
Melatonin Mitigates Cisplatin-Induced Ovarian Dysfunction via Altering Steroidogenesis, Inflammation, Apoptosis, Oxidative Stress, and PTEN/PI3K/Akt/mTOR/AMPK Signaling Pathway in Female Rats
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animals
2.3. Experimental Design and Sample Collection
2.4. Biochemical Analysis
2.5. Light Microscopic Analysis
2.6. Immunohistochemical Analysis
2.7. Morphometric Analysis
2.8. Real-Time Quantitative RT-PCR (qRT-PCR) Analysis
2.9. Statistical Analysis
3. Results
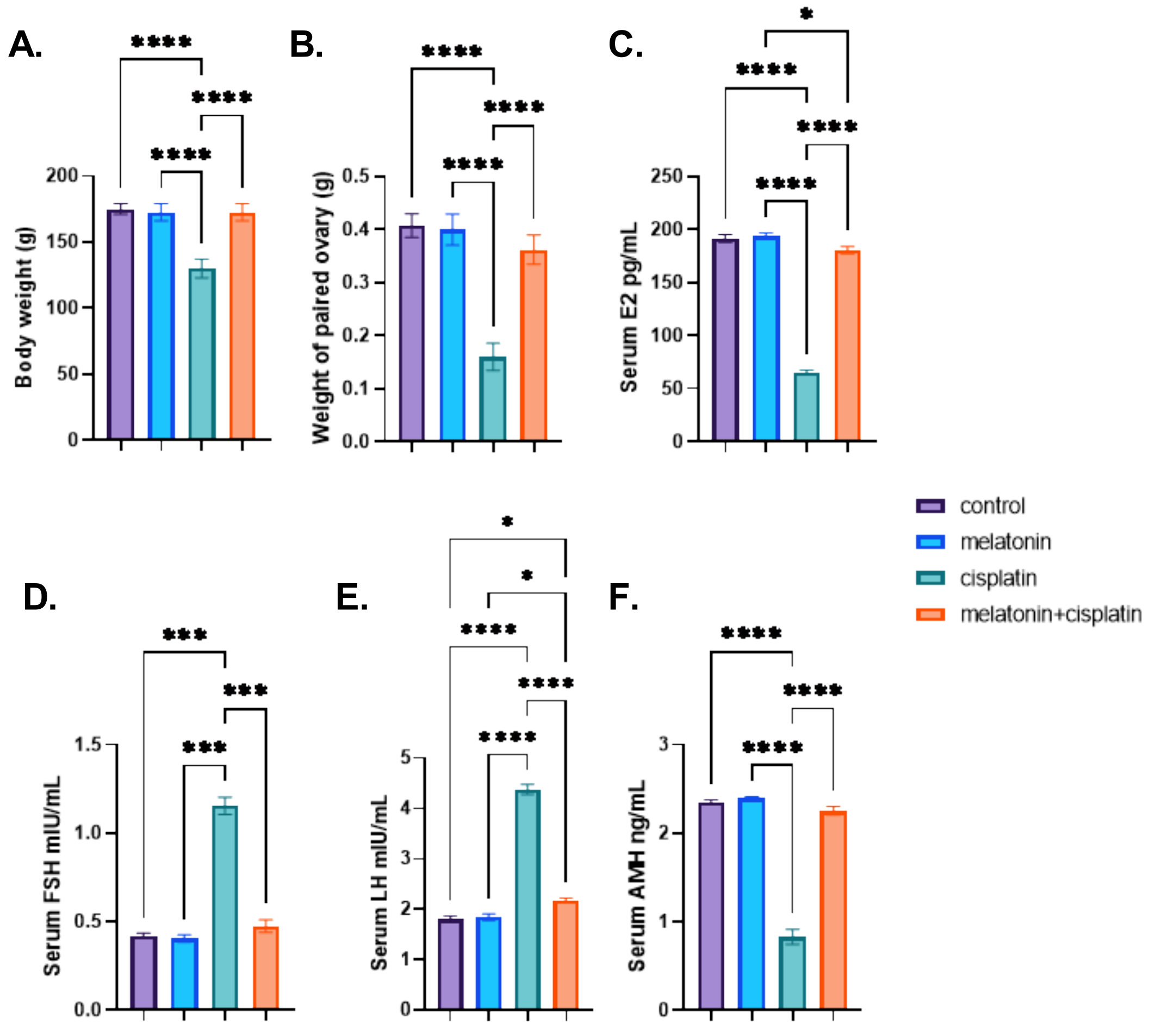
3.1. Body Weight, the Weight of Paired Ovary, and Serum Hormones
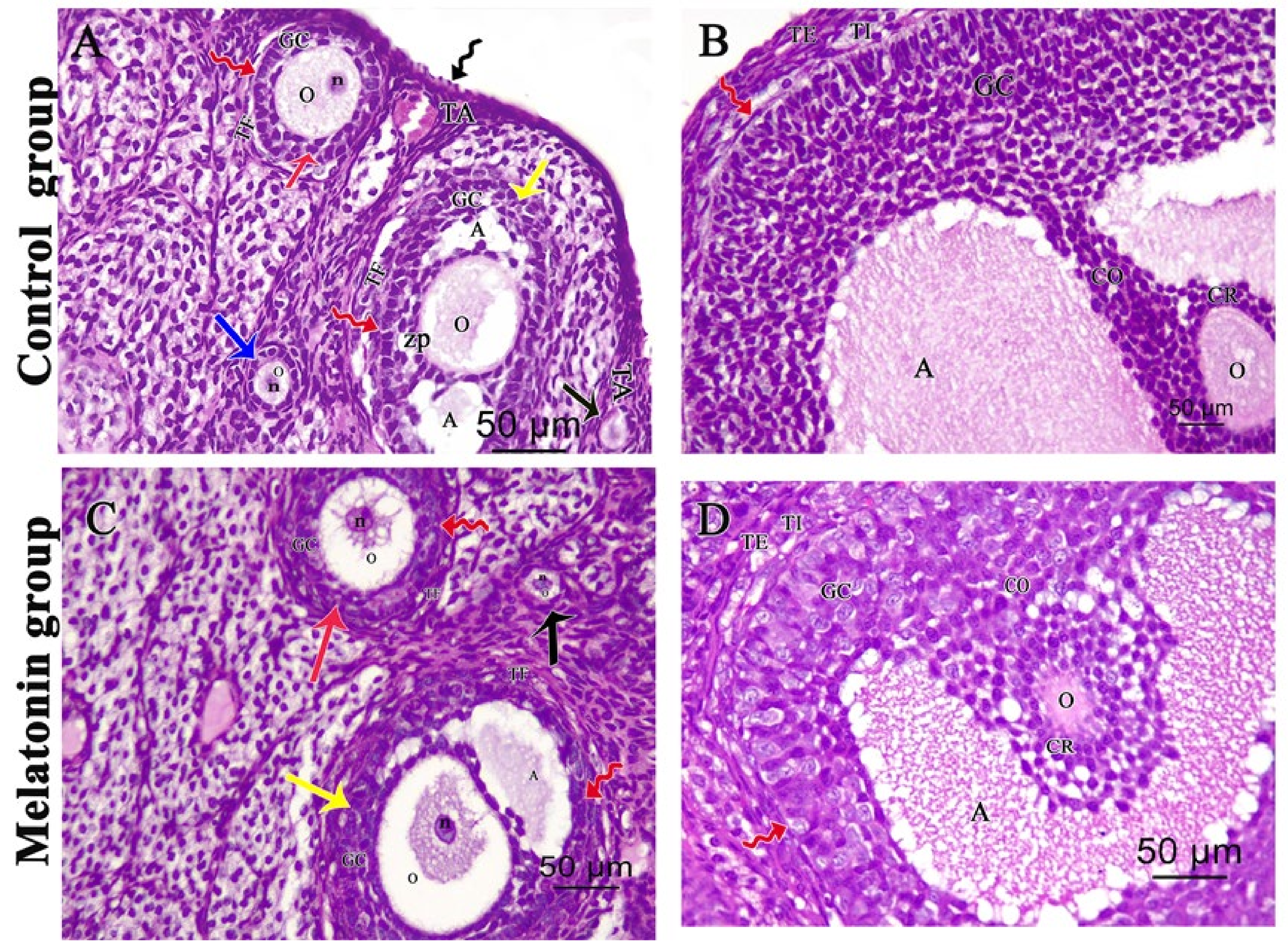
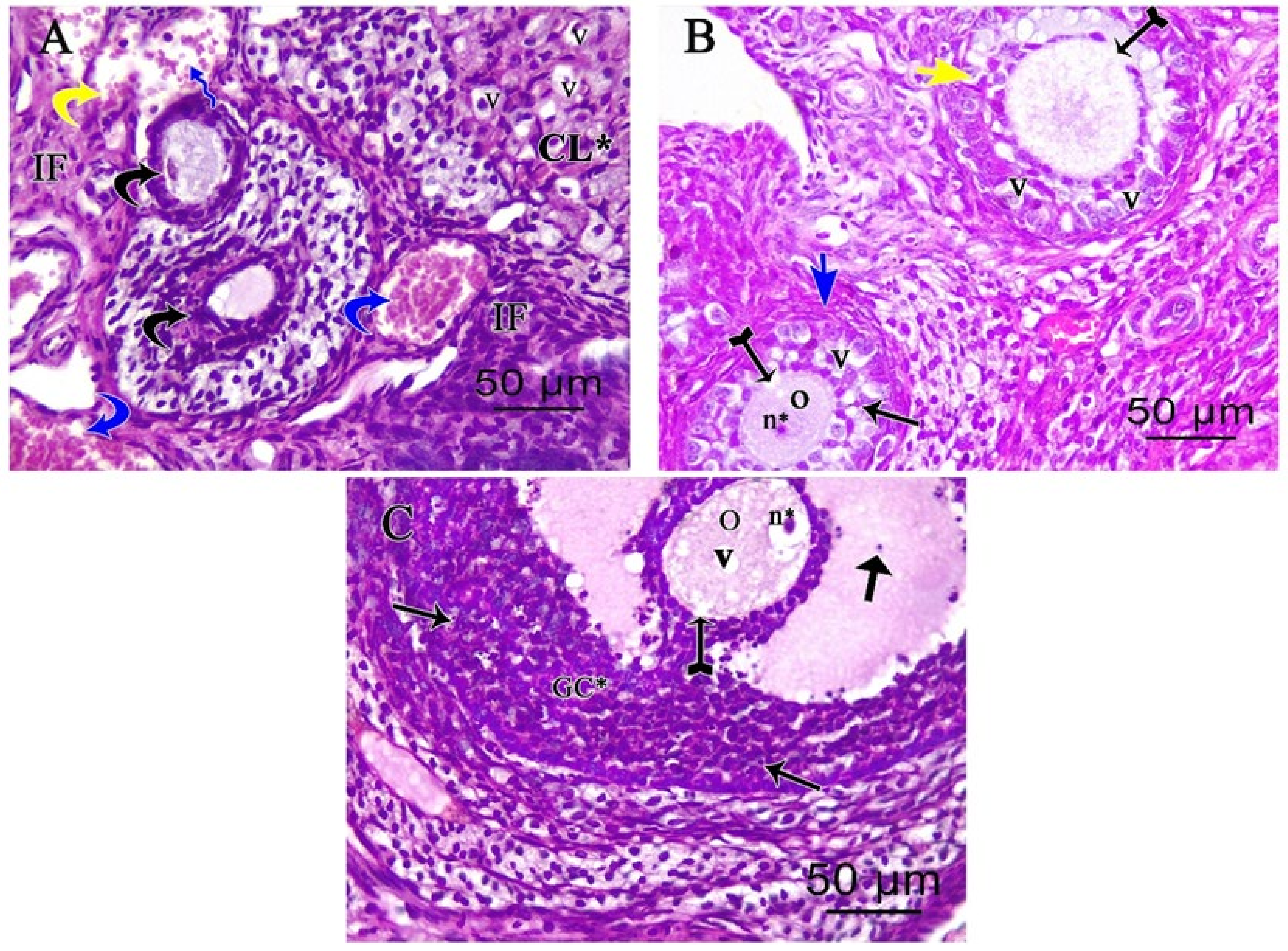
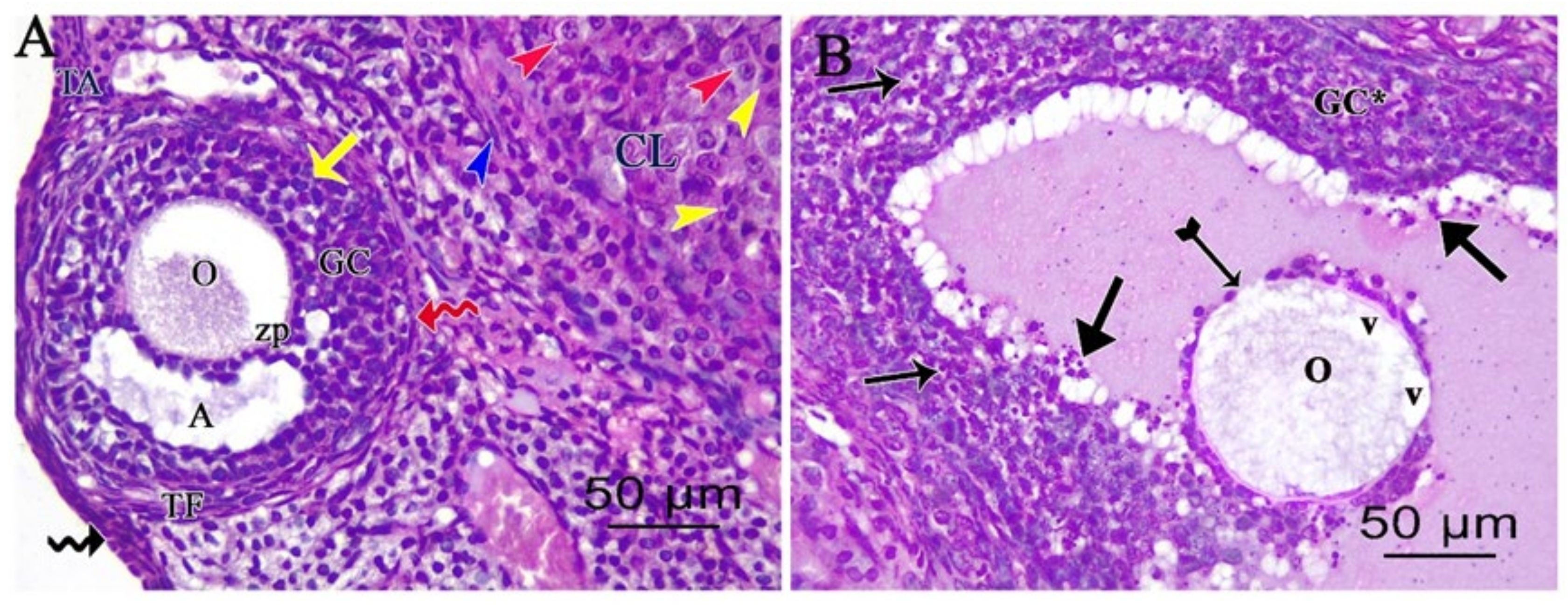
3.2. Light Microscopic Analysis of the Ovary
3.2.1. Ovarian Hematoxylin and Eosin (H&E)
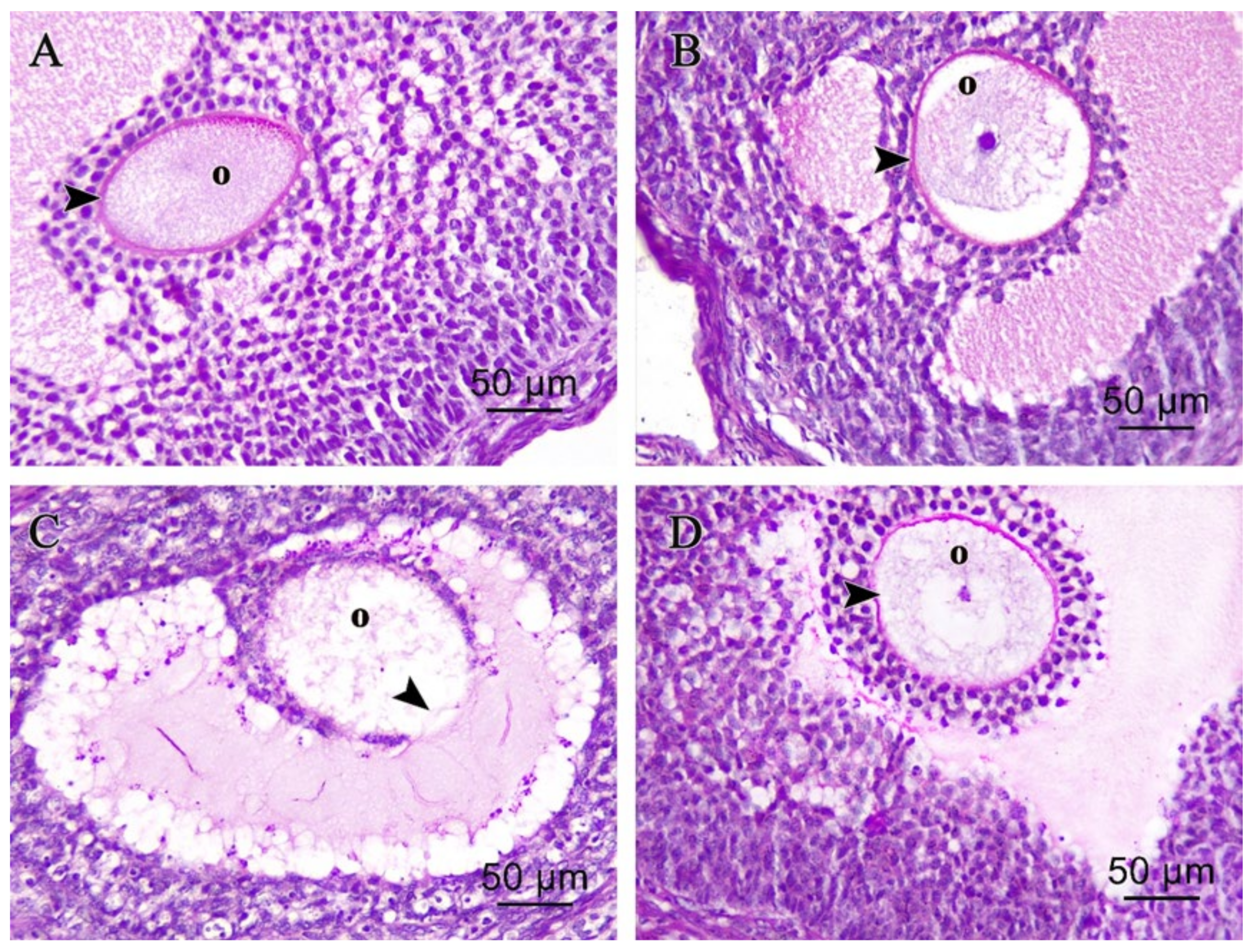
3.2.2. Ovarian Periodic Acid-Schiff Reagent (PAS)
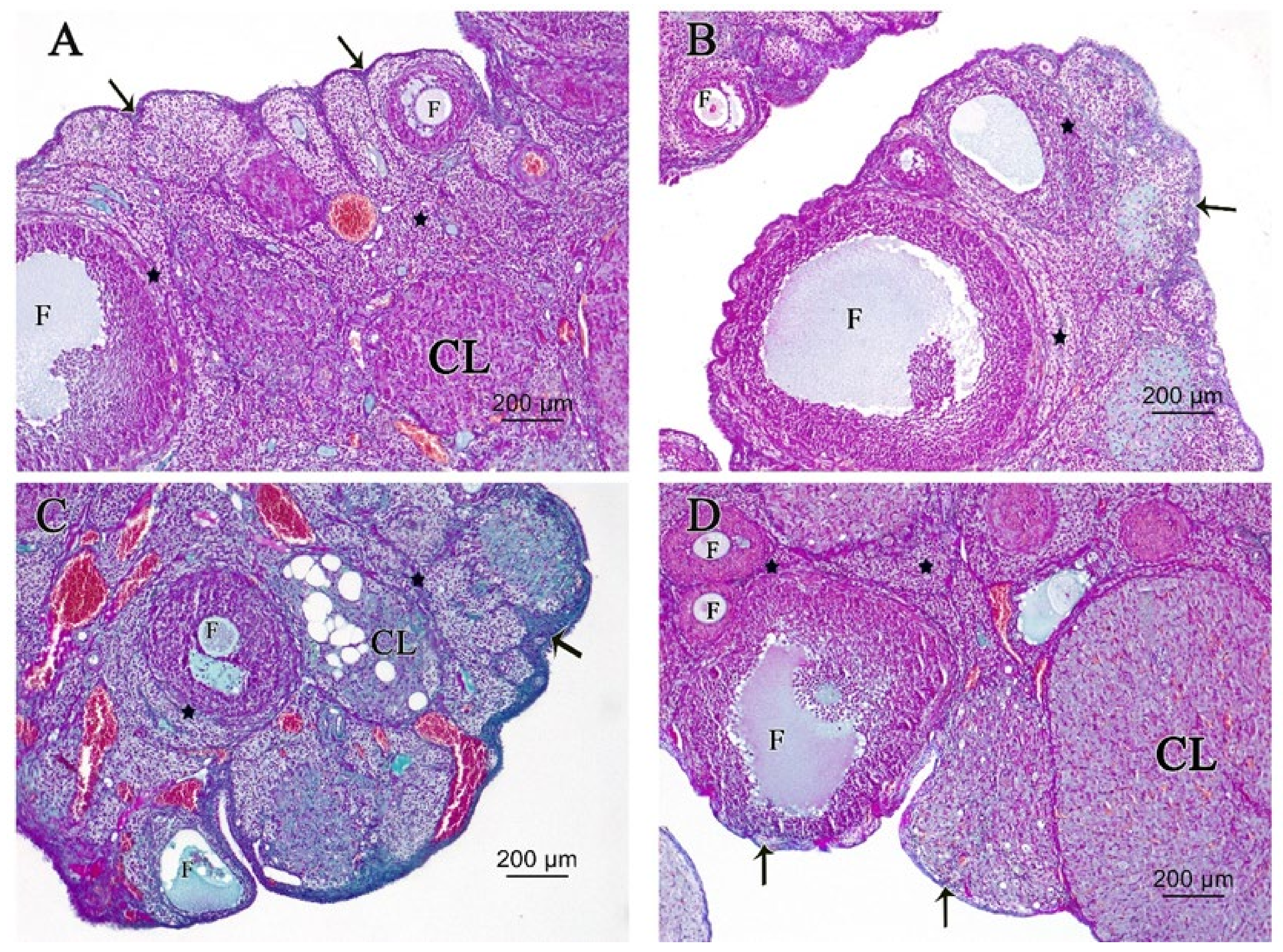
3.2.3. Ovarian Masson’s Trichrome (MT) Stain
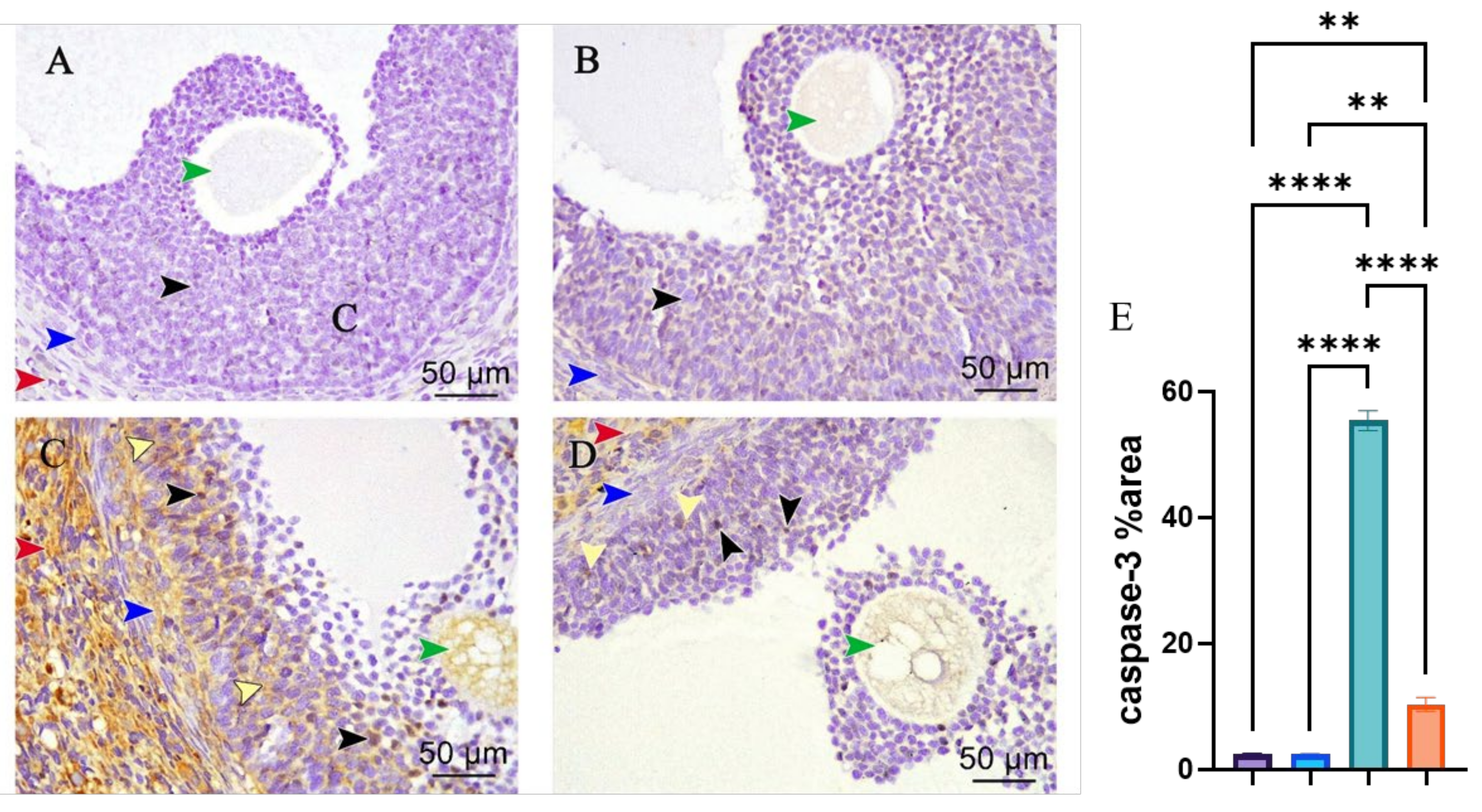
3.3. Caspase-3 Immunohistochemical Staining
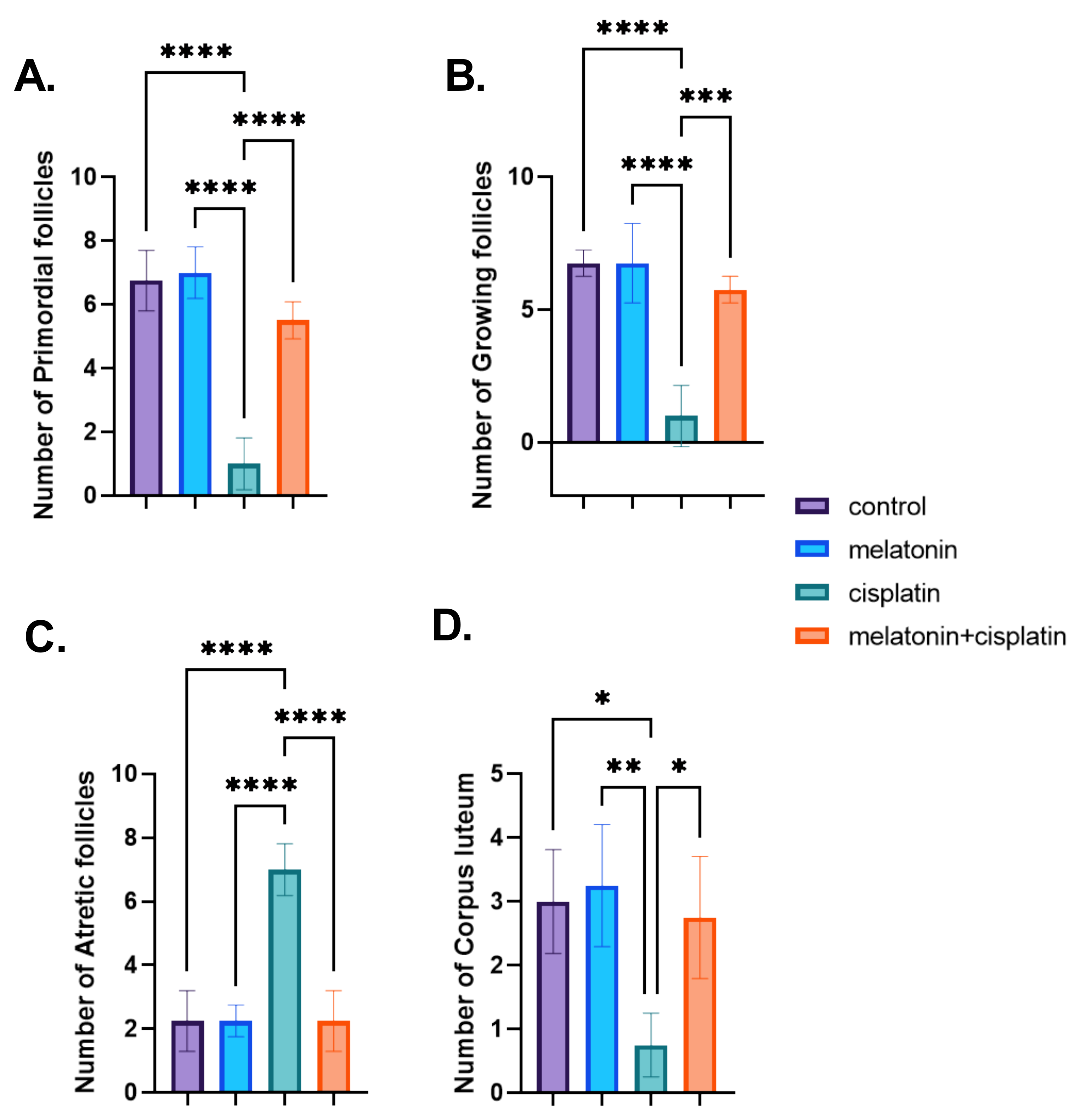
3.4. Morphometric Studies; Ovarian Follicles Counting
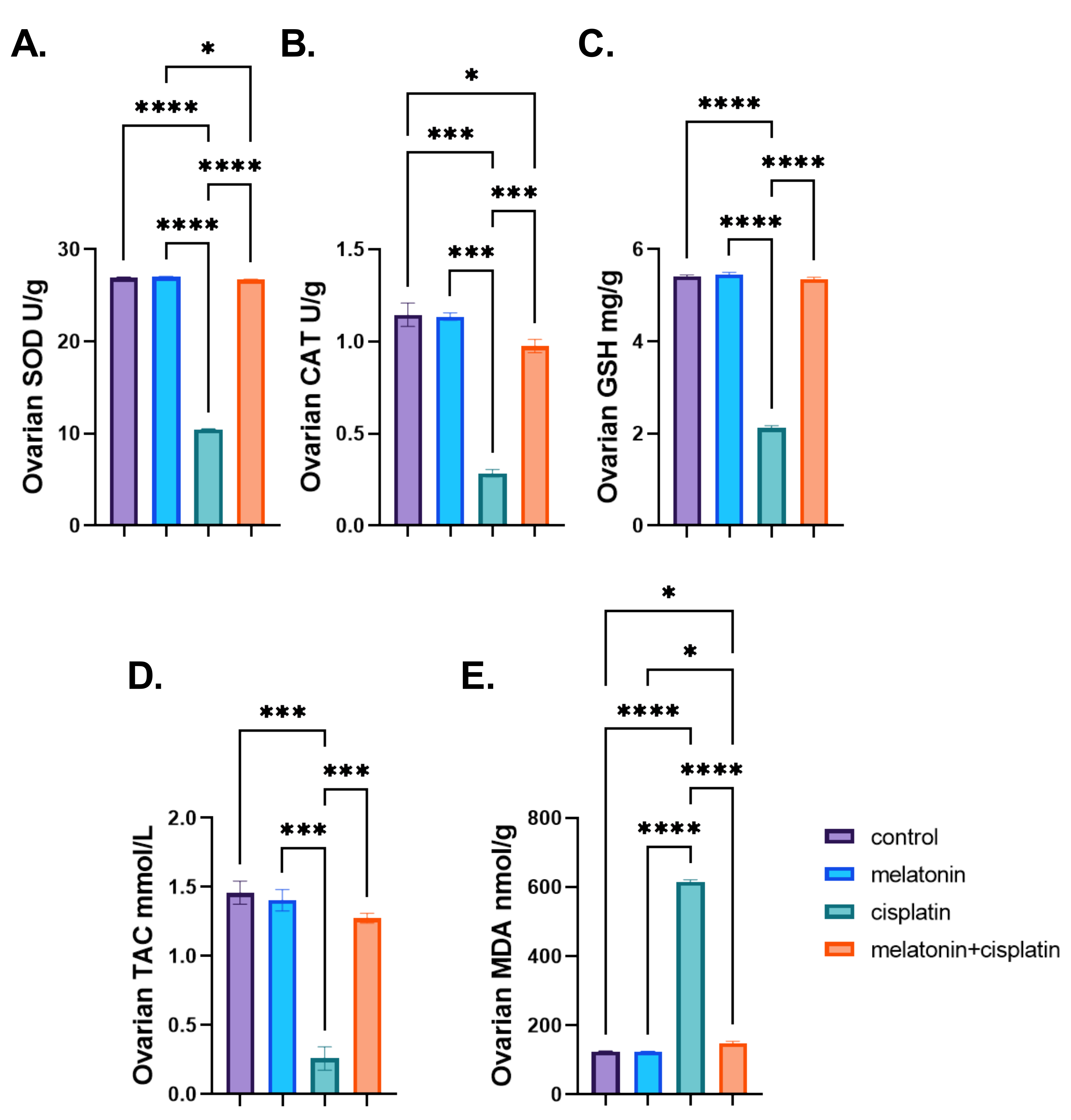
3.5. Ovarian Oxidative Stress Biomarkers
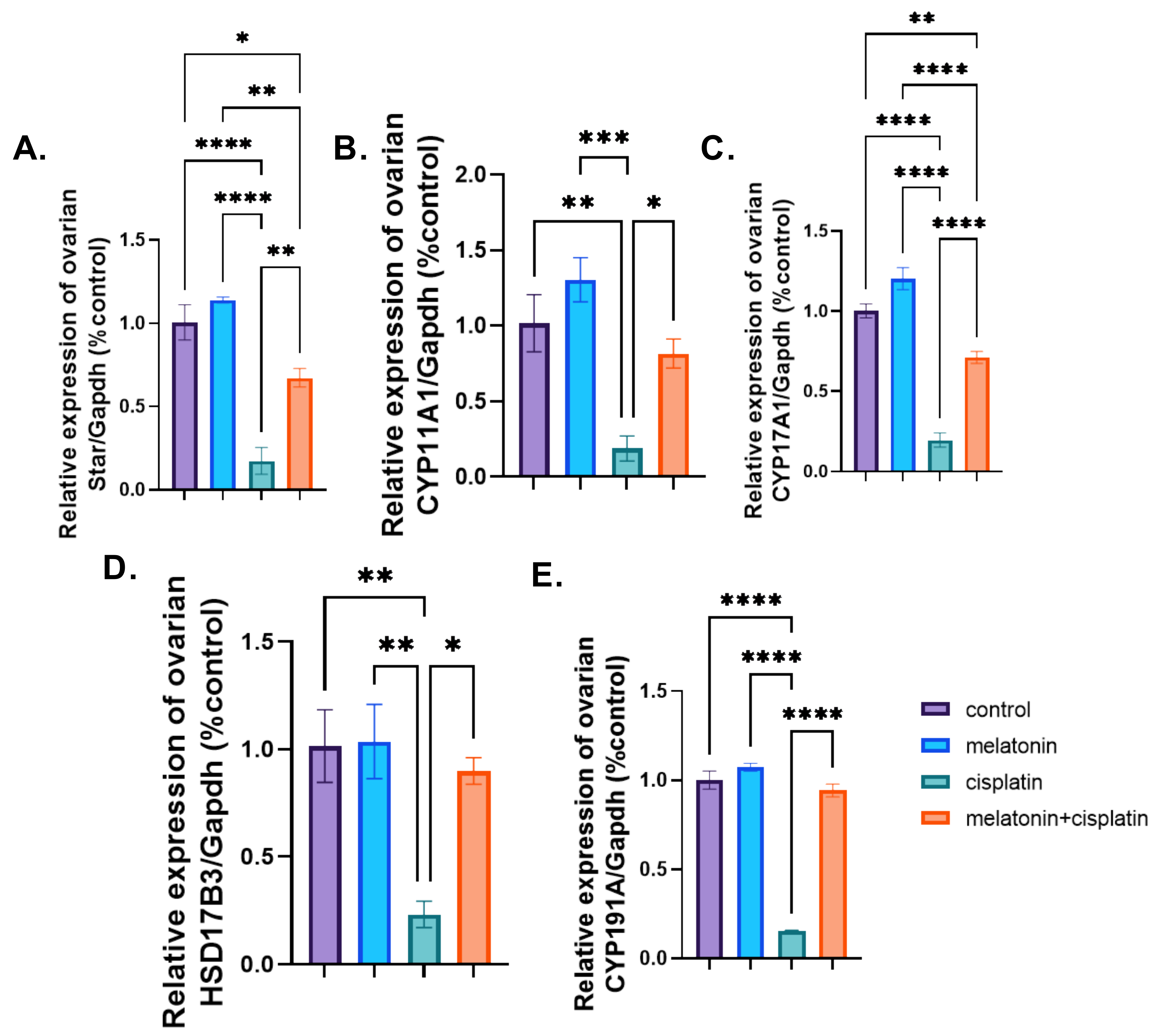
3.6. mRNA Expression of Ovarian Steroidogenic Pathway
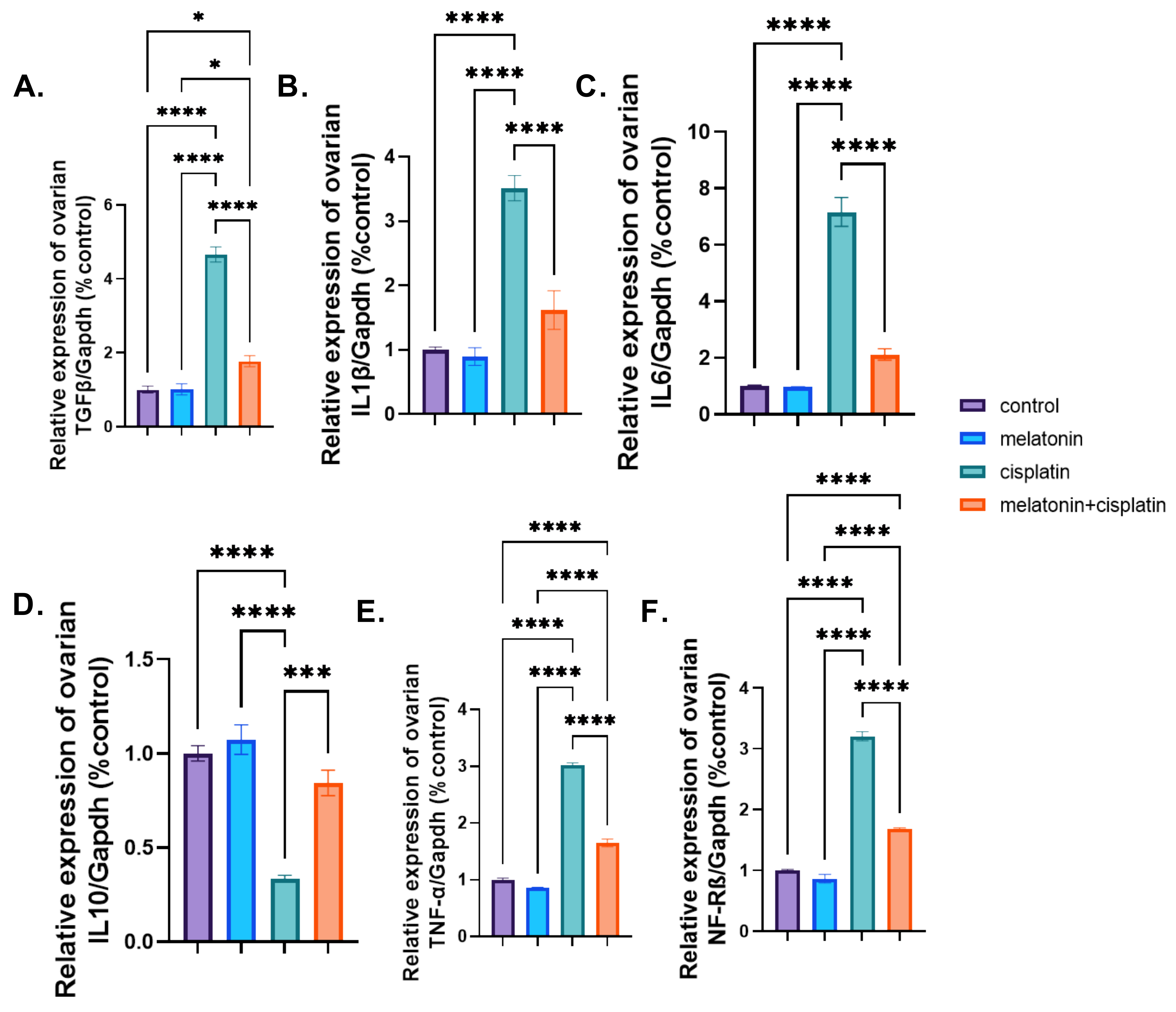
3.7. mRNA Expression of Proinflammatory and Anti-Inflammatory Markers
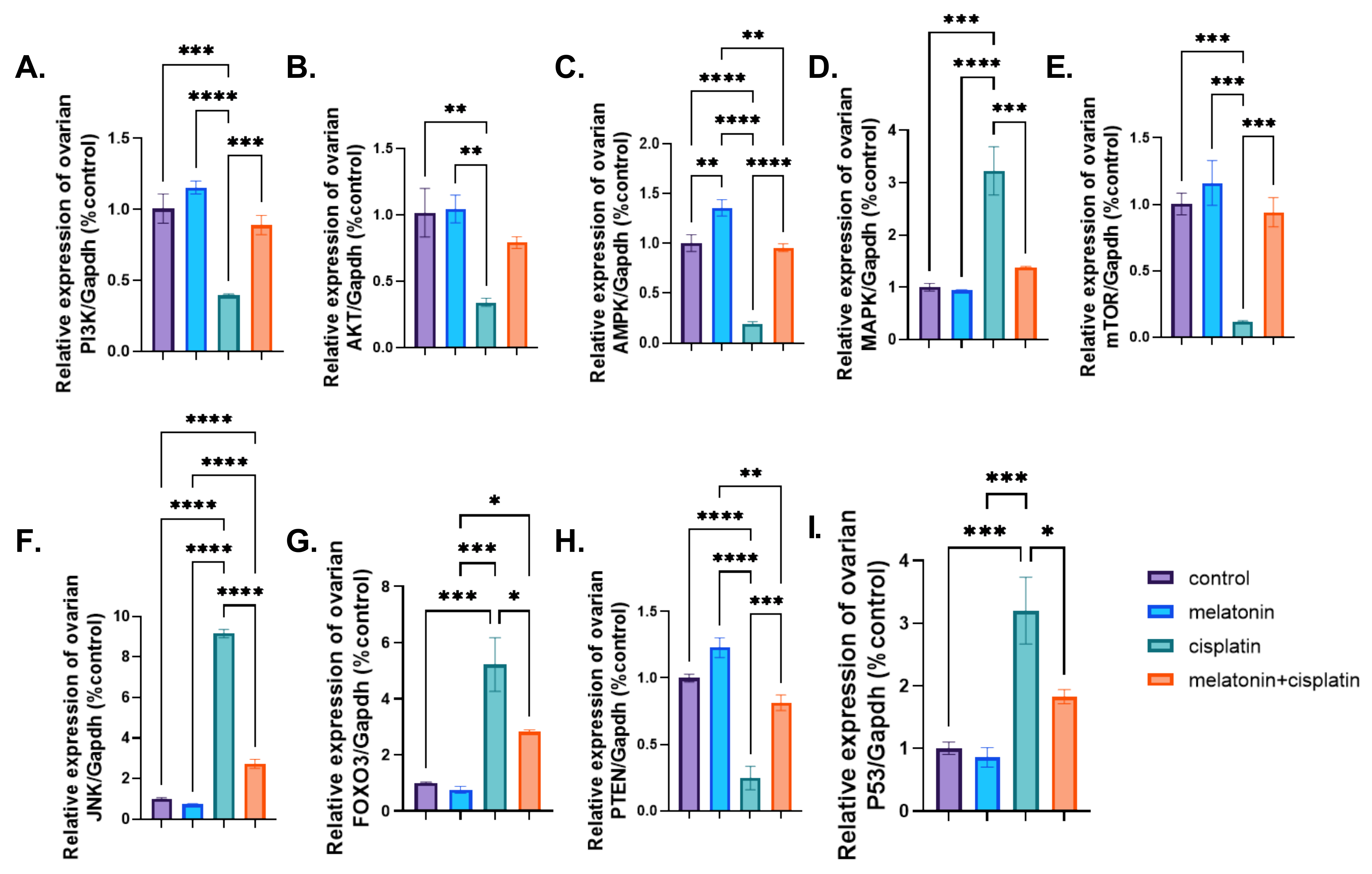
3.8. mRNA Expression of PI3K-Akt/mTOR/AMPK Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vassilakopoulou, M.; Boostandoost, E.; Papaxoinis, G.; de La Motte Rouge, T.; Khayat, D.; Psyrri, A. Anticancer treatment and fertility: Effect of therapeutic modalities on reproductive system and functions. Crit. Rev. Oncol. Hematol. 2016, 97, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Amorim, C.A.; Asiabi, P.; Ouni, E.; Vilela, J.; Camboni, A.; Chiti, M.C. Transplantation of Isolated Follicles and the Engineered Ovary. In Female and Male Fertility Preservation; Springer: Cham, Switzerland, 2022; pp. 353–367. [Google Scholar] [CrossRef]
- Liang, X.J.; Chen, C.; Zhao, Y.; Wang, P.C. Circumventing tumor resistance to chemotherapy by nanotechnology. Methods Mol. Biol. 2010, 596, 467–488. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, B.B.; Barberino, R.S.; Menezes, V.G.; O Monte, A.P.; Lucia Silva, R.S.; Palheta, R.C., Jr.; Rolim, L.A.; V Pereira, E.C.; Oliveira, R.G., Jr.; Roberto S Almeida, J.G.; et al. Amburana cearensis leaf extract protects against cisplatin-induced ovarian damage through regulation of p-PTEN and p-Akt proteins in mice. Iran. J. Basic Med. Sci. 2022, 25, 683–689. [Google Scholar] [PubMed]
- Gąsiorkiewicz, B.M.; Koczurkiewicz-Adamczyk, P.; Piska, K.; Pękala, E.; Mateusz Gąsiorkiewicz, B.; Koczurkiewicz-Adamczyk, P.; Piska, K.; Pękala, E.; Gąsiorkiewicz, B.M.; Koczurkiewicz-Adamczyk, P.; et al. Autophagy modulating agents as chemosensitizers for cisplatin therapy in cancer. Investig. New Drugs 2021, 39, 538–563. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.A.; Clatot, F.; Demeestere, I.; Lambertini, M.; Morgan, A.; Nelson, S.M.; Peccatori, F.; Cameron, D. Cancer survivorship: Reproductive health outcomes should be included in standard toxicity assessments. Eur. J. Cancer 2021, 144, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Haghi-Aminjan, H.; Asghari, M.H.; Farhood, B.; Rahimifard, M.; Hashemi Goradel, N.; Abdollahi, M. The role of melatonin on chemotherapy-induced reproductive toxicity. J. Pharm. Pharmacol. 2018, 70, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.C.; Liu, X.C.; Yang, S.H.; Song, L.L.; Zhou, S.J.; Deng, S.L.; Tian, L.; Cheng, L.Y. Melatonin Inhibits Oxidative Stress and Apoptosis in Cryopreserved Ovarian Tissues via Nrf2/HO-1 Signaling Pathway. Front. Mol. Biosci. 2020, 7, 163. [Google Scholar] [CrossRef]
- Xu, H.; Mu, X.; Ding, Y.; Tan, Q.; Liu, X.; He, J.; Gao, R.; Li, N.; Geng, Y.; Wang, Y.; et al. Melatonin alleviates benzo(a)pyrene-induced ovarian corpus luteum dysfunction by suppressing excessive oxidative stress and apoptosis. Ecotoxicol. Environ. Saf. 2021, 207, 111561. [Google Scholar] [CrossRef]
- Anadón, A.; Ares, I.; Martínez-Larrañaga, M.-R.; Martínez, M.-A. Melatonin: A safe nutraceutical and clinical agent. In Nutraceuticals; Academic Press: Cambridge, MA, USA, 2021; pp. 537–553. [Google Scholar] [CrossRef]
- Tamura, H.; Jozaki, M.; Tanabe, M.; Shirafuta, Y.; Mihara, Y.; Shinagawa, M.; Tamura, I.; Maekawa, R.; Sato, S.; Taketani, T.; et al. Importance of melatonin in assisted reproductive technology and ovarian aging. Int. J. Mol. Sci. 2020, 21, 1135. [Google Scholar] [CrossRef]
- Zhao, F.; Li, R.; Xiao, S.; Diao, H.; Viveiros, M.M.; Song, X.; Ye, X. Postweaning Exposure to Dietary Zearalenone, a Mycotoxin, Promotes Premature Onset of Puberty and Disrupts Early Pregnancy Events in Female Mice. Toxicol. Sci. 2013, 132, 431–442. [Google Scholar] [CrossRef]
- Kandemir, Y.B.; Aydin, C.; Gorgisen, G. The effects of melatonin on oxidative stress and prevention of primordial follicle loss via activation of mTOR pathway in the rat ovary. Cell. Mol. Biol. 2017, 63, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Caglayan, C.; Kandemir, F.M.; Yıldırım, S.; Kucukler, S.; Kılınc, M.A.; Saglam, Y.S. Zingerone ameliorates cisplatin-induced ovarian and uterine toxicity via suppression of sex hormone imbalances, oxidative stress, inflammation and apoptosis in female wistar rats. Biomed. Pharmacother. 2018, 102, 517–530. [Google Scholar] [CrossRef]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Bancroft, J.D.; Layton, C. Connective and other mesenchymal tissues with their stains. In Bancroft’s Theory and Practice of Histological Techniques, 8th ed.; Suvarna, S.K., Bancroft, J.D., Eds.; Elsevier Health Sciences: Amsterdam, The Netherland, 2018; pp. 53–175. [Google Scholar]
- Kececi, M.; Akpolat, M.; Gulle, K.; Gencer, E.; Sahbaz, A. Evaluation of preventive effect of shilajit on radiation-induced apoptosis on ovaries. Arch. Gynecol. Obstet. 2016, 293, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Abulfadle, K.A.; Edrees, H.; Hassan, N.H.; Mohammed, H.O. Spexin and Metformin Comparative Ameliorated Ovarian and Liver Function Changes in Letrozole-Induced Polycystic Ovary Syndrome in Rats (Histological, Biochemical, Immunohistochemical and Morphometric Study). Egypt. J. Histol. 2022, 45, 774–790. [Google Scholar] [CrossRef]
- Mohammadghasemi, F.; Jahromi, S.K.; Hajizadeh, H.; Homafar, M.A.; Saadat, N. The protective effects of exogenous melatonin on nicotine-induced changes in mouse ovarian follicles. J. Reprod. Infertil. 2012, 13, 143–150. [Google Scholar] [PubMed]
- Khamis, T.; Abdelalim, A.F.; Saeed, A.A.; Edress, N.M.; Nafea, A.; Ebian, H.F.; Algendy, R.; Hendawy, D.M.; Arisha, A.H.; Abdallah, S.H. Breast milk MSCs upregulated β-cells PDX1, Ngn3, and PCNA expression via remodeling ER stress /inflammatory /apoptotic signaling pathways in type 1 diabetic rats. Eur. J. Pharmacol. 2021, 905, 174188. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Stehlik-Barry, K.; Babinec, A.J. Data Analysis with IBM SPSS Statistics; Packt Publishing Ltd.: Birmingham, UK, 2017. [Google Scholar]
- Zhang, X.; Peng, X.; Wang, C.; Olatunji, O.J.; Famurewa, A. Tiliacora triandra attenuates cisplatin triggered hepatorenal and testicular toxicity in rats by modulating oxidative inflammation, apoptosis and endocrine deficit. Front. Biosci. 2022, 27, 44. [Google Scholar] [CrossRef]
- Shati, A.A. Resveratrol improves sperm parameter and testicular apoptosis in cisplatin-treated rats: Effects on ERK1/2, JNK, and Akt pathways. Syst. Biol. Reprod. Med. 2019, 65, 236–249. [Google Scholar] [CrossRef]
- Menczel Schrire, Z.; Phillips, C.L.; Chapman, J.L.; Duffy, S.L.; Wong, G.; D’Rozario, A.L.; Comas, M.; Raisin, I.; Saini, B.; Gordon, C.J.; et al. Safety of higher doses of melatonin in adults: A systematic review and meta-analysis. J. Pineal Res. 2022, 72, e12782. [Google Scholar] [CrossRef]
- Mousavi, S.A.; Heydari, K.; Mehravaran, H.; Saeedi, M.; Alizadeh-Navaei, R.; Hedayatizadeh-Omran, A.; Shamshirian, A. Melatonin effects on sleep quality and outcomes of COVID-19 patients: An open-label, randomized, controlled trial. J. Med. Virol. 2022, 94, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Said, R.S.; Mantawy, E.M.; El-Demerdash, E. Mechanistic perspective of protective effects of resveratrol against cisplatin-induced ovarian injury in rats: Emphasis on anti-inflammatory and anti-apoptotic effects. Naunyn. Schmiedeb. Arch. Pharmacol. 2019, 392, 1225–1238. [Google Scholar] [CrossRef] [PubMed]
- Usefzay, O.; Yari, S.; Amiri, P.; Hasanein, P. Evaluation of protective effects of methylene blue on cisplatin-induced nephrotoxicity. Biomed. Pharmacother. 2022, 150, 113023. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.T.; Zhou, J.; Wu, C.Y.; Zhang, W.; Shen, H.; Xu, J.D.; Zhang, Y.Q.; Long, F.; Li, S.L. Protective effects of Poria cocos and its components against cisplatin-induced intestinal injury. J. Ethnopharmacol. 2021, 269, 113722. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, H.E.L.; Hulail, M.A.E.; Mahmoud, S.M.; Yousef, D.M. Impact of cisplatin administration on cerebellar cortical structure and locomotor activity of infantile and juvenile albino rats: The role of oxidative stress. Anat. Sci. Int. 2022, 97, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Elbeltagy, A.; Mohamed, G.; Akeel, M.; Abdelaziz, K.; Elbakry, K.; Elsayed, A. Modulatory role of garlic (Allium sativum) extract against cisplatin-induced nephrotoxicity in female albino rats and their offspring. F1000Research 2022, 11, 504. [Google Scholar] [CrossRef]
- Wang, N.; Li, H.; Zhu, Y.; Li, N.; Chen, Z.J.; Zhang, C. Melatonin protects against epirubicin-induced ovarian damage. J. Reprod. Dev. 2020, 66, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, E.J.; Santacruz-Márquez, R.; Mourikes, V.E.; Neff, A.M.; Laws, M.J.; Flaws, J.A. Effects of Phthalate Mixtures on Ovarian Folliculogenesis and Steroidogenesis. Toxics 2022, 10, 251. [Google Scholar] [CrossRef]
- Dayangan Sayan, C.; Tulmac, O.B.; Karaca, G.; Ozkan, Z.S.; Yalcin, S.; Devrim, T.; Dindar Badem, N. Could erythropoietin reduce the ovarian damage of cisplatin in female rats? Gynecol. Endocrinol. 2018, 34, 309–313. [Google Scholar] [CrossRef]
- Cheng, J.C.; Fang, L.; Li, Y.; Wang, S.; Li, Y.; Yan, Y.; Jia, Q.; Wu, Z.; Wang, Z.; Han, X.; et al. Melatonin stimulates aromatase expression and estradiol production in human granulosa-lutein cells: Relevance for high serum estradiol levels in patients with ovarian hyperstimulation syndrome. Exp. Mol. Med. 2020, 52, 1341–1350. [Google Scholar] [CrossRef]
- Hashem, K.S.; Elkelawy, A.M.M.H.; Abd-Allah, S.; Helmy, N.A. Involvement of Mfn2, Bcl2/Bax signaling and mitochondrial viability in the potential protective effect of Royal jelly against mitochondria-mediated ovarian apoptosis by cisplatin in rats. Iran. J. Basic Med. Sci. 2020, 23, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.; Albahlol, I.A.; Wani, F.A.; Abd-Eltawab Tammam, A.; Kelleni, M.T.; Sayeed, M.U.; Abd El-Fadeal, N.M.; Mohamed, A.A. Resveratrol protects against cisplatin-induced ovarian and uterine toxicity in female rats by attenuating oxidative stress, inflammation and apoptosis: Resveratrol protects against cisplatin toxicity in rats. Chem. Biol. Interact. 2021, 338, 109402. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Ortiz, L.; Luderer, U. Charged iron particles, components of space radiation, destroy ovarian follicles. Hum. Reprod. 2016, 31, 1816–1826. [Google Scholar] [CrossRef]
- Wang, X.; Meng, K.; He, Y.; Wang, H.; Zhang, Y.; Quan, F. Melatonin stimulates STAR expression and progesterone production via activation of the PI3K/AKT pathway in bovine theca cells. Int. J. Biol. Sci. 2019, 15, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Shan, W.; Li, N.; Zhou, B.; Guo, E.; Xia, M.; Lu, H.; Wu, Y.; Chen, J.; Wang, B.; et al. Melatonin provides protection against cisplatin-induced ovarian damage and loss of fertility in mice. Reprod. Biomed. Online 2021, 42, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Kulhan, N.; Kulhan, M.; TÜRKLER, C.; Ata, N.; Kiremitli, T.; Kiremitli, S.; Cimen, F.E.R.D.A.; SÜLEYMAN, H.; Toprak, V. Effect of lycopene on oxidative ovarian damage induced by cisplatin in rats. Gen. Physiol. Biophys. 2019, 38, 253–258. [Google Scholar] [CrossRef]
- Ayazoglu Demir, E.; Mentese, A.; Kucuk, H.; Turkmen Alemdar, N.; Demir, S. p-Coumaric acid alleviates cisplatin-induced ovarian toxicity in rats. J. Obstet. Gynaecol. Res. 2022, 48, 411–419. [Google Scholar] [CrossRef]
- Şahin, H.Ö.; Duran, M.N.; Sılan, F.; Sılan, E.; Sıddıkoglu, D.; Kılınç, N. Investigation of TAp63 gene expression and follicle count using melatonin in cisplatin-induced ovarian toxicity. Int. J. Res. Med. Sci. 2021, 9, 658. [Google Scholar] [CrossRef]
- Yang, Q.; Zhu, J.; Luo, X.; Li, F.; Cong, L.; Wang, Y.; Sun, Y. Melatonin attenuates cadmium-induced ovulatory dysfunction by suppressing endoplasmic reticulum stress and cell apoptosis. Reprod. Biol. Endocrinol. 2019, 17, 1–8. [Google Scholar] [CrossRef]
- Walton, K.L.; Johnson, K.E.; Harrison, C.A. Targeting TGF-β Mediated SMAD Signaling for the Prevention of Fibrosis. Front. Pharmacol. 2017, 8, 461. [Google Scholar] [CrossRef]
- Sayed, A.A.; El-deek, S.E.M.; El-baz, M.A.; Sabry, D.; Al-rageaey, A.; Mansey, A.; Meligy, F.Y.; Abdelaziz, K. Exosomes Derived from Bone Marrow Mesenchymal Stem Cells Restore Cisplatin Induced Ovarian Damage by Promoting Stem Cell Survival, Meiotic, and Apoptotic Markers. J. Med. Med. Sci. 2017, 6, 116–130. [Google Scholar]
- Cui, L.; Bao, H.; Liu, Z.; Man, X.; Liu, H.; Hou, Y.; Luo, Q.; Wang, S.; Fu, Q.; Zhang, H. hUMSCs regulate the differentiation of ovarian stromal cells via TGF-β1/Smad3 signaling pathway to inhibit ovarian fibrosis to repair ovarian function in POI rats. Stem Cell Res. Ther. 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Goktepe, O.; Balcioglu, E.; Baran, M.; Cengiz, O.; Ceyhan, A.; Suna, P.A.; Bolat, D.; Yalcin, B.; Yay, A. Nonylphenol, Protective effects of melatonin on female rat ovary treated with nonylphenol. Biotech. Histochem. 2022, 1–7. [Google Scholar] [CrossRef] [PubMed]
- San-Miguel, B.; Fernández-Palanca, P.; Mauriz, J.L.; Tuñón, M.J.; González-Gallego, J. Beneficial effects of melatonin on liver fibrosis: A systematic review of current biological evidence. J. Cell. Physiol. 2022, 237, 2740–2757. [Google Scholar] [CrossRef]
- Hu, W.; Ma, Z.; Jiang, S.; Fan, C.; Deng, C.; Yan, X.; Di, S.; Lv, J.; Reiter, R.J.; Yang, Y. Melatonin: The dawning of a treatment for fibrosis? J. Pineal Res. 2016, 60, 121–131. [Google Scholar] [CrossRef]
- Abdellatief, S.A.; Galal, A.A.A.; Farouk, S.M.; Abdel-Daim, M.M. Ameliorative effect of parsley oil on cisplatin-induced hepato-cardiotoxicity: A biochemical, histopathological, and immunohistochemical study. Biomed. Pharmacother. 2017, 86, 482–491. [Google Scholar] [CrossRef]
- Deniz, G.Y.; Laloglu, E.; Altun, S.; Yiğit, N.; Gezer, A. Antioxidant and anti-apoptotic effects of vitexilactone on cisplatin-induced nephrotoxicity in rats. Biotech. Histochem. 2020, 95, 381–388. [Google Scholar] [CrossRef]
- Deng, C.C.; Zhang, J.P.; Huo, Y.N.; Xue, H.Y.; Wang, W.; Zhang, J.J.; Wang, X.Z. Melatonin alleviates the heat stress-induced impairment of Sertoli cells by reprogramming glucose metabolism. J. Pineal Res. 2022, 73, e12819. [Google Scholar] [CrossRef]
- Sayan, M.; Karabulut, D.; Özdamar, S. Assessment of the protective and therapeutic effect of melatonin against thioacetamide-induced acute liver damage. J. Biochem. Mol. Toxicol. 2020, 34, e22450. [Google Scholar] [CrossRef]
- Akçay, N.C.; Ömeroğlu, S.; Dizakar, S.Ö.A.; Kavutçu, M.; Türkoğlu, İ.; Eşmekaya, M.A.; Peker, T.V. The effects of melatonin on possible damage that will occur on adipocytokines and liver tissue by coadministration of fructose and bisphenol a (BPA). Environ. Sci. Pollut. Res. 2020, 27, 16231–16245. [Google Scholar] [CrossRef]
- Zhang, X.; Niu, Y.; Huang, Y. Melatonin inhibits cell proliferation in a rat model of breast hyperplasia by mediating the PTEN/AKT pathway. Oncol. Rep. 2021, 45, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rafiee, Z.; Moaiedi, M.Z.; Gorji, A.V.; Mansouri, E. p-Coumaric Acid Mitigates Doxorubicin-Induced Nephrotoxicity Through Suppression of Oxidative Stress, Inflammation and Apoptosis. Arch. Med. Res. 2020, 51, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Altındağ, F.; Meydan, İ. Evaluation of protective effects of gallic acid on cisplatin-induced testicular and epididymal damage. Andrologia 2021, 53, e14189. [Google Scholar] [CrossRef]
- Kara, O.; Kaymak, E.; Yakan, B. Protective effect of edaravone on cisplatin-induced injury in rat ovary. Arch. Gynecol. Obstet. 2022, 306, 1673–1678. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wang, B.; Xianghua, L.; Ningning, L.; Shi, K. Melatonin attenuates cisplatin-induced acute kidney injury in mice: Involvement of PPARα and fatty acid oxidation. Food Chem. Toxicol. 2022, 163, 112970. [Google Scholar] [CrossRef]
- Coşkun, N.; Hatipoǧlu, M.T.; Özoǧul, C.; Korkmaz, C.; Akyol, S.N.; Cilaker Micili, S.; Sanem Arik, G.; Erdoǧan, D. The protective effects of acetyl L-carnitine on testis gonadotoxicity induced by cisplatin in rats. Balkan Med. J. 2013, 30, 235–241. [Google Scholar] [CrossRef]
- Wu, J.Y.; Xiong, Y.; Xu, B.; Huang, Z.M.; Cheng, X.F. Dioscorea deltoidea Leaf Extract (DDLE) Attenuates Cisplatin-Induced Ovarian Injury via Regulation of Oxidative Stress. Dokl. Biochem. Biophys. 2021, 499, 282–288. [Google Scholar] [CrossRef]
- Algandaby, M.M. Quercetin attenuates cisplatin-induced ovarian toxicity in rats: Emphasis on anti-oxidant, anti-inflammatory and anti-apoptotic activities. Arab. J. Chem. 2021, 14, 103191. [Google Scholar] [CrossRef]
- Song, R.; Ritonga, F.N.; Yu, H.; Ding, C.; Zhao, X. Plant Melatonin: Regulatory and Protective Role. Horticulturae 2022, 8, 810. [Google Scholar] [CrossRef]
- Ristić, L.; Rančić, M.; Radović, M.; Krtinić, D.; Pavlović, M.; Ilić, B.; Milojković, M.; Živković, N.; Turković, K.; Sokolović, D. Melatonin inhibits apoptosis and oxidative tissue damage in cisplatin-induced pulmonary toxicity in rats. Arch. Med. Sci. 2020, 16. [Google Scholar] [CrossRef]
- de Araujo, J.G.; Serra, L.S.M.; Lauand, L.; Kückelhaus, S.A.S.; Sampaio, A.L.L. Protective effect of melatonin on cisplatin-induced ototoxicity in rats. Anticancer Res. 2019, 39, 2453–2458. [Google Scholar] [CrossRef] [PubMed]
- Zakria, M.; Ahmad, N.; Al Kury, L.T.; Alattar, A.; Uddin, Z.; Siraj, S.; Ullah, S.; Alshaman, R.; Khan, M.I.; Shah, F.A. Melatonin rescues the mice brain against cisplatin-induced neurodegeneration, an insight into antioxidant and anti-inflammatory effects. Neurotoxicology 2021, 87, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gentilin, E.; Simoni, E.; Candito, M.; Cazzador, D.; Astolfi, L. Cisplatin-Induced Ototoxicity: Updates on Molecular Targets. Trends Mol. Med. 2019, 25, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.R.; Zhu, W.N.; Tao, W.; Lin, W.Q.; Cheng, C.C.; Deng, H.; Ni, G.X. Moxibustion against Cyclophosphamide-Induced Premature Ovarian Failure in Rats through Inhibiting NLRP3-/Caspase-1-/GSDMD-Dependent Pyroptosis. Evidence-Based Complement. Altern. Med. 2021, 2021, 8874757. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Zhang, Y.; Tang, C.; Hou, Y.; Ai, X.; Chen, X.; Zhang, Y.; Wang, X.; Meng, X. Gallic acid: Pharmacological activities and molecular mechanisms involved in inflammation-related diseases. Biomed. Pharmacother. 2021, 133, 110985. [Google Scholar] [CrossRef]
- Ayres, L.S.; Berger, M.; de Oliveira Durli, I.C.L.; Kuhl, C.P.; Terraciano, P.B.; Garcez, T.N.A.; Dos Santos, B.G.; Guimarães, J.A.; Passos, E.P.; Cirne-Lima, E.O. Kallikrein-kinin system and oxidative stress in cisplatin-induced ovarian toxicity. Reprod. Toxicol. 2020, 93, 1–9. [Google Scholar] [CrossRef]
- Gui, H.; Jin, Y.; Lin, A.; Wang, P.; Wang, Y.; Zhu, H. Rosmarinic acid relieves cisplatin-induced ovary toxicity in female mice via suppression of oxidative stress and inflammation. J. Biochem. Mol. Toxicol. 2021, 35, e22839. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin and inflammation—Story of a double-edged blade. J. Pineal Res. 2018, 65, e12525. [Google Scholar] [CrossRef]
- Radogna, F.; Diederich, M.; Ghibelli, L. Melatonin: A pleiotropic molecule regulating inflammation. Biochem. Pharmacol. 2010, 80, 1844–1852. [Google Scholar] [CrossRef]
- Cardinali, D.P. Melatonin and Healthy Aging, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2021. [Google Scholar] [CrossRef]
- Kello, M.; Takac, P.; Kubatka, P.; Kuruc, T.; Petrova, K.; Mojzis, J. Oxidative stress-induced DNA damage and apoptosis in clove buds-treated MCF-7 cells. Biomolecules 2020, 10, 139. [Google Scholar] [CrossRef]
- Hao, Y.; Huang, Y.; Chen, J.; Li, J.; Yuan, Y.; Wang, M.; Han, L.; Xin, X.; Wang, H.; Lin, D.; et al. Exopolysaccharide from Cryptococcus heimaeyensis S20 induces autophagic cell death in non-small cell lung cancer cells via ROS/p38 and ROS/ERK signalling. Cell Prolif. 2020, 53, e12869. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Araki, S.; Tsuchiya, Y.; Watanabe, Y. Oxidative Stress Orchestrates MAPK and Nitric-Oxide Synthase Signal. Int. J. Mol. Sci. 2020, 21, 8750. [Google Scholar] [CrossRef] [PubMed]
- Mantawy, E.M.; Said, R.S.; Abdel-Aziz, A.K. Mechanistic approach of the inhibitory effect of chrysin on inflammatory and apoptotic events implicated in radiation-induced premature ovarian failure: Emphasis on TGF-β/MAPKs signaling pathway. Biomed. Pharmacother. 2019, 109, 293–303. [Google Scholar] [CrossRef]
- Guo, R.; Wang, G.; Zhao, A.; Gu, J.; Sun, X.; Hu, G. Paeoniflorin Protects against Ischemia-Induced Brain Damages in Rats via Inhibiting MAPKs/NF-κB-Mediated Inflammatory Responses. PLoS ONE 2012, 7, e49701. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Kwon, M.; Jung, J.; Nam, T. Protein extracted from Porphyra yezoensis prevents cisplatin-induced nephrotoxicity by downregulating the MAPK and NF-κB pathways. Int. J. Mol. Med. 2018, 41, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhao, Q.; Shao, Y.; Cao, M.Z.; Zhao, M.; Wang, D. Melatonin inhibits the inflammation and apoptosis in rats with diabetic retinopathy via MAPK pathway. Eur. Rev. Med. Pharmacol. Sci. 2021, 23, 1–8. [Google Scholar] [CrossRef]
- Cao, Z.; Fang, Y.; Lu, Y.; Tan, D.; Du, C.; Li, Y.; Ma, Q.; Yu, J.; Chen, M.; Zhou, C.; et al. Melatonin alleviates cadmium-induced liver injury by inhibiting the TXNIP-NLRP3 inflammasome. J. Pineal Res. 2017, 62, e12389. [Google Scholar] [CrossRef] [PubMed]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.W.; Hsu, K.C.; Lee, J.W.; Ham, M.; Huh, J.Y.; Shin, H.J.; Kim, W.S.; Kim, J.B. Berberine Suppresses Proinflammatory Responses through AMPK Activation in Macrophages. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E955–E964. [Google Scholar] [CrossRef]
- Chien, L.H.; Wu, C.T.; Deng, J.S.; Jiang, W.P.; Huang, W.C.; Huang, G.J. Salvianolic acid c protects against cisplatin-induced acute kidney injury through attenuation of inflammation, oxidative stress and apoptotic effects and activation of the CaMKK–AMPK–sirt1-associated signaling pathway in mouse models. Antioxidants 2021, 10, 1620. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, T.; Wang, C.; Meng, Q.; Huo, X.; Wang, C.; Sun, P.; Sun, H.; Ma, X.; Wu, J.; et al. Catalpol-Induced AMPK Activation Alleviates Cisplatin-Induced Nephrotoxicity through the Mitochondrial-Dependent Pathway without Compromising Its Anticancer Properties. Oxid. Med. Cell. Longev. 2021, 2021, 7467156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, Z.; Wang, J.; Lv, D.; Zhu, T.; Wang, F.; Tian, X.; Yao, Y.; Ji, P.; Liu, G. Melatonin regulates the activities of ovary and delays the fertility decline in female animals via MT1/AMPK pathway. J. Pineal Res. 2019, 66, e12550. [Google Scholar] [CrossRef] [PubMed]
- Luan, P.; Zhang, H.; Chen, X.; Zhu, Y.; Hu, G.; Cai, J. Ecotoxicology and Environmental Safety apoptosis and mitochondrial dysfunction through the AMPK-Sirt1-PGC-1 α axis in fish kidney cells (CIK). Ecotoxicol. Environ. Saf. 2022, 232, 113276. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Xie, Y.; Li, S.; Liang, Y.; Qiu, Q.; Lin, H.; Zhang, Q. Rapamycin Prevents cyclophosphamide-induced Over-activation of Primordial Follicle pool through PI3K/Akt/mTOR Signaling Pathway In Vivo. J. Ovarian Res. 2017, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mikaeili, S.; Rashidi, B.H.; Safa, M.; Najafi, A.; Sobhani, A.; Asadi, E.; Abbasi, M. Altered FoxO3 expression and apoptosis in granulosa cells of women with polycystic ovary syndrome. Arch. Gynecol. Obstet. 2016, 294, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.D. PTEN-PI3K pathway alterations in advanced prostate cancer and clinical implications. Prostate 2022, 82, S60–S72. [Google Scholar] [CrossRef] [PubMed]
- Tauber, S.; Steinbrenner, H.; Klotz, L.O. FoxO transcription factors in the control of redox homeostasis and fuel metabolism. In Oxidative Stress; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 315–330. [Google Scholar] [CrossRef]
- Yiming, Z.; Zhaoyi, L.; Jing, L.; Jinliang, W.; Zhiqiang, S.; Guangliang, S.; Shu, L. Cadmium induces the thymus apoptosis of pigs through ROS-dependent PTEN/PI3K/AKT signaling pathway. Environ. Sci. Pollut. Res. 2021, 28, 39982–39992. [Google Scholar] [CrossRef]
- Barberino, R.S.; Menezes, V.G.; Ribeiro, A.E.A.S.; Palheta, R.C.; Jiang, X.; Smitz, J.E.J.; Matos, M.H.T. Melatonin protects against cisplatin-induced ovarian damage in mice via the MT1 receptor and antioxidant activity. Biol. Reprod. 2017, 96, 1244–1255. [Google Scholar] [CrossRef]
- Lins, T.L.B.G.; Gouveia, B.B.; Barberino, R.S.; Silva, R.L.S.; Monte, A.P.O.; Pinto, J.G.C.; Campinho, D.S.P.; Palheta, R.C.; Matos, M.H.T. Rutin prevents cisplatin-induced ovarian damage via antioxidant activity and regulation of PTEN and FOXO3a phosphorylation in mouse model. Reprod. Toxicol. 2020, 98, 209–217. [Google Scholar] [CrossRef]
- Chang, E.M.; Lim, E.; Yoon, S.; Jeong, K.; Bae, S.; Lee, D.R.; Yoon, T.K.; Choi, Y.; Lee, W. Cisplatin induces overactivation of the dormant primordial follicle through PTEN/AKT/FOXO3α pathway which leads to loss of ovarian reserve in mice. PLoS ONE 2015, 10, e0144245. [Google Scholar] [CrossRef]
- Xie, F.; Zhang, J.; Zhai, M.; Liu, Y.; Hu, H.; Yu, Z.; Zhang, J.; Lin, S.; Liang, D.; Cao, Y. Melatonin ameliorates ovarian dysfunction by regulating autophagy in PCOS via the PI3K-Akt pathway. Reproduction 2021, 162, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Na, Y.; Hong, K.; Lee, S.; Moon, S.; Cho, M.; Park, M.; Lee, O.H.; Chang, E.M.; Lee, D.R.; et al. Synergistic effect of melatonin and ghrelin in preventing cisplatin-induced ovarian damage via regulation of FOXO3a phosphorylation and binding to the p27Kip1 promoter in primordial follicles. J. Pineal Res. 2017, 63, e12432. [Google Scholar] [CrossRef] [PubMed]
- Eldani, M.; Luan, Y.; Xu, P.C.; Bargar, T.; Kim, S.Y. Continuous treatment with cisplatin induces the oocyte death of primordial follicles without activation. FASEB J. 2020, 34, 13885–13899. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene & Accession Numbers | Forward Primer Sequence (5′ to 3′) | Reverse Primer Sequence (5′ to 3′) |
|---|---|---|
| PI3K (NM_053481.2) | CGAGAGTACGCTGTAGGCTG | AGAAACTGGCCAATCCTCCG |
| Pten (NM_031606.2) | ATACCAGGACCAGAGGAAACC | TTGTCATTATCCGCACGCTC |
| AKT (NM_033230.3) | GAAGGAGAAGGCCACAGGTC | TTCTGCAGGACACGGTTCTC |
| mTOR (NM_019906.2) | GCAATGGGCACGAGTTTGTT | AGTGTGTTCACCAGGCCAAA |
| FOXO3 (NM_017066.3) | AACAAAGGCAGCCTGCTAGT | TCGACGTTGCTGCTGGTATT |
| TGFβ1 (NM_021578.2) | AGGGCTACCATGCCAACTTC | CCACGTAGTAGACGATGGC |
| p38-MAPK (NM_019302.1) | TGGAGGTAACCAGGAGGGTT | AAGGCTGTCTTGTCGTAGGC |
| JNK (NM_053829.2) | TCCAGTTCTCGTACCCGCTA | AGCATGGCGTGACACAGTAA |
| AMPK (NM_023991.1) | GGCGTGTGAAGATCGGACA | GGCCTGTCAATTGATGTTCTCC |
| P53 (NM_030989.3) | GTTCGTGTTTGTGCCTGTCC | TGCTCTCTTTGCACTCCCTG |
| StAr (NM_031558.3) | CCCAAATGTCAAGGAAATCA | AGGCATCTCCCCAAAGTG |
| CYP11A1 (NM_017286.3) | AAGTATCCGTGATGTGGG | TCATACAGTGTCGCCTTTTCT |
| CYP17A1 (NM_012753.2) | TGGCTTTCCTGGTGCACAATC | TGAAAGTTGGTGTTCGGCTGAAG |
| CYP19A1 (NM_017085.2) | GCTGAGAGACGTGGAGACCTG | CTCTGTCACCAACAACAGTGTGG |
| HSD17B3 (NM_054007.1) | AGTGTGTGAGGTTCTCCCGGTACCT | TACAACATTGAGTCCATGTCTGGCCAG |
| GAPDH (NM_017008.4) | GGCACAGTCAAGGCTGAGAATG | TGGTGGTGAAGACGCCAGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Shahat, A.; Hulail, M.A.E.; Soliman, N.M.M.; Khamis, T.; Fericean, L.M.; Arisha, A.H.; Moawad, R.S. Melatonin Mitigates Cisplatin-Induced Ovarian Dysfunction via Altering Steroidogenesis, Inflammation, Apoptosis, Oxidative Stress, and PTEN/PI3K/Akt/mTOR/AMPK Signaling Pathway in Female Rats. Pharmaceutics 2022, 14, 2769. https://doi.org/10.3390/pharmaceutics14122769
Al-Shahat A, Hulail MAE, Soliman NMM, Khamis T, Fericean LM, Arisha AH, Moawad RS. Melatonin Mitigates Cisplatin-Induced Ovarian Dysfunction via Altering Steroidogenesis, Inflammation, Apoptosis, Oxidative Stress, and PTEN/PI3K/Akt/mTOR/AMPK Signaling Pathway in Female Rats. Pharmaceutics. 2022; 14(12):2769. https://doi.org/10.3390/pharmaceutics14122769
Chicago/Turabian StyleAl-Shahat, Amal, Mohey A. E. Hulail, Nada M. M. Soliman, Tarek Khamis, Liana Mihaela Fericean, Ahmed Hamed Arisha, and Rania S. Moawad. 2022. "Melatonin Mitigates Cisplatin-Induced Ovarian Dysfunction via Altering Steroidogenesis, Inflammation, Apoptosis, Oxidative Stress, and PTEN/PI3K/Akt/mTOR/AMPK Signaling Pathway in Female Rats" Pharmaceutics 14, no. 12: 2769. https://doi.org/10.3390/pharmaceutics14122769
APA StyleAl-Shahat, A., Hulail, M. A. E., Soliman, N. M. M., Khamis, T., Fericean, L. M., Arisha, A. H., & Moawad, R. S. (2022). Melatonin Mitigates Cisplatin-Induced Ovarian Dysfunction via Altering Steroidogenesis, Inflammation, Apoptosis, Oxidative Stress, and PTEN/PI3K/Akt/mTOR/AMPK Signaling Pathway in Female Rats. Pharmaceutics, 14(12), 2769. https://doi.org/10.3390/pharmaceutics14122769