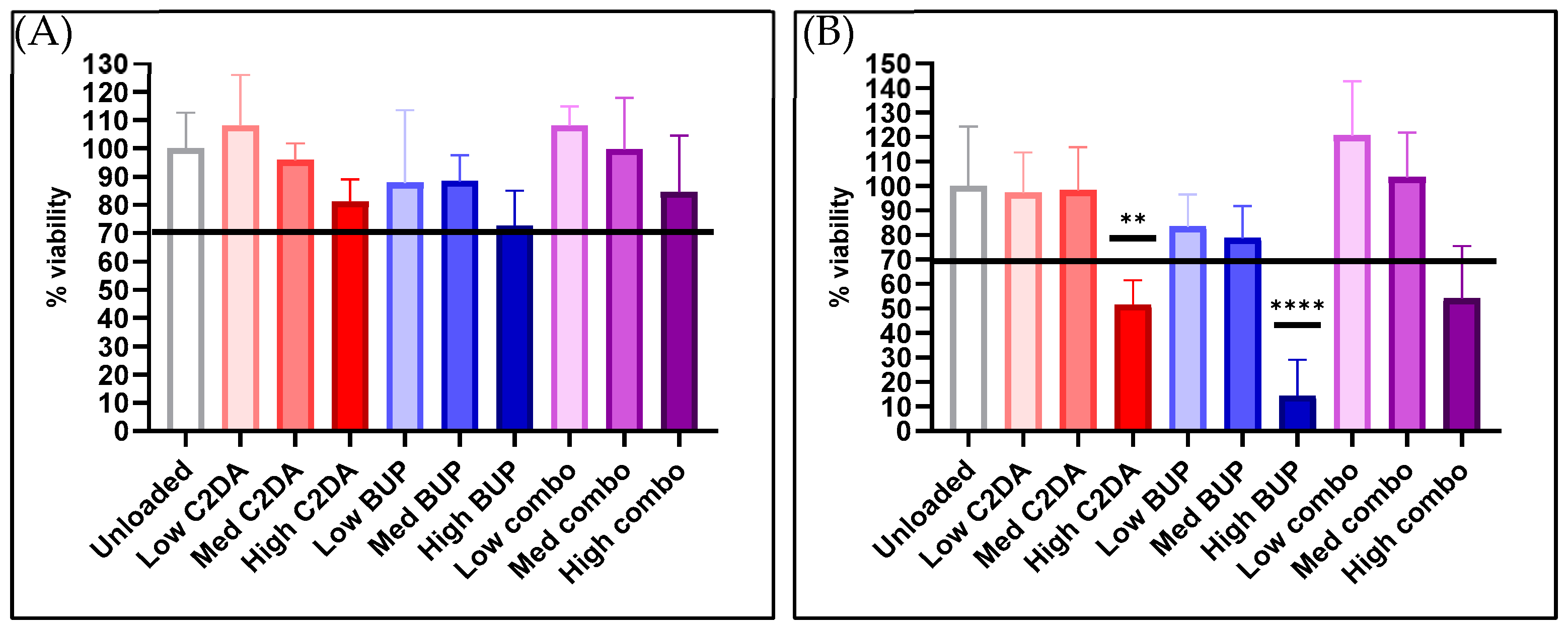
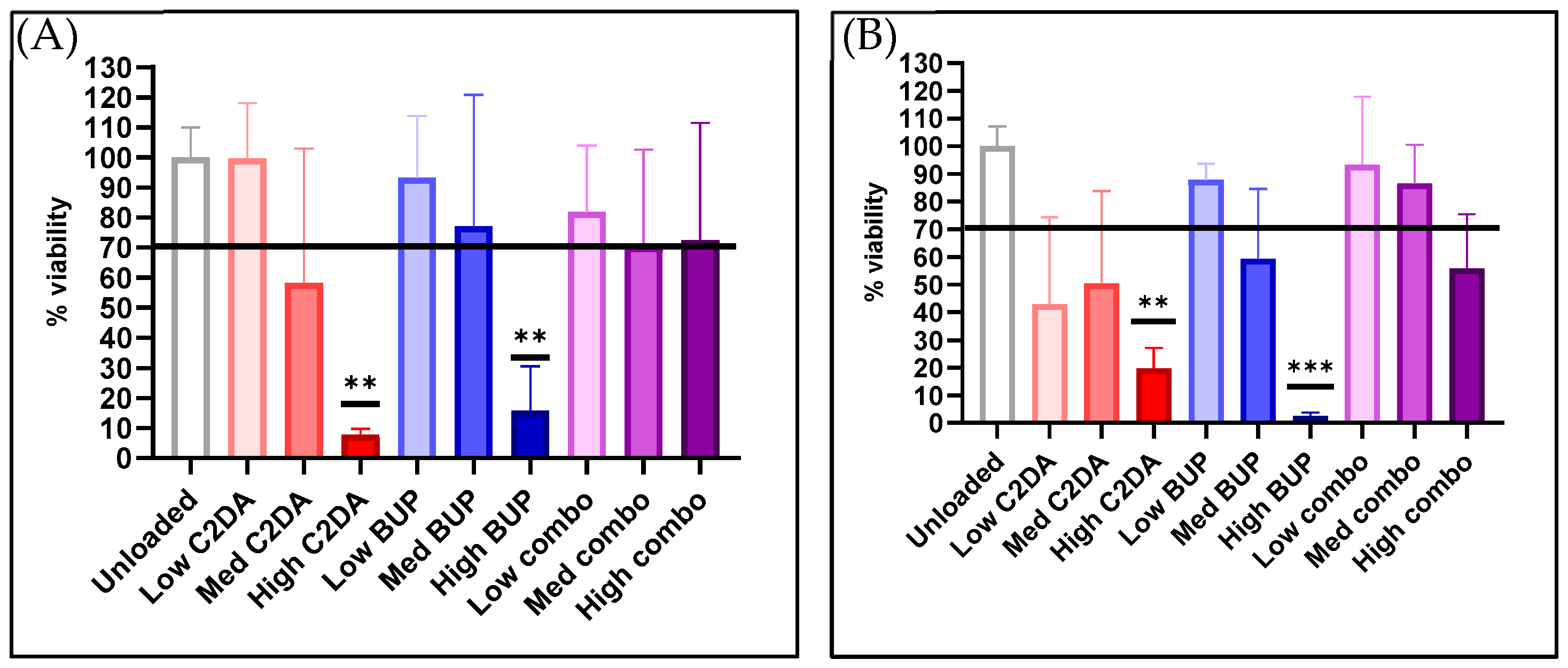
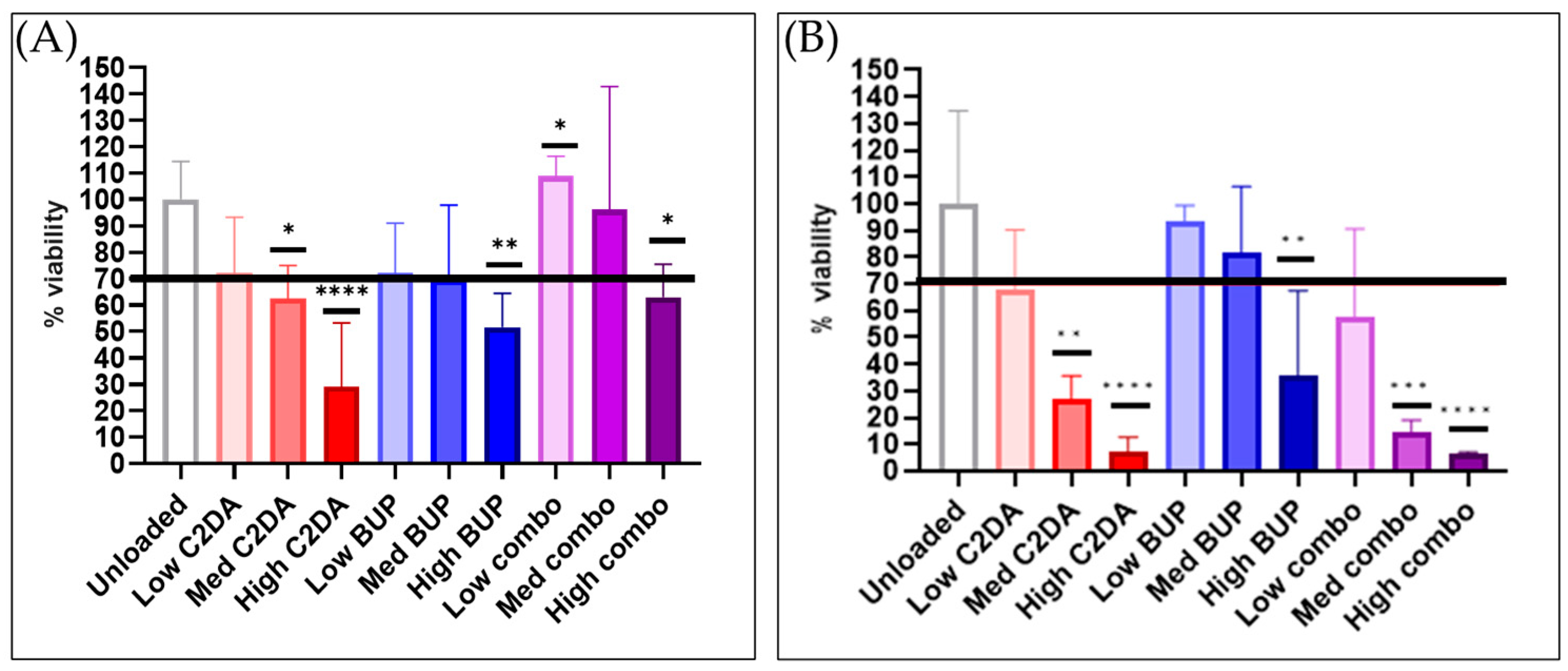
3.1.1. Cytocompatibility
3.1.2. Collagen Production
3.2. Normal Adult Human Epidermal Keratinocytes (NHEKs)
3.2.1. Cytocompatibility
3.2.2. Cytokine Production
3.3. RAW 264.7 Monocytes
3.3.1. Cytocompatibility
3.3.2. Cytokine Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gobi, R.; Ravichandiran, P.; Babu, R.S.; Yoo, D.J. Biopolymer and synthetic polymer-based nanocomposites in wound dressing applications: A review. Polymers 2021, 13, 1962. [Google Scholar] [CrossRef]
- Sahana, T.; Rekha, P. Biopolymers: Applications in wound healing and skin tissue engineering. Mol. Biol. Rep. 2018, 45, 2857–2867. [Google Scholar] [CrossRef]
- Weller, C.D.; Team, V.; Sussman, G. First-line interactive wound dressing update: A comprehensive review of the evidence. Front. Pharmacol. 2020, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, N.; Venkataraman, S.S.; Daniel, R.; Aravind, R.J.; Kumarakrishnan, V.B. Molecular biology of wound healing. J. Pharm. Bioallied Sci. 2012, 4, S334. [Google Scholar] [CrossRef] [PubMed]
- Darby, I.A.; Laverdet, B.; Bonté, F.; Desmoulière, A. Fibroblasts and myofibroblasts in wound healing. Clin. Cosmet. Investig. Dermatol. 2014, 7, 301. [Google Scholar]
- Ghahary, A.; Ghaffari, A. Role of keratinocyte–fibroblast cross-talk in development of hypertrophic scar. Wound Repair Regen. 2007, 15, S46–S53. [Google Scholar] [CrossRef]
- Jiang, Y.; Tsoi, L.C.; Billi, A.C.; Ward, N.L.; Harms, P.W.; Zeng, C.; Maverakis, E.; Kahlenberg, J.M.; Gudjonsson, J.E. Cytokinocytes: The diverse contribution of keratinocytes to immune responses in skin. JCI Insight 2020, 5, e142067. [Google Scholar] [CrossRef]
- Jennings, J.A.; Bumgardner, J.D. Chitosan Based Biomaterials Volume 1: Fundamentals; Woodhead Publishing: Sawston, UK, 2016. [Google Scholar]
- Jennings, J.A.; Beenken, K.E.; Parker, A.C.; Smith, J.K.; Courtney, H.S.; Smeltzer, M.S.; Haggard, W.O. Polymicrobial biofilm inhibition effects of acetate-buffered chitosan sponge delivery device. Macromol. Biosci. 2016, 16, 591–598. [Google Scholar] [CrossRef]
- Boles, L.; Alexander, C.; Pace, L.; Haggard, W.; Bumgardner, J.; Jennings, J. Development and Evaluation of an Injectable Chitosan/β-Glycerophosphate Paste as a Local Antibiotic Delivery System for Trauma Care. J. Funct. Biomater. 2018, 9, 56. [Google Scholar] [CrossRef]
- Harrison, Z.L.; Pace, L.R.; Brown, M.N.; Beenken, K.E.; Smeltzer, M.S.; Bumgardner, J.D.; Haggard, W.O.; Amber Jennings, J. Staphylococcal infection prevention using antibiotic-loaded mannitol–chitosan paste in a rabbit model of implant-associated osteomyelitis. J. Orthop. Res. 2021, 39, 2455–2464. [Google Scholar] [CrossRef]
- Pace, L.R.; Harrison, Z.L.; Brown, M.N.; Haggard, W.O.; Jennings, J.A. Characterization and anti-biofilm activity of mannitol-chitosan blended paste for local antibiotic delivery system. Mar. Drugs 2019, 17, 517. [Google Scholar]
- Harrison, Z.L.; Bumgardner, J.D.; Fujiwara, T.; Baker, D.L.; Jennings, J.A. In vitro evaluation of loaded chitosan membranes for pain relief and infection prevention. J. Biomed. Mater. Res. Part B Appl. Biomater. 2021, 109, 1735–1743. [Google Scholar] [CrossRef] [PubMed]
- Augustine, R.; Rehman, S.; Ahmed, R.; Zahid, A.; Sharifi, M.; Falahati, M.; Hasan, A. Electrospun chitosan membranes containing bioactive and therapeutic agents for enhanced wound healing. Int. J. Biol. Mol. 2020, 156, 153–170. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Wu, T.; Dai, Y.; Xia, Y. Electrospinning and electrospun nanofibers: Methods, materials, and applications. Chem. Rev. 2019, 119, 5298–5415. [Google Scholar] [CrossRef]
- Wu, C.; Su, H.; Tang, S.; Bumgardner, J.D. The stabilization of electrospun chitosan nanofibers by reversible acylation. Cellulose 2014, 21, 2549–2556. [Google Scholar] [CrossRef]
- Moore, D.C.; Bridenbaugh, L.D.; Thompson, G.E.; Balfour, R.I.; Horton, W.G. Bupivacaine: A review of 11,080 cases. Anesth. Analg. 1978, 57, 42–53. [Google Scholar] [CrossRef]
- Johnson, S.M.; Saint John, B.E.; Dine, A.P. Local anesthetics as antimicrobial agents: A review. Surg. Infect. 2008, 9, 205–213. [Google Scholar] [CrossRef]
- Marques, C.; Davies, D.; Sauer, K. Control of biofilms with the fatty acid signaling molecule cis-2-decenoic acid. Pharmaceuticals 2015, 8, 816–835. [Google Scholar] [CrossRef]
- Choi, L.R. In Vitro Evaluation of Chitosan Membranes Stabilized with Varying Acyl Lengths for Release of Therapeutics for Burn Wound Coverage, Infection Prevention, and Pain Relief; The University of Memphis: Memphis, TN, USA, 2022. [Google Scholar]
- Harrison, Z.L.; Awais, R.; Harris, M.; Raji, B.; Hoffman, B.C.; Baker, D.L.; Jennings, J.A. 2-Heptylcyclopropane-1-Carboxylic Acid Disperses and Inhibits Bacterial Biofilms. Front. Microbiol. 2021, 12, 1298. [Google Scholar] [CrossRef]
- ISO 10993-5; Biological Evaluation of Medical Devices—Part 5: Tests for In Vitro Cytotoxicity. ISO: Geneva, Switzerland, 2009.
- Chun, J.Y.; Kang, H.K.; Jeong, L.; Kang, Y.O.; Oh, J.-E.; Yeo, I.-S.; Jung, S.Y.; Park, W.H.; Min, B.-M. Epidermal cellular response to poly (vinyl alcohol) nanofibers containing silver nanoparticles. Colloids Surf. B Biointerfaces 2010, 78, 334–342. [Google Scholar] [CrossRef]
- Light, C.J. Cis-2-Decenoic Acid Signaling and Dispersion in Pseudomonas aeruginosa: The Role of Dispersed Cells in Virulence and Pathogenesis; State University of New York at Binghamton: Binghamton, NY, USA, 2017. [Google Scholar]
- Jennings, J.A.; Courtney, H.S.; Haggard, W.O. Cis-2-decenoic acid inhibits S. aureus growth and biofilm in vitro: A pilot study. Clin. Orthop. Relat. Res. 2012, 470, 2663–2670. [Google Scholar] [CrossRef] [PubMed]
- Harrison, Z.L. In Vitro Evaluation of Loaded Chitosan Membranes for Pain Relief and Infection Prevention; University of Memphis: Memphis, TN, USA, 2020. [Google Scholar]
- Böhmig, G.; Krieger, P.M.; Säemann, M.; Wenhardt, C.; Pohanka, E.; Zlabinger, G. n-Butyrate downregulates the stimulatory function of peripheral blood-derived antigen-presenting cells: A potential mechanism for modulating T-cell responses by short-chain fatty acids. Immunology 1997, 92, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Fedder, C.; Beck-Schimmer, B.; Aguirre, J.; Hasler, M.; Roth-Z’graggen, B.; Urner, M.; Kalberer, S.; Schlicker, A.; Votta-Velis, G.; Bonvini, J. In vitro exposure of human fibroblasts to local anaesthetics impairs cell growth. Clin. Exp. Immunol. 2010, 162, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.L.; Bainbridge, N.J.; Jordan, N.R.; Sharpe, J.R. The effect of topical analgesics on ex vivo skin growth and human keratinocyte and fibroblast behavior. Wound Repair Regen. 2009, 17, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Gray, A.; Marrero-Berrios, I.; Weinberg, J.; Manchikalapati, D.; SchianodiCola, J.; Schloss, R.S.; Yarmush, J. The effect of local anesthetic on pro-inflammatory macrophage modulation by mesenchymal stromal cells. Int. Immunopharmacol. 2016, 33, 48–54. [Google Scholar] [CrossRef]
- Horiguchi, T.; Shibata, M.A.; Ito, Y.; Eid, N.A.; Abe, M.; Otsuki, Y. Macrophage apoptosis in rat skeletal muscle treated with bupivacaine hydrochloride: Possible role of MCP-1. Muscle Nerve Off. J. Am. Assoc. Electrodiagn. Med. 2002, 26, 79–86. [Google Scholar] [CrossRef]
- Tsai, K.; Shi, H.; Parungao, R.; Naficy, S.; Ding, X.; Ding, X.; Hew, J.; Wang, X.; Chrzanowski, W.; Lavery, G.; et al. Skin 11beta-hydroxysteroid dehydrogenase type 1 enzyme expression regulates burn would healing and can be targeted to modify scar characteristics. Burn. Trauma 2023, 11, 14. [Google Scholar] [CrossRef]
- Khaliq, T.; Sohail, M.; Minhas, M.; Mahmood, A.; Munir, A.; Qalawlus, A.; Jabeen, N.; Kousar, M.; Anwar, Z. Hyaluronic acid/alginate-based biomimetic hydrogel membranes for accelerated diabetic wound repair. Int. J. Pharm. 2023, 643, 123244. [Google Scholar] [CrossRef]
- Harris, M.; Hansen, R.; Vidsudhiphan, P.; Koslo, J.; Thomas, J.; Watkins, B.; Allen, K. Effects of conjugated linoleic acids and docosahexaenoic acid on rat liver and reproductive tissue fatty acids, prostaglandins and matrix metalloproteinase production. Prostaglandins Leukot. Essent. Fat. Acids 2001, 65, 23–29. [Google Scholar] [CrossRef]
- Hankenson, K.D.; Watkins, B.A.; Schoenlein, I.A.; Allen, K.G.; Turek, J.J. Omega-3 fatty acids enhance ligament fibroblast collagen formation in association with changes in interleukin-6 production (44467). Proc. Soc. Exp. Biol. Med. 2000, 223, 88–95. [Google Scholar] [CrossRef]
- Abrão, J.; Fernandes, C.R.; White, P.F.; Shimano, A.C.; Okubo, R.; Lima, G.B.; Bachur, J.A.; Garcia, S.B. Effect of local anaesthetic infiltration with bupivacaine and ropivacaine on wound healing: A placebo-controlled study. Int. Wound J. 2014, 11, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Gröne, A. Keratinocytes and cytokines. Vet. Immunol. Immunopathol. 2002, 88, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Enk, C.D.; Mahanty, S.; Blauvelt, A.; Katz, S.I. UVB induces IL-12 transcription in human keratinocytes in vivo and in vitro. Photochem. Photobiol. 1996, 63, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Hanada, K.; Hashimoto, I. The study of ultraviolet B-induced apoptosis in cultured mouse keratinocytes and in mouse skin. J. Dermatol. Sci. 1996, 12, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Kovach, T.K.; Dighe, A.S.; Lobo, P.I.; Cui, Q. Interactions between MSCs and Immune Cells: Implications for Bone Healing. J. Immunol. Res. 2015, 2015, 752510. [Google Scholar] [CrossRef]
- Dąbrowska, A.M.; Słotwiński, R. The immune response to surgery and infection. Cent.-Eur. J. Immunol. 2014, 39, 532. [Google Scholar] [CrossRef]
- Cavaillon, J.M. Pro-versus anti-inflammatory cytokines: Myth or reality. Cell. Mol. Biol. 2001, 47, 695–702. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C2DA Loading | BUP Loading | |
|---|---|---|
| Unloaded | - | - |
| Low C2DA | 0.075 mg | - |
| Med C2DA | 0.15 mg | - |
| High C2DA | 0.3 mg | - |
| Low BUP | - | 0.25 mg |
| Med BUP | - | 0.5 mg |
| High BUP | - | 1.0 mg |
| Low combo | 0.075 mg | 0.25 mg |
| Medium combo | 0.15 mg | 0.5 mg |
| High combo | 0.3 mg | 1.0 mg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harrison, Z.; Montgomery, E.C.; Bush, J.R.; Gupta, N.; Bumgardner, J.D.; Fujiwara, T.; Baker, D.L.; Jennings, J.A. Cis-2-Decenoic Acid and Bupivacaine Delivered from Electrospun Chitosan Membranes Increase Cytokine Production in Dermal and Inflammatory Cell Lines. Pharmaceutics 2023, 15, 2476. https://doi.org/10.3390/pharmaceutics15102476
Harrison Z, Montgomery EC, Bush JR, Gupta N, Bumgardner JD, Fujiwara T, Baker DL, Jennings JA. Cis-2-Decenoic Acid and Bupivacaine Delivered from Electrospun Chitosan Membranes Increase Cytokine Production in Dermal and Inflammatory Cell Lines. Pharmaceutics. 2023; 15(10):2476. https://doi.org/10.3390/pharmaceutics15102476
Chicago/Turabian StyleHarrison, Zoe, Emily C. Montgomery, Joshua R. Bush, Nidhi Gupta, Joel D. Bumgardner, Tomoko Fujiwara, Daniel L. Baker, and Jessica Amber Jennings. 2023. "Cis-2-Decenoic Acid and Bupivacaine Delivered from Electrospun Chitosan Membranes Increase Cytokine Production in Dermal and Inflammatory Cell Lines" Pharmaceutics 15, no. 10: 2476. https://doi.org/10.3390/pharmaceutics15102476
APA StyleHarrison, Z., Montgomery, E. C., Bush, J. R., Gupta, N., Bumgardner, J. D., Fujiwara, T., Baker, D. L., & Jennings, J. A. (2023). Cis-2-Decenoic Acid and Bupivacaine Delivered from Electrospun Chitosan Membranes Increase Cytokine Production in Dermal and Inflammatory Cell Lines. Pharmaceutics, 15(10), 2476. https://doi.org/10.3390/pharmaceutics15102476