Evaluation of Tobramycin and Ciprofloxacin as a Synergistic Combination Against Hypermutable Pseudomonas Aeruginosa Strains via Mechanism-Based Modelling

 ,
,  , ,
, , 
Abstract
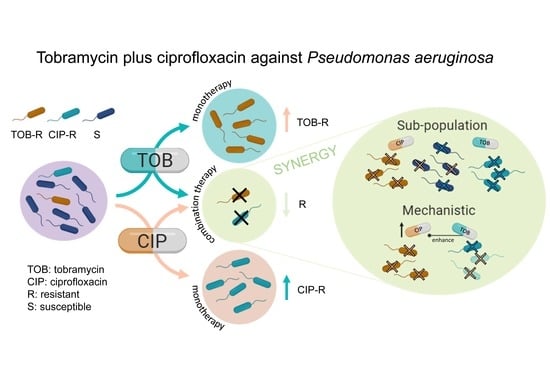
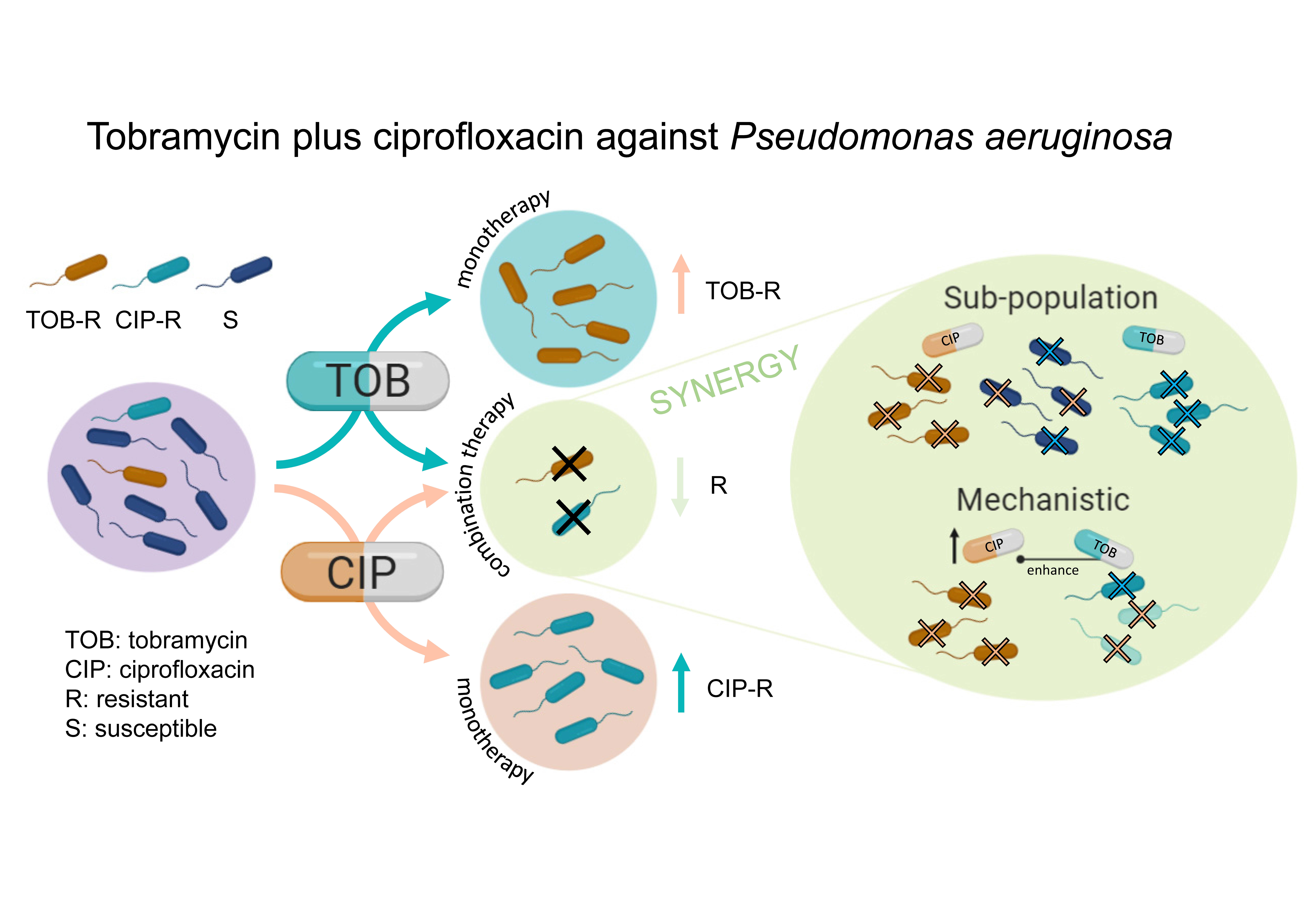
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Antibiotics Tested
2.2. Static Concentration Time-Kill Assays
2.3. Viable Counting of the Total and Resistant Populations
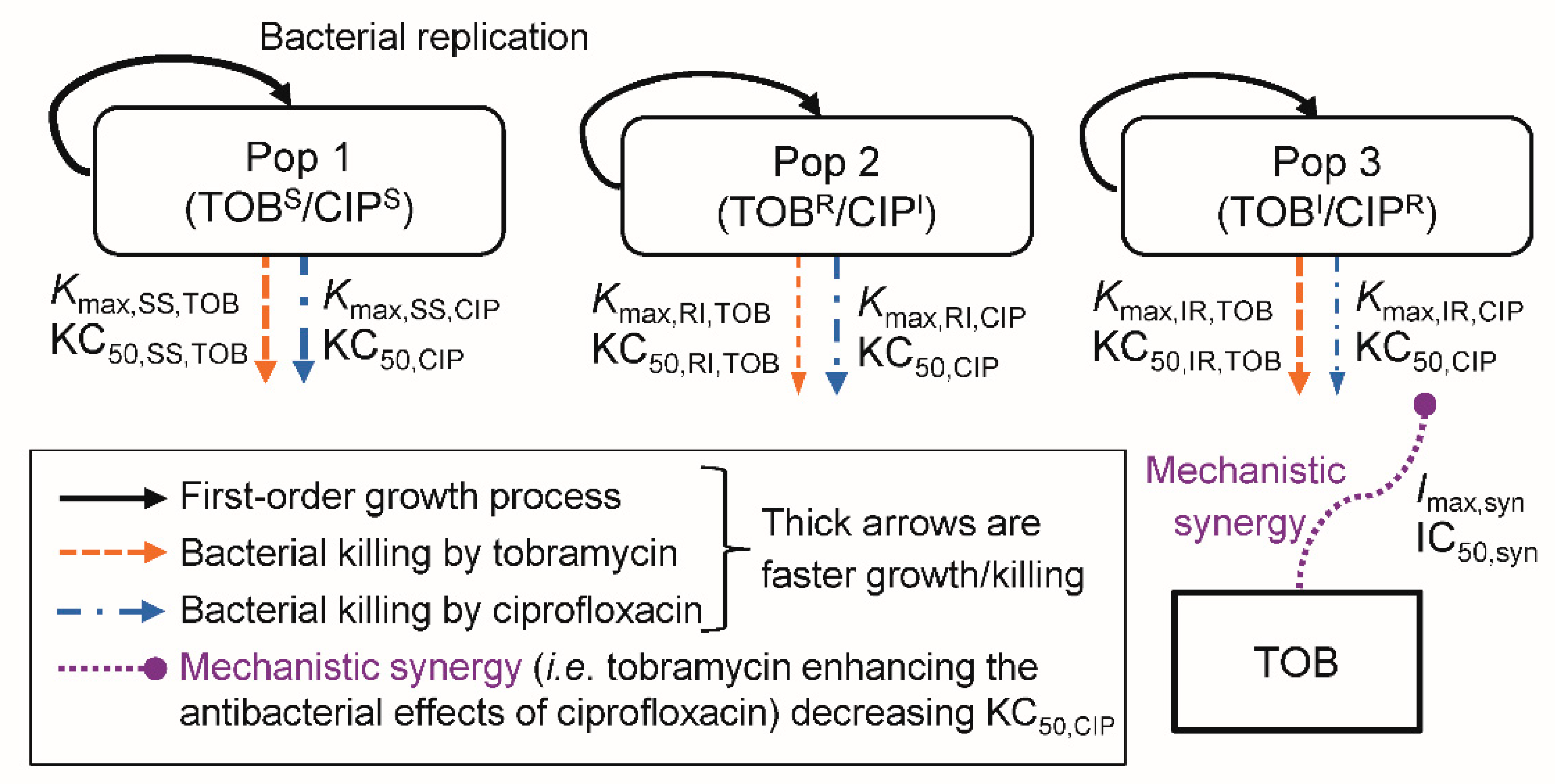
2.4. Mechanism-Based Modelling of Bacterial Killing and Resistance
2.4.1. The Life-Cycle Growth Model
2.4.2. Synergy Modelling
2.4.3. Less-Susceptible Bacterial Populations
2.4.4. Initial Conditions and Observation Model
3. Results and Discussion
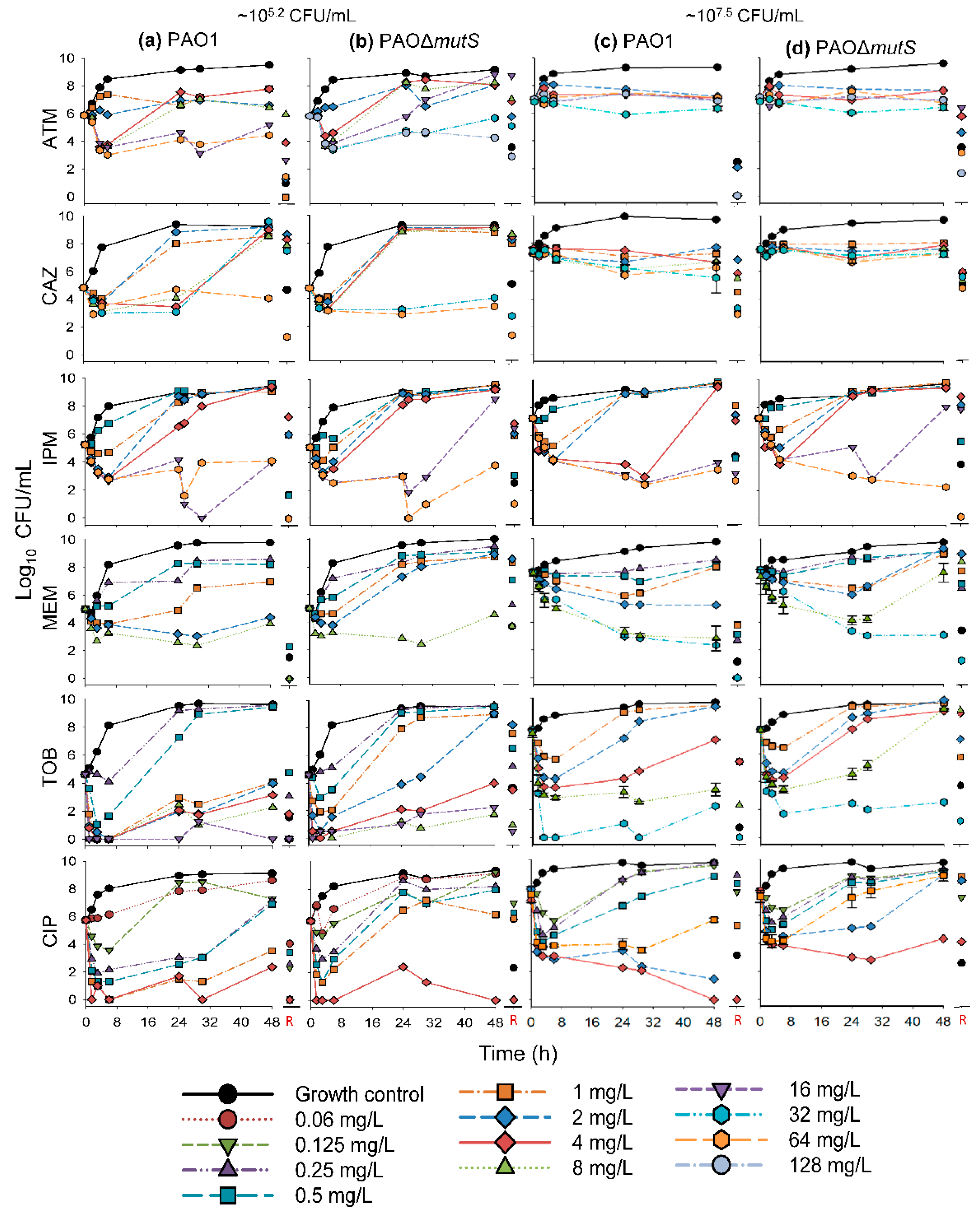
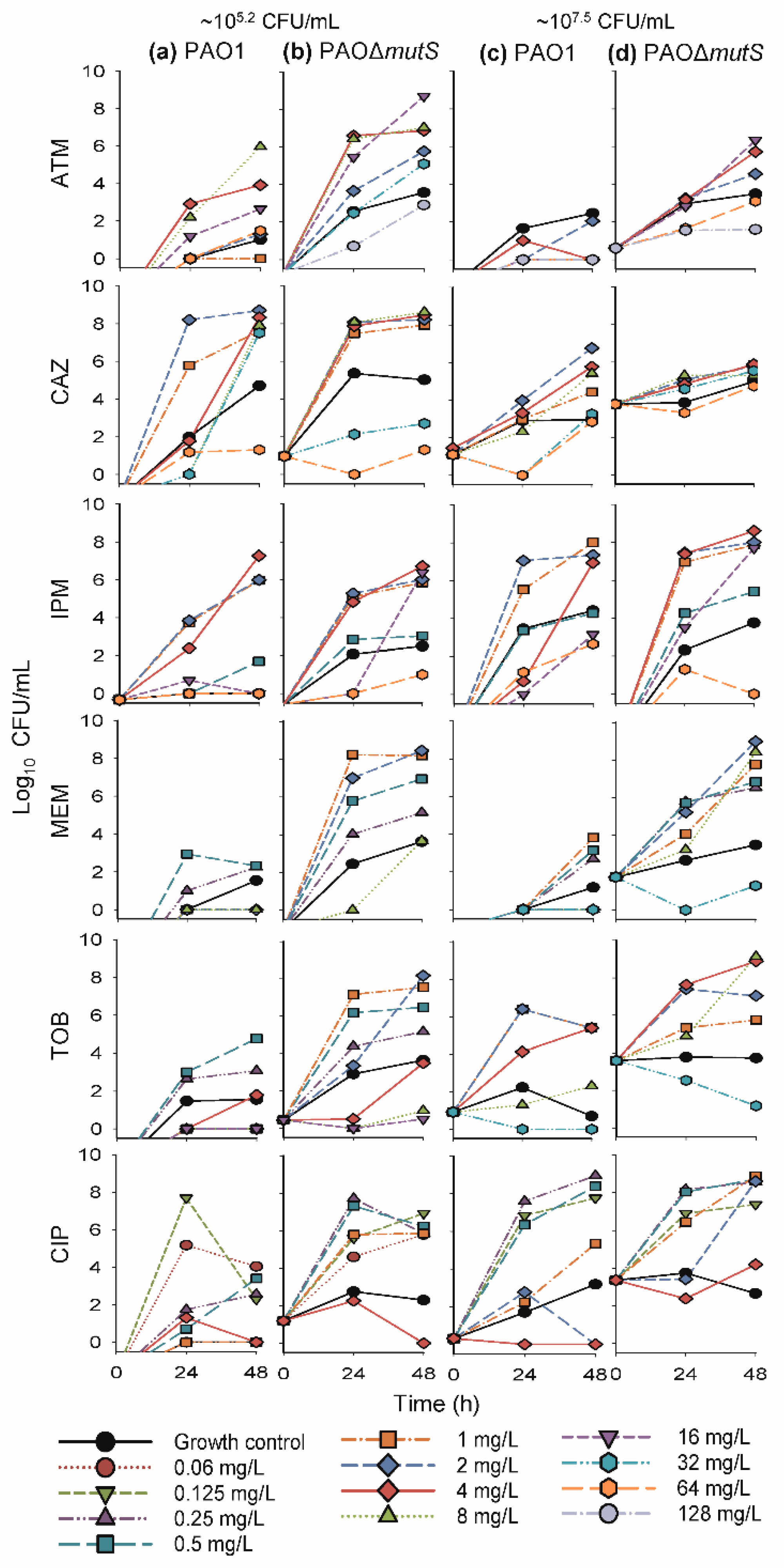
3.1. Antibacterial Effect of Common Antipseudomonal Antibiotics in Monotherapy vs PAO1 and PAO∆mutS
3.1.1. Antibacterial Effect of Beta-Lactam Antibiotics in Monotherapy
3.1.2. Antibacterial Effect of Fast-Acting Antipseudomonal Antibiotics in Monotherapy
3.2. Antibacterial Effect of Two Fast-Acting Antipseudomonal Antibiotics in Combination
3.2.1. Antibacterial Effect of Tobramycin and Ciprofloxacin against the Three Hypermutator Strains
3.2.2. Mechanism-Based Mathematical Modelling of the Tobramycin and Ciprofloxacin Combination
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bhagirath, A.Y.; Li, Y.; Somayajula, D.; Dadashi, M.; Badr, S.; Duan, K. Cystic fibrosis lung environment and Pseudomonas aeruginosa infection. BMC Pulm. Med. 2016, 16, 174. [Google Scholar] [CrossRef] [PubMed]
- Macia, M.D.; Blanquer, D.; Togores, B.; Sauleda, J.; Perez, J.L.; Oliver, A. Hypermutation is a key factor in development of multiple-antimicrobial resistance in Pseudomonas aeruginosa strains causing chronic lung infections. Antimicrob. Agents Chemother. 2005, 49, 3382–3386. [Google Scholar] [CrossRef] [PubMed]
- Oliver, A.; Canton, R.; Campo, P.; Baquero, F.; Blazquez, J. High frequency of hypermutable Pseudomonas aeruginosa in cystic fibrosis lung infection. Science 2000, 288, 1251–1254. [Google Scholar] [CrossRef] [PubMed]
- Smania, A.M.; Segura, I.; Pezza, R.J.; Becerra, C.; Albesa, I.; Argarana, C.E. Emergence of phenotypic variants upon mismatch repair disruption in Pseudomonas aeruginosa. Microbiology 2004, 150, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Bals, R.; Hubert, D.; Tummler, B. Antibiotic treatment of CF lung disease: From bench to bedside. J. Cyst. Fibros 2011, 10 (Suppl. 2), S146–S151. [Google Scholar] [CrossRef]
- Cabot, G.; Zamorano, L.; Moya, B.; Juan, C.; Navas, A.; Blazquez, J.; Oliver, A. Evolution of Pseudomonas aeruginosa antimicrobial resistance and fitness under low and high mutation rates. Antimicrob. Agents Chemother. 2016, 60, 1767–1778. [Google Scholar] [CrossRef] [PubMed]
- Oliver, A.; Levin, B.R.; Juan, C.; Baquero, F.; Blazquez, J. Hypermutation and the preexistence of antibiotic-resistant Pseudomonas aeruginosa mutants: Implications for susceptibility testing and treatment of chronic infections. Antimicrob. Agents Chemother. 2004, 48, 4226–4233. [Google Scholar] [CrossRef] [PubMed]
- Mulet, X.; Macia, M.D.; Mena, A.; Juan, C.; Perez, J.L.; Oliver, A. Azithromycin in Pseudomonas aeruginosa biofilms: Bactericidal activity and selection of nfxB mutants. Antimicrob. Agents Chemother. 2009, 53, 1552–1560. [Google Scholar] [CrossRef]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef]
- Rees, V.E.; Deveson Lucas, D.S.; Lopez-Causape, C.; Huang, Y.; Kotsimbos, T.; Bulitta, J.B.; Rees, M.C.; Barugahare, A.; Peleg, A.Y.; Nation, R.L.; et al. Characterization of hypermutator Pseudomonas aeruginosa isolates from patients with cystic fibrosis in Australia. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically: Approved Standard Ninth Edition M07-A9; CLSI: Wayne, PA, USA, 2012. [Google Scholar]
- Rees, V.E.; Bulitta, J.B.; Nation, R.L.; Tsuji, B.T.; Sorgel, F.; Landersdorfer, C.B. Shape does matter: Short high-concentration exposure minimizes resistance emergence for fluoroquinolones in Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2015, 70, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Viaene, E.; Chanteux, H.; Servais, H.; Mingeot-Leclercq, M.P.; Tulkens, P.M. Comparative stability studies of antipseudomonal beta-lactams for potential administration through portable elastomeric pumps (home therapy for cystic fibrosis patients) and motor-operated syringes (intensive care units). Antimicrob. Agents Chemother. 2002, 46, 2327–2332. [Google Scholar] [PubMed]
- Keel, R.A.; Sutherland, C.A.; Crandon, J.L.; Nicolau, D.P. Stability of doripenem, imipenem and meropenem at elevated room temperatures. Int. J. Antimicrob. Agents 2011, 37, 184–185. [Google Scholar] [CrossRef] [PubMed]
- Bulitta, J.B.; Bingolbali, A.; Shin, B.S.; Landersdorfer, C.B. Development of a new pre- and post-processing tool (SADAPT-TRAN) for nonlinear mixed-effects modeling in S-ADAPT. AAPS J. 2011, 13, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Bulitta, J.B.; Yang, J.C.; Yohonn, L.; Ly, N.S.; Brown, S.V.; D’Hondt, R.E.; Jusko, W.J.; Forrest, A.; Tsuji, B.T. Attenuation of colistin bactericidal activity by high inoculum of Pseudomonas aeruginosa characterized by a new mechanism-based population pharmacodynamic model. Antimicrob. Agents Chemother. 2010, 54, 2051–2062. [Google Scholar] [CrossRef] [PubMed]
- Bulitta, J.B.; Duffull, S.B.; Kinzig-Schippers, M.; Holzgrabe, U.; Stephan, U.; Drusano, G.L.; Sorgel, F. Systematic comparison of the population pharmacokinetics and pharmacodynamics of piperacillin in cystic fibrosis patients and healthy volunteers. Antimicrob. Agents Chemother. 2007, 51, 2497–2507. [Google Scholar] [CrossRef] [PubMed]
- Landersdorfer, C.B.; Kirkpatrick, C.M.; Kinzig-Schippers, M.; Bulitta, J.B.; Holzgrabe, U.; Drusano, G.L.; Sorgel, F. Population pharmacokinetics at two dose levels and pharmacodynamic profiling of flucloxacillin. Antimicrob. Agents Chemother. 2007, 51, 3290–3297. [Google Scholar] [CrossRef]
- Tsuji, B.T.; Okusanya, O.O.; Bulitta, J.B.; Forrest, A.; Bhavnani, S.M.; Fernandez, P.B.; Ambrose, P.G. Application of pharmacokinetic-pharmacodynamic modeling and the justification of a novel fusidic acid dosing regimen: Raising Lazarus from the dead. Clin. Infect. Dis. 2011, 52 (Suppl. 7), S513–S519. [Google Scholar] [CrossRef]
- Maidhof, H.; Johannsen, L.; Labischinski, H.; Giesbrecht, P. Onset of penicillin-induced bacteriolysis in staphylococci is cell cycle dependent. J. Bacteriol. 1989, 171, 2252–2257. [Google Scholar]
- Landersdorfer, C.B.; Ly, N.S.; Xu, H.; Tsuji, B.T.; Bulitta, J.B. Quantifying subpopulation synergy for antibiotic combinations via mechanism-based modeling and a sequential dosing design. Antimicrob. Agents Chemother. 2013, 57, 2343–2351. [Google Scholar] [CrossRef]
- Bulitta, J.B.; Ly, N.S.; Yang, J.C.; Forrest, A.; Jusko, W.J.; Tsuji, B.T. Development and qualification of a pharmacodynamic model for the pronounced inoculum effect of ceftazidime against Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2009, 53, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, B.T.; Bulitta, J.B.; Brown, T.; Forrest, A.; Kelchlin, P.A.; Holden, P.N.; Peloquin, C.A.; Skerlos, L.; Hanna, D. Pharmacodynamics of early, high-dose linezolid against vancomycin-resistant enterococci with elevated MICs and pre-existing genetic mutations. J. Antimicrob. Chemother. 2012, 67, 2182–2190. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Landersdorfer, C.B.; London, D.; Meng, R.; Lim, C.U.; Lin, C.; Lin, S.; Tang, H.Y.; Brown, D.; Van Scoy, B.; et al. Pharmacodynamic modeling of anti-cancer activity of tetraiodothyroacetic acid in a perfused cell culture system. PLoS Comput. Biol. 2011, 7, e1001073. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Bulitta, J.B.; Nation, R.L.; Landersdorfer, C.B. Optimization of synergistic combination regimens against carbapenem- and aminoglycoside-resistant clinical Pseudomonas aeruginosa isolates via mechanism-based pharmacokinetic/pharmacodynamic modeling. Antimicrob. Agents Chemother. 2017, 61, e01011-16. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Landersdorfer, C.B.; Nation, R.L.; Boyce, J.D.; Bulitta, J.B. Novel approach to optimize synergistic carbapenem-aminoglycoside combinations against carbapenem-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2015, 59, 2286–2298. [Google Scholar] [CrossRef]
- Rees, V.E.; Yadav, R.; Rogers, K.E.; Bulitta, J.B.; Wirth, V.; Oliver, A.; Boyce, J.D.; Peleg, A.Y.; Nation, R.L.; Landersdorfer, C.B. Meropenem combined with ciprofloxacin combats hypermutable Pseudomonas aeruginosa from respiratory infections of cystic fibrosis patients. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Ly, N.S.; Bulitta, J.B.; Rao, G.G.; Landersdorfer, C.B.; Holden, P.N.; Forrest, A.; Bergen, P.J.; Nation, R.L.; Li, J.; Tsuji, B.T. Colistin and doripenem combinations against Pseudomonas aeruginosa: Profiling the time course of synergistic killing and prevention of resistance. J. Antimicrob. Chemother. 2015, 70, 1434–1442. [Google Scholar] [CrossRef]
- Rees, V.E.; Bulitta, J.B.; Oliver, A.; Tsuji, B.T.; Rayner, C.R.; Nation, R.L.; Landersdorfer, C.B. Resistance suppression by high-intensity, short-duration aminoglycoside exposure against hypermutable and non-hypermutable Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2016, 71, 3157–3167. [Google Scholar] [CrossRef]
- Mouton, J.W.; Muller, A.E.; Canton, R.; Giske, C.G.; Kahlmeter, G.; Turnidge, J. MIC-based dose adjustment: Facts and fables. J. Antimicrob. Chemother. 2018, 73, 564–568. [Google Scholar] [CrossRef]
- Grayson, M.L.; Cosgrove, S.E.; Crowe, S.; Hope, W.; McCarthy, J.S.; Mills, J.; Mouton, J.W.; Paterson, D.L. Kucers’ The Use of Antibiotics: A Clinical Review of Antibacterial, Antifungal, Antiparasitic, and Antiviral Drugs, 7th ed.; Grayson, M.L., Ed.; CRC Press: London, UK, 2017; Volume 1, p. 194. [Google Scholar]
- Eng, R.H.; Smith, S.M.; Cherubin, C. Inoculum effect of new beta-lactam antibiotics on Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1984, 26, 42–47. [Google Scholar] [CrossRef]
- Henrichfreise, B.; Wiegand, I.; Luhmer-Becker, I.; Wiedemann, B. Development of resistance in wild-type and hypermutable Pseudomonas aeruginosa strains exposed to clinical pharmacokinetic profiles of meropenem and ceftazidime simulated in vitro. Antimicrob. Agents Chemother. 2007, 51, 3642–3649. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Tomida, J.; Kawamura, Y. Responses of Pseudomonas aeruginosa to antimicrobials. Front. Microbiol. 2014, 4, 422. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Jonker, M.J.; Moustakas, I.; Brul, S.; Ter Kuile, B.H. Dynamics of mutations during development of resistance by Pseudomonas aeruginosa against five antibiotics. Antimicrob. Agents Chemother. 2016, 60, 4229–4236. [Google Scholar] [CrossRef] [PubMed]
- Macia, M.D.; Borrell, N.; Segura, M.; Gomez, C.; Perez, J.L.; Oliver, A. Efficacy and potential for resistance selection of antipseudomonal treatments in a mouse model of lung infection by hypermutable Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2006, 50, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Poole, K. Aminoglycoside resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2005, 49, 479–487. [Google Scholar] [CrossRef]
- Poole, K. Pseudomonas aeruginosa: Resistance to the max. Front Microbiol. 2011, 2, 65. [Google Scholar] [CrossRef]
- Bulitta, J.B.; Ly, N.S.; Landersdorfer, C.B.; Wanigaratne, N.A.; Velkov, T.; Yadav, R.; Oliver, A.; Martin, L.; Shin, B.S.; Forrest, A.; et al. Two mechanisms of killing of Pseudomonas aeruginosa by tobramycin assessed at multiple inocula via mechanism-based modeling. Antimicrob. Agents Chemother. 2015, 59, 2315–2327. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAO1/PAOΔmutS 1 | CW19 2 | CW44 2 | |
|---|---|---|---|
| aztreonam | 4 | - | - |
| ceftazidime | 2 | - | - |
| imipenem | 2 | - | - |
| meropenem | 1 | - | - |
| tobramycin | 0.5 | 1 | 1 |
| ciprofloxacin | 0.125 | 0.5 | 0.19 |
| Parameter | Symbol (unit) | Population Mean Value (SE[%]) | |||
|---|---|---|---|---|---|
| PAO1 | PAOΔmutS | CW19 | CW44 | ||
| Bacterial growth and subpopulations | |||||
| Initial inoculum | Log10 CFU0 | 7.09 (2.05%) | 7.62 (1.71%) | 7.87 (2.75%) | 7.26 (1.39%) |
| Maximum population size | Log10 CFUmax | 9.33 (2.00%) | 9.01 (1.79%) | 9.00 (1.31%) | 8.94 (1.01%) |
| Mean generation time (MGT) | |||||
| TOBS/CIPS | 1/k12,SS (min) | 50.9 (4.55%) 1 | 55.9 (4.81%) 1 | 115 (6.26%) 1 | 124 (6.48%) 1 |
| TOBR/CIPI | 1/k12,RI (min) | 340 (13.1%) | 254 (19.4%) | 327 (4.96%) | 141 (8.57%) |
| TOBI/CIPR | 1/k12,IR (min) | 50.9 (4.55%) 1 | 55.9 (4.81%) 1 | 115 (6.26%) 1 | 124 (6.48%) 1 |
| Log10 mutation frequencies | |||||
| TOB | Log10 MFTOB | −3.68 (5.43%) | −3.93 (6.52%) | −3.3 (16.6%) | −4.81 (4.01%) |
| CIP | Log10 MFCIP | −7.68 (4.04%) | −8.39 (4.60%) | −5.79 (12.7%) | −7.47 (5.12%) |
| Killing by TOB | |||||
| Maximum killing rate constant | |||||
| TOBS/CIPS | Kmax,SS,TOB (h−1) | 12.2 (11.2%) | 10.8 (34.1%) | 5.18 (36.1%) | 3.26 (19.4%) |
| TOBR/CIPI | Kmax,RI,TOB (h−1) | 0.251 (22.9%) | 0.305 (42.1%) | 0.354 (15.1%) | 0.60 (18.0%) |
| TOBI/CIPR | Kmax,IR,TOB (h−1) | 1.17 (15.1%) | 0.367 (47.2%) | 0.690 (62.5%) | 5.02 (20.0%) |
| TOB concentration causing 50% of Kmax,TOB | KC50,TOB (mg/L) | 3.68 (21.5%) | 2.11 (41.9%) 2 | 53.4 (10.1%) | 18.5 (7.14%) |
| 7.33 (17.2%) 3 | |||||
| 4.50 (14.9%) 4 | |||||
| Hill coefficient for TOB | HILLTOB | 0.790 (17.9%) 5 | |||
| Killing by CIP | |||||
| Maximum killing rate constant | |||||
| TOBS/CIPS | Kmax,SS,CIP (h−1) | 16.4 (9.95%) | 17.1 (14.2%) | 2.43 (11.7%) | 5.11 (13.5%) |
| TOBR/CIPI | Kmax,RI,CIP (h−1) | 0.392 (20.5%) | 0.307 (33.1%) | 0.730 (17.5%) | 1.14 (18.6%) |
| TOBI/CIPR | Kmax,IR,CIP (h−1) | 1.83 (13.6%) | 0.812 (9.26%) | 0.562 (13.4%) | 0.226 (21.4%) |
| CIP concentration causing 50% of Kmax,CIP | KC50,CIP (mg/L) | 1.29 (24.3%) | 1.09 (29.6%) | 7.07 (41.3%) | 8.30 (11.6%) |
| Mechanistic synergy | |||||
| Maximum fractional decrease of KC50,CIP via mechanistic synergy6 | Imax,SYN | 1 (fixed) | 1 (fixed) | 1 (fixed) | 1 (fixed) |
| TOB concentration causing 50% of Imax,SYN | IC50,SYN (mg/L) | 2.16 (11.0%) | 1.10 (33.9%) | 2.01 (10.9%) | 1.74 (20.0%) |
| Residual variability | |||||
| SD of residual error on log10 scale | |||||
| Total population | SDCFU | 0.303 (10.2%) | 0.383 (12.1%) | 0.296 (16.5%) | 0.273 (10.1%) |
| Population on TOB plates | SDCFU,TOB | 1.05 (24.1%) | 0.401 (22.0%) | 0.883 (14.1%) | 0.197 (18.9%) |
| Population on CIP plates | SDCFU,CIP | 3.64 (26.4%) | 1.18 (14.4%) | 0.586 (37.1%) | 0.740 (31.1%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rees, V.E.; Bulitta, J.B.; Oliver, A.; Nation, R.L.; Landersdorfer, C.B. Evaluation of Tobramycin and Ciprofloxacin as a Synergistic Combination Against Hypermutable Pseudomonas Aeruginosa Strains via Mechanism-Based Modelling. Pharmaceutics 2019, 11, 470. https://doi.org/10.3390/pharmaceutics11090470
Rees VE, Bulitta JB, Oliver A, Nation RL, Landersdorfer CB. Evaluation of Tobramycin and Ciprofloxacin as a Synergistic Combination Against Hypermutable Pseudomonas Aeruginosa Strains via Mechanism-Based Modelling. Pharmaceutics. 2019; 11(9):470. https://doi.org/10.3390/pharmaceutics11090470
Chicago/Turabian StyleRees, Vanessa E., Jürgen B. Bulitta, Antonio Oliver, Roger L. Nation, and Cornelia B. Landersdorfer. 2019. "Evaluation of Tobramycin and Ciprofloxacin as a Synergistic Combination Against Hypermutable Pseudomonas Aeruginosa Strains via Mechanism-Based Modelling" Pharmaceutics 11, no. 9: 470. https://doi.org/10.3390/pharmaceutics11090470
APA StyleRees, V. E., Bulitta, J. B., Oliver, A., Nation, R. L., & Landersdorfer, C. B. (2019). Evaluation of Tobramycin and Ciprofloxacin as a Synergistic Combination Against Hypermutable Pseudomonas Aeruginosa Strains via Mechanism-Based Modelling. Pharmaceutics, 11(9), 470. https://doi.org/10.3390/pharmaceutics11090470