
Honey: An Advanced Antimicrobial and Wound Healing Biomaterial for Tissue Engineering Applications

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
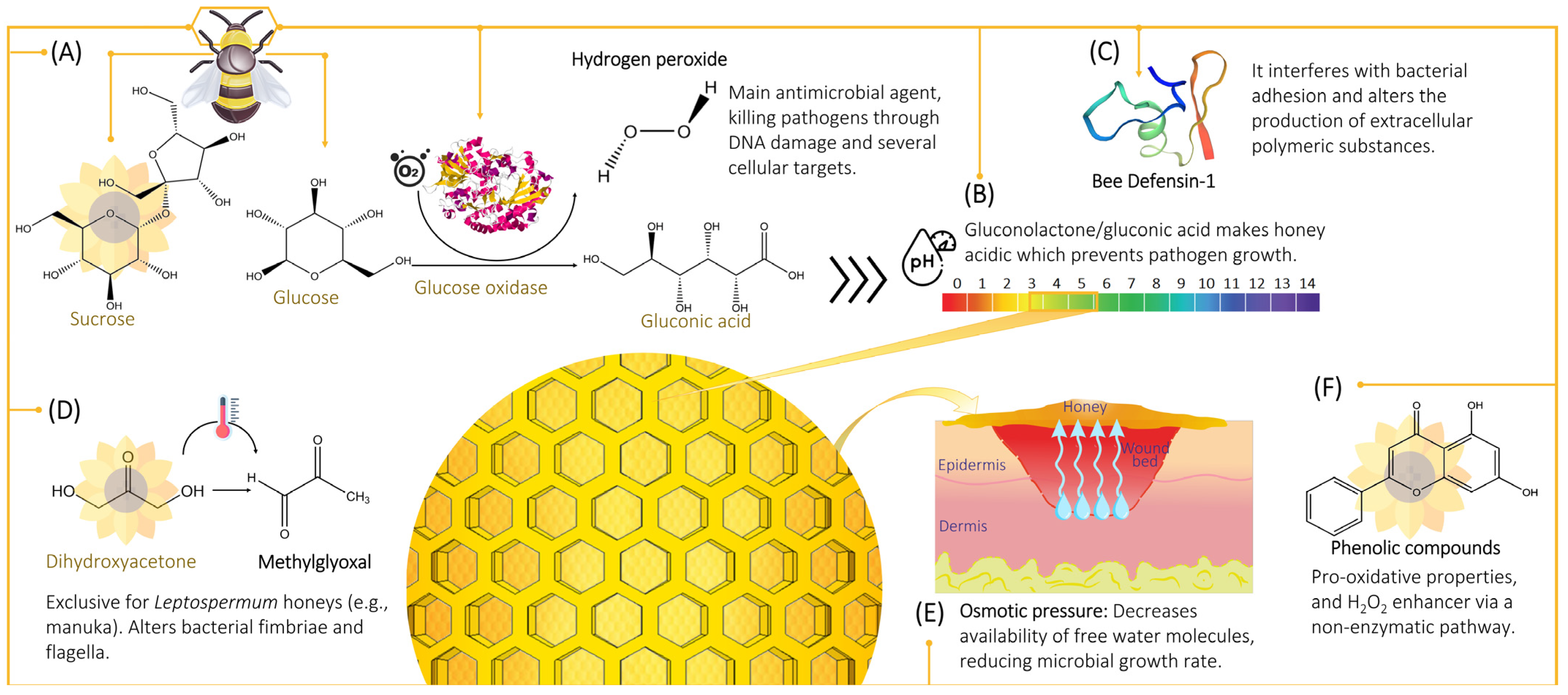
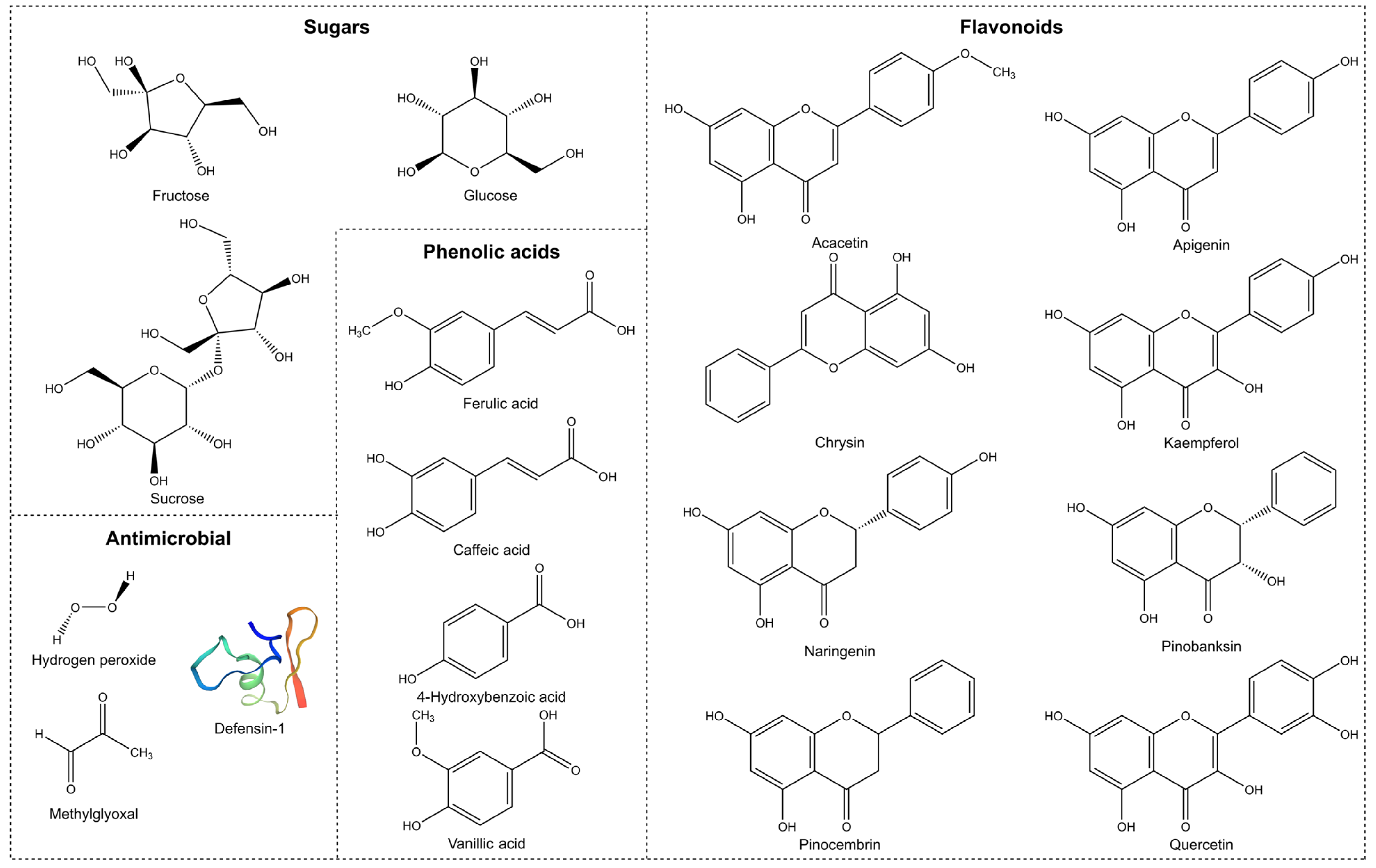
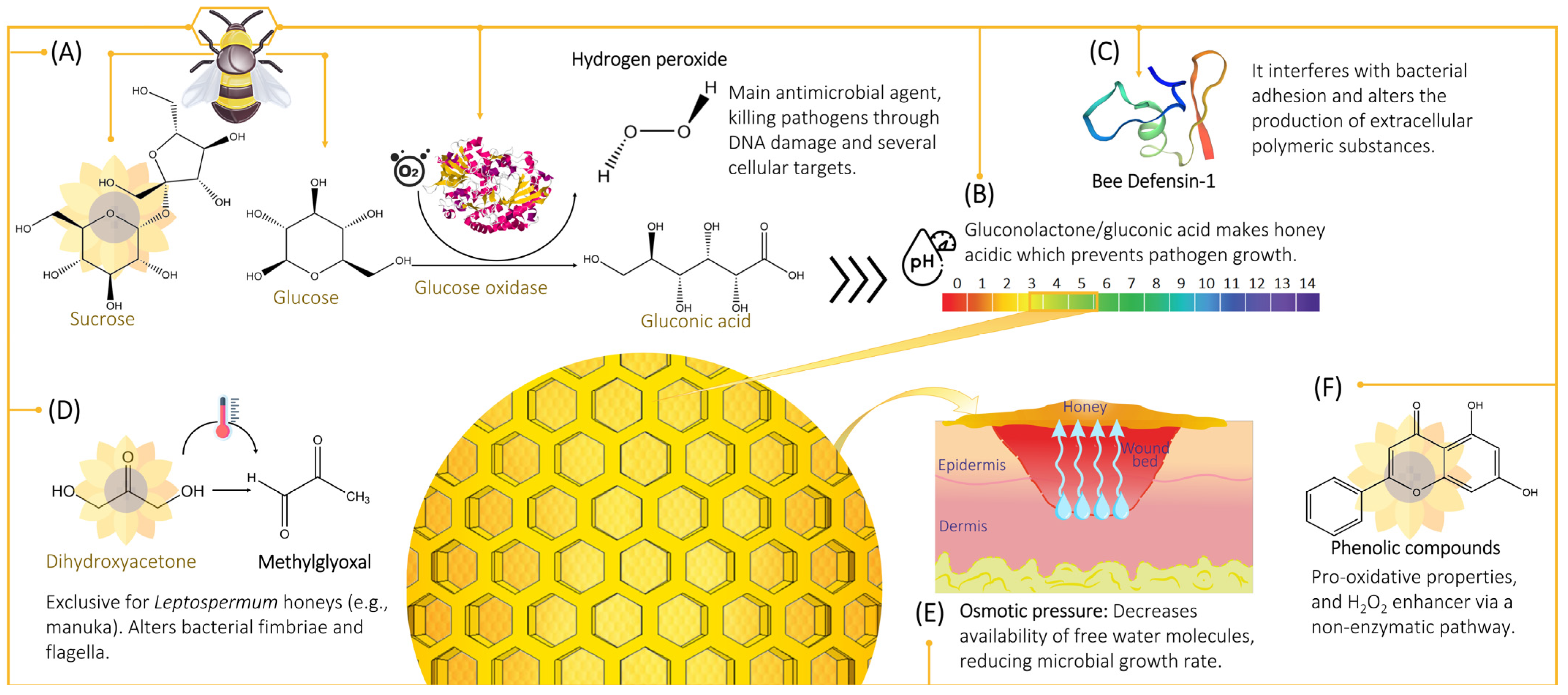
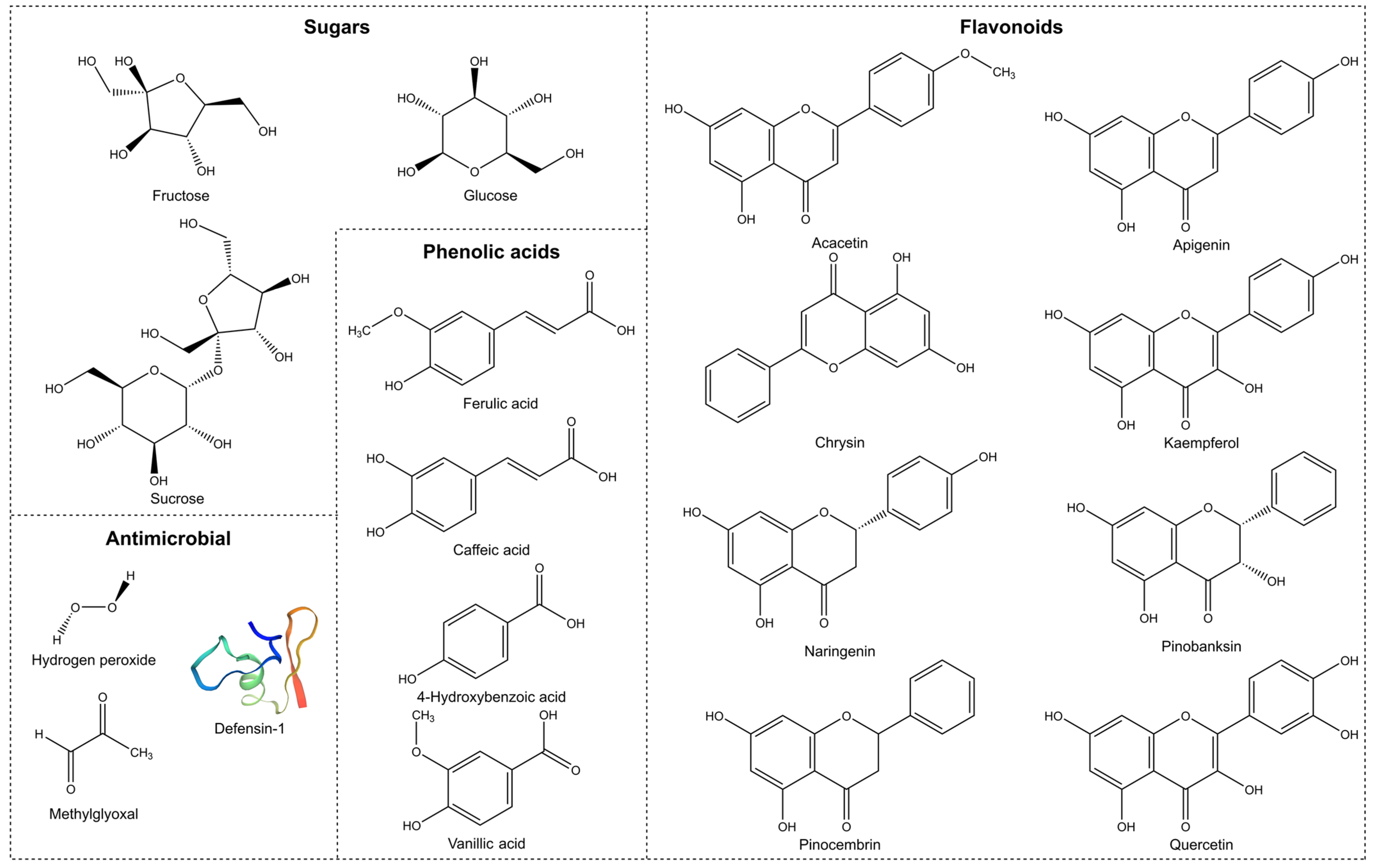
2. Antimicrobial Properties
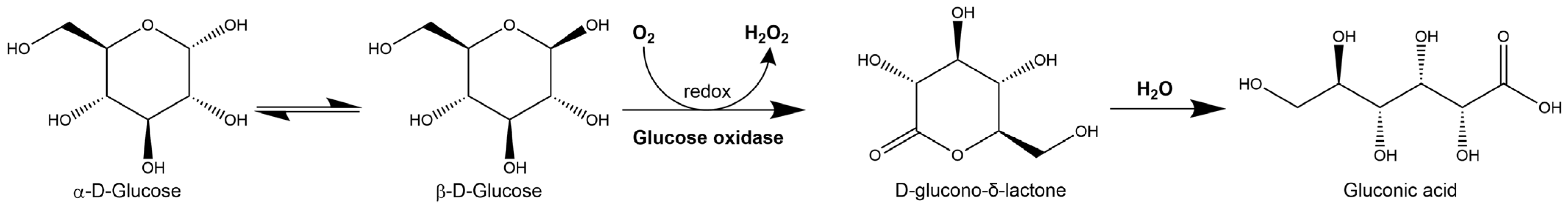
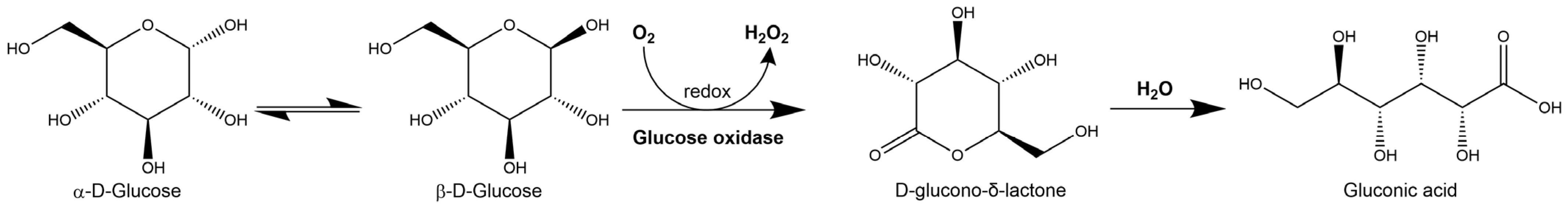
2.1. Hydrogen Peroxide
2.1.1. Hydrogen Peroxide Production
2.1.2. Cytotoxicity Mechanism of Hydrogen Peroxide
2.2. Non-Peroxide Antimicrobial Activity
2.2.1. Osmotic Effect
2.2.2. Acidity and pH
2.2.3. Phenolic Content
2.2.4. Defensin-1
2.2.5. Methylglyoxal
2.3. Antibacterial Activity
2.4. Anti-Fungal Activity
2.5. Antiviral Activity
2.6. Commercial Medical-Grade Honey
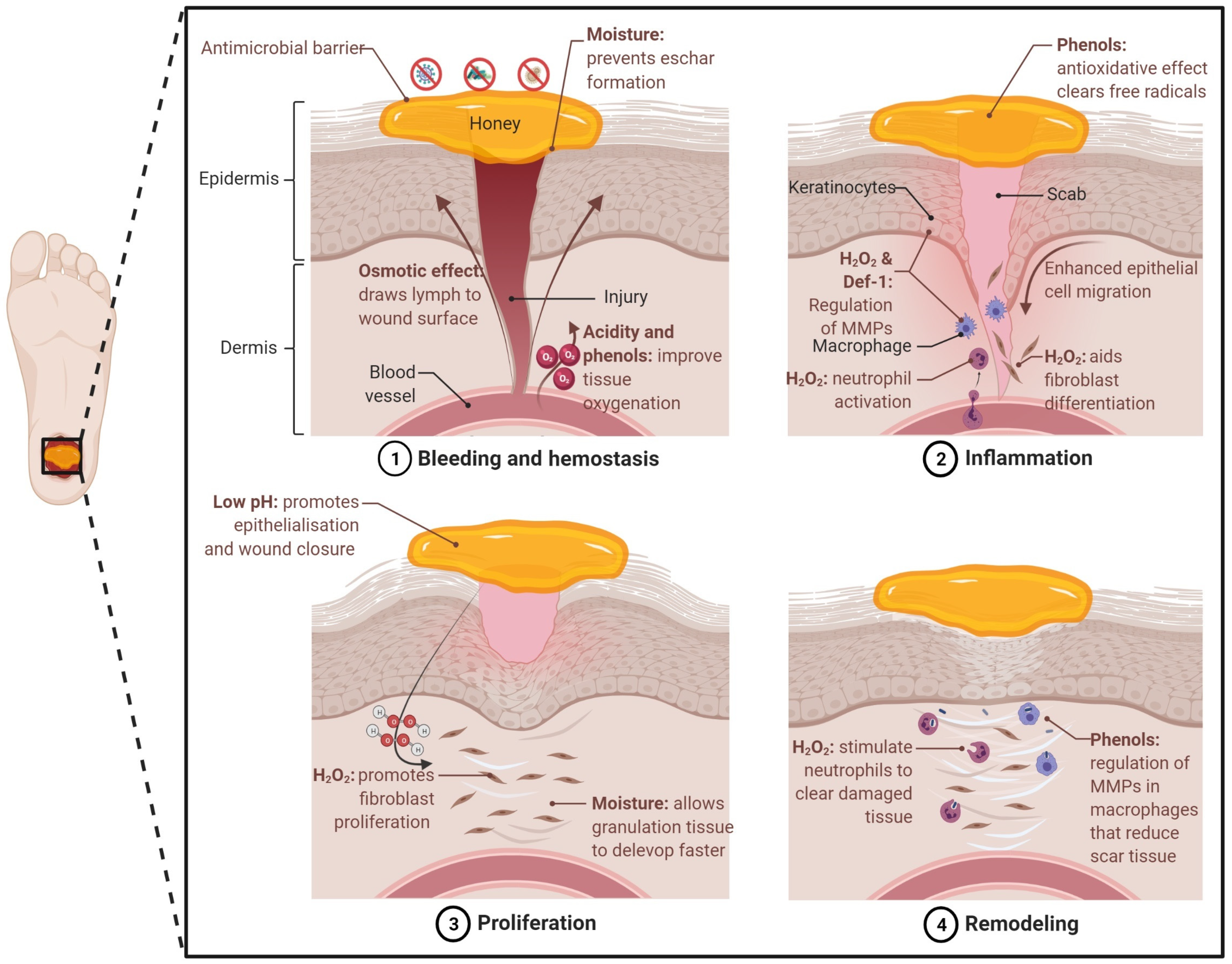
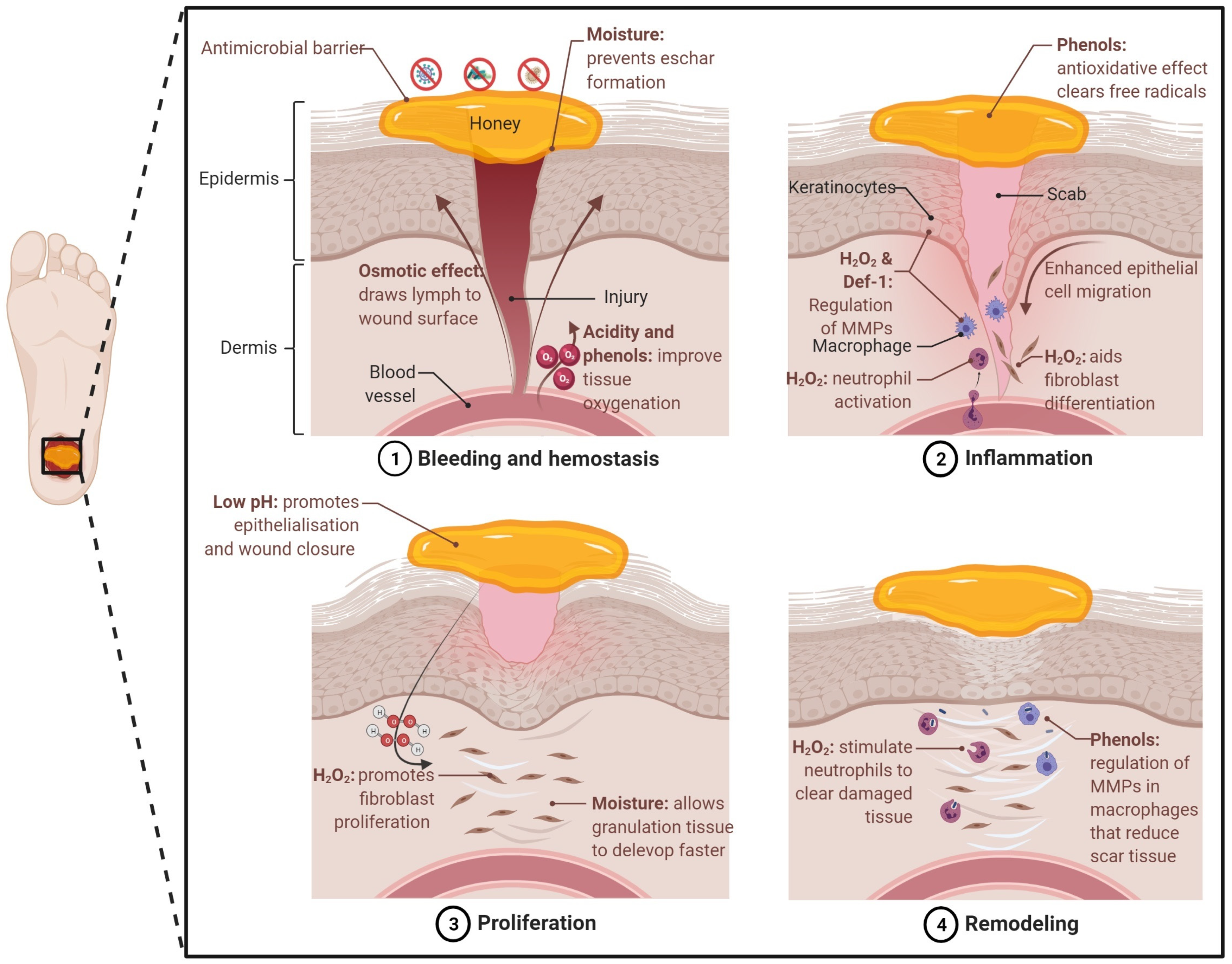
3. Honey as a Wound Healing and Tissue Regenerative Agent
Honey for Tissue Engineering Applications
4. Conclusions and Research Challenges
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Majno, G. The healing hand. Man and wound in the ancient world. Med. Hist. 1975, 20, 461. [Google Scholar]
- Roope, L.S.J.; Smith, R.D.; Pouwels, K.B.; Buchanan, J.; Abel, L.; Eibich, P.; Butler, C.C.; Tan, P.S.; Walker, A.S.; Robotham, J.V.; et al. The challenge of antimicrobial resistance: What economics can contribute. Science 2019, 364, eaau4679. [Google Scholar] [CrossRef]
- Nathan, C.; Cars, O. Antibiotic Resistance—Problems, Progress, and Prospects. N. Engl. J. Med. 2014, 371, 1761–1763. [Google Scholar] [CrossRef]
- Nathan, C. Resisting antimicrobial resistance. Nat. Rev. Microbiol. 2020, 18, 259–260. [Google Scholar] [CrossRef]
- Soroye, P.; Newbold, T.; Kerr, J. Climate change contributes to widespread declines among bumble bees across continents. Science 2020, 367, 685–688. [Google Scholar] [CrossRef]
- Woodcock, B.A.; Bullock, J.M.; Shore, R.F.; Heard, M.S.; Pereira, M.G.; Redhead, J.; Ridding, L.; Dean, H.; Sleep, D.; Henrys, P.; et al. Country-specific effects of neonicotinoid pesticides on honey bees and wild bees. Science 2017, 356, 1393. [Google Scholar] [CrossRef] [Green Version]
- Brettell, L.E.; Martin, S.J. Oldest Varroa tolerant honey bee population provides insight into the origins of the global decline of honey bees. Sci. Rep. 2017, 7, 45953. [Google Scholar] [CrossRef] [Green Version]
- Rangel, J.; Traver, B.; Stoner, M.; Hatter, A.; Trevelline, B.; Garza, C.; Shepherd, T.; Seeley, T.D.; Wenzel, J. Genetic diversity of wild and managed honey bees (Apis mellifera) in Southwestern Pennsylvania, and prevalence of the microsporidian gut pathogens Nosema ceranae and N. Apis. Apidologie 2020, 51, 802–814. [Google Scholar] [CrossRef]
- Villa, J.D.; Bustamante, D.M.; Dunkley, J.P.; Escobar, L.A. Changes in Honey Bee (Hymenoptera: Apidae) Colony Swarming and Survival Pre- and Postarrival of Varroa destructor (Mesostigmata: Varroidae) in Louisiana. Ann. Entomol. Soc. Am. 2008, 101, 867–871. [Google Scholar] [CrossRef] [Green Version]
- Raymann, K.; Moran, N.A. The role of the gut microbiome in health and disease of adult honey bee workers. Curr. Opin. Insect Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef]
- Alghamdi, B.A.; Alshumrani, E.S.; Saeed, M.S.B.; Rawas, G.M.; Alharthi, N.T.; Baeshen, M.N.; Helmi, N.M.; Alam, M.Z.; Suhail, M. Analysis of sugar composition and pesticides using HPLC and GC–MS techniques in honey samples collected from Saudi Arabian markets. Saudi J. Biol. Sci. 2020, 27, 3720–3726. [Google Scholar] [CrossRef]
- Amor, D.M. Composition, Properties and Uses of Honey: A Literature Survey; British Food Manufacturing Industries Research Association: Leatherhead, UK, 1978. [Google Scholar]
- White, J.W.; Riethof, M.L.; Subers, M.H.; Kushnir, I. Composition of American honeys. Tech. Bull. Dep. Agric. 1962, 1261, 157–158. [Google Scholar]
- Pasupuleti, V.R.; Sammugam, L.; Ramesh, N.; Gan, S.H. Honey, Propolis, and Royal Jelly: A Comprehensive Review of Their Biological Actions and Health Benefits. Oxid. Med. Cell. Longev. 2017, 2017, 21. [Google Scholar] [CrossRef]
- Van Ketel, B.A. Festnummer der berichten van den niederlandsche maatschappij. Bevord. Der Pharm. 1892, 67, 96. [Google Scholar]
- Molan, P.C. Why honey is effective as a medicine. 2. The scientific explanation of its effects. Bee World 2001, 82, 22–40. [Google Scholar] [CrossRef]
- Molan, P.C. The Antibacterial Activity of Honey. 1. The Nature of The Antibacterial Activity. Bee World 1992, 73, 5–28. [Google Scholar] [CrossRef]
- Cooke, J.; Dryden, M.; Patton, T.; Brennan, J.; Barrett, J. The antimicrobial activity of prototype modified honeys that generate reactive oxygen species (ROS) hydrogen peroxide. BMC Res. Notes 2015, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Dryden, M.; Lockyer, G.; Saeed, K.; Cooke, J. Engineered honey: In vitro antimicrobial activity of a novel topical wound care treatment. J. Glob. Antimicrob. Resist. 2014, 2, 168–172. [Google Scholar] [CrossRef] [Green Version]
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Leaper, D.; Georgopoulos, N.T. Reactive oxygen species (ROS) and wound healing: The functional role of ROS and emerging ROS-modulating technologies for augmentation of the healing process. Int. Wound J. 2017, 14, 89–96. [Google Scholar] [CrossRef]
- Mandal, M.D.; Mandal, S. Honey: Its medicinal property and antibacterial activity. Asian Pac. J. Trop. Biomed. 2011, 1, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Nolan, V.C.; Harrison, J.; Cox, J.A.G. Dissecting the Antimicrobial Composition of Honey. Antibiotics 2019, 8, 251. [Google Scholar] [CrossRef] [Green Version]
- Kuś, P.M.; Szweda, P.; Jerković, I.; Tuberoso, C.I.G. Activity of Polish unifloral honeys against pathogenic bacteria and its correlation with colour, phenolic content, antioxidant capacity and other parameters. Lett. Appl. Microbiol. 2016, 62, 269–276. [Google Scholar] [CrossRef]
- Wong, C.M.; Wong, K.H.; Chen, X.D. Glucose oxidase: Natural occurrence, function, properties and industrial applications. Appl. Microbiol. Biotechnol. 2008, 78, 927–938. [Google Scholar] [CrossRef]
- Cebrero, G.; Sanhueza, O.; Pezoa, M.; Báez, M.E.; Martínez, J.; Báez, M.; Fuentes, E. Relationship among the minor constituents, antibacterial activity and geographical origin of honey: A multifactor perspective. Food Chem. 2020, 315, 126296. [Google Scholar] [CrossRef]
- Bucekova, M.; Valachova, I.; Kohutova, L.; Prochazka, E.; Klaudiny, J.; Majtan, J. Honeybee glucose oxidase—Its expression in honeybee workers and comparative analyses of its content and H2O2-mediated antibacterial activity in natural honeys. Naturwissenschaften 2014, 101, 661–670. [Google Scholar] [CrossRef]
- Molan, P.C. Potential of Honey in the Treatment of Wounds and Burns. Am. J. Clin. Dermatol. 2001, 2, 13–19. [Google Scholar] [CrossRef]
- Vandamme, L.; Heyneman, A.; Hoeksema, H.; Verbelen, J.; Monstrey, S. Honey in modern wound care: A systematic review. Burns 2013, 39, 1514–1525. [Google Scholar] [CrossRef]
- Hixon, K.R.; Klein, R.C.; Eberlin, C.T.; Linder, H.R.; Ona, W.J.; Gonzalez, H.; Sell, S.A. A Critical Review and Perspective of Honey in Tissue Engineering and Clinical Wound Healing. Adv. Wound Care 2019, 8, 403–415. [Google Scholar] [CrossRef]
- Ding, Y.; Li, W.; Zhang, F.; Liu, Z.; Zanjanizadeh Ezazi, N.; Liu, D.; Santos, H.A. Electrospun Fibrous Architectures for Drug Delivery, Tissue Engineering and Cancer Therapy. Adv. Funct. Mater. 2019, 29, 1802852. [Google Scholar] [CrossRef]
- Li, J.; Mooney, D.J. Designing hydrogels for controlled drug delivery. Nat. Rev. Mater. 2016, 1, 16071. [Google Scholar] [CrossRef]
- Rossi, M.; Marrazzo, P. The Potential of Honeybee Products for Biomaterial Applications. Biomimetics 2021, 6, 6. [Google Scholar] [CrossRef]
- Minden-Birkenmaier, B.A.; Bowlin, G.L. Honey-Based Templates in Wound Healing and Tissue Engineering. Bioengineering 2018, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Love, N.R.; Chen, Y.Y.; Ishibashi, S.; Kritsiligkou, P.; Lea, R.; Koh, Y.; Gallop, J.L.; Dorey, K.; Amaya, E. Amputation-induced reactive oxygen species are required for successful Xenopus tadpole tail regeneration. Nat. Cell Biol. 2013, 15, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Defensin-1 Apis Mellifera (Honeybee) P17722. Available online: https://swissmodel.expasy.org/repository/uniprot/P17722 (accessed on 6 March 2021).
- Dold, H.; Du, D.H.; Dziao, S.T. Nachweis antibakterieller, hitze-und lichtempfindlicher Hemmungsstoffe (Inhibine) im Naturhonig (Blütenhonig). Z. Hyg. Infekt. 1937, 120, 155–167. [Google Scholar] [CrossRef]
- Prica, M. The Bactericidal Action of Honey. Z. Hyg. Infekt. 1938, 120, 437–443. [Google Scholar]
- Plachy, E. Studie über die bakterizide Wirkung des Naturhonigs (Blüten und Blatthonig) aus verschiedenen Höhenlagen sowie einige Untersuchungen über die Eigenschaft der antibakteriellen Hemmungstoffe (Inhibine) im Naturhonig. Zentbl. Bakt. ParasitKde Abt. II 1944, 106, 401–419. [Google Scholar]
- Dold, H.; Witzenhausen, R. Ein Verfahren zur Beurteilung der örtlichen inhibitorischen (keimvermehrungshemmenden) Wirkung von Honigsorten verschiedener Herkunft. Z. Hyg. Infekt. 1955, 141, 333–337. [Google Scholar] [CrossRef]
- Adcock, D. The Effect of Catalase on the Inhibine and Peroxide Values of Various Honeys. J. Apic. Res. 1962, 1, 38–40. [Google Scholar] [CrossRef]
- White, J.W.; Subers, M.H.; Schepartz, A.I. The identification of inhibine, the antibacterial factor in honey, as hydrogen peroxide and its origin in a honey glucose-oxidase system. Biochim. Biophys. Acta (BBA) Spec. Sect. Enzymol. Subj. 1963, 73, 57–70. [Google Scholar] [CrossRef]
- Molan, P.C. The Antibacterial Activity of Honey. 2. Variation in The Potency of the Antibacterial Activity. Bee World 1992, 73, 59–76. [Google Scholar] [CrossRef]
- Dustmann, J.H. Antibacterial effect of honey. Apiacta 1979, 14, 7–11. [Google Scholar]
- Scott, D.O.N. Oxidoreductases. In Enzymes in Food Processing, 2nd ed.; Reed, G., Ed.; Academic Press: Cambridge, MA, USA, 1975; pp. 219–254. [Google Scholar]
- Reed, G. Enzymes in Food Processing; Academic Press: Cambridge, MA, USA, 1966; pp. 176–181. [Google Scholar]
- Gauhe, A. Über ein glukoseoxydierendes Enzym in der Pharynxdrüse der Honigbiene. Z. Vgl. Physiol. 1940, 28, 211–253. [Google Scholar] [CrossRef]
- Schepartz, A.I.; Subers, M.H. The glucose oxidase of honey I. Purification and some general properties of the enzyme. Biochim. Biophys. Acta 1964, 85, 228–237. [Google Scholar] [CrossRef]
- Maurizio, A. From the Raw Material to the Finished Product: Honey. Bee World 1962, 43, 66–81. [Google Scholar] [CrossRef]
- White, J.W.; Subers, M.H. Studies on Honey Inhibine. 2. A Chemical Assay. J. Apic. Res. 1963, 2, 93–100. [Google Scholar] [CrossRef]
- Brudzynski, K.; Abubaker, K.; St-Martin, L.; Castle, A. Re-examining the role of hydrogen peroxide in bacteriostatic and bactericidal activities of honey. Front. Microbiol. 2011, 2, 9. [Google Scholar] [CrossRef] [Green Version]
- Roth, L.A.; Kwan, S.; Sporns, P. Use of a Disc-Assay System to Detect Oxytetracycline Residues in Honey. J. Food Prot. 1986, 49, 436–441. [Google Scholar] [CrossRef]
- Bang, L.M.; Buntting, C.; Molan, P. The effect of dilution on the rate of hydrogen peroxide production in honey and its implications for wound healing. J. Altern. Complement Med. 2003, 9, 267–273. [Google Scholar] [CrossRef] [Green Version]
- Dustmann, J.H. Über die Katalaseaktivität in Bienenhonig aus der Tracht der Heidekrautgewächse (Ericaceae). Z. Lebensm.-Unters. Forsch. 1971, 145, 294–295. [Google Scholar] [CrossRef]
- Weston, R.J. The contribution of catalase and other natural products to the antibacterial activity of honey: A review. Food Chem. 2000, 71, 235–239. [Google Scholar] [CrossRef]
- Turner, F. Hydrogen Peroxide and Other Oxidant Disinfectants; Lea & Febiger: Philadelphia, PA, USA, 1983; pp. 240–280. [Google Scholar]
- Imlay, J.A.; Linn, S. Bimodal pattern of killing of DNA-repair-defective or anoxically grown Escherichia coli by hydrogen peroxide. J. Bacteriol. 1986, 166, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Imlay, J.; Linn, S. DNA damage and oxygen radical toxicity. Science 1988, 240, 1302–1309. [Google Scholar] [CrossRef] [Green Version]
- Haber, F.; Weiss, J. The Catalytic Decomposition of Hydrogen Peroxide by Iron Salts. Proc. R. Soc. Lond. Ser. A 1934, 147, 332–351. [Google Scholar] [CrossRef]
- Liochev, S.I. The mechanism of “Fenton-like” reactions and their importance for biological systems. A biologist’s view. Met. Ions Biol. Syst. 1999, 36, 1–39. [Google Scholar]
- Linley, E.; Denyer, S.P.; McDonnell, G.; Simons, C.; Maillard, J.-Y. Use of hydrogen peroxide as a biocide: New consideration of its mechanisms of biocidal action. J. Antimicrob. Chemother. 2012, 67, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.C. Antimicrobial reactive oxygen and nitrogen species: Concepts and controversies. Nat. Rev. Microbiol. 2004, 2, 820–832. [Google Scholar] [CrossRef]
- Tamarit, J.; Cabiscol, E.; Ros, J. Identification of the Major Oxidatively Damaged Proteins in Escherichia coli Cells Exposed to Oxidative Stress. J. Biol. Chem. 1998, 273, 3027–3032. [Google Scholar] [CrossRef] [Green Version]
- Albaridi, N.A. Antibacterial Potency of Honey. Int. J. Microbiol. 2019, 2019, 2464507. [Google Scholar] [CrossRef]
- Schroeder, A.; Horn, H.; Pieper, H.J. The correlation between moisture content and water activity (a (w)) in honey. Dtsch. Lebensm. Rundsch. 2005, 101, 139–142. [Google Scholar]
- Chen, C.C. Relationship between Water Activity and Moisture Content in Floral Honey. Foods 2019, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Chirife, J.; Zamora, M.C.; Motto, A. The correlation between water activity and % moisture in honey: Fundamental aspects and application to Argentine honeys. J. Food Eng. 2006, 72, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Gleiter, R.A.; Horn, H.; Isengard, H.D. Influence of type and state of crystallisation on the water activity of honey. Food Chem. 2006, 96, 441–445. [Google Scholar] [CrossRef]
- Medved’ova, A.; Havlikova, A.; Lehotova, V.; Valik, L. Staphylococcus aureus 2064 growth as affected by temperature and reduced water activity. Ital. J. Food Saf. 2019, 8, 188–193. [Google Scholar] [CrossRef]
- Yatsunami, K.; Echigo, T. Antibacterial activity of honey and royal jelly. Honeybee Sci. 1984, 5, 125–130. [Google Scholar]
- Bogdanov, S. Antibacterial substances in honey. Artik. Swiss Bee Res. Cent. Switz. 1997, 17, 74–76. [Google Scholar]
- Wahdan, H.A.L. Causes of the antimicrobial activity of honey. Infection 1998, 26, 30–35. [Google Scholar] [CrossRef]
- Chang, X.; Wang, J.H.; Yang, S.H.; Chen, S.; Song, Y.J. Antioxidative, antibrowning and antibacterial activities of sixteen floral honeys. Food Funct. 2011, 2, 541–546. [Google Scholar] [CrossRef]
- Viuda-Martos, M.; Ruiz-Navajas, Y.; Fernandez-Lopez, J.; Perez-Alvarez, J.A. Functional Properties of Honey, Propolis, and Royal Jelly. J. Food Sci. 2008, 73, R117–R124. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Estevinho, L.; Pereira, A.P.; Moreira, L.; Dias, L.G.; Pereira, E. Antioxidant and antimicrobial effects of phenolic compounds extracts of Northeast Portugal honey. Food Chem. Toxicol. 2008, 46, 3774–3779. [Google Scholar] [CrossRef]
- Andrade, P.; Ferreres, F.; Amaral, M.T. Analysis of Honey Phenolic Acids by HPLC, Its Application to Honey Botanical Characterization. J. Liq. Chromatogr. Relat. Technol. 1997, 20, 2281–2288. [Google Scholar] [CrossRef]
- Metzner, J.; Bekemeier, H.; Paintz, M.; Schneidewind, E. On the antimicrobial activity of propolis and propolis constituents (author’s transl). Die Pharm. 1979, 34, 97–102. [Google Scholar]
- Ferreres, F.; Ortiz, A.; Silva, C.; Garcia-Viguera, C.; Tomás-Barberán, F.A.; Tomás-Lorente, F. Flavonoids of “La Alcarria” honey A study of their botanical origin. Z. Lebensm. Unters. Forsch. 1992, 194, 139–143. [Google Scholar] [CrossRef]
- McLoone, P.; Warnock, M.; Fyfe, L. Honey: A realistic antimicrobial for disorders of the skin. J. Microbiol. Immunol. Infect. 2016, 49, 161–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheller, S.; Szaflarski, J.; Tustanowski, J.; Nolewajka, E.; Stojko, A. Biological properties and clinical application of propolis. I. Some physico-chemical properties of propolis. Arzneim.-Forsch. 1977, 27, 889–890. [Google Scholar]
- Kwakman, P.H.S.; Zaat, S.A.J. Antibacterial components of honey. IUBMB Life 2012, 64, 48–55. [Google Scholar] [CrossRef]
- Al-Waili, N.; Al-Ghamdi, A.; Ansari, M.J.; Al-Attal, Y.; Salom, K. Synergistic effects of honey and propolis toward drug multi-resistant Staphylococcus aureus, Escherichia coli and Candida albicans isolates in single and polymicrobial cultures. Int. J. Med. Sci. 2012, 9, 793–800. [Google Scholar] [CrossRef] [Green Version]
- Przybyłek, I.; Karpiński, T.M. Antibacterial Properties of Propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef] [Green Version]
- Bucekova, M.; Buriova, M.; Pekarik, L.; Majtan, V.; Majtan, J. Phytochemicals-mediated production of hydrogen peroxide is crucial for high antibacterial activity of honeydew honey. Sci. Rep. 2018, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Bucekova, M.; Jardekova, L.; Juricova, V.; Bugarova, V.; Di Marco, G.; Gismondi, A.; Leonardi, D.; Farkasovska, J.; Godocikova, J.; Laho, M.; et al. Antibacterial Activity of Different Blossom Honeys: New Findings. Molecules 2019, 24, 1573. [Google Scholar] [CrossRef] [Green Version]
- Bucekova, M.; Sojka, M.; Valachova, I.; Martinotti, S.; Ranzato, E.; Szep, Z.; Majtan, V.; Klaudiny, J.; Majtan, J. Bee-derived antibacterial peptide, defensin-1, promotes wound re-epithelialisation in vitro and in vivo. Sci. Rep. 2017, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Kwakman, P.H.S.; Velde, A.A.T.; de Boer, L.; Speijer, D.; Christina Vandenbroucke-Grauls, M.J.; Zaat, S.A.J. How honey kills bacteria. FASEB J. 2010, 24, 2576–2582. [Google Scholar] [CrossRef] [Green Version]
- Klaudiny, J.; Albert, T.; Bachanova, K.; Kopernick, J.; Simuth, J. Two structurally different defensin genes, one of them encoding a novel defensin isoform, are expressed in honeybee Apis mellifera. Insect Biochem. Mol. Biol. 2005, 35, 11–22. [Google Scholar] [CrossRef]
- Sojka, M.; Valachova, I.; Bucekova, M.; Majtan, J. Antibiofilm efficacy of honey and bee-derived defensin-1 on multispecies wound biofilm. J. Med. Microbiol. 2016, 65, 337–344. [Google Scholar] [CrossRef]
- Čeřovský, V.; Bém, R. Lucifensins, the Insect Defensins of Biomedical Importance: The Story behind Maggot Therapy. Pharmaceuticals 2014, 7, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Valachova, I.; Bucekova, M.; Majtan, J. Quantification of Bee-Derived Peptide Defensin-1 in Honey by Competitive Enzyme-Linked Immunosorbent Assay, a New Approach in Honey Quality Control. Czech. J. Food Sci. 2016, 34, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.L.; Molan, P.C.; Reid, G.M. A Survey of the Antibacterial Activity of Some New Zealand Honeys. J. Pharm. Pharmacol. 1991, 43, 817–822. [Google Scholar] [CrossRef]
- Weigel, K.U.; Opitz, T.; Henle, T. Studies on the occurrence and formation of 1,2-dicarbonyls in honey. Eur. Food Res. Technol. 2004, 218, 151. [Google Scholar] [CrossRef]
- Mavric, E.; Wittmann, S.; Barth, G.; Henle, T. Identification and quantification of methylglyoxal as the dominant antibacterial constituent of Manuka (Leptospermum scoparium) honeys from New Zealand. Mol. Nutr. Food Res. 2008, 52, 483–489. [Google Scholar] [CrossRef]
- Grainger, M.N.C.; Manley-Harris, M.; Lane, J.R.; Field, R.J. Kinetics of conversion of dihydroxyacetone to methylglyoxal in New Zealand mānuka honey: Part I—Honey systems. Food Chem. 2016, 202, 484–491. [Google Scholar] [CrossRef]
- Shapla, U.M.; Solayman, M.; Alam, N.; Khalil, M.I.; Gan, S.H. 5-Hydroxymethylfurfural (HMF) levels in honey and other food products: Effects on bees and human health. Chem. Cent. J. 2018, 12, 35. [Google Scholar] [CrossRef]
- Suortti, T.; Malkki, Y. Antimicrobial Activities of Heated Glucose and Fructose Solutions and Their Elucidation by High-Performance Liquid-Chromatography. Food Chem. 1984, 15, 165–173. [Google Scholar] [CrossRef]
- Parmar, M.S.; Wexler, P. Methylglyoxal. In Encyclopedia of Toxicology, 3rd ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 302–305. [Google Scholar]
- Majtan, J. Methylglyoxal—A Potential Risk Factor of Manuka Honey in Healing of Diabetic Ulcers. Evid. Based Complement. Altern. Med. 2011, 2011, 295494. [Google Scholar] [CrossRef] [Green Version]
- Cooper, R.; Jenkins, L. A Comparison Between Medical Grade Honey and Table Honeys in Relation to Antimicrobial Efficacy. Wounds 2009, 21, 29–36. [Google Scholar]
- Kwakman, P.H.S.; Velde, A.A.T.; de Boer, L.; Vandenbroucke-Grauls, C.; Zaat, S.A.J. Two Major Medicinal Honeys Have Different Mechanisms of Bactericidal Activity. PLoS ONE 2011, 6, e17709. [Google Scholar] [CrossRef] [Green Version]
- Rabie, E.; Serem, J.C.; Oberholzer, H.M.; Gaspar, A.R.M.; Bester, M.J. How methylglyoxal kills bacteria: An ultrastructural study. Ultrastruct. Pathol. 2016, 40, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Schalkwijk, C.G.; Stehouwer, C.D.A. Methylglyoxal, a Highly Reactive Dicarbonyl Compound, in Diabetes, Its Vascular Complications, and Other Age-Related Diseases. Physiol. Rev. 2019, 100, 407–461. [Google Scholar] [CrossRef]
- Hanssen, N.M.J.; Scheijen, J.L.J.M.; Jorsal, A.; Parving, H.-H.; Tarnow, L.; Rossing, P.; Stehouwer, C.D.A.; Schalkwijk, C.G. Higher Plasma Methylglyoxal Levels Are Associated with Incident Cardiovascular Disease in Individuals with Type 1 Diabetes: A 12-Year Follow-up Study. Diabetes 2017, 66, 2278. [Google Scholar] [CrossRef] [Green Version]
- Berlanga, J.; Cibrian, D.; GuillÉN, I.; Freyre, F.; Alba, J.S.; Lopez-Saura, P.; Merino, N.; Aldama, A.; Quintela, A.M.; Triana, M.E.; et al. Methylglyoxal administration induces diabetes-like microvascular changes and perturbs the healing process of cutaneous wounds. Clin. Sci. 2005, 109, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Shen, M.; Kuang, H.; Liu, X.; Zhang, C.; Tian, Y.; Miao, X.; Xu, X. The botanical sources, entomological proteome and antibiotic properties of wild honey. Innov. Food Sci. Emerg. Technol. 2021, 67, 102589. [Google Scholar] [CrossRef]
- Cooper, R.A.; Molan, P.C.; Harding, K.G. The sensitivity to honey of Gram-positive cocci of clinical significance isolated from wounds. J. Appl. Microbiol. 2002, 93, 857–863. [Google Scholar] [CrossRef]
- Anthimidou, E.; Mossialos, D. Antibacterial Activity of Greek and Cypriot Honeys Against Staphylococcus aureus and Pseudomonas aeruginosa in Comparison to Manuka Honey. J. Med. Food 2013, 16, 42–47. [Google Scholar] [CrossRef]
- Grecka, K.; Kus, P.M.; Worobo, R.W.; Szweda, P. Study of the Anti-Staphylococcal Potential of Honeys Produced in Northern Poland. Molecules 2018, 23, 260. [Google Scholar] [CrossRef] [Green Version]
- Grego, E.; Robino, P.; Tramuta, C.; Giusto, G.; Boi, M.; Colombo, R.; Serra, G.; Chiado-Cutin, S.; Gandini, M.; Nebbia, P. Evaluation of antimicrobial activity of Italian honey for wound healing application in veterinary medicine. Schweiz. Arch. Tierheilkd. 2016, 158, 521–527. [Google Scholar] [CrossRef]
- Anand, S.; Deighton, M.; Livanos, G.; Morrison, P.D.; Pang, E.C.K.; Mantri, N. Antimicrobial Activity of Agastache Honey and Characterization of Its Bioactive Compounds in Comparison with Important Commercial Honeys. Front. Microbiol. 2019, 10, 263. [Google Scholar] [CrossRef]
- Sherlock, O.; Dolan, A.; Athman, R.; Power, A.; Gethin, G.; Cowman, S.; Humphreys, H. Comparison of the antimicrobial activity of Ulmo honey from Chile and Manuka honey against methicillin-resistant Staphylococcus aureus, Escherichia coli and Pseudomonas aeruginosa. BMC Complement. Altern. Med. 2010, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.T.; Rahman, R.A.; Gan, S.H.; Halim, A.S.; Hassan, S.A.; Sulaiman, S.A.; Kirnpal-Kaur, B.S. The antibacterial properties of Malaysian tualang honey against wound and enteric microorganisms in comparison to manuka honey. BMC Complement. Altern. Med. 2009, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Ng, W.-J.; Sit, N.-W.; Ooi, P.A.; Ee, K.-Y.; Lim, T.-M. The Antibacterial Potential of Honeydew Honey Produced by Stingless Bee (Heterotrigona itama) against Antibiotic Resistant Bacteria. Antibiotics 2020, 9, 871. [Google Scholar] [CrossRef]
- Jenkins, R.; Cooper, R. Improving Antibiotic Activity against Wound Pathogens with Manuka Honey In Vitro. PLoS ONE 2012, 7, e45600. [Google Scholar] [CrossRef] [Green Version]
- Müller, P.; Alber, D.G.; Turnbull, L.; Schlothauer, R.C.; Carter, D.A.; Whitchurch, C.B.; Harry, E.J. Synergism between Medihoney and Rifampicin against Methicillin-Resistant Staphylococcus aureus (MRSA). PLoS ONE 2013, 8, e57679. [Google Scholar] [CrossRef]
- Jenkins, R.E.; Cooper, R. Synergy between oxacillin and manuka honey sensitizes methicillin-resistant Staphylococcus aureus to oxacillin. J. Antimicrob. Chemother. 2012, 67, 1405–1407. [Google Scholar] [CrossRef] [Green Version]
- Campeau, M.E.M.; Patel, R. Antibiofilm Activity of Manuka Honey in Combination with Antibiotics. Int. J. Bacteriol. 2014, 2014, 795281. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.; Ribeiro, H.G.; Silva, A.C.; Silva, M.D.; Sousa, J.C.; Rodrigues, C.F.; Melo, L.D.R.; Henriques, A.F.; Sillankorva, S. Synergistic Antimicrobial Interaction between Honey and Phage against Escherichia coli Biofilms. Front. Microbiol. 2017, 8, 2407. [Google Scholar] [CrossRef]
- Scorzoni, L.; Silva, A.; Marcos, C.M.; Assato, P.A.; de Melo, W.; de Oliveira, H.C.; Costa-Orlandi, C.B.; Mendes-Giannini, M.J.S.; Fusco-Almeida, A.M. Antifungal Therapy: New Advances in the Understanding and Treatment of Mycosis. Front. Microbiol. 2017, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole Antifungal Resistance in Candida albicans and Emerging Non-albicans Candida Species. Front. Microbiol. 2017, 7, 2173. [Google Scholar] [CrossRef] [Green Version]
- Jeffery-Smith, A.; Taori, S.K.; Schelenz, S.; Jeffery, K.; Johnson, E.M.; Borman, A.; Manuel, R.; Brown, C.S. Candida auris: A Review of the Literature. Clin. Microbiol. Rev. 2018, 31, e00029-17. [Google Scholar] [CrossRef] [Green Version]
- Groll, A.H.; Grist, L.M. Current challenges in the diagnosis and management of invasive fungal infections: Report from the 15th International Symposium on Infections in the Immunocompromised Host: Thessaloniki, Greece, 22–25 June 2008. Int. J. Antimicrob. Agents 2009, 33, 101–104. [Google Scholar] [CrossRef]
- Perlin, D.S.; Shor, E.; Zhao, Y. Update on Antifungal Drug Resistance. Curr. Clin. Microbiol. 2015, 2, 84–95. [Google Scholar] [CrossRef]
- Ansari, M.J.; Al-Ghamdi, A.; Usmani, S.; Al-Waili, N.S.; Sharma, D.; Nuru, A.; Al-Attal, Y. Effect of Jujube Honey on Candida albicans Growth and Biofilm Formation. Arch. Med. Res. 2013, 44, 352–360. [Google Scholar] [CrossRef]
- Haydak, M.H.; Crane, E.; Duisberg, H.; Cochnauer, T.A.; Morse, R.A.; White, J.W.; Wix, P. Biological properties of honey. In Honey: A Comprehensive Survey; William Heinemann: London, UK, 1975; pp. 258–266. [Google Scholar]
- Irish, J.; Carter, D.A.; Shokohi, T.; Blair, S.E. Honey has an antifungal effect against Candida species. Med. Mycol. 2006, 44, 289–291. [Google Scholar] [CrossRef] [Green Version]
- Katiraee, F.; Mahmodi, R.; Mardani, K.; Babaei, E. Antifungal Activity of Iranian Honeybees Against Candida, Aspergillus Species And Trichophyton Rubrum. J. Food Process Preserv. 2014, 38, 2078–2082. [Google Scholar] [CrossRef]
- Anand, S.; Deighton, M.; Livanos, G.; Pang, E.C.K.; Mantri, N. Agastache honey has superior antifungal activity in comparison with important commercial honeys. Sci. Rep. 2019, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Arai, M.; Tomoda, H.; Okuda, T.; Wang, H.Y.; Tabata, N.; Masuma, R.; Yamaguchi, Y.; Omura, S. Funicone-related compounds, potentiators of antifungal miconazole activity, produced by Talaromyces flavus FKI-0076. J. Antibiot. 2002, 55, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Kubo, A.; Lunde, C.S.; Kubo, I. Antimicrobial Activity of The Olive Oil Flavor Compounds. J. Agric. Food Chem. 1995, 43, 1629–1633. [Google Scholar] [CrossRef]
- Israili, Z.H. Antimicrobial Properties of Honey. Am. J. Ther. 2014, 21, 304–323. [Google Scholar] [CrossRef] [PubMed]
- Zeina, B.; Othman, O.; Al-Assad, S. Effect of Honey versus Thyme on Rubella Virus Survival in Vitro. J. Altern. Complement. Med. 1996, 2, 345–348. [Google Scholar] [CrossRef]
- NICE. Cough (Acute): Antimicrobial Prescribing. Available online: https://www.nice.org.uk/guidance/ng120/resources/cough-acute-antimicrobial-prescribing-pdf-66141652166341 (accessed on 26 August 2020).
- Al-Waili, N.S. Topical honey application vs. acyclovir for the treatment of recurrent herpes simplex lesions. Med. Sci. Monit. 2004, 10, MT94–MT98. [Google Scholar]
- Semprini, A.; Singer, J.; Braithwaite, I.; Shortt, N.; Thayabaran, D.; McConnell, M.; Weatherall, M.; Beasley, R. Kanuka honey versus aciclovir for the topical treatment of herpes simplex labialis: A randomised controlled trial. BMJ Open 2019, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Miguel, M.G.; Antunes, M.D.; Faleiro, M.L. Honey as a Complementary Medicine. Integr. Med. Insights 2017, 12, 15. [Google Scholar] [CrossRef] [Green Version]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Critchfield, J.W.; Butera, S.T.; Folks, T.M. Inhibition of HIV activation in latently infected cells by flavonoid compounds. Aids Res. Hum. Retrovir. 1996, 12, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Debiaggi, M.; Tateo, F.; Pagani, L.; Luini, M.; Romero, E. Effects of Propolis Flavonoids on Virus Infectivity and Replication. Microbiologica 1990, 13, 207–213. [Google Scholar] [PubMed]
- Schnitzler, P.; Neuner, A.; Nolkemper, S.; Zundel, C.; Nowack, H.; Sensch, K.H.; Reichling, J. Antiviral Activity and Mode of Action of Propolis Extracts and Selected Compounds. Phytother. Res. 2010, 24, S20–S28. [Google Scholar] [CrossRef] [PubMed]
- Al-Waili, N.; Salom, K.; Al-Ghamdi, A.; Ansari, M.J. Antibiotic, pesticide, and microbial contaminants of honey: Human health hazards. Sci. World J. 2012, 2012, 930849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remize, F.; Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Chapter 4—Spore-Forming Bacteria. In The Microbiological Quality of Food; Woodhead Publishing: Thorston, UK, 2017; pp. 99–120. [Google Scholar]
- Hermanns, R.; Mateescu, C.; Thrasyvoulou, A.; Tananaki, C.; Wagener, F.A.D.T.G.; Cremers, N.A.J. Defining the standards for medical grade honey. J. Apic. Res. 2020, 59, 125–135. [Google Scholar] [CrossRef]
- Horniackova, M.; Bucekova, M.; Valachova, I.; Majtan, J. Effect of gamma radiation on the antibacterial and antibiofilm activity of honeydew honey. Eur. Food Res. Technol. 2017, 243, 81–88. [Google Scholar] [CrossRef]
- Oxford Health NHS Foundation Trust. Medical Honey Simplified. Available online: https://www.oxfordhealth.nhs.uk/wp-content/uploads/2015/08/Medical_Honey_Simplified_-_Patients-leaflet.pdf (accessed on 2 June 2020).
- Bong, J.; Prijic, G.; Braggins, T.J.; Schlothauer, R.C.; Stephens, J.M.; Loomes, K.M. Leptosperin is a distinct and detectable fluorophore in Leptospermum honeys. Food Chem. 2017, 214, 102–109. [Google Scholar] [CrossRef]
- UMFHA. Scientific Breakthrough Identifies Genuine Mānuka Honey. Available online: https://www.umf.org.nz/wp-content/myimages/2017/02/Press-Release-UK-News-FINAL.pdf (accessed on 27 May 2020).
- UMFHA. About the UMF Honey Association. Available online: https://www.umf.org.nz/what-we-do/ (accessed on 27 May 2020).
- Manuka Health. What Is MGO. Available online: https://www.manukahealth.co.nz/en-nz/manuka-honey/what-is-mgo/ (accessed on 27 May 2020).
- Comvita. MGO & UMF Explained. Available online: https://www.comvita.co.uk/store/new_comvita/pages/ingredients-uk/learn-about/mgo-and-umf.jsp (accessed on 27 May 2020).
- Girma, A.; Seo, W.; She, R.C. Antibacterial activity of varying UMF-graded Manuka honeys. PLoS ONE 2019, 14, 9. [Google Scholar] [CrossRef] [Green Version]
- NHS Supply Chain. About Us. Available online: https://www.supplychain.nhs.uk/about-us/ (accessed on 2 June 2020).
- Advancis Medical. Activon® Manuka Honey. Available online: https://uk.advancismedical.com/products/activon-manuka-honey (accessed on 17 July 2020).
- Cooper, R.; Jenkings, L.; Rowlands, R. Inhibition of biofilms through the use of manuka honey. Wounds UK 2011, 7, 24–32. [Google Scholar]
- Rafter, L.; Reynolds, T.; Collier, M.; Rafter, M.; West, M. A clinical evaluation of Algivon® Plus manuka honey dressings for chronic wounds. Wounds UK 2017, 13, 132–140. [Google Scholar]
- Edwards, J. A prospective evaluation of the use of honey dressings to manage burn wounds. Wounds UK 2013, 9, 102–106. [Google Scholar]
- Cryer, S. The use of manuka honey to facilitate the autoamputation of fingertip necrosis. Wounds UK 2016, 12, 66–70. [Google Scholar]
- Welland Medical. Aurum with Manuka Honey. Available online: https://wellandmedical.com/brand_type/aurum/ (accessed on 25 August 2020).
- Martin-Skurr, K. Case Study: Pyoderma Gangrenosum and the Effects of Manuka Honey. Available online: https://wellandmedical.com/wp-content/uploads/2019/02/Case-study_NZ-Pyoderma-Gangrenosum-Manuka-Honey.pdf (accessed on 15 September 2020).
- L-Mesitran. Products. Available online: https://l-mesitran.com/eu/en/products (accessed on 16 July 2020).
- Boggust, A. Evaluation of L-Mesitran® dressings in the treatment of minor burns and scalds at a paediatric emergency department. Wounds UK 2013, 9, 114–117. [Google Scholar]
- Stephen-Haynes, J.; Callaghan, R. Properties of honey: Its mode of action and clinical outcomes. Wounds UK 2011, 7, 50–57. [Google Scholar]
- International, M. ManukaDress—Sterile Honey Wound Dressing Range. Available online: https://medicareplus.co.uk/product/manukadress/ (accessed on 1 November 2020).
- Sack, A.; Pleat, J.; Cobley, T.; Mackie, I.; Estala, C.; Stephen-Haynes, J.; Callaghan, R.; Taylor, S.; Tucker, J.; Elston, H. Case studies series: ManukaDress IG and ManukaDress IG Max for the management of burns. Wounds UK 2018, 14, 110–117. [Google Scholar]
- Callaghan, R. Treating difficult-to-debride wounds using a manuka honey dressing: A case study evaluation. Wounds UK 2014, 10, 104–109. [Google Scholar]
- Bradbury, S.; Callaghan, R.; Ivins, N. ManukaDress Made Easy. Wounds UK 2014, 10, 1–6. [Google Scholar]
- Derma Science Europe. Antibacterial Dressing Medihoney®: Antibacterial Medical Honey. Available online: https://media.supplychain.nhs.uk/media/documents/ELY302/Marketing/48251_ELY302_3.pdf.pdf (accessed on 17 July 2020).
- Robson, V.; Dodd, S.; Thomas, S. Standardized antibacterial honey (Medihoney (TM)) with standard therapy in wound care: Randomized clinical trial. J. Adv. Nurs. 2009, 65, 565–575. [Google Scholar] [CrossRef]
- Johnson, D.W.; van Eps, C.; Mudge, D.W.; Wiggins, K.J.; Armstrong, K.; Hawley, C.M.; Campbell, S.B.; Isbel, N.M.; Nimmo, G.R.; Gibbs, H. Randomized, controlled trial of topical exit-site application of honey (Medihoney) versus Mupirocin for the prevention of catheter-associated infections in hemodialysis patients. J. Am. Soc. Nephrol. 2005, 16, 1456–1462. [Google Scholar] [CrossRef] [Green Version]
- Kestrel Health Information. MEDIHONEY® Calcium Alginate Dressing. Available online: https://www.woundsource.com/product/medihoney-calcium-alginate-dressing (accessed on 24 March 2020).
- Jull, A.; Walker, N.; Parag, V.; Molan, P.; Rodgers, A. Randomized clinical trial of honey-impregnated dressings for venous leg ulcers. Br. J. Surg. 2008, 95, 175–182. [Google Scholar] [CrossRef]
- Derma Sciences Europe. Antibacterial Dressing Medihoney®: Barrier Cream. Available online: https://media.supplychain.nhs.uk/media/documents/ELY289/Marketing/48248_ELY289.pdf.pdf (accessed on 13 July 2020).
- Nijhuis, W.A.; Houwing, R.H.; Van der Zwet, W.C.; Jansman, F.G.A. A randomised trial of honey barrier cream versus zinc oxide ointment. Br. J. Nurs. 2012, 21, S10–S13. [Google Scholar] [CrossRef]
- Comvita. Antibacterial Wound Gel™ 25g. Available online: https://www.comvita.co.uk/product/antibacterial-wound-gel%E2%84%A2-25g/6011 (accessed on 15 June 2020).
- Robson, V.; Yorke, J.; Sen, R.A.; Lowe, D.; Rogers, S.N. Randomised controlled feasibility trial on the use of medical grade honey following microvascular free tissue transfer to reduce the incidence of wound infection. Br. J. Oral Maxillofac. Surg. 2012, 50, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Matoke Holdings. Surgihoney RO. Available online: https://www.surgihoneyro.com/ (accessed on 13 April 2020).
- Dryden, M.; Goddard, C.; Madadi, A.; Heard, M.; Saeed, K.; Cooke, J. Using antimicrobial surgihoney to prevent caesarean wound infection. Br. J. Midwifery 2014, 22, 111–115. [Google Scholar] [CrossRef]
- Dryden, M.; Tawse, C.; Adams, J.; Howard, A.; Saeed, K.; Cooke, J. The use of Surgihoney to prevent or eradicate bacterial colonisation in dressing oncology long vascular lines. J. Wound Care 2014, 23, 338–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dryden, M.; Dickinson, A.; Brooks, J.; Hudgell, L.; Saeed, K.; Cutting, K.F. A multi-centre clinical evaluation of reactive oxygen topical wound gel in 114 wounds. J. Wound Care 2016, 25, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Halstead, F.D.; Webber, M.A.; Rauf, M.; Burt, R.; Dryden, M.; Oppenheim, A. In vitro activity of an engineered honey, medical-grade honeys, and antimicrobial wound dressings against biofilm-producing clinical bacterial isolates. J. Wound Care 2016, 25, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Oryan, A.; Alemzadeh, E.; Mohammadi, A.A. Application of honey as a protective material in maintaining the viability of adipose stem cells in burn wound healing: A histological, molecular and biochemical study. Tissue Cell 2019, 61, 89–97. [Google Scholar] [CrossRef]
- Suleman, L. Extracellular Bacterial Proteases in Chronic Wounds: A Potential Therapeutic Target? Adv. Wound Care 2015, 5, 455–463. [Google Scholar] [CrossRef] [Green Version]
- McCarty, S.M.; Cochrane, C.A.; Clegg, P.D.; Percival, S.L. The role of endogenous and exogenous enzymes in chronic wounds: A focus on the implications of aberrant levels of both host and bacterial proteases in wound healing. Wound Repair Regen. 2012, 20, 125–136. [Google Scholar] [CrossRef]
- Hunt, T.K. The Physiology of Wound-Healing. Ann. Emerg. Med. 1988, 17, 1265–1273. [Google Scholar] [CrossRef]
- Winter, G.D. Formation of the scab and the rate of epithelization of superficial wounds in the skin of the young domestic Pig. Nature 1962, 193, 293–294. [Google Scholar] [CrossRef] [PubMed]
- Svensjo, T.; Pomahac, B.; Yao, F.; Slama, J.; Eriksson, E. Accelerated healing of full-thickness skin wounds in a wet environment. Plast. Reconstr. Surg. 2000, 106, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, S.; Ranzato, E. Honey, Wound Repair and Regenerative Medicine. J. Func. Biomater. 2018, 9, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, T.; Eichenlaub, E.H.; Angel, M.F.; Levin, M.; Futrell, J.W. Topical Acidification Promotes Healing of Experimental Deep Partial Thickness Skin Burns: A Randomized Double-Blind Preliminary-Study. Burns 1985, 12, 84–90. [Google Scholar] [CrossRef]
- Greener, B.; Hughes, A.A.; Bannister, N.P.; Douglass, J. Proteases and pH in chronic wounds. J. Wound Care 2005, 14, 59–61. [Google Scholar] [CrossRef]
- Lönnqvist, S.; Emanuelsson, P.; Kratz, G. Influence of acidic pH on keratinocyte function and re-epithelialisation of human in vitro wounds. J. Plast. Surg. Hand Surg. 2015, 49, 346–352. [Google Scholar] [CrossRef]
- Kruse, C.R.; Singh, M.; Targosinski, S.; Sinha, I.; Sørensen, J.A.; Eriksson, E.; Nuutila, K. The effect of pH on cell viability, cell migration, cell proliferation, wound closure, and wound reepithelialization: In vitro and in vivo study. Wound Repair Regen. 2017, 25, 260–269. [Google Scholar] [CrossRef]
- Lambeth, J.D. NOX enzymes and the biology of reactive oxygen. Nat. Rev. Immunol. 2004, 4, 181–189. [Google Scholar] [CrossRef]
- Suh, Y.A.; Arnold, R.S.; Lassegue, B.; Shi, J.; Xu, X.X.; Sorescu, D.; Chung, A.B.; Griendling, K.K.; Lambeth, J.D. Cell transformation by the superoxide-generating oxidase Mox1. Nature 1999, 401, 79–82. [Google Scholar] [CrossRef]
- Lambeth, J.D.; Cheng, G.J.; Arnold, R.S.; Edens, W.A. Novel homologs of gp91phox. Trends Biochem. Sci. 2000, 25, 459–461. [Google Scholar] [CrossRef]
- Al-Jadi, A.M.; Enchang, F.K.; Yusoff, K.M. The effect of Malaysian honey and its major components on the proliferation of cultured fibroblasts. Turk. J. Med. Sci. 2014, 44, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Chung, L.Y.; Schmidt, R.J.; Andrews, A.M.; Turner, T.D. A Study Of Hydrogen-Peroxide Generation By, And Antioxidant Activity Of, Granuflex(Tm) (Duoderm(Tm)) Hydrocolloid Granules And Some Other Hydrogel Hydrocolloid Wound Management Materials. Br. J. Dermatol. 1993, 129, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Stricklin, G.P.; Jeffrey, J.J.; Roswit, W.T.; Eisen, A.Z. Human-Skin Fibroblast Procollagenase: Mechanisms of Activation by Organomercurials And Trypsin. Biochemistry 1983, 22, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Witkosarsat, V.; Descampslatscha, B. Neutrophil-Derived Oxidants and Proteinases as Immunomodulatory Mediators in Inflammation. Mediat. Inflamm. 1994, 3, 257–273. [Google Scholar] [CrossRef] [PubMed]
- Caley, M.P.; Martins, V.L.C.; O’Toole, E.A. Metalloproteinases and Wound Healing. Adv. Wound Care 2015, 4, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M. Restructuring of the extracellular matrix in diabetic wounds and healing: A perspective. Pharmacol. Res. 2016, 107, 243–248. [Google Scholar] [CrossRef]
- Majtan, J.; Kumar, P.; Majtan, T.; Walls, A.F.; Klaudiny, J. Effect of honey and its major royal jelly protein 1 on cytokine and MMP-9 mRNA transcripts in human keratinocytes. Exp. Dermatol. 2010, 19, E73–E79. [Google Scholar] [CrossRef]
- Majtan, J.; Bohova, J.; Garcia-Villalba, R.; Tomas-Barberan, F.A.; Madakova, Z.; Majtan, T.; Majtan, V.; Klaudiny, J. Fir honeydew honey flavonoids inhibit TNF-α-induced MMP-9 expression in human keratinocytes: A new action of honey in wound healing. Arch. Dermatol. Res. 2013, 305, 619–627. [Google Scholar] [CrossRef]
- Santibanez, J.F.; Obradovic, H.; Kukolj, T.; Krstic, J. Transforming growth factor-β, matrix metalloproteinases, and urokinase-type plasminogen activator interaction in the cancer epithelial to mesenchymal transition. Dev. Dyn. 2018, 247, 382–395. [Google Scholar] [CrossRef] [Green Version]
- Cano Sanchez, M.; Lancel, S.; Boulanger, E.; Neviere, R. Targeting Oxidative Stress and Mitochondrial Dysfunction in the Treatment of Impaired Wound Healing: A Systematic Review. Antioxidants 2018, 7, 98. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, M.; Werner, S. Oxidative stress in normal and impaired wound repair. Pharmacol. Res. 2008, 58, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Platzer, M.; Kiese, S.; Tybussek, T.; Herfellner, T.; Schneider, F.; Schweiggert-Weisz, U.; Eisner, P. Radical Scavenging Mechanisms of Phenolic Compounds: A Quantitative Structure-Property Relationship (QSPR) Study. Front. Nutr. 2022, 9, 663. [Google Scholar] [CrossRef] [PubMed]
- Mathew, S.; Abraham, T.E.; Zakaria, Z.A. Reactivity of phenolic compounds towards free radicals under in vitro conditions. J. Food Sci. Technol. 2015, 52, 5790–5798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, T.; Neuman, R.A.; Weinberg, A. Is postburn dermal ischaemia enhanced by oxygen free radicals? Burns 1989, 15, 291–294. [Google Scholar] [CrossRef]
- Tanaka, H.; Hanumadass, M.; Matsuda, H.; Shimazaki, S.; Walter, R.J.; Matsuda, T. Hemodynamic Effects of Delayed Initiation of Antioxidant Therapy (Beginning Two Hours After Burn) in Extensive Third-Degree Burns. J. Burn. Care Rehabil. 1995, 16, 610–615. [Google Scholar] [CrossRef]
- Tang, Y.; Lan, X.; Liang, C.; Zhong, Z.; Xie, R.; Zhou, Y.; Miao, X.; Wang, H.; Wang, W. Honey loaded alginate/PVA nanofibrous membrane as potential bioactive wound dressing. Carbohydr. Polym. 2019, 219, 113–120. [Google Scholar] [CrossRef]
- Yang, X.; Fan, L.; Ma, L.; Wang, Y.; Lin, S.; Yu, F.; Pan, X.; Luo, G.; Zhang, D.; Wang, H. Green electrospun Manuka honey/silk fibroin fibrous matrices as potential wound dressing. Mater. Des. 2017, 119, 76–84. [Google Scholar] [CrossRef]
- Khan, M.Q.; Lee, H.; Khatri, Z.; Kharaghani, D.; Khatri, M.; Ishikawa, T.; Im, S.-S.; Kim, I.S. Fabrication and characterization of nanofibers of honey/poly(1,4-cyclohexane dimethylene isosorbide trephthalate) by electrospinning. Mater. Sci. Eng. C 2017, 81, 247–251. [Google Scholar] [CrossRef]
- Ullah, A.; Ullah, S.; Khan, M.Q.; Hashmi, M.; Nam, P.D.; Kato, Y.; Tamada, Y.; Kim, I.S. Manuka honey incorporated cellulose acetate nanofibrous mats: Fabrication and in vitro evaluation as a potential wound dressing. Int. J. Biol. Macromol. 2020, 155, 479–489. [Google Scholar] [CrossRef]
- Minden-Birkenmaier, B.A.; Neuhalfen, R.M.; Janowiak, B.E.; Sell, S.A. Preliminary Investigation and Characterization of Electrospun Polycaprolactone and Manuka Honey Scaffolds for Dermal Repair. J. Eng. Fibers Fabr. 2015, 10, 155892501501000406. [Google Scholar] [CrossRef]
- Mancuso, E.; Tonda-Turo, C.; Ceresa, C.; Pensabene, V.; Connell, S.D.; Fracchia, L.; Gentile, P. Potential of Manuka Honey as a Natural Polyelectrolyte to Develop Biomimetic Nanostructured Meshes with Antimicrobial Properties. Front. Bioeng. Biotechnol. 2019, 7, 344. [Google Scholar] [CrossRef] [PubMed]
- Kadakia, P.U.; Growney Kalaf, E.A.; Dunn, A.J.; Shornick, L.P.; Sell, S.A. Comparison of silk fibroin electrospun scaffolds with poloxamer and honey additives for burn wound applications. J. Bioact. Compat. Polym. 2017, 33, 79–94. [Google Scholar] [CrossRef]
- Schuhladen, K.; Raghu, S.N.V.; Liverani, L.; Neščáková, Z.; Boccaccini, A.R. Production of a novel poly(ɛ-caprolactone)-methylcellulose electrospun wound dressing by incorporating bioactive glass and Manuka honey. J. Biomed. Mater. Res. Part B Appl. Biomater. 2021, 109, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Hixon, K.R.; Bogner, S.J.; Ronning-Arnesen, G.; Janowiak, B.E.; Sell, S.A. Investigating Manuka Honey Antibacterial Properties When Incorporated into Cryogel, Hydrogel, and Electrospun Tissue Engineering Scaffolds. Gels 2019, 5, 21. [Google Scholar] [CrossRef] [Green Version]
- Hixon, K.R.; Lu, T.; McBride-Gagyi, S.H.; Janowiak, B.E.; Sell, S.A. A comparison of tissue engineering scaffolds incorporated with Manuka honey of varying UMF. BioMed Res. Int. 2017, 2017, 4843065. [Google Scholar] [CrossRef] [Green Version]
- Maleki, H.; Gharehaghaji, A.A.; Dijkstra, P.J. A novel honey-based nanofibrous scaffold for wound dressing application. J. Appl. Polym. Sci. 2013, 127, 4086–4092. [Google Scholar] [CrossRef]
- Balaji, A.; Jaganathan, S.K.; Ismail, A.F.; Rajasekar, R. Fabrication and hemocompatibility assessment of novel polyurethane-based bio-nanofibrous dressing loaded with honey and Carica papaya extract for the management of burn injuries. Int. J. Nanomed. 2016, 11, 4339–4355. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, R.; Ghosh, A.; Barui, A.; Datta, P. Repositing honey incorporated electrospun nanofiber membranes to provide anti-oxidant, anti-bacterial and anti-inflammatory microenvironment for wound regeneration. J. Mater. Sci. Mater. Med. 2018, 29, 31. [Google Scholar] [CrossRef]
- Sarhan, W.A.; Azzazy, H.M.E. High concentration honey chitosan electrospun nanofibers: Biocompatibility and antibacterial effects. Carbohydr. Polym. 2015, 122, 135–143. [Google Scholar] [CrossRef]
- Sarhan, W.A.; Azzazy, H.M.E.; El-Sherbiny, I.M. The effect of increasing honey concentration on the properties of the honey/polyvinyl alcohol/chitosan nanofibers. Mater. Sci. Eng. C 2016, 67, 276–284. [Google Scholar] [CrossRef]
- Ghalei, S.; Li, J.; Douglass, M.; Garren, M.; Handa, H. Synergistic Approach to Develop Antibacterial Electrospun Scaffolds Using Honey and S-Nitroso-N-acetyl Penicillamine. ACS Biomater. Sci. Eng. 2021, 7, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Gaydhane, M.K.; Kanuganti, J.S.; Sharma, C.S. Honey and curcumin loaded multilayered polyvinylalcohol/cellulose acetate electrospun nanofibrous mat for wound healing. J. Mater. Res. 2020, 35, 600–609. [Google Scholar] [CrossRef]
- Kanimozhi, S.; Kathiresan, G.; Kathalingam, A.; Kim, H.-S.; Doss, M.N.R. Organic nanocomposite Band-Aid for chronic wound healing: A novel honey-based nanofibrous scaffold. Appl. Nanosci. 2020, 10, 1639–1652. [Google Scholar] [CrossRef]
- Parin, F.N.; Terzioğlu, P.; Sicak, Y.; Yildirim, K.; Öztürk, M. Pine honey–loaded electrospun poly (vinyl alcohol)/gelatin nanofibers with antioxidant properties. J. Text. Inst. 2021, 112, 628–635. [Google Scholar] [CrossRef]
- Shahid, M.A.; Ali, A.; Uddin, M.N.; Miah, S.; Islam, S.M.; Mohebbullah, M.; Jamal, M.S.I. Antibacterial wound dressing electrospun nanofibrous material from polyvinyl alcohol, honey and Curcumin longa extract. J. Ind. Text. 2020, 51, 455–469. [Google Scholar] [CrossRef]
- Minden-Birkenmaier, B.A.; Smith, R.A.; Radic, M.Z.; van der Merwe, M.; Bowlin, G.L. Manuka Honey Reduces NETosis on an Electrospun Template Within a Therapeutic Window. Polymers 2020, 12, 1430. [Google Scholar] [CrossRef]
- Abou Zekry, S.S.; Abdellatif, A.; Azzazy, H.M.E. Fabrication of pomegranate/honey nanofibers for use as antibacterial wound dressings. Wound Med. 2020, 28, 100181. [Google Scholar] [CrossRef]
- Naeimi, A.; Payandeh, M.; Ghara, A.R.; Ghadi, F.E. In vivo evaluation of the wound healing properties of bio-nanofiber chitosan/polyvinyl alcohol incorporating honey and Nepeta dschuparensis. Carbohydr. Polym. 2020, 240, 116315. [Google Scholar] [CrossRef]
- Ionescu, O.M.; Mignon, A.; Iacob, A.T.; Simionescu, N.; Confederat, L.G.; Tuchilus, C.; Profire, L. New Hyaluronic Acid/Polyethylene Oxide-Based Electrospun Nanofibers: Design, Characterization and In Vitro Biological Evaluation. Polymers 2021, 13, 1291. [Google Scholar] [CrossRef]
- Zhang, Q.; Lin, Z.; Zhang, W.; Huang, T.; Jiang, J.; Ren, Y.; Zhang, R.; Li, W.; Zhang, X.; Tu, Q. Fabrication of green poly(vinyl alcohol) nanofibers using natural deep eutectic solvent for fast-dissolving drug delivery. RSC Adv. 2021, 11, 1012–1021. [Google Scholar] [CrossRef]
- Tavakoli, J.; Tang, Y. Honey/PVA hybrid wound dressings with controlled release of antibioticsStructural, physico-mechanical and in-vitro biomedical studies. Mater. Sci. Eng. C 2017, 77, 318–325. [Google Scholar] [CrossRef]
- Park, J.-S.; An, S.-J.; Jeong, S.-I.; Gwon, H.-J.; Lim, Y.-M.; Nho, Y.-C. Chestnut Honey Impregnated Carboxymethyl Cellulose Hydrogel for Diabetic Ulcer Healing. Polymers 2017, 9, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giusto, G.; Vercelli, C.; Comino, F.; Caramello, V.; Tursi, M.; Gandini, M. A new, easy-to-make pectin-honey hydrogel enhances wound healing in rats. BMC Complement. Altern. Med. 2017, 17, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonifacio, M.A.; Cometa, S.; Cochis, A.; Gentile, P.; Ferreira, A.M.; Azzimonti, B.; Procino, G.; Ceci, E.; Rimondini, L.; De Giglio, E. Antibacterial effectiveness meets improved mechanical properties: Manuka honey/gellan gum composite hydrogels for cartilage repair. Carbohydr. Polym. 2018, 198, 462–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonifacio, M.A.; Cochis, A.; Cometa, S.; Scalzone, A.; Gentile, P.; Procino, G.; Milano, S.; Scalia, A.C.; Rimondini, L.; De Giglio, E. Advances in cartilage repair: The influence of inorganic clays to improve mechanical and healing properties of antibacterial Gellan gum-Manuka honey hydrogels. Mater. Sci. Eng. C 2020, 108, 110444. [Google Scholar] [CrossRef] [PubMed]
- El-Kased, R.F.; Amer, R.I.; Attia, D.; Elmazar, M.M. Honey-based hydrogel: In vitro and comparative In vivo evaluation for burn wound healing. Sci. Rep. 2017, 7, 9692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Zhu, X.K.; Xue, X.T.; Wu, D.Y. Hydrogel sheets of chitosan, honey and gelatin as burn wound dressings. Carbohydr. Polym. 2012, 88, 75–83. [Google Scholar] [CrossRef]
- Shamloo, A.; Aghababaie, Z.; Afjoul, H.; Jami, M.; Bidgoli, M.R.; Vossoughi, M.; Ramazani, A.; Kamyabhesari, K. Fabrication and evaluation of chitosan/gelatin/PVA hydrogel incorporating honey for wound healing applications: An in vitro, in vivo study. Int. J. Pharm. 2021, 592, 120068. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Rajput, M.; Barui, A.; Chatterjee, S.S.; Pal, N.K.; Chatterjee, J.; Mukherjee, R. Dual cross-linked honey coupled 3D antimicrobial alginate hydrogels for cutaneous wound healing. Mater. Sci. Eng. C 2020, 116, 111218. [Google Scholar] [CrossRef] [PubMed]
- Lahooti, B.; Khorram, M.; Karimi, G.; Mohammadi, A.; Emami, A. Modeling and optimization of antibacterial activity of the chitosan-based hydrogel films using central composite design. J. Biomed. Mater. Res. Part A 2016, 104, 2544–2553. [Google Scholar] [CrossRef]
- Giusto, G.; Beretta, G.; Vercelli, C.; Valle, E.; Iussich, S.; Borghi, R.; Odetti, P.; Monacelli, F.; Tramuta, C.; Grego, E.; et al. Pectin-honey hydrogel: Characterization, antimicrobial activity and biocompatibility. Bio-Med. Mater. Eng. 2018, 29, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Afshari, M.J.; Sheikh, N.; Afarideh, H. PVA/CM-chitosan/honey hydrogels prepared by using the combined technique of irradiation followed by freeze-thawing. Radiat. Phys. Chem. 2015, 113, 28–35. [Google Scholar] [CrossRef]
- Hixon, K.R.; Lu, T.; Carletta, M.N.; McBride-Gagyi, S.H.; Janowiak, B.E.; Sell, S.A. A preliminary in vitro evaluation of the bioactive potential of cryogel scaffolds incorporated with Manuka honey for the treatment of chronic bone infections. J. Biomed. Mater. Res. Part B Appl. 2018, 106, 1918–1933. [Google Scholar] [CrossRef] [PubMed]
- Neres Santos, A.M.; Duarte Moreira, A.P.; Piler Carvalho, C.W.; Luchese, R.; Ribeiro, E.; McGuinness, G.B.; Fernandes Mendes, M.; Nunes Oliveira, R. Physically Cross-Linked Gels of PVA with Natural Polymers as Matrices for Manuka Honey Release in Wound-Care Applications. Materials 2019, 12, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saberian, M.; Seyedjafari, E.; Zargar, S.J.; Mahdavi, F.S.; Sanaei-rad, P. Fabrication and characterization of alginate/chitosan hydrogel combined with honey and aloe vera for wound dressing applications. J. Appl. Polym. Sci. 2021, 138, 51398. [Google Scholar] [CrossRef]
- Rajput, M.; Mandal, M.; Anura, A.; Mukhopadhyay, A.; Subramanian, B.; Paul, R.R.; Chatterjee, J. Honey loaded silk fibroin 3D porous scaffold facilitates homeostatic full-thickness wound healing. Materialia 2020, 12, 100703. [Google Scholar] [CrossRef]
- Schuhladen, K.; Mukoo, P.; Liverani, L.; Neščáková, Z.; Boccaccini, A.R. Manuka honey and bioactive glass impart methylcellulose foams with antibacterial effects for wound-healing applications. Biomed. Mater. 2020, 15, 065002. [Google Scholar] [CrossRef] [Green Version]
- Rathinamoorthy, R.; Sasikala, L. In vivo—Wound healing studies of Leptospermum scoparium honey loaded chitosan bioactive wound dressing. Wound Med. 2019, 26, 100162. [Google Scholar] [CrossRef]
- Hall, T.J.; Azoidis, I.; Barroso, I.A.; Hughes, E.A.B.; Grover, L.M.; Cox, S.C. Formulation of an antimicrobial superabsorbent powder that gels in situ to produce reactive oxygen. Mater. Sci. Eng. C 2021, 118, 111479. [Google Scholar] [CrossRef]
- Hall, T.J.; Hughes, E.A.B.; Sajjad, H.; Kuehne, S.A.; Grant, M.M.; Grover, L.M.; Cox, S.C. Formulation of a reactive oxygen producing calcium sulphate cement as an anti-bacterial hard tissue scaffold. Sci. Rep. 2021, 11, 4491. [Google Scholar] [CrossRef]
- Datta, S.; Sarkar, R.; Vyas, V.; Bhutoria, S.; Barui, A.; Roy Chowdhury, A.; Datta, P. Alginate-honey bioinks with improved cell responses for applications as bioprinted tissue engineered constructs. J. Mater. Res. 2018, 33, 2029–2039. [Google Scholar] [CrossRef]
- Andriotis, E.G.; Eleftheriadis, G.K.; Karavasili, C.; Fatouros, D.G. Development of Bio-Active Patches Based on Pectin for the Treatment of Ulcers and Wounds Using 3D-Bioprinting Technology. Pharmaceutics 2020, 12, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Li, T.; Han, Y.; Li, F.; Liu, Y. Recent development of electrospun wound dressing. Curr. Opin. Biomed. Eng. 2021, 17, 100247. [Google Scholar] [CrossRef]
- da Silva, L.P.; Reis, R.L.; Correlo, V.M.; Marques, A.P. Hydrogel-Based Strategies to Advance Therapies for Chronic Skin Wounds. Annu. Rev. Biomed. Eng. 2019, 21, 145–169. [Google Scholar] [CrossRef] [Green Version]
- Palmese, L.L.; Thapa, R.K.; Sullivan, M.O.; Kiick, K.L. Hybrid hydrogels for biomedical applications. Curr. Opin. Chem. Eng. 2019, 24, 143–157. [Google Scholar] [CrossRef]
- Liu, M.; Lu, J.; Müller, P.; Turnbull, L.; Burke, C.M.; Schlothauer, R.C.; Carter, D.A.; Whitchurch, C.B.; Harry, E.J. Antibiotic-specific differences in the response of Staphylococcus aureus to treatment with antimicrobials combined with manuka honey. Front. Microbiol. 2015, 5, 779. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.Y.; Cokcetin, N.N.; Lu, J.; Turnbull, L.; Carter, D.A.; Whitchurch, C.B.; Harry, E.J. Rifampicin-Manuka Honey Combinations Are Superior to Other Antibiotic-Manuka Honey Combinations in Eradicating Staphylococcus aureus Biofilms. Front. Microbiol. 2018, 8, 2653. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.; Millonig, G.; Waite, G.N. The GOX/CAT system: A novel enzymatic method to independently control hydrogen peroxide and hypoxia in cell culture. Adv. Med. Sci. 2009, 54, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Suvarnapathaki, S.; Wu, X.; Lantigua, D.; Nguyen, M.A.; Camci-Unal, G. Breathing life into engineered tissues using oxygen-releasing biomaterials. NPG Asia Mater. 2019, 11, 65. [Google Scholar] [CrossRef] [Green Version]
- Erdem, A.; Darabi, M.A.; Nasiri, R.; Sangabathuni, S.; Ertas, Y.N.; Alem, H.; Hosseini, V.; Shamloo, A.; Nasr, A.S.; Ahadian, S.; et al. 3D Bioprinting of Oxygenated Cell-Laden Gelatin Methacryloyl Constructs. Adv. Healthc. Mater. 2020, 9, 1901794. [Google Scholar] [CrossRef]
- Harrison, B.S.; Eberli, D.; Lee, S.J.; Atala, A.; Yoo, J.J. Oxygen producing biomaterials for tissue regeneration. Biomaterials 2007, 28, 4628–4634. [Google Scholar] [CrossRef] [PubMed]
- Wijekoon, A.; Fountas-Davis, N.; Leipzig, N.D. Fluorinated methacrylamide chitosan hydrogel systems as adaptable oxygen carriers for wound healing. Acta Biomater. 2013, 9, 5653–5664. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.S.; Fountas-Davis, N.; Huang, H.; Michelle Evancho-Chapman, M.; Fulton, J.A.; Shriver, L.P.; Leipzig, N.D. Fluorinated methacrylamide chitosan hydrogels enhance collagen synthesis in wound healing through increased oxygen availability. Acta Biomater. 2016, 36, 164–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dogan-Guner, E.M.; Mohamed, H.; Orbey, N.; Goodyear, N. Stabilization and controlled release of micro-encapsulated hydrogen peroxide for wound treatment applications. J. Appl. Microbiol. 2019, 126, 965–972. [Google Scholar] [CrossRef]
- Abuelgasim, H.; Albury, C.; Lee, J. Effectiveness of honey for symptomatic relief in upper respiratory tract infections: A systematic review and meta-analysis. BMJ Evid.-Based Med. 2020, 26, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Duffy, C.; Sorolla, A.; Wang, E.; Golden, E.; Woodward, E.; Davern, K.; Ho, D.; Johnstone, E.; Pfleger, K.; Redfern, A.; et al. Honeybee venom and melittin suppress growth factor receptor activation in HER2-enriched and triple-negative breast cancer. NPJ Precis. Oncol. 2020, 4, 24. [Google Scholar] [CrossRef] [PubMed]
- Comvita. UMF™ Manuka Honey. Available online: https://www.comvita.co.uk/category/umf-manuka-honey/range00001 (accessed on 28 August 2020).
- Egmonthoney. Products. Available online: https://egmonthoney.co.uk/collections/all (accessed on 28 August 2020).
- Manuka Doctor. Manuka Honey. Available online: https://www.manukadoctor.co.uk/collections/manuka-honey (accessed on 28 August 2020).
- Savant Distribution. MGO Manuka Honey. Available online: https://www.manukahealthuk.com/shop/mgo-manuka-honey.html (accessed on 28 August 2020).
- Manuka Pharm. Manuka Honey. Available online: https://www.manukapharm.co.uk/collections/manuka-honey (accessed on 28 August 2020).
- Melora. Melora Manuka Honey. Available online: https://www.melora.co.uk/collections/manuka-honey-food (accessed on 28 August 2020).
- Optima Health and Nutrition. Premium Select Manuka Honey. Available online: https://optimah.com/pages/pure-gold-premium-manuka-honey (accessed on 28 August 2020).
- Goslinski, M.; Nowak, D.; Klebukowska, L. Antioxidant properties and antimicrobial activity of manuka honey versus Polish honeys. J. Food Sci. Technol. Mysore 2020, 57, 1269–1277. [Google Scholar] [CrossRef]
- Miorin, P.L.; Junior, N.C.L.; Custodio, A.R.; Bretz, W.A.; Marcucci, M.C. Antibacterial activity of honey and propolis from Apis mellifera and Tetragonisca angustula against Staphylococcus aureus. J. Appl. Microbiol. 2003, 95, 913–920. [Google Scholar] [CrossRef]
- Boukraa, L. Additive Activity of Royal Jelly and Honey Against Pseudomonas aeruginosa. Altern. Med. Rev. 2008, 13, 330–333. [Google Scholar]
- Hamid, Z.; Mohamad, I.; Harun, A.; Salim, R.; Sulaiman, S.A. Antifungal Effect of Three Local Malaysian Honeys on Selected Pathogenic Fungi of Otomycosis: An in vitro Evaluation. J. Young Pharm. 2018, 10, 414–417. [Google Scholar] [CrossRef] [Green Version]
- Hau-Yama, N.E.; Magana-Ortiz, D.; Oliva, A.I.; Ortiz-Vazquez, E. Antifungal activity of honey from stingless bee Melipona beecheii against Candida albicans. J. Apic. Res. 2020, 59, 12–18. [Google Scholar] [CrossRef]
- Feas, X.; Estevinho, M.L. A Survey of the In Vitro Antifungal Activity of Heather (Erica sp.) Organic Honey. J. Med. Food 2011, 14, 1284–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erturk, O.; Karakas, F.P.; Pehlivan, D.; Nas, N. The Antibacterial and Antifungal Effects of Rhododendron Derived Mad Honey and Extracts of Four Rhododendron Species. Turk. J. Biol. 2009, 33, 151–158. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product | Manufacturer | Description | Indications | Mechanism of Action | Ref. | Clinical Evidence |
|---|---|---|---|---|---|---|
| Activon® Manuka Honey Tube | Advancis Medical | 100% medical-grade manuka honey | Any wound type but especially sloughy, necrotic, and malodorous wounds, including: pressure ulcers, leg ulcers, diabetic ulcers, surgical wounds, burns, graft sites, infected wounds, cavity wounds and sinuses | Debrides necrotic tissue; can be used in dressings or directly into cavities. | [154] | Inhibition of in vitro formation of clinically important Gram-positive bacteria biofilms [155]. Blistering and cellulitis on a type 2 diabetic patient; paediatric burn; foot ulceration; grade 5 sacral wound [154] |
| Activon® Tulle | Advancis Medical | Knitted viscose mesh dressing impregnated with 100% manuka honey | Granulating or shallow wounds, good when debriding or de-sloughing small areas of necrotic or sloughy tissue | Creates a moist healing environment, eliminates wound odour, and provides antibacterial action | [154] | Overgranulated grade 3 and 4 pressure ulcers; extensive leg cellulitis; venous ulcer; chronic wound infections; necrotic foot [154] |
| Algivon® Plus | Advancis Medical | Reinforced alginate dressing impregnated with 100% manuka honey | Pressure, leg and diabetic ulcers, surgical wounds, burns, graft sites and infected wounds. Ideal for wetter wounds | Absorbs exudate. Debrides, removes slough, and reduces bacterial load | [154] | Chronic wounds [156]; burn wound management [157] |
| Algivon® Plus Ribbon | Advancis Medical | Reinforced alginate ribbon impregnated with 100% manuka honey | Cavities, sinuses, pressure ulcers, leg ulcers, diabetic ulcers, surgical wounds, burns, graft sites, and infected wounds | Absorb exudates. Debrides, removes slough, and reduces bacterial load | [154] | Autoamputation of fingertip necrosis [158] |
| Aurum® ostomy bags | Welland Medical Ltd. | Medical-grade manuka honey added to the hydrocolloid | Stoma care | Kills bacteria, suppresses inflammation, and stimulates the growth of cells to promote healthy skin around the stoma | [159] | Pyoderma gangrenosum around ileostomy [160] |
| L-Mesitran® Border | Aspen Medical Europe Ltd. | Combined hydrogel and honey (30%) pad on a strong fixation layer | Chronic wounds, such as: pressure ulcers; superficial and partial-thickness burns; venous, arterial, and diabetic ulcers. | Exudate absorption. Donates moisture to rehydrate dry tissue. Antibacterial properties. Helps to maintain a moist wound environment | [161] | Paediatric minor burns and scalds [162] |
| L-Mesitran® Hydro | Aspen Medical Europe Ltd. | Sterile, semi-permeable hydrogel dressing containing 30% honey with vitamin C and E, as well as an acrylic polymer gel and water, with a polyurethane film backing | Low to moderate exuding wounds, including: chronic wounds (pressure ulcers, venous and diabetic ulcers), superficial and acute wounds (cuts, abrasions and donor sites), superficial and partial-thickness burns (first- and second-degree), fungating wounds, acute wounds, e.g., donor sites, surgical wounds, cuts and abrasions | Donates moisture to rehydrate dry tissue. Antibacterial properties. Helps to maintain a moist wound environment | [161] | Paediatric minor burns and scalds [162]. Fungating wounds [163] |
| L-Mesitran® Ointment | Aspen Medical Europe Ltd. | Ointment with 48% medical-grade honey, medical-grade hypoallergenic lanolin, oils, and vitamins | Superficial, acute, and chronic wounds. Superficial and partial-thickness burns. Fungating wounds (to help deodorise and debride). Colonised acute wounds and (postoperative) surgical wounds | Aids debridement and reduce bacterial colonisation | [161] | Skin tears; irritation and inflammation [163] |
| ManukaDress IG | Medicareplus International | Wound dressing made with 100% Leptospermum scoparium sterile honey from New Zealand. Non-adherent impregnated gauze | Leg and pressure ulcers, first- and second-degree burns, diabetic foot ulcers, surgical and trauma wounds | Osmotic activity that promotes autolytic debridement and helps maintain a moist wound environment | [164] | Burn management [165]. Difficult-to-debride wounds [166]. Necrotic pressure ulcer; recurrent venous leg ulceration [167] |
| Medihoney® Antibacterial Honey | Derma Sciences—Comvita | 100% sterilised medical-grade manuka honey | All types of wounds with low to moderate exudate, including: deep, sinus, necrotic, infected, surgic and malodorous wounds® | Creates an antibacterial environment (MGO). Autolytic debridement on sloughy and necrotic tissue. Removes malodour. Provides a moist environment. | [168] | Wound healing [169]; prevention of catheter-associated infections in haemodialysis patients [170] |
| Medihoney® Apinate Dressing | Derma Sciences—Comvita | Calcium alginate dressing impregnated with 100% medical-grade manuka honey | Moderately to heavily exuding wounds such as: diabetic foot ulcers, leg ulcers, pressure ulcers (partial- and full-thickness), first- and second-degree partial-thickness burns, donor sites and traumatic or surgical wounds. | Promotes a moisture-balanced environment. Osmotic potential draws fluid through the wound to the surface. Low pH of 3.5–4.5. | [171] | Venous leg ulcers [172] |
| Medihoney® Barrier Cream | Derma Sciences—Comvita | Barrier cream containing 30% medical-grade manuka honey | Use to protect skin from breakdown (e.g., skin damaged by irradiation treatment or in wet areas due to incontinence). Additionally, to prevent damage caused by shear and friction | Maintains skin moisture and pH. | [173] | Treatment for intertrigo in large skin folds [174] |
| Medihoney® Antibacterial Wound Gel™ | Derma Sciences—Comvita | Antibacterial wound gel: 80% medical-grade manuka honey with natural waxes and oils | Surface wounds with low to moderate exudate and partial- and full-thickness wounds, including burns, cuts, grazes, and eczema wounds | Creates a moist, low-pH environment. Cleans the wound through osmotic effect. Reduces the risk of infection (MGO) | [175] | Reduction in incidence of wound infection after microvascular free tissue reconstruction [176] |
| SurgihoneyRO™ | Matoke Holdings Ltd. | Antimicrobial wound gel utilising bioengineered honey to deliver Reactive Oxygen® (RO™) | Infected, chronic (diabetic foot, pressure, and leg ulcers) and acute (surgical, traumatic and abrasions wounds, cuts, burns, donor and recipient sites) wounds | Controlled release of hydrogen peroxide release for antimicrobial activity. Promotes debridement and new tissue growth | [177] | Prevention of caesarean wound infection; prevention/eradication of bacterial colonies in dressing oncology long vascular lines; ulcers, surgical wounds and trauma wounds [178,179,180]. In vitro activity against biofilm-producing clinical bacterial isolates [181] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yupanqui Mieles, J.; Vyas, C.; Aslan, E.; Humphreys, G.; Diver, C.; Bartolo, P. Honey: An Advanced Antimicrobial and Wound Healing Biomaterial for Tissue Engineering Applications. Pharmaceutics 2022, 14, 1663. https://doi.org/10.3390/pharmaceutics14081663
Yupanqui Mieles J, Vyas C, Aslan E, Humphreys G, Diver C, Bartolo P. Honey: An Advanced Antimicrobial and Wound Healing Biomaterial for Tissue Engineering Applications. Pharmaceutics. 2022; 14(8):1663. https://doi.org/10.3390/pharmaceutics14081663
Chicago/Turabian StyleYupanqui Mieles, Joel, Cian Vyas, Enes Aslan, Gavin Humphreys, Carl Diver, and Paulo Bartolo. 2022. "Honey: An Advanced Antimicrobial and Wound Healing Biomaterial for Tissue Engineering Applications" Pharmaceutics 14, no. 8: 1663. https://doi.org/10.3390/pharmaceutics14081663
APA StyleYupanqui Mieles, J., Vyas, C., Aslan, E., Humphreys, G., Diver, C., & Bartolo, P. (2022). Honey: An Advanced Antimicrobial and Wound Healing Biomaterial for Tissue Engineering Applications. Pharmaceutics, 14(8), 1663. https://doi.org/10.3390/pharmaceutics14081663