
Withania somnifera: Progress towards a Pharmaceutical Agent for Immunomodulation and Cancer Therapeutics

 ,
, 
 and
and 
Abstract
:1. Introduction
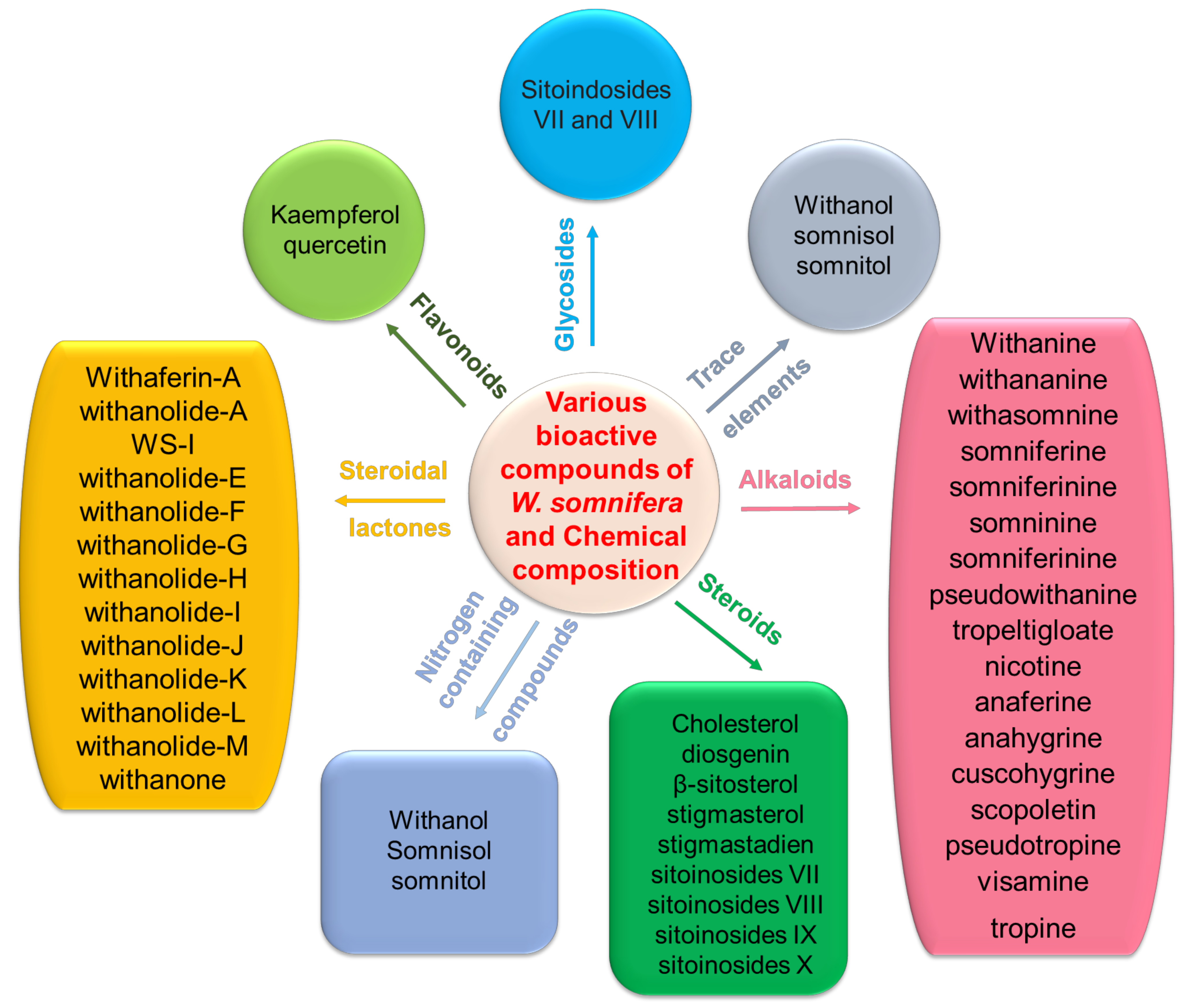
2. Biological and Chemical Properties of Withania somnifera
2.1. Sources and Chemical Properties of Withania somnifera
2.2. Toxicity of Withania somnifera

{kind=link}
{kind=link}
{kind=link}
| Bioactive Molecules | Part Used | Type of Cancer | Medicinal Value of the Bioactive Molecules | Ref. |
|---|---|---|---|---|
| Withaferin A | Leaves | Breast cancer cell lines MDA-MB-231 and MCF-7 | In vitro, WA inhibited the expression of ER, RET, and HSF1 while increasing the expression of phospho-p38 MAPK, p53, and p21 in MCF-7 breast cancer cells | [19] |
| Breast cancer cells and mice model | Inhibit cell proliferation, reduced tumor growth and induce FOXO3a- and Bim-dependent apoptosis | [20] | ||
| Ovarian cancer cells | Inhibits cell growth, inducing apoptosis, and cell cycle arrest and targeting Notch1 and Notch3 down regulates | [55] | ||
| Breast cancer cell lines, 4T1 (mouse breast), Nu/nu mice, Balb/c mice, SCID mice | Chemoprevention and reduced tumor growth | [56] | ||
| Leukemia U937 cells | Induces apoptosis by activating caspase 3, JNK, and Akt signaling pathways | [57] | ||
| Human renal cancer cells (Caki cells) | Increased radiation-induced apoptosis by ROS generation, inhibits the expression of Bcl-2 and dephosphorylation of Akt | [21] | ||
| Human cancer cell lines Balb/c nude mice | Increased expression of p53 transcription factor, suppression of tumour growth and apoptosis | [58] | ||
| Osteogenic sarcoma (U2OS) and fibrosarcoma (HT1080) cells | In vitro and in vivo anticancer activity | [59] | ||
| Root | Balb/C mice | WA reduced macrophage production of pro-inflammatory cytokines, tumor weight, granulocytic MDSC number, and MDSC potential to inhibit antigen-driven activation of CD4+ and CD8+ T cells | [22] | |
| Prostate PC-3 xenografts in nude mice | Inhibition of the proteasomal chymotrypsin-like activity and tumor growth | [60] | ||
| DRO81-(medullary thyroid) and nu/nu mice | Reduced tumor growth and inhibited total and phospho-RET levels at the protein level | [61] | ||
| Malignant pleural mesothelioma (MPM), MPM (H2373, H2452, H2461, H226 and AB12) cells and BALB/c mice | Inhibits the proteasome activity in mesothelioma in vitro and in vivo and reduces tumor growth | [62] | ||
| Pancreatic cancer cell lines Panc-1, MiaPaCa2 and BxPc3 and Panc-1 xenografts mice | Showed potent cytotoxicity against pancreatic cancer cells in vitro, reduced tumor growth and targeted heat shock protein 90 | [63] | ||
| 7,12-dimethylbenz[a]anthracene (DMBA) induced oral carcinogenesis in Syrian golden hamsters | Exhibit anti-lipid peroxidative and antioxidant activity | [64] | ||
| Breast cancer cell lines MCF-7 and SUM159 | Exhibit antiproliferative activity and Induce apoptosis | [65] | ||
| Root | Sarcoma 180, Animal model | Reduced tumor growth | [66] | |
| NR | Cervical cancer cells CaSki, HeLa, SiHa, C33a and athymic nu/nu mice | WA significantly reduced tumor growth inhibit expression of HPV E6/E7 oncogenes and restores the p53 and induces apoptosis | [67] | |
| Breast cancer cell lines and MMTV-neu mice | Inhibit the expression of aldehyde dehydrogenase (ALDH1), CD44 (high)/CD24 (low)/epithelial-specific antigen-positive (ESA+) along with Oct4, SOX-2, Nanog mRNA and inhibition of cancer stem cell growth | [23] | ||
| Colon cancer cell lines C57BL/6-APCMin/+ | Inhibit the expression of interleukin-6, COX-2, TNF-α, pAKT, Notch1, NF-κB and Ki67 | [24] | ||
| Colorectal cancer cells HCT-116 and RKO | Exhibit cell cycle arrest and ROS-dependent mitochondrial dysfunction-mediated apoptosis | [68] | ||
| Root | Gliobastoma multiforme, nu/nu mice | GBM neurosphere collapsed at nM concentrations | [69] | |
| NR | Panc-1, SW1990, MIAPaCa-2, AsPC-1 and BxPc-3 and xenografts mouse model | Reduced tumor growth, activation of proteasome inhibition, and enhancement of ubiquitinated protein accumulation, resulting in ER stress-mediated apoptosis | [70] | |
| Leaves | Lymphoid and myeloid leukemia cells | Induces mitochondrial apoptosis by activating the p38 MAPK cascade | [71] | |
| Myeloid leukemia HL-60 cells | Early ROS generation and mitochondrial dysfunction | [72] | ||
| NR | Prostate cancer cells and PC-3 xenografts | Par-4-Dependent Apoptosis | [73] | |
| Fruit | Liver cancer cells HepG2 | Remarkable changes in the chromatin structure (fragmentation, uniform condensation) | [74] | |
| Root | HUVEC cells | Inhibition of NF-κB by interference with the ubiquitin-mediated proteasome pathway by increasing levels of poly-ubiquitinated proteins | [75] | |
| Leaves | HepG2 (hepatocellular carcinoma) | Increased the expression of Caspase-3; caspase-8, caspase-9, upregulated antioxidant activity and decreased TNF-α level | [76] | |
| Telomerase plus, telomerase negative, ALT (JFCF-1 l and JFCF-4D) | Exhibit cytotoxicity, cause DNA damage, and promote telomere dysfunction | [77] | ||
| NR | Melanoma cells (Lu1205, M14, Mel501 and SK28) | Inhibit cell proliferation, induces apoptosis; downregulated ROS productions and Bcl-2 expression | [78] | |
| NSCLC cancer cell lines A549, CL141, H441, CL97, H1975, CL152, and H1299 and NOD/SCID mice | Reduced lung CSC growth and spheroid formation capacity, mTOR/STAT3 signaling downregulation, and EGFR inhibition | [24,79] | ||
| B cell lymphoma cell line (Lymphatic systems) and Balb/c mice | Decreased cell survival, heat shock protein (Hsp) 90, key kinases and cell cycle regulators | [80] | ||
| Human A549 and U937 cells | Inhibited cell adhesion and reduces the expression of ICAM-1 and VCAM-1 TNF-α and NF-κB | [25] | ||
| Prostate cancer cell lines (PC-3; DU-145 LNCaP) | Promoted cell death and inhibited the expression of c-Fos and heat-shock proteins (HSPs) | [81] | ||
| Leaves | Colorectal cancer cell lines (SW480 and HCT116) | Inhibited cell proliferation, induce apoptosis, cell cycle G2/M arrest and associated with proteasomal degradation of Mad2 and Cdc20 | [82] | |
| Withaferin A and withanone | NR | U2OS (osteosarcoma) and TIG (normal skin fibroblast) cells | Reduced cell viability and induces p53 expression | [83] |
| Withaferin A and CAPE | NR | Human ovarian cancer (SKOV3 and OKV-18 and SKGII, SKGIIIb, ME180) and cervical (HeLa) cancer cells | Exhibited antiproliferative activity and induced apoptosis, increased p53, and downregulated mortalin | [84] |
| Withaferin A and withanone | NR | Hepatocarcinoma HUH-6 and HUH-7 cells | Reduced cell viability and induces apoptosis | [85] |
| 27-acetoxy-4b,6a-dihydroxy-5b-chloro-1-oxowitha-2, 24-dienolide. 5b,6b,14a,15a-diepoxy 4b,27-dihydroxy-1- oxowitha-2,24-dienolide & Withaferin A | Fresh aerial parts | Lung cancer cell line NCI-H460 | Anti-cancer efficacy against human lung cancer cells and growth inhibition | [86] |
| L-asparaginase | Fruits | Human leukemia cells | Exhibited inhibitory effect against lymphoblastic leukemia | [87] |
| Withaferin A and Withanolide D | Root | B16F-10 melanoma cells in C57BL/6 mice | Exhibited significant antitumor activity | [88] |
| Withanolide A | Root | Balb/C mice | Upregulated the Th1 response, CD4 and CD8 numbers, and enhances the activity of natural killer (NK) cells | [89] |
| Withanolide A, Withanoside IV, and Withanoside VI | Root | Human neuroblastoma SH-SY5Y cell line | Activate neurite outgrowth in the SH-SY5Y cell line | [90] |
| Triethylene glycol | Leaves | Different human cancer cells and CD1-ICR mice and Balb/c nude mice | ASH-WEX and TEG are selectively cytotoxic to cancer cells and activate the tumor suppressor proteins p53 and pRB | [91] |
| 27-desoxy-24,25-dihydrowithaferin A | Leaves | Lung (NCI-H460), colon (HCT-116), central nervous system (SF-268) and breast (MCF-7) human tumor cell lines | Reduced viability and inhibited cell proliferation | [92] |
| 27-Oglucopyranosylviscosalactone B | Leaves | Lung (NCI-H460), colon (HCT-116), central nervous system (SF-268) and breast (MCF-7) human tumor cell lines | Reduced viability and inhibited cell proliferation | [92] |
| 3-azido withaferin A | NR | Human cervical (HeLa and prostate (PC-3) cancer cells and C57/BL6J mice | By modulating extracellular Par-4, it prevents cancer cell invasion and angiogenesis | [93] |
| 4,16-dihydroxy-5h,6h-epoxyphysagulin D | Leaves | Lung (NCI-H460), colon (HCT-116), central nervous system (SF-268) and breast (MCF-7) human tumor cell lines | Reduced viability and inhibited cell proliferation | [92] |
| 4β-Hydroxywithanolide E | Aerial parts (stems and leaves) | Human breast cancer cells (MDA-MB-231 and MCF-7) | Inhibition of NF-κB activation | [94] |
| 4β-hydroxywithanolide E, Withaferin A | NR | Triple-negative breast cancer (TNBC) MDA-MB-231 cells | Inhibit cell viability, cell cycle arrest and apoptosis/necrosis | [95] |
| Combination of cucurbitacin B and withanone CucWi-N | NR | A549; TIG-3 and athymic balb/c nude mice | Reduced tumor growth, induces cellular senescence and decreases the expression of Cyclin E, Lamin A/C, CDK2, Cyclin D, CDK4, phosphorylated RB, mortalin and an increase in p53 | [26] |
| Diacetylwithaferin A | Leaves | Lung (NCI-H460), colon (HCT-116), central nervous system (SF-268) and breast (MCF-7) human tumor cell lines | Inhibition of cell proliferation and decrease the expression of COX-2 | [92] |
| Physagulin D (1→ 6)-h-Dglucopyranosyl-(1→4)-h-Dglucopyranoside | Leaves | Lung (NCI-H460), colon (HCT-116), central nervous system (SF-268) and breast (MCF-7) human tumor cell lines | Inhibition of cell proliferation reduces viability and decrease the expression of COX-2 | [92] |
| Viscosalactone B | Leaves | Lung (NCI-H460), colon (HCT-116), central nervous system (SF-268) and breast (MCF-7) human tumor cell lines | Inhibition of cell proliferation reduces viability and decreases the expression of COX-2 | [92] |
| 27-desoxy-24,25-dihydrowithaferin A | Leaves | Lung (NCI-H460), colon (HCT-116), central nervous system (SF-268) and breast (MCF-7) tumor cell lines | Inhibition of cell proliferation and reduced viability | [92] |
| Withanolide analogue | NR | Breast cancer cells (SK-Br-3 and MCF7/BUS) | Inhibition of cell proliferation and upregulation the expression of β-tubulin | [96] |
| Withanolide D | Leaves | Myeloid (K562) and lymphoid (MOLT-4) cells | Induced apoptosis and cell killing through JNK and p38MAPK activation | [97] |
| Withanolide D | Root | Multiple myeloma CSCs and RPMI 8226 cell | Inhibition of cell proliferation and cell death induces apoptosis | [98] |
| Withanone and withaferin A (20:1) | NR | Metastatic cancer cells A172, IMR32, YKG1, MCF7, HT1080, U20S and Nude mice | Inhibition of cell proliferation and downregulation the expression of hnRNP-K, VEGF, and metalloproteases | |
| Withanoside IV | Leaves | Lung (NCI-H460), colon (HCT-116), central nervous system (SF-268) and breast (MCF-7) human tumor cell lines | Inhibition of cell proliferation and decrease the expression of COX-2 | [92] |
| Withania somnifera leaf extract | Leaves | Human glioma cell line (YKG1, U118MG and A172) | Inhibition of cell proliferation and increased the expression of NCAM and mortalin | [99] |
| Withania somnifera leaf extract and withaferin | Root | Human normal fibroblasts (TIG-3), breast carcinoma (MCF7), colon carcinoma (HCT116) | Increased DNA damage, oxidative stress, and downregulation of TPX2, TFAP2A, LHX3, and ING1 | [100] |
| Withania somnifera root extract | Root | Human prostate cancer cells (LNCaP and 22Rv1) | Inhibition of cell proliferation, fatty acid synthesis and downregulation of the expression of c-Myc and p-Akt (S473) | [101] |
| Withania somnifera root extract | Root | Prostate cancer cells (PC3) | Inhibition of cell proliferation, cell cycle arrest in G2/M phase and downregulation of the expression of IL-8 and COX-2 | [102] |
| Withania somnifera roots extract and cisplatin | Root | Breast (MDA-MB-231) and colon (HT-29) cancer cells | Inhibition of cell proliferation, increased mitochondrial dysfunction, and ROS | [103] |
2.3. Pharmacokinetic Studies and Bioavailability of Withania somnifera
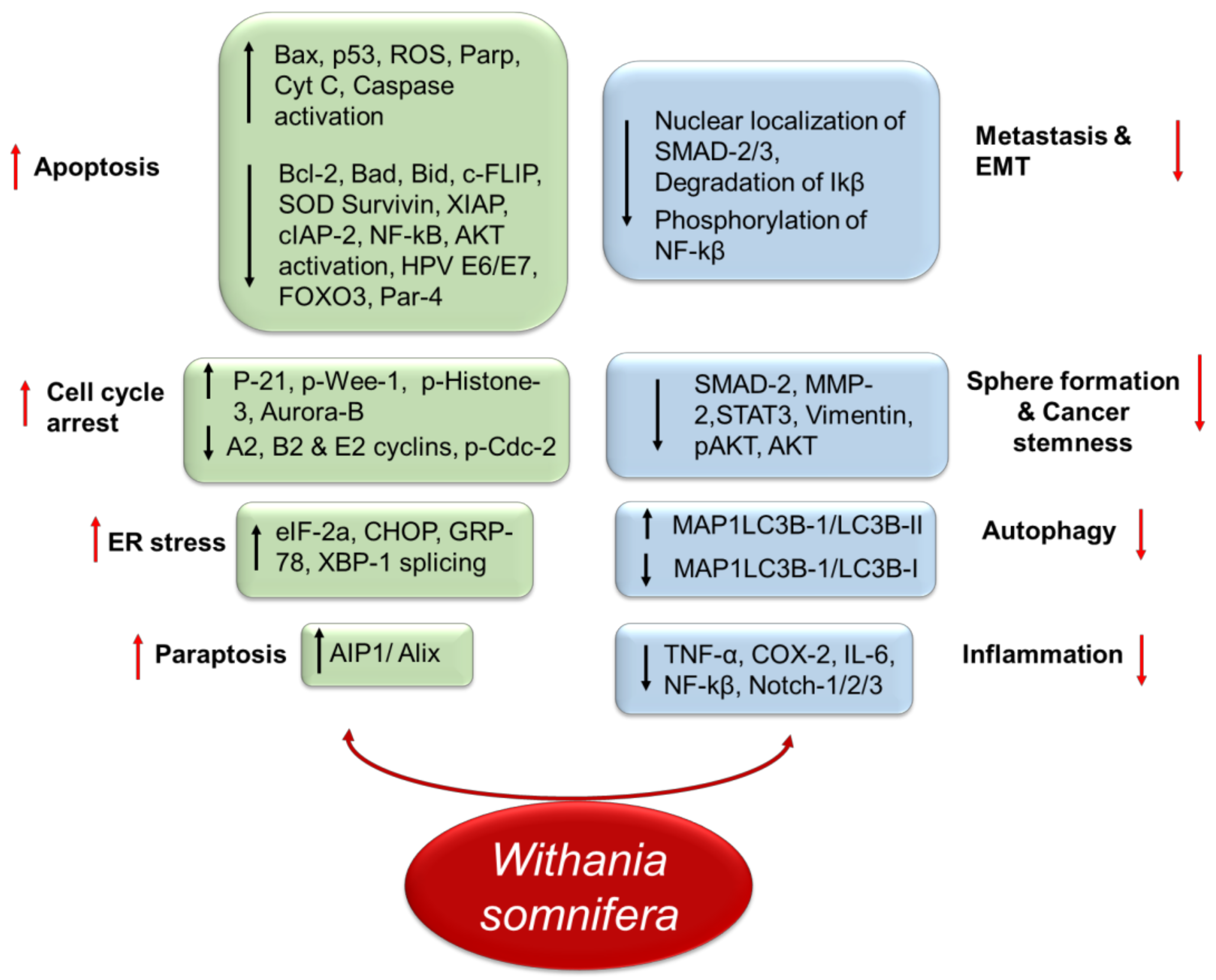
3. Role of Withania somnifera in Cancer
3.1. Lung Cancer
3.2. Breast Cancer
3.3. Prostate Cancer
3.4. Colon Cancer
3.5. Leukemia
3.6. Other Cancers
3.7. Chemosensitization and Synergistic Actions of Withania somnifera
| Cancer | Cell Line | Targeted Molecular Mechanisms | Ref. |
|---|---|---|---|
| Lung cancer | A549 | Cell cycle arrest ↑; PI3K/Akt pathway↓ | [109] |
| H1299 and A549 | TGF-β and TNF-α induced EMT ↓; nuclear translocation of Smad 2/3 and NF-κB ↓ | [110] | |
| H1299, CL141, CL149, and A549 | ROS, autophagy, and apoptosis ↑; mTOR/STAT3 signaling ↓ | [79] | |
| Breast cancer | MCF7 and MDA-MB-231 | G2/M phase cell cycle arrest ↑; ROS generation and apoptosis ↑; ER-a, XIAP, cIAP-2 and survivin ↓ | [19,20] |
| MCF7 and MDA-MB-231 | Cell migration, EMT and invasion ↓; IL6 induced STAT3 activation ↓; Notch2 and Notch4 ↑; mitochondrial membrane potential ↓ | [100,122,125] | |
| SUM-159 and MCF-7 | Mammosphere formation ↓, ALDH1 activity ↓, bCSCs↓; | [23] | |
| Glioblastomas | GL26, U251, and U87 | Cell proliferation ↓; G2/M phase cell cycle arrest ↑; ROS generation ↑; Akt/mTOR and MAPK pathway ↓ | [151,152] |
| Microglial | BV2 | Nrf-2 and HO-1 ↑; filopodia formation ↓ | [153] |
| Neuroblastomas | IMR-32, U87-MG, C6, GBM39, and GBM2 | Cell proliferation ↓; G0/G1 cell cycle arrest ↑; Cyclin D1 ↓; p-Akt, PSA-NCAM, Bcl-xL, MMP-2, MMP-9 ↓ | [69,154,155] |
| Oral | CAL27 and Ca9-22 | Cell proliferation ↓; G1 phase cell cycle arrest ↑; ROS generation, DNA damage and mitochondrial membrane depolarization ↑ | [137] |
| Osteosarcoma | U2OS and MG-63 | Cell proliferation ↓; G2/M phase cell cycle arrest ↑; cyclin B1, cyclin A ↓; p-Chk1, p-Chk2 ↑ | [138] |
| Leukemia | THP-1, HL-60, MDS-L, and Ramos | Apoptosis↑; G2/M phase cell cycle arrest ↑; ROS ↑ | [134,135] |
| Prostate | DU 145 and PC3, | Cell proliferation ↓; G2/M Phase cell cycle arrest ↑; ROS and autophagy ↑ | [81,127] |
| Ovarian cancer | CaOV3, SKOV3, and A2780 | Cell proliferation ↓; apoptosis ↑; ROS ↑; G2/M cell cycle arrest↑; Notch1, Notch2, otch3, Bcl-2, Akt ↓ | [141,142] |
| Melanoma | Lu1205, M14, Mel501, and SK28 | Cell viability ↓; apoptosis ↑; ROS↑; DNA fragmentation and mitochondrial membrane depolarization ↑ | [78] |
| Gastric cancer | AGS | Cell viability ↓; Apoptosis ↑; G2/M cell cycle arrest ↑; ROS ↑; Cell migration and invasion ↓ | [139] |
| Gastrointestinal | UP-LN1 | Apoptosis ↑; CXCR4/CXCL12 and STAT3/IL-6 axis ↓ | [156] |
| Thyroid cancers | SW1736 and BCPAP | BRAF, Raf-1 and, ERK ↓; cell cycle arrest at G2/M phase ↑ | [157] |
3.8. Clinical Trials
3.9. Immunomodulatory Activity and Hematopoiesis Actions of Withania somnifera
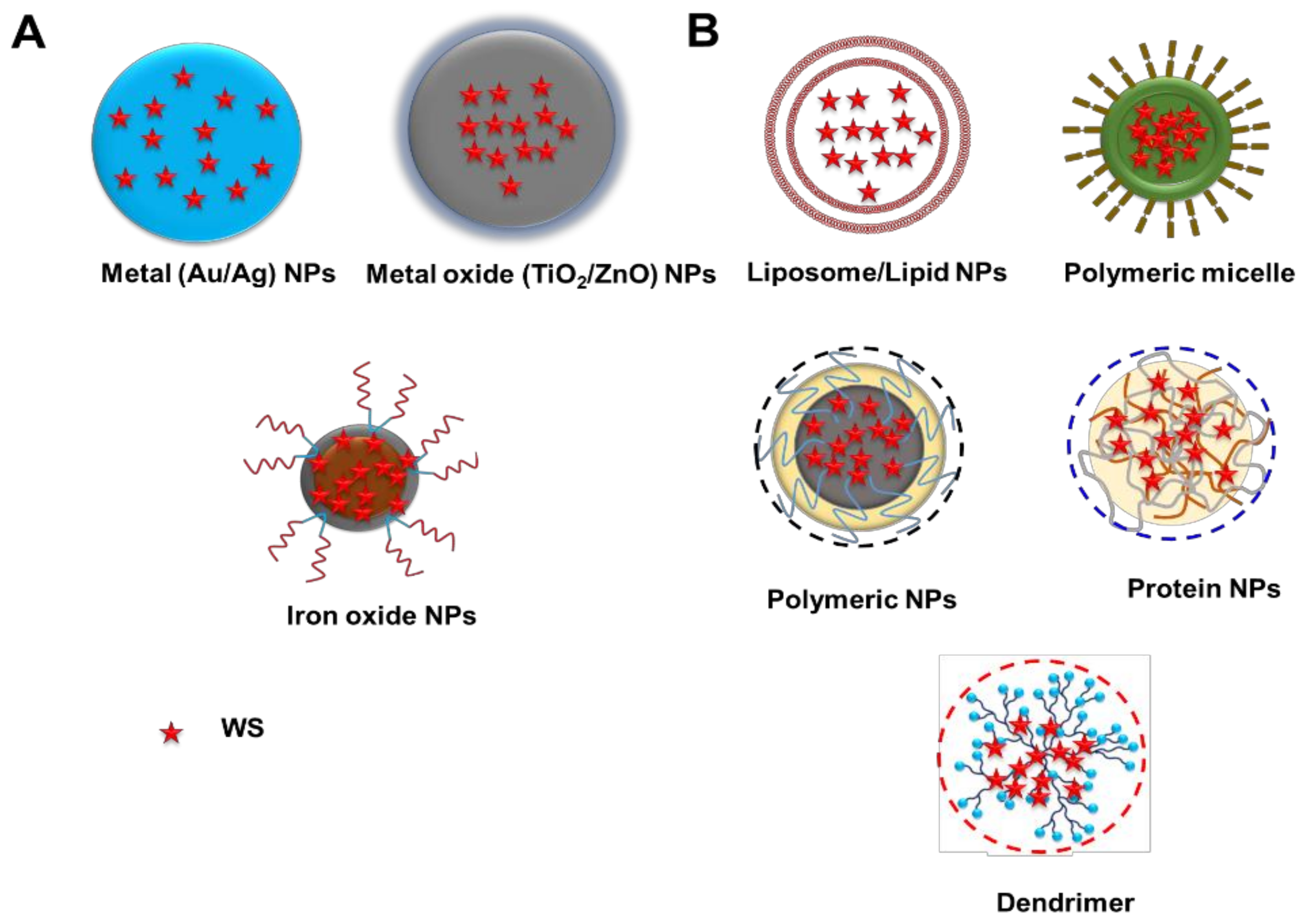
4. Nanotechnology-Based Strategies for the Delivery of WS
4.1. Inorganic Nanoparticles
4.1.1. Gold Nanoparticles (AuNPs)
4.1.2. Titanium Oxide Nanoparticles (TiO2 NPs)
4.1.3. Silver Nanoparticles (AgNPs)
4.1.4. Zinc Oxide Nanoparticles (ZnO NPs)
4.2. Organic Nanoparticles
5. Future Perspective of WS Delivery
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- The Centers for Disease Control and Prevention, an Update on Cancer Deaths in the United States. Available online: https://www.cdc.gov/cancer/dcpc/research/update-on-cancer-deaths/index.htm (accessed on 3 March 2022).
- Banik, K.; Harsha, C.; Bordoloi, D.; Lalduhsaki Sailo, B.; Sethi, G.; Leong, H.C.; Arfuso, F.; Mishra, S.; Wang, L.; Kumar, A.P.; et al. Therapeutic potential of gambogic acid, a caged xanthone, to target cancer. Cancer Lett. 2018, 416, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Banik, K.; Bordoloi, D.; Harsha, C.; Sailo, B.L.; Padmavathi, G.; Roy, N.K.; Gupta, S.C.; Aggarwal, B.B. Googling the Guggul (Commiphora and Boswellia) for Prevention of Chronic Diseases. Front. Pharmacol. 2018, 9, 686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manu, K.A.; Shanmugam, M.K.; Ramachandran, L.; Li, F.; Siveen, K.S.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; Arfuso, F.; Kumar, A.P.; et al. Isorhamnetin augments the anti-tumor effect of capecitabine through the negative regulation of NF-κB signaling cascade in gastric cancer. Cancer Lett. 2015, 363, 28–36. [Google Scholar] [CrossRef]
- Manu, K.A.; Shanmugam, M.K.; Ramachandran, L.; Li, F.; Fong, C.W.; Kumar, A.P.; Tan, P.; Sethi, G. First evidence that γ-tocotrienol inhibits the growth of human gastric cancer and chemosensitizes it to capecitabine in a xenograft mouse model through the modulation of NF-κB pathway. Clin. Cancer Res. 2012, 18, 2220–2229. [Google Scholar] [CrossRef] [Green Version]
- Seca, A.M.L.; Pinto, D. Plant Secondary Metabolites as Anticancer Agents: Successes in Clinical Trials and Therapeutic Application. Int. J. Mol. Sci. 2018, 19, 263. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, M.K.; Lee, J.H.; Chai, E.Z.; Kanchi, M.M.; Kar, S.; Arfuso, F.; Dharmarajan, A.; Kumar, A.P.; Ramar, P.S.; Looi, C.Y.; et al. Cancer prevention and therapy through the modulation of transcription factors by bioactive natural compounds. Semin. Cancer Biol. 2016, 40–41, 35–47. [Google Scholar] [CrossRef]
- Patel, S.M.; Nagulapalli Venkata, K.C.; Bhattacharyya, P.; Sethi, G.; Bishayee, A. Potential of neem (Azadirachta indica L.) for prevention and treatment of oncologic diseases. Semin. Cancer Biol. 2016, 40–41, 100–115. [Google Scholar] [CrossRef]
- Singh, N.; Bhalla, M.; de Jager, P.; Gilca, M. An overview on ashwagandha: A Rasayana (rejuvenator) of Ayurveda. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 208–213. [Google Scholar] [CrossRef]
- Dhuley, J.N. Effect of ashwagandha on lipid peroxidation in stress-induced animals. J. Ethnopharmacol. 1998, 60, 173–178. [Google Scholar] [CrossRef]
- Mehta, V.; Chander, H.; Munshi, A. Mechanisms of Anti-Tumor Activity of Withania somnifera (Ashwagandha). Nutr. Cancer 2021, 73, 914–926. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Jogee, P.S.; Agarkar, G.; dos Santos, C.A. Anticancer activities of Withania somnifera: Current research, formulations, and future perspectives. Pharm. Biol. 2016, 54, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, R.; Khalil, R.; Green, R.; Mohapatra, S.S.; Mohapatra, S. Withania somnifera (Ashwagandha) and Withaferin A: Potential in Integrative Oncology. Int. J. Mol. Sci. 2019, 20, 5310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashyap, V.K.; Dhasmana, A.; Yallapu, M.M.; Chauhan, S.C.; Jaggi, M. Withania somnifera as a potential future drug molecule for COVID-19. Future Drug Discov. 2020, 2, Fdd50. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.A.; Farshori, N.N.; Al-Oqail, M.M.; Pant, A.B.; Al-Khedhairy, A.A. Neuroprotective Effects of Withania somnifera on 4-Hydroxynonenal Induced Cell Death in Human Neuroblastoma SH-SY5Y Cells Through ROS Inhibition and Apoptotic Mitochondrial Pathway. Neurochem. Res. 2021, 46, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.Y.; Wang, Y.Y.; Lan, T.H.; Lin, L.C.; Yuan, S.F.; Tang, J.Y.; Chang, H.W. Low Dose Combined Treatment with Ultraviolet-C and Withaferin a Enhances Selective Killing of Oral Cancer Cells. Antioxidants 2020, 9, 1120. [Google Scholar] [CrossRef] [PubMed]
- Heidari, Z.; Mahmoudzadeh-Sagheb, H.; Sarbishegi, M.; Gorgich, E.A.C. Withania coagulans extract attenuates oxidative stress-mediated apoptosis of cerebellar purkinje neurons after ischemia/reperfusion injury. Metab. Brain Dis. 2021, 36, 1699–1708. [Google Scholar] [CrossRef]
- Zhang, X.; Mukerji, R.; Samadi, A.K.; Cohen, M.S. Down-regulation of estrogen receptor-alpha and rearranged during transfection tyrosine kinase is associated with withaferin a-induced apoptosis in MCF-7 breast cancer cells. BMC Complement. Altern. Med. 2011, 11, 84. [Google Scholar] [CrossRef] [Green Version]
- Stan, S.D.; Hahm, E.-R.; Warin, R.; Singh, S.V. Withaferin A causes FOXO3a-and Bim-dependent apoptosis and inhibits growth of human breast cancer cells in vivo. Cancer Res. 2008, 68, 7661–7669. [Google Scholar] [CrossRef] [Green Version]
- Yang, E.S.; Choi, M.J.; Kim, J.H.; Choi, K.S.; Kwon, T.K. Withaferin A enhances radiation-induced apoptosis in Caki cells through induction of reactive oxygen species, Bcl-2 downregulation and Akt inhibition. Chem.-Biol. Interact. 2011, 190, 9–15. [Google Scholar] [CrossRef]
- Sinha, P.; Ostrand-Rosenberg, S. Myeloid-derived suppressor cell function is reduced by Withaferin A, a potent and abundant component of Withania somnifera root extract. Cancer Immunol. Immunother. 2013, 62, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Singh, S.V. Mammary cancer chemoprevention by withaferin A is accompanied by in vivo suppression of self-renewal of cancer stem cells. Cancer Prev. Res. 2014, 7, 738–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekaran, B.; Pal, D.; Kolluru, V.; Tyagi, A.; Baby, B.; Dahiya, N.R.; Youssef, K.; Alatassi, H.; Ankem, M.K.; Sharma, A.K. The chemopreventive effect of withaferin A on spontaneous and inflammation-associated colon carcinogenesis models. Carcinogenesis 2018, 39, 1537–1547. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Kwon, T.K. Withaferin A inhibits tumor necrosis factor-α-induced expression of cell adhesion molecules by inactivation of Akt and NF-κB in human pulmonary epithelial cells. Int. Immunopharmacol. 2009, 9, 614–619. [Google Scholar] [CrossRef]
- Garg, S.; Huifu, H.; Kumari, A.; Sundar, D.; Kaul, S.C.; Wadhwa, R. Induction of Senescence in Cancer Cells by a Novel Combination of Cucurbitacin B and Withanone: Molecular Mechanism and Therapeutic Potential. J. Gerontol. Ser. A 2019, 75, 1031–1041. [Google Scholar] [CrossRef]
- Dubey, S.; Yoon, H.; Cohen, M.S.; Nagarkatti, P.; Nagarkatti, M.; Karan, D. Withaferin A Associated Differential Regulation of Inflammatory Cytokines. Front. Immunol. 2018, 9, 195. [Google Scholar] [CrossRef] [Green Version]
- Tewari, D.; Chander, V.; Dhyani, A.; Sahu, S.; Gupta, P.; Patni, P.; Kalick, L.S.; Bishayee, A. Withania somnifera (L.) Dunal: Phytochemistry, structure-activity relationship, and anticancer potential. Phytomedicine 2022, 98, 153949. [Google Scholar] [CrossRef]
- Choudhary, D.; Bhattacharyya, S.; Bose, S. Efficacy and Safety of Ashwagandha (Withania somnifera (L.) Dunal) Root Extract in Improving Memory and Cognitive Functions. J. Diet Suppl. 2017, 14, 599–612. [Google Scholar] [CrossRef]
- Choudhary, D.; Bhattacharyya, S.; Joshi, K. Body Weight Management in Adults Under Chronic Stress Through Treatment With Ashwagandha Root Extract: A Double-Blind, Randomized, Placebo-Controlled Trial. J. Evid. Based Complement. Altern. Med. 2017, 22, 96–106. [Google Scholar] [CrossRef] [Green Version]
- Salve, J.; Pate, S.; Debnath, K.; Langade, D. Adaptogenic and Anxiolytic Effects of Ashwagandha Root Extract in Healthy Adults: A Double-blind, Randomized, Placebo-controlled Clinical Study. Cureus 2019, 11, e6466. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekhar, K.; Kapoor, J.; Anishetty, S. A prospective, randomized double-blind, placebo-controlled study of safety and efficacy of a high-concentration full-spectrum extract of ashwagandha root in reducing stress and anxiety in adults. Indian J. Psychol. Med. 2012, 34, 255–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, S.K.; Dhir, A. Withania somnifera: An Indian ginseng. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Dar, N.J.; Hamid, A.; Ahmad, M. Pharmacologic overview of Withania somnifera, the Indian Ginseng. Cell Mol. Life Sci. 2015, 72, 4445–4460. [Google Scholar] [CrossRef] [PubMed]
- Gorelick, J.; Rosenberg, R.; Smotrich, A.; Hanuš, L.; Bernstein, N. Hypoglycemic activity of withanolides and elicitated Withania somnifera. Phytochemistry 2015, 116, 283–289. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, S. Evaluation of anti-inflammatory effect of Withania somnifera root on collagen-induced arthritis in rats. Pharm. Biol. 2014, 52, 308–320. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, I.R.; Arya, D.S.; Gupta, S.K. Withania somnifera provides cardioprotection and attenuates ischemia-reperfusion induced apoptosis. Clin. Nutr. 2008, 27, 635–642. [Google Scholar] [CrossRef]
- RajaSankar, S.; Manivasagam, T.; Sankar, V.; Prakash, S.; Muthusamy, R.; Krishnamurti, A.; Surendran, S. Withania somnifera root extract improves catecholamines and physiological abnormalities seen in a Parkinson’s disease model mouse. J. Ethnopharmacol. 2009, 125, 369–373. [Google Scholar] [CrossRef]
- Bhatnagar, M.; Goel, I.; Roy, T.; Shukla, S.D.; Khurana, S. Complete Comparison Display (CCD) evaluation of ethanol extracts of Centella asiatica and Withania somnifera shows that they can non-synergistically ameliorate biochemical and behavioural damages in MPTP induced Parkinson’s model of mice. PLoS ONE 2017, 12, e0177254. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, S.; Dayakar, A.; Veronica, J.; Sundar, S.; Maurya, R. An in vitro study of apoptotic like death in Leishmania donovani promastigotes by withanolides. Parasitol. Int. 2013, 62, 253–261. [Google Scholar] [CrossRef]
- Sachdeva, H.; Sehgal, R.; Kaur, S. Studies on the protective and immunomodulatory efficacy of Withania somnifera along with cisplatin against experimental visceral leishmaniasis. Parasitol. Res. 2013, 112, 2269–2280. [Google Scholar] [CrossRef]
- Reuland, D.J.; Khademi, S.; Castle, C.J.; Irwin, D.C.; McCord, J.M.; Miller, B.F.; Hamilton, K.L. Upregulation of phase II enzymes through phytochemical activation of Nrf2 protects cardiomyocytes against oxidant stress. Free Radic. Biol. Med. 2013, 56, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Budhiraja, R.D.; Sudhir, S. Review of biological activity of withanolides. J. Sci. Ind. Res. 1987, 46, 488–491. [Google Scholar]
- Rastogi RP, M.B. Compendium of Indian Medicinal Plants; Central Drug Research Institute: New Delhi, India, 1998; p. 6. [Google Scholar]
- Bone, K. Clinical applications of Ayurvedic and Chinese herbs. In Monograph for the Western Herbal Practitioner; Phytotherapy Press: Warwick, QLD, Australia, 1996; pp. 137–141. [Google Scholar]
- Elsakka, M.; Grigorescu, E.; Stănescu, U.; Stănescu, U.; Dorneanu, V. New data referring to chemistry of Withania somnifera species. Rev. Med. Chir. Soc. Med. Nat. Iasi 1990, 94, 385–387. [Google Scholar]
- Srivastava, D.N.; Deshpande, S.M. Gas chromatographic identification of fatty acids, fatty alcohols, and hydrocarbons of Convolvulus pluricaulis (Chois). J. Am. Oil Chem. Soc. 1975, 52, 318–319. [Google Scholar] [CrossRef]
- Vedi, M.; Sabina, E.P. Assessment of hepatoprotective and nephroprotective potential of withaferin A on bromobenzene-induced injury in Swiss albino mice: Possible involvement of mitochondrial dysfunction and inflammation. Cell Biol. Toxicol. 2016, 32, 373–390. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Guo, R.; Gan, L.; Lau, W.B.; Cao, X.; Zhao, J.; Ma, X.; Christopher, T.A.; Lopez, B.L.; Wang, Y. Withaferin A inhibits apoptosis via activated Akt-mediated inhibition of oxidative stress. Life Sci. 2018, 211, 91–101. [Google Scholar] [CrossRef]
- Tiruveedi, V.L.; Bale, S.; Khurana, A.; Godugu, C. Withaferin A, a novel compound of Indian ginseng (Withania somnifera), ameliorates Cerulein-induced acute pancreatitis: Possible role of oxidative stress and inflammation. Phytother. Res. 2018, 32, 2586–2596. [Google Scholar] [CrossRef]
- Palliyaguru, D.L.; Chartoumpekis, D.V.; Wakabayashi, N.; Skoko, J.J.; Yagishita, Y.; Singh, S.V.; Kensler, T.W. Withaferin A induces Nrf2-dependent protection against liver injury: Role of Keap1-independent mechanisms. Free Radic. Biol. Med. 2016, 101, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Bale, S.; Pulivendala, G.; Godugu, C. Withaferin A attenuates bleomycin-induced scleroderma by targeting FoxO3a and NF-κβ signaling: Connecting fibrosis and inflammation. Biofactors 2018, 44, 507–517. [Google Scholar] [CrossRef]
- Anwar, M.F.; Yadav, D.; Rastogi, S.; Arora, I.; Khar, R.K.; Chander, J.; Samim, M. Modulation of liver and kidney toxicity by herb Withania somnifera for silver nanoparticles: A novel approach for harmonizing between safety and use of nanoparticles. Protoplasma 2015, 252, 547–558. [Google Scholar] [CrossRef]
- Biswal, B.M.; Sulaiman, S.A.; Ismail, H.C.; Zakaria, H.; Musa, K.I. Effect of Withania somnifera (Ashwagandha) on the development of chemotherapy-induced fatigue and quality of life in breast cancer patients. Integr. Cancer Ther. 2013, 12, 312–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Samadi, A.K.; Roby, K.F.; Timmermann, B.; Cohen, M.S. Inhibition of cell growth and induction of apoptosis in ovarian carcinoma cell lines CaOV3 and SKOV3 by natural withanolide Withaferin A. Gynecol. Oncol. 2012, 124, 606–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaiparambil, J.T.; Bender, L.; Ganesh, T.; Kline, E.; Patel, P.; Liu, Y.; Tighiouart, M.; Vertino, P.M.; Harvey, R.D.; Garcia, A.; et al. Withaferin A inhibits breast cancer invasion and metastasis at sub-cytotoxic doses by inducing vimentin disassembly and serine 56 phosphorylation. Int. J. Cancer 2011, 129, 2744–2755. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Lee, T.-J.; Kim, S.H.; Choi, Y.H.; Lee, S.H.; Lee, J.M.; Kim, Y.-H.; Park, J.-W.; Kwon, T.K. Induction of apoptosis by withaferin A in human leukemia U937 cells through down-regulation of Akt phosphorylation. Apoptosis 2008, 13, 1494–1504. [Google Scholar] [CrossRef]
- Widodo, N.; Kaur, K.; Shrestha, B.G.; Takagi, Y.; Ishii, T.; Wadhwa, R.; Kaul, S.C. Selective killing of cancer cells by leaf extract of Ashwagandha: Identification of a tumor-inhibitory factor and the first molecular insights to its effect. Clin. Cancer Res. 2007, 13, 2298–2306. [Google Scholar] [CrossRef] [Green Version]
- Widodo, N.; Takagi, Y.; Shrestha, B.G.; Ishii, T.; Kaul, S.C.; Wadhwa, R. Selective killing of cancer cells by leaf extract of Ashwagandha: Components, activity and pathway analyses. Cancer Lett. 2008, 262, 37–47. [Google Scholar] [CrossRef]
- Yang, H.; Shi, G.; Dou, Q.P. The tumor proteasome is a primary target for the natural anticancer compound Withaferin A isolated from “Indian winter cherry”. Mol. Pharmacol. 2007, 71, 426–437. [Google Scholar] [CrossRef]
- Samadi, A.K.; Mukerji, R.; Shah, A.; Timmermann, B.N.; Cohen, M.S. A novel RET inhibitor with potent efficacy against medullary thyroid cancer in vivo. Surgery 2010, 148, 1228–1236. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wang, Y.; Cheryan, V.T.; Wu, W.; Cui, C.Q.; Polin, L.A.; Pass, H.I.; Dou, Q.P.; Rishi, A.K.; Wali, A. Withaferin A inhibits the proteasome activity in mesothelioma in vitro and in vivo. PLoS ONE 2012, 7, e41214. [Google Scholar] [CrossRef]
- Yu, Y.; Hamza, A.; Zhang, T.; Gu, M.; Zou, P.; Newman, B.; Li, Y.; Gunatilaka, A.A.; Zhan, C.G.; Sun, D. Withaferin A targets heat shock protein 90 in pancreatic cancer cells. Biochem. Pharmacol. 2010, 79, 542–551. [Google Scholar] [CrossRef] [Green Version]
- Manoharan, S.; Panjamurthy, K.; Menon, V.P.; Balakrishnan, S.; Alias, L.M. Protective effect of Withaferin-A on tumour formation in 7,12-dimethylbenz[a]anthracene induced oral carcinogenesis in hamsters. Indian J. Exp. Biol. 2009, 47, 16–23. [Google Scholar] [PubMed]
- Hahm, E.R.; Lee, J.; Singh, S.V. Role of mitogen-activated protein kinases and Mcl-1 in apoptosis induction by withaferin A in human breast cancer cells. Mol. Carcinog. 2014, 53, 907–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devi, P.U.; Sharada, A.C.; Solomon, F.E.; Kamath, M.S. In vivo growth inhibitory effect of Withania somnifera (Ashwagandha) on a transplantable mouse tumor, Sarcoma 180. Indian J. Exp. Biol. 1992, 30, 169–172. [Google Scholar] [PubMed]
- Munagala, R.; Kausar, H.; Munjal, C.; Gupta, R.C. Withaferin A induces p53-dependent apoptosis by repression of HPV oncogenes and upregulation of tumor suppressor proteins in human cervical cancer cells. Carcinogenesis 2011, 32, 1697–1705. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Miao, Y.; Liu, S. Withaferin A induces apoptosis by ROS-dependent mitochondrial dysfunction in human colorectal cancer cells. Biochem. Biophys. Res. Commun. 2018, 503, 2363–2369. [Google Scholar] [CrossRef]
- Chang, E.; Pohling, C.; Natarajan, A.; Witney, T.H.; Kaur, J.; Xu, L.; Gowrishankar, G.; D’Souza, A.L.; Murty, S.; Schick, S.; et al. AshwaMAX and Withaferin A inhibits gliomas in cellular and murine orthotopic models. J. Neurooncol. 2016, 126, 253–264. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhu, F.; Jiang, J.; Sun, C.; Zhong, Q.; Shen, M.; Wang, X.; Tian, R.; Shi, C.; Xu, M.; et al. Simultaneous inhibition of the ubiquitin-proteasome system and autophagy enhances apoptosis induced by ER stress aggravators in human pancreatic cancer cells. Autophagy 2016, 12, 1521–1537. [Google Scholar] [CrossRef] [Green Version]
- Mandal, C.; Dutta, A.; Mallick, A.; Chandra, S.; Misra, L.; Sangwan, R.S.; Mandal, C. Withaferin A induces apoptosis by activating p38 mitogen-activated protein kinase signaling cascade in leukemic cells of lymphoid and myeloid origin through mitochondrial death cascade. Apoptosis 2008, 13, 1450–1464. [Google Scholar] [CrossRef]
- Malik, F.; Kumar, A.; Bhushan, S.; Khan, S.; Bhatia, A.; Suri, K.A.; Qazi, G.N.; Singh, J. Reactive oxygen species generation and mitochondrial dysfunction in the apoptotic cell death of human myeloid leukemia HL-60 cells by a dietary compound withaferin A with concomitant protection by N-acetyl cysteine. Apoptosis 2007, 12, 2115–2133. [Google Scholar] [CrossRef]
- Srinivasan, S.; Ranga, R.S.; Burikhanov, R.; Han, S.S.; Chendil, D. Par-4-dependent apoptosis by the dietary compound withaferin A in prostate cancer cells. Cancer Res. 2007, 67, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Abutaha, N. In vitro antiproliferative activity of partially purified Withania somnifera fruit extract on different cancer cell lines. J. Balkan Union Oncol. 2015, 20, 625–630. [Google Scholar]
- Mohan, R.; Hammers, H.J.; Bargagna-Mohan, P.; Zhan, X.H.; Herbstritt, C.J.; Ruiz, A.; Zhang, L.; Hanson, A.D.; Conner, B.P.; Rougas, J.; et al. Withaferin A is a potent inhibitor of angiogenesis. Angiogenesis 2004, 7, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Mofed, D.; Zekri, A.-R.; El-Sayed, N.; Rahouma, M.; Sabet, S. Antioxidant activity and apoptotic induction as mechanisms of action of Withania somnifera (Ashwagandha) against a hepatocellular carcinoma cell line. J. Int. Med. Res. 2018, 46, 1358–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Katiyar, S.P.; Sundar, D.; Kaul, Z.; Miyako, E.; Zhang, Z.; Kaul, S.C.; Reddel, R.R.; Wadhwa, R. Withaferin-A kills cancer cells with and without telomerase: Chemical, computational and experimental evidences. Cell Death Dis. 2017, 8, e2755. [Google Scholar] [CrossRef] [Green Version]
- Mayola, E.; Gallerne, C.; Esposti, D.D.; Martel, C.; Pervaiz, S.; Larue, L.; Debuire, B.; Lemoine, A.; Brenner, C.; Lemaire, C. Withaferin A induces apoptosis in human melanoma cells through generation of reactive oxygen species and down-regulation of Bcl-2. Apoptosis 2011, 16, 1014–1027. [Google Scholar] [CrossRef]
- Hsu, J.H.; Chang, P.M.; Cheng, T.S.; Kuo, Y.L.; Wu, A.T.; Tran, T.H.; Yang, Y.H.; Chen, J.M.; Tsai, Y.C.; Chu, Y.S.; et al. Identification of Withaferin A as a Potential Candidate for Anti-Cancer Therapy in Non-Small Cell Lung Cancer. Cancers 2019, 11, 1003. [Google Scholar] [CrossRef] [Green Version]
- McKenna, M.K.; Gachuki, B.W.; Alhakeem, S.S.; Oben, K.N.; Rangnekar, V.M.; Gupta, R.C.; Bondada, S. Anti-cancer activity of withaferin A in B-cell lymphoma. Cancer Biol. Ther. 2015, 16, 1088–1098. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, Y.; Okuzaki, D.; Fukushima, K.; Mukai, S.; Ohno, S.; Ozaki, Y.; Yabuta, N.; Nojima, H. Withaferin A Induces Cell Death Selectively in Androgen-Independent Prostate Cancer Cells but Not in Normal Fibroblast Cells. PLoS ONE 2015, 10, e0134137. [Google Scholar] [CrossRef]
- Das, T.; Roy, K.S.; Chakrabarti, T.; Mukhopadhyay, S.; Roychoudhury, S. Withaferin A modulates the Spindle assembly checkpoint by degradation of Mad2-Cdc20 complex in colorectal cancer cell lines. Biochem. Pharmacol. 2014, 91, 31–39. [Google Scholar] [CrossRef]
- Vaishnavi, K.; Saxena, N.; Shah, N.; Singh, R.; Manjunath, K.; Uthayakumar, M.; Kanaujia, S.P.; Kaul, S.C.; Sekar, K.; Wadhwa, R. Differential Activities of the Two Closely Related Withanolides, Withaferin A and Withanone: Bioinformatics and Experimental Evidences. PLoS ONE 2012, 7, e44419. [Google Scholar] [CrossRef] [Green Version]
- Sari, A.N.; Bhargava, P.; Dhanjal, J.K.; Putri, J.F.; Radhakrishnan, N.; Shefrin, S.; Ishida, Y.; Terao, K.; Sundar, D.; Kaul, S.C.; et al. Combination of Withaferin-A and CAPE Provides Superior Anticancer Potency: Bioinformatics and Experimental Evidence to Their Molecular Targets and Mechanism of Action. Cancers 2020, 12, 1160. [Google Scholar] [CrossRef] [PubMed]
- Sundar, D.; Yu, Y.; Katiyar, S.P.; Putri, J.F.; Dhanjal, J.K.; Wang, J.; Sari, A.N.; Kolettas, E.; Kaul, S.C.; Wadhwa, R. Wild type p53 function in p53Y220C mutant harboring cells by treatment with Ashwagandha derived anticancer withanolides: Bioinformatics and experimental evidence. J. Exp. Clin. Cancer Res. 2019, 38, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, M.I.; Hussain, S.; Yousuf, S.; Dar, A.; Mudassar; Attaur, R. Chlorinated and diepoxy withanolides from Withania somnifera and their cytotoxic effects against human lung cancer cell line. Phytochemistry 2010, 71, 2205–2209. [Google Scholar] [CrossRef]
- Oza, V.P.; Parmar, P.P.; Kumar, S.; Subramanian, R.B. Anticancer properties of highly purified L-asparaginase from Withania somnifera L. against acute lymphoblastic leukemia. Appl. Biochem. Biotechnol. 2010, 160, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- Leyon, P.V.; Kuttan, G. Effect of Withania somnifera on B16F-10 melanoma induced metastasis in mice. Phytother. Res. 2004, 18, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Bani, S.; Gautam, M.; Sheikh, F.A.; Khan, B.; Satti, N.K.; Suri, K.A.; Qazi, G.N.; Patwardhan, B. Selective Th1 up-regulating activity of Withania somnifera aqueous extract in an experimental system using flow cytometry. J. Ethnopharmacol. 2006, 107, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Nakamura, N.; Hattori, M.; Kuboyama, T.; Tohda, C.; Komatsu, K. Withanolide derivatives from the roots of Withania somnifera and their neurite outgrowth activities. Chem. Pharm. Bull. 2002, 50, 760–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadhwa, R.; Singh, R.; Gao, R.; Shah, N.; Widodo, N.; Nakamoto, T.; Ishida, Y.; Terao, K.; Kaul, S.C. Water Extract of Ashwagandha Leaves Has Anticancer Activity: Identification of an Active Component and Its Mechanism of Action. PLoS ONE 2013, 8, e77189. [Google Scholar] [CrossRef]
- Jayaprakasam, B.; Zhang, Y.; Seeram, N.P.; Nair, M.G. Growth inhibition of human tumor cell lines by withanolides from Withania somnifera leaves. Life Sci. 2003, 74, 125–132. [Google Scholar] [CrossRef]
- Rah, B.; Amin, H.; Yousuf, K.; Khan, S.; Jamwal, G.; Mukherjee, D.; Goswami, A. A novel MMP-2 inhibitor 3-azidowithaferin A (3-azidoWA) abrogates cancer cell invasion and angiogenesis by modulating extracellular Par-4. PLoS ONE 2012, 7, e44039. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.-C.; Tsai, Y.-L.; Wu, Y.-C.; Chang, F.-R.; Liu, M.-H.; Chen, W.-Y.; Wu, C.-C. Withanolides-Induced Breast Cancer Cell Death Is Correlated with Their Ability to Inhibit Heat Protein 90. PLoS ONE 2012, 7, e37764. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-C.; Hu, H.-H.; Chang, F.-R.; Tsai, J.-Y.; Kuo, C.-Y.; Wu, Y.-C.; Wu, C.-C. Different effects of 4β-hydroxywithanolide E and withaferin A, two withanolides from Solanaceae plants, on the Akt signaling pathway in human breast cancer cells. Phytomedicine 2019, 53, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.K.; Kumar, S.; Saloni; Singh, H.; Kim, M.H.; Sharma, P.; Misra, S.; Khan, F. Molecular docking, QSAR and ADMET studies of withanolide analogs against breast cancer. Drug Des. Dev. Ther. 2017, 11, 1859–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Mandal, C.; Sangwan, R.; Chandra, S.; Mandal, C. Withanolide D induces apoptosis in leukemia by targeting the activation of neutral sphingomyelinase-ceramide cascade mediated by synergistic activation of c-Jun N-terminal kinase and p38 mitogen-activated protein kinase. Mol. Cancer 2010, 9, 239. [Google Scholar] [CrossRef] [Green Version]
- Issa, M.E.; Wijeratne, E.M.K.; Gunatilaka, A.A.L.; Cuendet, M. Withanolide D Exhibits Similar Cytostatic Effect in Drug-Resistant and Drug-Sensitive Multiple Myeloma Cells. Front. Pharmacol. 2017, 8, 610. [Google Scholar] [CrossRef] [Green Version]
- Kataria, H.; Shah, N.; Kaul, S.C.; Wadhwa, R.; Kaur, G. Water extract of ashwagandha leaves limits proliferation and migration, and induces differentiation in glioma cells. Evid.-Based Complement. Altern. 2011, 2011, 267614. [Google Scholar] [CrossRef] [Green Version]
- Widodo, N.; Priyandoko, D.; Shah, N.; Wadhwa, R.; Kaul, S.C. Selective Killing of Cancer Cells by Ashwagandha Leaf Extract and Its Component Withanone Involves ROS Signaling. PLoS ONE 2010, 5, e13536. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Singh, K.B.; Hahm, E.-R.; Lokeshwar, B.L.; Singh, S.V. Withania somnifera root extract inhibits fatty acid synthesis in prostate cancer cells. J. Tradit. Complement. Med. 2020, 10, 188–197. [Google Scholar] [CrossRef]
- Setty Balakrishnan, A.; Nathan, A.A.; Kumar, M.; Ramamoorthy, S.; Ramia Mothilal, S.K. Withania somnifera targets interleukin-8 and cyclooxygenase-2 in human prostate cancer progression. Prostate Int. 2017, 5, 75–83. [Google Scholar] [CrossRef]
- Henley, A.B.; Yang, L.; Chuang, K.-L.; Sahuri-Arisoylu, M.; Wu, L.-H.; Bligh, S.W.A.; Bell, J.D. Withania somnifera Root Extract Enhances Chemotherapy through ‘Priming’. PLoS ONE 2017, 12, e0170917. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Harsha, C.; Banik, K.; Vikkurthi, R.; Sailo, B.L.; Bordoloi, D.; Gupta, S.C.; Aggarwal, B.B. Is curcumin bioavailability a problem in humans: Lessons from clinical trials. Expert. Opin. Drug Metab. Toxicol. 2019, 15, 705–733. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Jiang, W.; Guo, Z.; Wang, Z.; Huang, M.; Zhong, G.; Liang, C.; Pei, X.; Dai, R. Studies on oral bioavailability and first-pass metabolism of withaferin A in rats using LC-MS/MS and Q-TRAP. Biomed. Chromatogr. 2019, 33, e4573. [Google Scholar] [CrossRef]
- Patil, D.; Gautam, M.; Mishra, S.; Karupothula, S.; Gairola, S.; Jadhav, S.; Pawar, S.; Patwardhan, B. Determination of withaferin A and withanolide A in mice plasma using high-performance liquid chromatography-tandem mass spectrometry: Application to pharmacokinetics after oral administration of Withania somnifera aqueous extract. J. Pharm. Biomed. Anal. 2013, 80, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Senthilnathan, P.; Padmavathi, R.; Banu, S.M.; Sakthisekaran, D. Enhancement of antitumor effect of paclitaxel in combination with immunomodulatory Withania somnifera on benzo (a) pyrene induced experimental lung cancer. Chem.-Biol. Interact. 2006, 159, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Senthilnathan, P.; Padmavathi, R.; Magesh, V.; Sakthisekaran, D. Chemotherapeutic efficacy of paclitaxel in combination with Withania somnifera on benzo (a) pyrene-induced experimental lung cancer. Cancer Sci. 2006, 97, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Sheng, Z.-Y.; Chen, Y.; Bai, C. Effect of Withaferin A on A549 cellular proliferation and apoptosis in non-small cell lung cancer. Asian Pac. J. Cancer Prev. 2014, 15, 1711–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyakulaga, A.H.; Aqil, F.; Munagala, R.; Gupta, R.C. Withaferin A inhibits Epithelial to Mesenchymal Transition in Non-Small Cell Lung Cancer Cells. Sci. Rep. 2018, 8, 15737. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Arpino, G.; Milano, M.; De Placido, S. Features of aggressive breast cancer. Breast 2015, 24, 594–600. [Google Scholar] [CrossRef] [Green Version]
- Stan, S.D.; Zeng, Y.; Singh, S.V. Ayurvedic medicine constituent withaferin a causes G2 and M phase cell cycle arrest in human breast cancer cells. Nutr. Cancer 2008, 60, 51–60. [Google Scholar] [CrossRef]
- Hahm, E.-R.; Singh, S.V. Withaferin A-induced apoptosis in human breast cancer cells is associated with suppression of inhibitor of apoptosis family protein expression. Cancer Lett. 2013, 334, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahm, E.-R.; Moura, M.B.; Kelley, E.E.; Van Houten, B.; Shiva, S.; Singh, S.V. Withaferin A-induced apoptosis in human breast cancer cells is mediated by reactive oxygen species. PLoS ONE 2011, 6, e23354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargagna-Mohan, P.; Hamza, A.; Kim, Y.-e.; Ho, Y.K.A.; Mor-Vaknin, N.; Wendschlag, N.; Liu, J.; Evans, R.M.; Markovitz, D.M.; Zhan, C.-G. The tumor inhibitor and antiangiogenic agent withaferin A targets the intermediate filament protein vimentin. Chem. Biol. 2007, 14, 623–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Garcia, A.; Xu, S.; Powell, D.R.; Vertino, P.M.; Singh, S.; Marcus, A.I. Withania somnifera root extract inhibits mammary cancer metastasis and epithelial to mesenchymal transition. PLoS ONE 2013, 8, e75069. [Google Scholar] [CrossRef] [Green Version]
- Khazal, K.F.; Hill, D.L. Withania somnifera extract reduces the invasiveness of MDA-MB-231 breast cancer and inhibits cytokines associated with metastasis. J. Cancer Metastasis Treat. 2015, 1, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Khazal, K.F.; Samuel, T.; Hill, D.L.; Grubbs, C.J. Effect of an extract of Withania somnifera root on estrogen receptor-positive mammary carcinomas. Anticancer Res. 2013, 33, 1519–1523. [Google Scholar]
- Hahm, E.R.; Lee, J.; Huang, Y.; Singh, S.V. Withaferin a suppresses estrogen receptor-α expression in human breast cancer cells. Mol. Carcinog. 2011, 50, 614–624. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Hahm, E.R.; Marcus, A.I.; Singh, S.V. Withaferin A inhibits experimental epithelial-mesenchymal transition in MCF-10A cells and suppresses vimentin protein level in vivo in breast tumors. Mol. Carcinog. 2015, 54, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Hahm, E.-R.; Singh, S.V. Withaferin A inhibits activation of signal transducer and activator of transcription 3 in human breast cancer cells. Carcinogenesis 2010, 31, 1991–1998. [Google Scholar] [CrossRef] [Green Version]
- Mulabagal, V.; Subbaraju, G.V.; Rao, C.V.; Sivaramakrishna, C.; DeWitt, D.L.; Holmes, D.; Sung, B.; Aggarwal, B.B.; Tsay, H.S.; Nair, M.G. Withanolide sulfoxide from Aswagandha roots inhibits nuclear transcription factor-kappa-B, cyclooxygenase and tumor cell proliferation. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2009, 23, 987–992. [Google Scholar] [CrossRef]
- Lee, J.; Sehrawat, A.; Singh, S.V. Withaferin A causes activation of Notch2 and Notch4 in human breast cancer cells. Breast Cancer Res. Treat. 2012, 136, 45–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagalingam, A.; Kuppusamy, P.; Singh, S.V.; Sharma, D.; Saxena, N.K. Mechanistic elucidation of the antitumor properties of withaferin a in breast cancer. Cancer Res. 2014, 74, 2617–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vel Szic, K.S.; Declerck, K.; Crans, R.A.; Diddens, J.; Scherf, D.B.; Gerhäuser, C.; Berghe, W.V. Epigenetic silencing of triple negative breast cancer hallmarks by Withaferin A. Oncotarget 2017, 8, 40434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, R.V.; Suman, S.; Das, T.P.; Luevano, J.E.; Damodaran, C. Withaferin A, a steroidal lactone from Withania somnifera, induces mitotic catastrophe and growth arrest in prostate cancer cells. J. Nat. Prod. 2013, 76, 1909–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunimasa, K.; Nagano, T.; Shimono, Y.; Dokuni, R.; Kiriu, T.; Tokunaga, S.; Tamura, D.; Yamamoto, M.; Tachihara, M.; Kobayashi, K.; et al. Glucose metabolism-targeted therapy and withaferin A are effective for epidermal growth factor receptor tyrosine kinase inhibitor-induced drug-tolerant persisters. Cancer Sci. 2017, 108, 1368–1377. [Google Scholar] [CrossRef] [Green Version]
- Muralikrishnan, G.; Dinda, A.K.; Shakeel, F. Immunomodulatory effects of Withania somnifera on azoxymethane induced experimental colon cancer in mice. Immunol. Investig. 2010, 39, 688–698. [Google Scholar] [CrossRef]
- Koduru, S.; Kumar, R.; Srinivasan, S.; Evers, M.B.; Damodaran, C. Notch-1 inhibition by Withaferin-A: A therapeutic target against colon carcinogenesis. Mol. Cancer Ther. 2010, 9, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.Y.; Kim, B.-W. Withaferin-A inhibits colon cancer cell growth by blocking STAT3 transcriptional activity. J. Cancer Prev. 2015, 20, 185. [Google Scholar] [CrossRef] [Green Version]
- Alnuqaydan, A.M.; Rah, B.; Almutary, A.G.; Chauhan, S.S. Synergistic antitumor effect of 5-fluorouracil and withaferin-A induces endoplasmic reticulum stress-mediated autophagy and apoptosis in colorectal cancer cells. Am. J. Cancer Res. 2020, 10, 799. [Google Scholar]
- Yang, E.S.; Choi, M.J.; Kim, J.H.; Choi, K.S.; Kwon, T.K. Combination of withaferin A and X-ray irradiation enhances apoptosis in U937 cells. Toxicol. In Vitro 2011, 25, 1803–1810. [Google Scholar] [CrossRef]
- Okamoto, S.; Tsujioka, T.; Suemori, S.; Kida, J.; Kondo, T.; Tohyama, Y.; Tohyama, K. Withaferin A suppresses the growth of myelodysplasia and leukemia cell lines by inhibiting cell cycle progression. Cancer Sci. 2016, 107, 1302–1314. [Google Scholar] [CrossRef] [PubMed]
- Turrini, E.; Calcabrini, C.; Sestili, P.; Catanzaro, E.; de Gianni, E.; Diaz, A.R.; Hrelia, P.; Tacchini, M.; Guerrini, A.; Canonico, B.; et al. Withania somnifera Induces Cytotoxic and Cytostatic Effects on Human T Leukemia Cells. Toxins 2016, 8, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, J.; Gupta, S.K.; Dinda, A.K. Withania somnifera root extract prevents DMBA-induced squamous cell carcinoma of skin in Swiss albino mice. Nutr. Cancer. 2002, 42, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-W.; Li, R.-N.; Wang, H.-R.; Liu, J.-R.; Tang, J.-Y.; Huang, H.-W.; Chan, Y.-H.; Yen, C.-Y. Withaferin A induces oxidative stress-mediated apoptosis and DNA damage in oral cancer cells. Front. Physiol. 2017, 8, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, T.Z.; Wang, G.S. Antiproliferation potential of withaferin A on human osteosarcoma cells via the inhibition of G2/M checkpoint proteins. Exp. Ther. Med. 2015, 10, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.; Kim, T.H.; Hwang, E.H.; Chang, K.T.; Hong, J.J.; Park, J.H. Withaferin A inhibits the proliferation of gastric cancer cells by inducing G2/M cell cycle arrest and apoptosis. Oncol. Lett. 2017, 14, 416–422. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhu, F.; Jiang, J.; Sun, C.; Wang, X.; Shen, M.; Tian, R.; Shi, C.; Xu, M.; Peng, F. Synergistic antitumor activity of withaferin A combined with oxaliplatin triggers reactive oxygen species-mediated inactivation of the PI3K/AKT pathway in human pancreatic cancer cells. Cancer lett. 2015, 357, 219–230. [Google Scholar] [CrossRef]
- Fong, M.Y.; Jin, S.; Rane, M.; Singh, R.K.; Gupta, R.; Kakar, S.S. Withaferin A synergizes the therapeutic effect of doxorubicin through ROS-mediated autophagy in ovarian cancer. PLoS ONE 2012, 7, e42265. [Google Scholar] [CrossRef] [Green Version]
- Kakar, S.S.; Ratajczak, M.Z.; Powell, K.S.; Moghadamfalahi, M.; Miller, D.M.; Batra, S.K.; Singh, S.K. Withaferin a alone and in combination with cisplatin suppresses growth and metastasis of ovarian cancer by targeting putative cancer stem cells. PLoS ONE 2014, 9, e107596. [Google Scholar] [CrossRef]
- Lee, D.H.; Lim, I.-H.; Sung, E.-G.; Kim, J.-Y.; Song, I.-H.; Park, Y.K.; Lee, T.-J. Withaferin A inhibits matrix metalloproteinase-9 activity by suppressing the Akt signaling pathway. Oncol. Rep. 2013, 30, 933–938. [Google Scholar] [CrossRef] [Green Version]
- Um, H.J.; Min, K.-j.; Kim, D.E.; Kwon, T.K. Withaferin A inhibits JAK/STAT3 signaling and induces apoptosis of human renal carcinoma Caki cells. Biochem. Biophys. Res. Commun. 2012, 427, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.J.; Park, E.J.; Min, K.J.; Park, J.W.; Kwon, T.K. Endoplasmic reticulum stress mediates withaferin A-induced apoptosis in human renal carcinoma cells. Toxicol In Vitro 2011, 25, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Heyninck, K.; Lahtela-Kakkonen, M.; Van der Veken, P.; Haegeman, G.; Berghe, W.V. Withaferin A inhibits NF-kappaB activation by targeting cysteine 179 in IKKβ. Biochem. Pharmacol. 2014, 91, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.K.; Pandey, K.; Panda, M.; Spinella, M.J.; Rengasamy, K.R.R.; Biswal, B.K. The potential of retinoids for combination therapy of lung cancer: Updates and future directions. Pharmacol. Res. 2019, 147, 104331. [Google Scholar] [CrossRef]
- Kyakulaga, A.H.; Aqil, F.; Munagala, R.; Gupta, R.C. Synergistic combinations of paclitaxel and withaferin A against human non-small cell lung cancer cells. Oncotarget 2020, 11, 1399–1416. [Google Scholar] [CrossRef] [Green Version]
- Suttana, W.; Mankhetkorn, S.; Poompimon, W.; Palagani, A.; Zhokhov, S.; Gerlo, S.; Haegeman, G.; Berghe, W.V. Differential chemosensitization of P-glycoprotein overexpressing K562/Adr cells by withaferin A and Siamois polyphenols. Mol. Cancer 2010, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- Rah, B.; ur Rasool, R.; Nayak, D.; Yousuf, S.K.; Mukherjee, D.; Kumar, L.D.; Goswami, A. PAWR-mediated suppression of BCL2 promotes switching of 3-azido withaferin A (3-AWA)-induced autophagy to apoptosis in prostate cancer cells. Autophagy 2015, 11, 314–331. [Google Scholar] [CrossRef]
- Grogan, P.T.; Sleder, K.D.; Samadi, A.K.; Zhang, H.; Timmermann, B.N.; Cohen, M.S. Cytotoxicity of withaferin A in glioblastomas involves induction of an oxidative stress-mediated heat shock response while altering Akt/mTOR and MAPK signaling pathways. Investig. New Drugs 2013, 31, 545–557. [Google Scholar] [CrossRef] [Green Version]
- Grogan, P.T.; Sarkaria, J.N.; Timmermann, B.N.; Cohen, M.S. Oxidative cytotoxic agent withaferin A resensitizes temozolomide-resistant glioblastomas via MGMT depletion and induces apoptosis through Akt/mTOR pathway inhibitory modulation. Investig. New Drugs. 2014, 32, 604–617. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.Y.; Li, R.; Cui, J.; Hannink, M.; Gu, Z.; Fritsche, K.L.; Lubahn, D.B.; Simonyi, A. Withania somnifera and Its Withanolides Attenuate Oxidative and Inflammatory Responses and Up-Regulate Antioxidant Responses in BV-2 Microglial Cells. Neuromolecular Med. 2016, 18, 241–252. [Google Scholar] [CrossRef]
- Kataria, H.; Wadhwa, R.; Kaul, S.C.; Kaur, G. Withania somnifera water extract as a potential candidate for differentiation based therapy of human neuroblastomas. PLoS ONE 2013, 8, e55316. [Google Scholar] [CrossRef] [PubMed]
- Kataria, H.; Kumar, S.; Chaudhary, H.; Kaur, G. Withania somnifera Suppresses Tumor Growth of Intracranial Allograft of Glioma Cells. Mol. Neurobiol. 2016, 53, 4143–4158. [Google Scholar] [CrossRef] [PubMed]
- Ting, L.-L.; Chou, A.S.-B.; Hsieh, C.-H.; Hsiung, S.-C.; Pang, S.-T.; Liao, S.-K. Withaferin A targeting both cancer stem cells and metastatic cancer stem cells in the UP-LN1 carcinoma cell model. J. Cancer Metastatis Treat. 2016, 2, 29–40. [Google Scholar]
- Cohen, S.M.; Mukerji, R.; Timmermann, B.N.; Samadi, A.K.; Cohen, M.S. A novel combination of withaferin A and sorafenib shows synergistic efficacy against both papillary and anaplastic thyroid cancers. Am. J. Surg. 2012, 204, 895–900. [Google Scholar] [CrossRef] [PubMed]
- Pilot Study of Curcumin Formulation and Ashwagandha Extract in Advanced Osteosarcoma (OSCAT). Available online: https://clinicaltrials.gov/ct2/show/results/NCT00689195?term=Withania+somnifera&cond=cancer&draw=2&rank=3 (accessed on 3 March 2022).
- Bhat, J.; Damle, A.; Vaishnav, P.P.; Albers, R.; Joshi, M.; Banerjee, G. In vivo enhancement of natural killer cell activity through tea fortified with Ayurvedic herbs. Phytother. Res. 2010, 24, 129–135. [Google Scholar] [CrossRef]
- Ashwagandha: Effects on Stress, Inflammation and Immune Cell Activation. Available online: https://clinicaltrials.gov/ct2/show/NCT00817752?term=Withania+somnifera&cond=cancer&draw=2&rank=2 (accessed on 3 March 2022).
- Chengappa, K.N.R.; Brar, J.S.; Gannon, J.M.; Schlicht, P.J. Adjunctive Use of a Standardized Extract of Withania somnifera (Ashwagandha) to Treat Symptom Exacerbation in Schizophrenia: A Randomized, Double-Blind, Placebo-Controlled Study. J. Clin. Psychiatry 2018, 79, 22496. [Google Scholar] [CrossRef] [Green Version]
- Mandlik Ingawale, D.S.; Namdeo, A.G. Pharmacological evaluation of Ashwagandha highlighting its healthcare claims, safety, and toxicity aspects. J. Diet Suppl. 2021, 18, 183–226. [Google Scholar] [CrossRef]
- Khanal, P.; Chikhale, R.; Dey, Y.N.; Pasha, I.; Chand, S.; Gurav, N.; Ayyanar, M.; Patil, B.M.; Gurav, S. Withanolides from Withania somnifera as an immunity booster and their therapeutic options against COVID-19. J. Biomol. Struct. Dyn. 2021. [Google Scholar] [CrossRef]
- Chandran, U.; Patwardhan, B. Network ethnopharmacological evaluation of the immunomodulatory activity of Withania somnifera. J. Ethnopharmacol. 2017, 197, 250–256. [Google Scholar] [CrossRef]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; May, M.J.; Kopp, E.B. NF-kappa B and Rel proteins: Evolutionarily conserved mediators of immune responses. Annu Rev. Immunol. 1998, 16, 225–260. [Google Scholar] [CrossRef] [PubMed]
- Vanden Berghe, W.; Sabbe, L.; Kaileh, M.; Haegeman, G.; Heyninck, K. Molecular insight in the multifunctional activities of Withaferin A. Biochem. Pharmacol. 2012, 84, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Iuvone, T.; Esposito, G.; Capasso, F.; Izzo, A.A. Induction of nitric oxide synthase expression by Withania somnifera in macrophages. Life Sci. 2003, 72, 1617–1625. [Google Scholar] [CrossRef]
- Min, K.J.; Choi, K.; Kwon, T.K. Withaferin A down-regulates lipopolysaccharide-induced cyclooxygenase-2 expression and PGE2 production through the inhibition of STAT1/3 activation in microglial cells. Int. Immunopharmacol. 2011, 11, 1137–1142. [Google Scholar] [CrossRef]
- Singh, D.; Aggarwal, A.; Maurya, R.; Naik, S. Withania somnifera inhibits NF-kappaB and AP-1 transcription factors in human peripheral blood and synovial fluid mononuclear cells. Phytother. Res. 2007, 21, 905–913. [Google Scholar] [CrossRef]
- Russo, A.; Izzo, A.A.; Cardile, V.; Borrelli, F.; Vanella, A. Indian medicinal plants as antiradicals and DNA cleavage protectors. Phytomedicine 2001, 8, 125–132. [Google Scholar] [CrossRef]
- Davis, L.; Kuttan, G. Immunomodulatory activity of Withania somnifera. J. Ethnopharmacol. 2000, 71, 193–200. [Google Scholar] [CrossRef]
- Ziauddin, M.; Phansalkar, N.; Patki, P.; Diwanay, S.; Patwardhan, B. Studies on the immunomodulatory effects of Ashwagandha. J. Ethnopharmacol. 1996, 50, 69–76. [Google Scholar] [CrossRef]
- Oh, J.H.; Lee, T.J.; Park, J.W.; Kwon, T.K. Withaferin A inhibits iNOS expression and nitric oxide production by Akt inactivation and down-regulating LPS-induced activity of NF-kappaB in RAW 264.7 cells. Eur. J. Pharmacol. 2008, 599, 11–17. [Google Scholar] [CrossRef]
- Chauhan, D.S.; Dhasmana, A.; Laskar, P.; Prasad, R.; Jain, N.K.; Srivastava, R.; Jaggi, M.; Chauhan, S.C.; Yallapu, M.M. Nanotechnology synergized immunoengineering for cancer. Eur. J. Pharm. Biopharm. 2021, 163, 72–101. [Google Scholar] [CrossRef] [PubMed]
- Shetty, A.; Nagesh, P.K.B.; Setua, S.; Hafeez, B.B.; Jaggi, M.; Yallapu, M.M.; Chauhan, S.C. Novel Paclitaxel Nanoformulation Impairs De Novo Lipid Synthesis in Pancreatic Cancer Cells and Enhances Gemcitabine Efficacy. ACS Omega 2020, 5, 8982–8991. [Google Scholar] [CrossRef] [PubMed]
- Samanta, K.; Setua, S.; Kumari, S.; Jaggi, M.; Yallapu, M.M.; Chauhan, S.C. Gemcitabine Combination Nano Therapies for Pancreatic Cancer. Pharmaceutics 2019, 11, 574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massey, A.E.; Sikander, M.; Chauhan, N.; Kumari, S.; Setua, S.; Shetty, A.B.; Mandil, H.; Kashyap, V.K.; Khan, S.; Jaggi, M.; et al. Next-generation paclitaxel-nanoparticle formulation for pancreatic cancer treatment. Nanomedicine 2019, 20, 102027. [Google Scholar] [CrossRef]
- Kim, B.; Park, J.E.; Im, E.; Cho, Y.; Lee, J.; Lee, H.J.; Sim, D.Y.; Park, W.Y.; Shim, B.S.; Kim, S.H. Recent Advances in Nanotechnology with Nano-Phytochemicals: Molecular Mechanisms and Clinical Implications in Cancer Progression. Int. J. Mol. Sci. 2021, 22, 3571. [Google Scholar] [CrossRef]
- Salama, L.; Pastor, E.R.; Stone, T.; Mousa, S.A. Emerging Nanopharmaceuticals and Nanonutraceuticals in Cancer Management. Biomedicines 2020, 8, 347. [Google Scholar] [CrossRef]
- Granja, A.; Frias, I.; Neves, A.R.; Pinheiro, M.; Reis, S. Therapeutic Potential of Epigallocatechin Gallate Nanodelivery Systems. BioMed Res. Int. 2017, 2017, 5813793. [Google Scholar] [CrossRef]
- Khan, T.; Gurav, P. PhytoNanotechnology: Enhancing Delivery of Plant Based Anti-cancer Drugs. Front. Pharmacol. 2018, 8, 1002. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, S.; Tavares, A.J.; Dai, Q.; Ohta, S.; Audet, J.; Dvorak, H.F.; Chan, W.C.W. Analysis of nanoparticle delivery to tumours. Nat. Rev. Mater. 2016, 1, 16014. [Google Scholar] [CrossRef]
- Khan, S.; Setua, S.; Kumari, S.; Dan, N.; Massey, A.; Hafeez, B.B.; Yallapu, M.M.; Stiles, Z.E.; Alabkaa, A.; Yue, J.; et al. Superparamagnetic iron oxide nanoparticles of curcumin enhance gemcitabine therapeutic response in pancreatic cancer. Biomaterials 2019, 208, 83–97. [Google Scholar] [CrossRef]
- Nagesh, P.K.B.; Chowdhury, P.; Hatami, E.; Boya, V.K.N.; Kashyap, V.K.; Khan, S.; Hafeez, B.B.; Chauhan, S.C.; Jaggi, M.; Yallapu, M.M. miRNA-205 Nanoformulation Sensitizes Prostate Cancer Cells to Chemotherapy. Cancers 2018, 10, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanomedicine Market Is Estimated To Be Valued At $350.8 Billion By 2025: Grand View Research, Inc. Available online: https://www.grandviewresearch.com/press-release/global-nanomedicine-market (accessed on 3 March 2022).
- Anselmo, A.C.; Mitragotri, S. A Review of Clinical Translation of Inorganic Nanoparticles. AAPS J. 2015, 17, 1041–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Yan, L.; Gu, Z.; Zhao, Y. Strategies based on metal-based nanoparticles for hypoxic-tumor radiotherapy. Chem. Sci. 2019, 10, 6932–6943. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.; Dong, B.; Yu, J.; Hou, Y. Inherent multifunctional inorganic nanomaterials for imaging-guided cancer therapy. Nano Today 2019, 26, 108–122. [Google Scholar] [CrossRef]
- Hess, K.L.; Medintz, I.L.; Jewell, C.M. Designing inorganic nanomaterials for vaccines and immunotherapies. Nano Today 2019, 27, 73–98. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Zhang, X.; Gu, Z.; Zhao, Y. Recent advances in upconversion nanoparticles-based multifunctional nanocomposites for combined cancer therapy. Adv. Mater. 2015, 27, 7692–7712. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Hyeon, T. Applications of inorganic nanoparticles as therapeutic agents. Nanotechnology 2013, 25, 012001. [Google Scholar] [CrossRef]
- Erathodiyil, N.; Ying, J.Y. Functionalization of inorganic nanoparticles for bioimaging applications. Acc. Chem. Res. 2011, 44, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Na, H.B.; Song, I.C.; Hyeon, T. Inorganic nanoparticles for MRI contrast agents. Adv. Mater. 2009, 21, 2133–2148. [Google Scholar] [CrossRef]
- Liu, J.; Zheng, X.; Yan, L.; Zhou, L.; Tian, G.; Yin, W.; Wang, L.; Liu, Y.; Hu, Z.; Gu, Z. Bismuth sulfide nanorods as a precision nanomedicine for in vivo multimodal imaging-guided photothermal therapy of tumor. ACS Nano 2015, 9, 696–707. [Google Scholar] [CrossRef]
- Tian, G.; Gu, Z.; Zhou, L.; Yin, W.; Liu, X.; Yan, L.; Jin, S.; Ren, W.; Xing, G.; Li, S. Mn2+ dopant-controlled synthesis of NaYF4: Yb/Er upconversion nanoparticles for in vivo imaging and drug delivery. Adv. Mater. 2012, 24, 1226–1231. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Gu, Z.; Yan, L.; Yong, Y.; Yi, X.; Zhang, X.; Liu, J.; Wu, R.; Ge, C.; Chen, C.; et al. Poly(Vinylpyrollidone)- and Selenocysteine-Modified Bi(2) Se(3) Nanoparticles Enhance Radiotherapy Efficacy in Tumors and Promote Radioprotection in Normal Tissues. Adv. Mater. 2017, 29, 1701268. [Google Scholar] [CrossRef]
- Samia, A.C.; Chen, X.; Burda, C. Semiconductor quantum dots for photodynamic therapy. J. Am. Chem. Soc. 2003, 125, 15736–15737. [Google Scholar] [CrossRef] [PubMed]
- Son, S.J.; Bai, X.; Lee, S.B. Inorganic hollow nanoparticles and nanotubes in nanomedicine Part 1. Drug/gene delivery applications. Drug Discov. Today 2007, 12, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yan, L.; Wang, X.; Dong, X.; Zhou, R.; Gu, Z.; Zhao, Y. Tumor Microenvironment-Responsive Cu(2)(OH)PO(4) Nanocrystals for Selective and Controllable Radiosentization via the X-ray-Triggered Fenton-like Reaction. Nano Lett. 2019, 19, 1749–1757. [Google Scholar] [CrossRef]
- Gu, Z.; Zhu, S.; Yan, L.; Zhao, F.; Zhao, Y. Graphene-Based Smart Platforms for Combined Cancer Therapy. Adv. Mater. 2019, 31, e1800662. [Google Scholar] [CrossRef]
- Du, Z.; Zhang, X.; Guo, Z.; Xie, J.; Dong, X.; Zhu, S.; Du, J.; Gu, Z.; Zhao, Y. X-Ray-Controlled Generation of Peroxynitrite Based on Nanosized LiLuF(4): Ce(3+) Scintillators and their Applications for Radiosensitization. Adv. Mater. 2018, 30, e1804046. [Google Scholar] [CrossRef]
- Chen, H.; Gu, Z.; An, H.; Chen, C.; Chen, J.; Cui, R.; Chen, S.; Chen, W.; Chen, X.; Chen, X. Precise nanomedicine for intelligent therapy of cancer. Sci. China Chem. 2018, 61, 1503–1552. [Google Scholar] [CrossRef]
- Bobo, D.; Robinson, K.J.; Islam, J.; Thurecht, K.J.; Corrie, S.R. Nanoparticle-based medicines: A review of FDA-approved materials and clinical trials to date. Pharm. Res. 2016, 33, 2373–2387. [Google Scholar] [CrossRef]
- Lim, Z.-Z.J.; Li, J.-E.J.; Ng, C.-T.; Yung, L.-Y.L.; Bay, B.-H. Gold nanoparticles in cancer therapy. Acta Pharmacol. Sin. 2011, 32, 983–990. [Google Scholar] [CrossRef] [Green Version]
- Muddineti, O.S.; Ghosh, B.; Biswas, S. Current trends in using polymer coated gold nanoparticles for cancer therapy. Int. J. Pharm. 2015, 484, 252–267. [Google Scholar] [CrossRef] [PubMed]
- Tabassam, Q.; Mehmood, T.; Raza, A.R.; Ullah, A.; Saeed, F.; Anjum, F.M. Synthesis, Characterization and Anti-Cancer Therapeutic Potential of Withanolide-A with 20nm sAuNPs Conjugates Against SKBR3 Breast Cancer Cell Line. Int. J. Nanomed. 2020, 15, 6649–6658. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Naydenov, N.G.; Dozmorov, M.G.; Koblinski, J.E.; Ivanov, A.I. Anillin regulates breast cancer cell migration, growth, and metastasis by non-canonical mechanisms involving control of cell stemness and differentiation. Breast Cancer Res. 2020, 22, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devanand Venkatasubbu, G.; Ramasamy, S.; Ramakrishnan, V.; Kumar, J. Folate targeted PEGylated titanium dioxide nanoparticles as a nanocarrier for targeted paclitaxel drug delivery. Adv. Powder Technol. 2013, 24, 947–954. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Kobayashi, H.; Narita, T.; Kanehira, K.; Sonezaki, S.; Kudo, N.; Kubota, Y.; Terasaka, S.; Houkin, K. Sonodynamic therapy using water-dispersed TiO2-polyethylene glycol compound on glioma cells: Comparison of cytotoxic mechanism with photodynamic therapy. Ultrason. Sonochem. 2011, 18, 1197–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Hashimoto, K.; Fujishima, A.; Chikuni, M.; Kojima, E.; Kitamura, A.; Shimohigoshi, M.; Watanabe, T. Light-induced amphiphilic surfaces. Nature 1997, 388, 431–432. [Google Scholar] [CrossRef]
- Yin, Z.F.; Wu, L.; Yang, H.G.; Su, Y.H. Recent progress in biomedical applications of titanium dioxide. Phys. Chem. Chem. Phys. 2013, 15, 4844–4858. [Google Scholar] [CrossRef]
- Xu, P.; Wang, R.; Ouyang, J.; Chen, B. A new strategy for TiO2 whiskers mediated multi-mode cancer treatment. Nanoscale Res. Lett. 2015, 10, 94. [Google Scholar] [CrossRef] [Green Version]
- Liu, E.; Zhou, Y.; Liu, Z.; Li, J.; Zhang, D.; Chen, J.; Cai, Z. Cisplatin loaded hyaluronic acid modified TiO 2 nanoparticles for neoadjuvant chemotherapy of ovarian cancer. J. Nanomater. 2015, 16, 275. [Google Scholar]
- Kulshrestha, S.; Qayyum, S.; Khan, A.U. Antibiofilm efficacy of green synthesized graphene oxide-silver nanocomposite using Lagerstroemia speciosa floral extract: A comparative study on inhibition of gram-positive and gram-negative biofilms. Microb. Pathog. 2017, 103, 167–177. [Google Scholar] [CrossRef]
- Rajkumari, J.; Magdalane, C.M.; Siddhardha, B.; Madhavan, J.; Ramalingam, G.; Al-Dhabi, N.A.; Arasu, M.V.; Ghilan, A.K.M.; Duraipandiayan, V.; Kaviyarasu, K. Synthesis of titanium oxide nanoparticles using Aloe barbadensis mill and evaluation of its antibiofilm potential against Pseudomonas aeruginosa PAO1. J. Photochem. Photobiol. B 2019, 201, 111667. [Google Scholar] [CrossRef] [PubMed]
- Madadi, Z.; Bagheri Lotfabad, T. Aqueous Extract of Acanthophyllum laxiusculum Roots as a Renewable Resource for Green synthesis of nano-sized titanium dioxide using Sol-gel Method. Adv. Ceram. Prog. 2016, 2, 26–31. [Google Scholar] [CrossRef]
- Kashale, A.A.; Gattu, K.P.; Ghule, K.; Ingole, V.H.; Dhanayat, S.; Sharma, R.; Chang, J.-Y.; Ghule, A.V. Biomediated green synthesis of TiO2 nanoparticles for lithium ion battery application. Compos. B Eng. 2016, 99, 297–304. [Google Scholar] [CrossRef]
- Bischoff, B.L.; Anderson, M.A. Peptization Process in the Sol-Gel Preparation of Porous Anatase (TiO2). Chem. Mater. 1995, 7, 1772–1778. [Google Scholar] [CrossRef]
- Reinke, M.; Ponomarev, E.; Kuzminykh, Y.; Hoffmann, P. Combinatorial Characterization of TiO2 Chemical Vapor Deposition Utilizing Titanium Isopropoxide. ACS Comb. Sci. 2015, 17, 413–420. [Google Scholar] [CrossRef]
- Sakai, H.; Kawahara, H.; Shimazaki, M.; Abe, M. Preparation of Ultrafine Titanium Dioxide Particles Using Hydrolysis and Condensation Reactions in the Inner Aqueous Phase of Reversed Micelles: Effect of Alcohol Addition. Langmuir 1998, 14, 2208–2212. [Google Scholar] [CrossRef]
- Chen, H.; Wang, Y.; Dong, S. An Effective Hydrothermal Route for the Synthesis of Multiple PDDA-Protected Noble-Metal Nanostructures. Inorg. Chem. 2007, 46, 10587–10593. [Google Scholar] [CrossRef]
- Leshuk, T.; Parviz, R.; Everett, P.; Krishnakumar, H.; Varin, R.A.; Gu, F. Photocatalytic activity of hydrogenated TiO2. ACS Appl. Mater. Interfaces 2013, 5, 1892–1895. [Google Scholar] [CrossRef]
- Schneider, J.; Matsuoka, M.; Takeuchi, M.; Zhang, J.; Horiuchi, Y.; Anpo, M.; Bahnemann, D.W. Understanding TiO2 photocatalysis: Mechanisms and materials. Chem. Rev. 2014, 114, 9919–9986. [Google Scholar] [CrossRef]
- Anderson, C.; Bard, A.J. Improved Photocatalytic Activity and Characterization of Mixed TiO2/SiO2 and TiO2/Al2O3 Materials. J. Phys. Chem. 1997, 101, 2611–2616. [Google Scholar] [CrossRef]
- Benavides, J.A.; Trudeau, C.P.; Gerlein, L.F.; Cloutier, S.G. Laser Selective Photoactivation of Amorphous TiO2 Films to Anatase and/or Rutile Crystalline Phases. ACS Appl. Energy Mater. 2018, 1, 3607–3613. [Google Scholar] [CrossRef]
- Carta, D.; Salaoru, I.; Khiat, A.; Regoutz, A.; Mitterbauer, C.; Harrison, N.M.; Prodromakis, T. Investigation of the Switching Mechanism in TiO2-Based RRAM: A Two-Dimensional EDX Approach. ACS Appl. Mater. Interfaces 2016, 8, 19605–19611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Mora-Seró, I.; De Angelis, F.; Bisquert, J.; Wang, P. Titanium Dioxide Nanomaterials for Photovoltaic Applications. Chem. Rev. 2014, 114, 10095–10130. [Google Scholar] [CrossRef] [PubMed]
- Maheswari, P.; Harish, S.; Navaneethan, M.; Muthamizhchelvan, C.; Ponnusamy, S.; Hayakawa, Y. Bio-modified TiO(2) nanoparticles with Withania somnifera, Eclipta prostrata and Glycyrrhiza glabra for anticancer and antibacterial applications. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 108, 110457. [Google Scholar] [CrossRef] [PubMed]
- Al-Shabib, N.A.; Husain, F.M.; Qais, F.A.; Ahmad, N.; Khan, A.; Alyousef, A.A.; Arshad, M.; Noor, S.; Khan, J.M.; Alam, P.; et al. Phyto-Mediated Synthesis of Porous Titanium Dioxide Nanoparticles From Withania somnifera Root Extract: Broad-Spectrum Attenuation of Biofilm and Cytotoxic Properties Against HepG2 Cell Lines. Front. Microbiol. 2020, 11, 1680. [Google Scholar] [CrossRef]
- Xu, L.; Wang, Y.-Y.; Huang, J.; Chen, C.-Y.; Wang, Z.-X.; Xie, H. Silver nanoparticles: Synthesis, medical applications and biosafety. Theranostics 2020, 10, 8996–9031. [Google Scholar] [CrossRef]
- Abbasi, E.; Milani, M.; Fekri Aval, S.; Kouhi, M.; Akbarzadeh, A.; Tayefi Nasrabadi, H.; Nikasa, P.; Joo, S.W.; Hanifehpour, Y.; Nejati-Koshki, K.; et al. Silver nanoparticles: Synthesis methods, bio-applications and properties. Crit. Rev. Microbiol. 2016, 42, 173–180. [Google Scholar] [CrossRef]
- Mousavi, S.M.; Hashemi, S.A.; Ghasemi, Y.; Atapour, A.; Amani, A.M.; Savar Dashtaki, A.; Babapoor, A.; Arjmand, O. Green synthesis of silver nanoparticles toward bio and medical applications: Review study. Artif. Cells Nanomed. Biotechnol. 2018, 46, S855–S872. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Amatya, R.; Hwang, S.; Lee, S.; Min, K.A.; Shin, M.C. BSA-Silver Nanoparticles: A Potential Multimodal Therapeutics for Conventional and Photothermal Treatment of Skin Cancer. Pharmaceutics 2021, 13, 575. [Google Scholar] [CrossRef]
- Tripathi, D.; Modi, A.; Narayan, G.; Rai, S.P. Green and cost effective synthesis of silver nanoparticles from endangered medicinal plant Withania coagulans and their potential biomedical properties. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 100, 152–164. [Google Scholar] [CrossRef]
- Gaurav, I.; Singh, T.; Thakur, A.; Kumar, G.; Rathee, P.; Kumari, P.; Sweta, K. Synthesis, In-Vitro and In-Silico Evaluation of Silver Nanoparticles with Root Extract of Withania somnifera for Antibacterial Activity via Binding of Penicillin-Binding Protein-4. Curr. Pharm. Biotechnol. 2020, 21, 1674–1687. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, S.; Sood, H.; Saxena, S.; Chaurasia, O.P. Green synthesis of silver nanoparticles using Rhodiola imbricata and Withania somnifera root extract and their potential catalytic, antioxidant, cytotoxic and growth-promoting activities. Bioprocess Biosyst. Eng. 2022, 45, 365–380. [Google Scholar] [CrossRef] [PubMed]
- Bisht, G.; Rayamajhi, S. ZnO Nanoparticles: A Promising Anticancer Agent. Nanobiomedicine 2016, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Nayak, T.R.; Hong, H.; Cai, W. Biomedical applications of zinc oxide nanomaterials. Curr. Mol. Med. 2013, 13, 1633–1645. [Google Scholar] [CrossRef] [Green Version]
- Kołodziejczak-Radzimska, A.; Jesionowski, T. Zinc Oxide-From Synthesis to Application: A Review. Materials 2014, 7, 2833–2881. [Google Scholar] [CrossRef] [Green Version]
- Vasuki, K.; Manimekalai, R. NIR light active ternary modified ZnO nanocomposites for combined cancer therapy. Heliyon 2019, 5, e02729. [Google Scholar] [CrossRef]
- Kumar, J.; Mitra, M.D.; Hussain, A.; Kaul, G. Exploration of immunomodulatory and protective effect of Withania somnifera on trace metal oxide (zinc oxide nanoparticles) induced toxicity in Balb/c mice. Mol. Biol. Rep. 2019, 46, 2447–2459. [Google Scholar] [CrossRef]
- Ventola, C.L. Progress in nanomedicine: Approved and investigational nanodrugs. Pharm. Ther. 2017, 42, 742. [Google Scholar]
- Ventola, C.L. The nanomedicine revolution: Part 1: Emerging concepts. Pharm. Ther. 2012, 37, 512. [Google Scholar]
- Fenske, D.B.; Cullis, P.R. Liposomal nanomedicines. Expert. Opin. Drug Deliv. 2008, 5, 25–44. [Google Scholar] [CrossRef]
- Min, Y.; Caster, J.M.; Eblan, M.J.; Wang, A.Z. Clinical translation of nanomedicine. Chem. Rev. 2015, 115, 11147–11190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Yan, L.; Wang, X.; Zhu, S.; Chen, C.; Gu, Z.; Zhao, Y. Progress, challenges, and future of nanomedicine. Nano Today 2020, 35, 101008. [Google Scholar] [CrossRef]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S.Y.; Sood, A.K.; Hua, S. Advances and challenges of liposome assisted drug delivery. Front. Pharmacol. 2015, 6, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Szoka, F. Steric stabilization of fusogenic liposomes by a low-pH sensitive PEG− diortho ester− lipid conjugate. Bioconjugate Chem. 2001, 12, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Bibi, S.; Lattmann, E.; Mohammed, A.R.; Perrie, Y. Trigger release liposome systems: Local and remote controlled delivery? J. Microencapsul. 2012, 29, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Woodle, M.C.; Lasic, D.D. Sterically stabilized liposomes. Biochim. Biophys. Acta (BBA)-Rev. Biomembr. 1992, 1113, 171–199. [Google Scholar] [CrossRef]
- Allen, T.M.; Hansen, C.; Rutledge, J. Liposomes with prolonged circulation times: Factors affecting uptake by reticuloendothelial and other tissues. Biochim. Biophys. Acta Biomembr. 1989, 981, 27–35. [Google Scholar] [CrossRef]
- Hua, S.; Wu, S.Y. The use of lipid-based nanocarriers for targeted pain therapies. Front. Pharmacol. 2013, 4, 143. [Google Scholar] [CrossRef] [Green Version]
- Kraft, J.C.; Freeling, J.P.; Wang, Z.; Ho, R.J. Emerging research and clinical development trends of liposome and lipid nanoparticle drug delivery systems. J. Pharm. Sci. 2014, 103, 29–52. [Google Scholar] [CrossRef] [Green Version]
- Bulbake, U.; Doppalapudi, S.; Kommineni, N.; Khan, W. Liposomal formulations in clinical use: An updated review. Pharmaceutics 2017, 9, 12. [Google Scholar] [CrossRef]
- Liu, D.; He, C.; Wang, A.Z.; Lin, W. Application of liposomal technologies for delivery of platinum analogs in oncology. Int. J. Nanomed. 2013, 8, 3309. [Google Scholar]
- Ko, A.H.; Tempero, M.A.; Shan, Y.S.; Su, W.C.; Lin, Y.L.; Dito, E.; Ong, A.; Wang, Y.W.; Yeh, C.G.; Chen, L.T. A multinational phase 2 study of nanoliposomal irinotecan sucrosofate (PEP02, MM-398) for patients with gemcitabine-refractory metastatic pancreatic cancer. Br. J. Cancer. 2013, 109, 920–925. [Google Scholar] [CrossRef] [PubMed]
- Koudelka, Š.; Turánek, J. Liposomal paclitaxel formulations. J. Control. Release 2012, 163, 322–334. [Google Scholar] [CrossRef] [PubMed]
- Deeken, J.F.; Slack, R.; Weiss, G.J.; Ramanathan, R.K.; Pishvaian, M.J.; Hwang, J.; Lewandowski, K.; Subramaniam, D.; He, A.R.; Cotarla, I. A phase I study of liposomal-encapsulated docetaxel (LE-DT) in patients with advanced solid tumor malignancies. Cancer Chemother. Pharmacol. 2013, 71, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Belfiore, L.; Saunders, D.N.; Ranson, M.; Thurecht, K.J.; Storm, G.; Vine, K.L. Towards clinical translation of ligand-functionalized liposomes in targeted cancer therapy: Challenges and opportunities. J. Control. Release 2018, 277, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapra, P.; Tyagi, P.; Allen, T.M. Ligand-targeted liposomes for cancer treatment. Curr. Drug Deliv. 2005, 2, 369–381. [Google Scholar] [CrossRef]
- Cheng, C.-C.; Chang, F.-C.; Kao, W.-Y.; Hwang, S.-M.; Liao, L.-C.; Chang, Y.-J.; Liang, M.-C.; Chen, J.-K.; Lee, D.-J. Highly efficient drug delivery systems based on functional supramolecular polymers: In vitro evaluation. Acta Biomater. 2016, 33, 194–202. [Google Scholar] [CrossRef]
- Hassan, S.; Prakash, G.; Ozturk, A.B.; Saghazadeh, S.; Sohail, M.F.; Seo, J.; Dokmeci, M.R.; Zhang, Y.S.; Khademhosseini, A. Evolution and clinical translation of drug delivery nanomaterials. Nano Today 2017, 15, 91–106. [Google Scholar] [CrossRef]
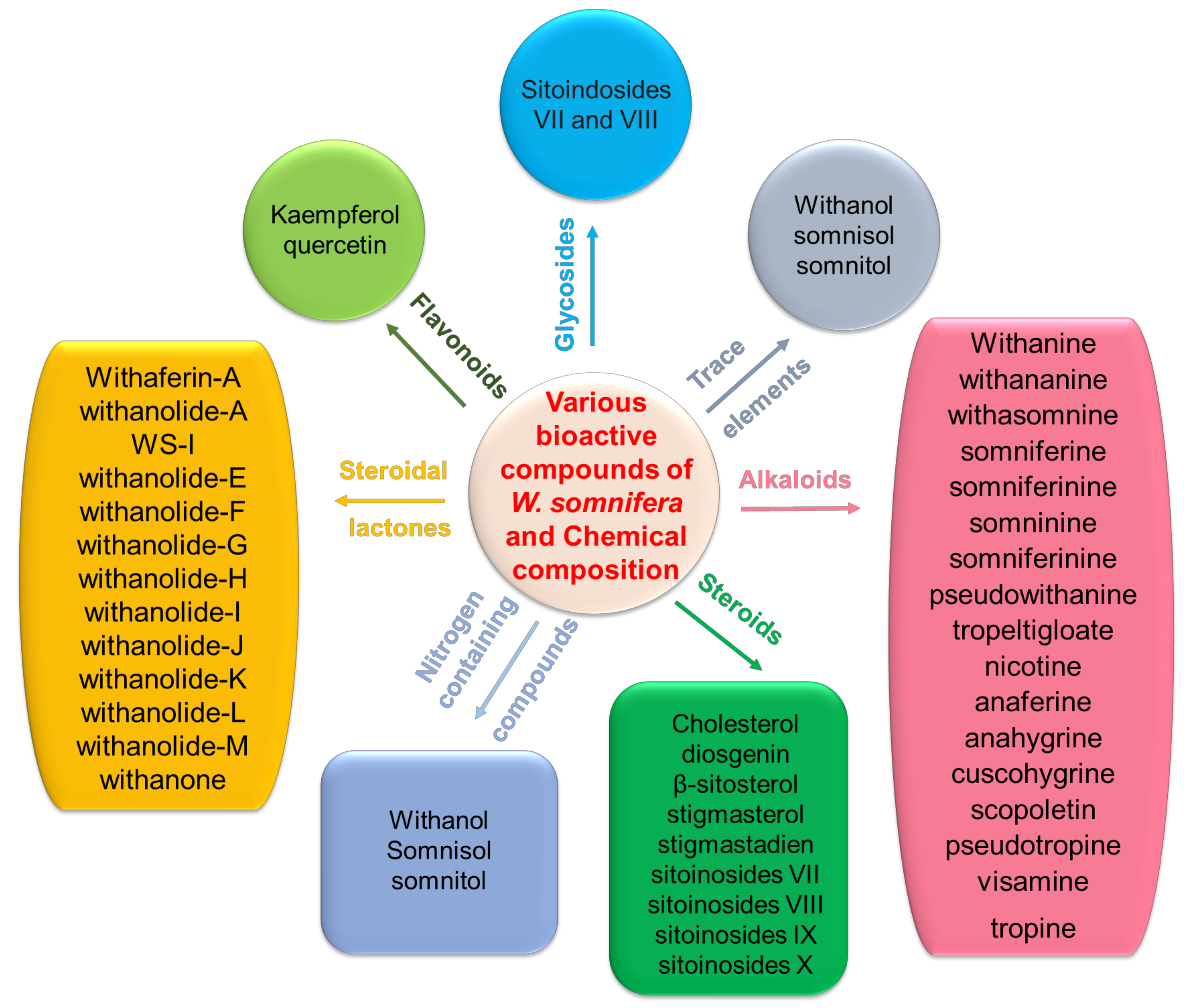
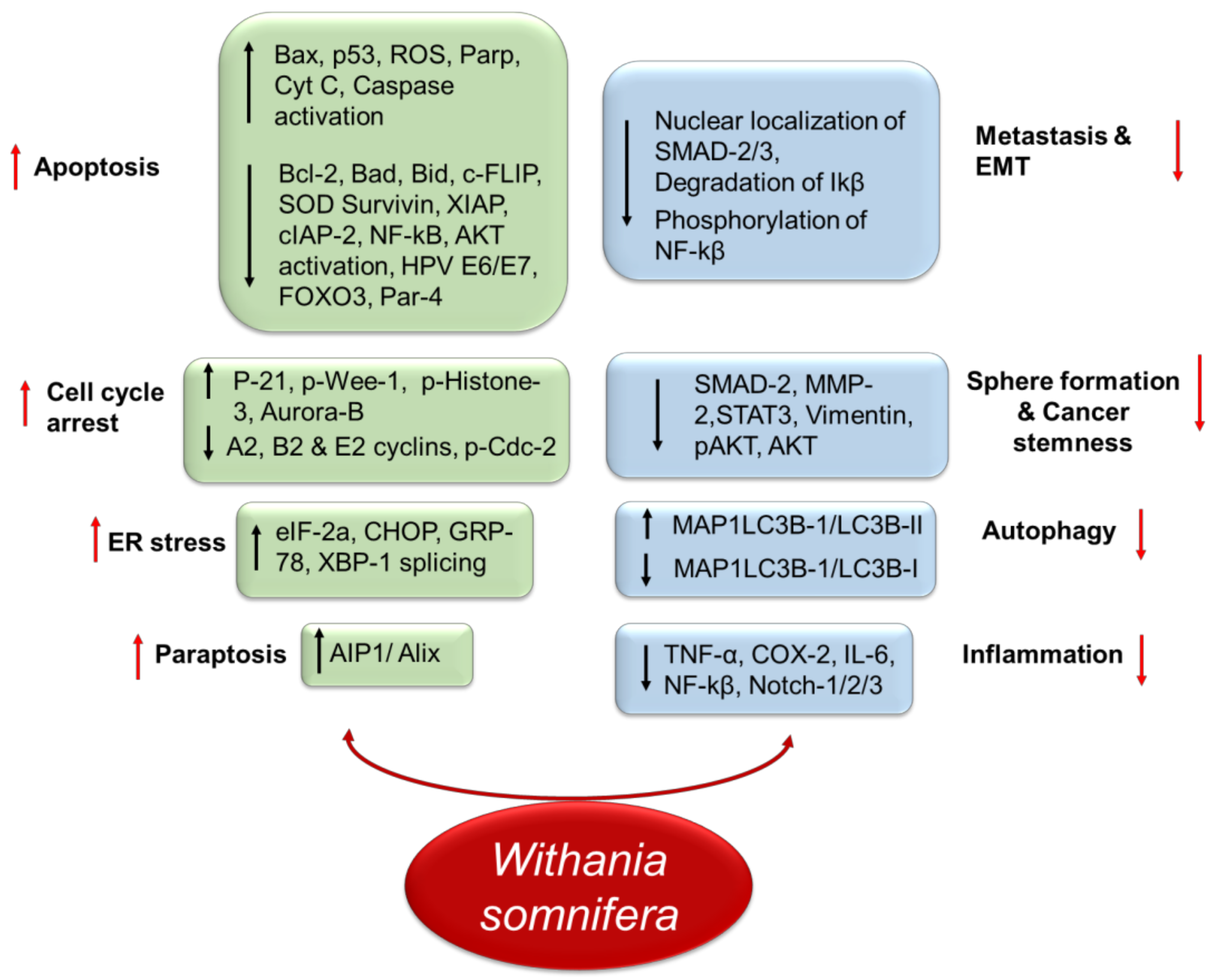
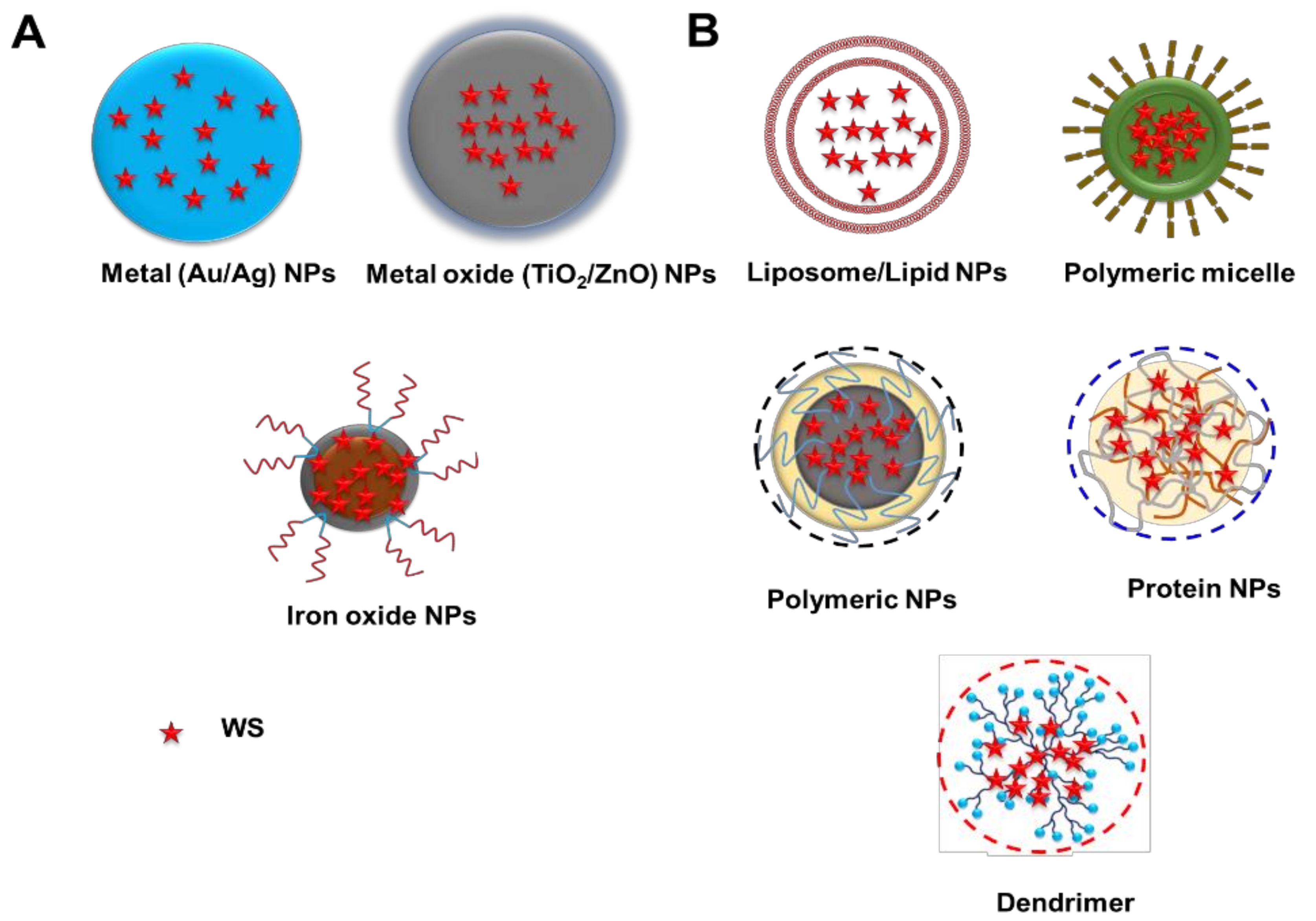
| Conditions | Used Alone or in Combination | ClinicalTrials.gov Identifier | No. of Enrolled Patients | Outcome | Ref. |
|---|---|---|---|---|---|
| Breast Cancer | Root extract | NA | 100 | Improvement of quality of life and chemotherapy induced fatigue | [54] |
| Advanced Osteosarcoma | CUR formulation and Ashwagandha Extract | NCT00689195 | 24 | Detailed is not available | [158] |
| Stress and Anxiety | Root extract | NA | 64 | Reduced stress and Anxiety improve self-assessed quality of life | [32] |
| NK Cell Activity | Polyherbal formulation | NA | 32 | Enhancement of NK cell activity | [159] |
| Stress, Inflammation, and Immune Cell Activation | 3 mL of Ashwagandha for 5 days | NCT00817752 | 25 | Detailed is not available | [160] |
| Schizophrenia | WS extract | NCT01793935 | 68 | Significant benefit for people with schizophrenia exacerbation | [161] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kashyap, V.K.; Peasah-Darkwah, G.; Dhasmana, A.; Jaggi, M.; Yallapu, M.M.; Chauhan, S.C. Withania somnifera: Progress towards a Pharmaceutical Agent for Immunomodulation and Cancer Therapeutics. Pharmaceutics 2022, 14, 611. https://doi.org/10.3390/pharmaceutics14030611
Kashyap VK, Peasah-Darkwah G, Dhasmana A, Jaggi M, Yallapu MM, Chauhan SC. Withania somnifera: Progress towards a Pharmaceutical Agent for Immunomodulation and Cancer Therapeutics. Pharmaceutics. 2022; 14(3):611. https://doi.org/10.3390/pharmaceutics14030611
Chicago/Turabian StyleKashyap, Vivek K., Godwin Peasah-Darkwah, Anupam Dhasmana, Meena Jaggi, Murali M. Yallapu, and Subhash C. Chauhan. 2022. "Withania somnifera: Progress towards a Pharmaceutical Agent for Immunomodulation and Cancer Therapeutics" Pharmaceutics 14, no. 3: 611. https://doi.org/10.3390/pharmaceutics14030611
APA StyleKashyap, V. K., Peasah-Darkwah, G., Dhasmana, A., Jaggi, M., Yallapu, M. M., & Chauhan, S. C. (2022). Withania somnifera: Progress towards a Pharmaceutical Agent for Immunomodulation and Cancer Therapeutics. Pharmaceutics, 14(3), 611. https://doi.org/10.3390/pharmaceutics14030611