
Can Plant Viruses Cross the Kingdom Border and Be Pathogenic to Humans?


Abstract
:1. Introduction
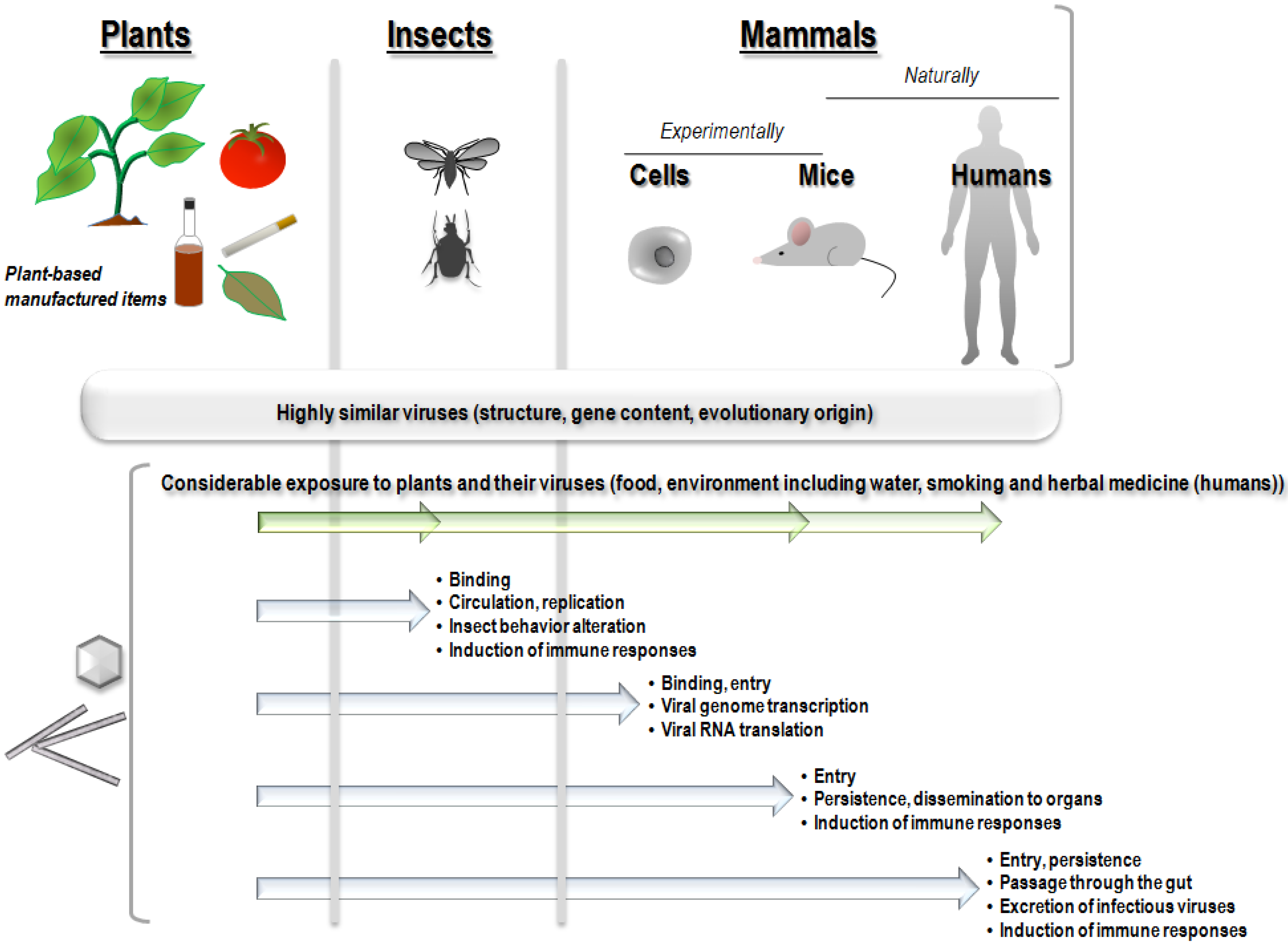
2. The Nature of the Border that Segregates Viruses in Plants or Animals
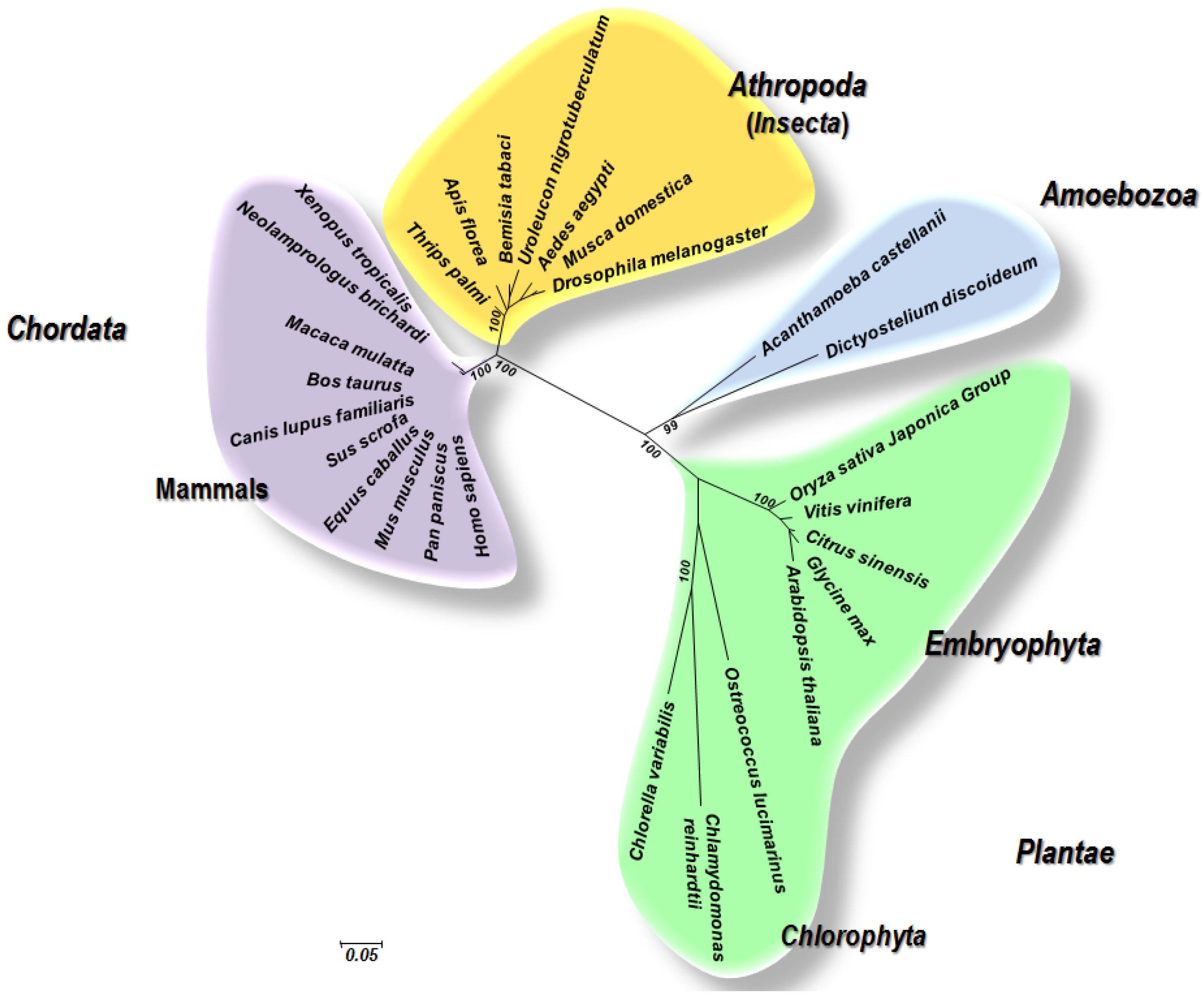
3. Close Relatedness between Plant and Animal Viruses and Evidence for Common Origins

{kind=link}
{kind=link}
{kind=link}
| Genome | Genome Segmentation | Virion Shape | Family | Sub-Family | Genus | Hosts |
|---|---|---|---|---|---|---|
| RNA, single-stranded, positive | Monopartite | Icosahedral | Tymoviridae | Tymovirus | Plants | |
| Monopartite | Maculavirus | Plants | ||||
| Monopartite | Marafivirus | Plants and insects | ||||
| RNA, single-stranded, negative or | Monopartite | Pleomorphic, globular | Bunyaviridae | Hantavirus | Mammals | |
| ambisense | Monopartite | Bunyavirus | Mammals | |||
| Monopartite | Nairovirus | Mammals | ||||
| Monopartite | Phlebovirus | Mammals | ||||
| Tripartite | Tospovirus | Plants and insects | ||||
| RNA, single-stranded negative | Monopartite | bacilliform | Rhabdoviridae | Lyssavirus | Vertebrates | |
| Monopartite | Vesiculovirus | Vertebrates | ||||
| Monopartite | Ephemerovirus | Vertebrates | ||||
| Monopartite | Novirhabdovirus | Vertebrates (fishes) | ||||
| Monopartite | Cytorhabdovirus | Plants and insects | ||||
| Monopartite | Nucleorhabdovirus | Plants and insects | ||||
| Bipartite | Dichorhabdovirus | Plants | ||||
| Multipartite | Flexuous | - | Tenuivirus | Plants and insects | ||
| RNA, double-stranded | Monopartite | icosahedral | Reoviridae | Sedoreovirinae | Cardoreovirus | Crustaceans |
| Monopartite | Mimoreovirus | Photosynthetic marine protists | ||||
| Monopartite | Orbivirus | Arthropod, vertebrates | ||||
| Monopartite | Rotavirus | Vertebrates | ||||
| Monopartite | Seadornavirus | Invertebrates and vertebrates | ||||
| Multipartite | Phytoreovirus | Plants and insects | ||||
| Monopartite | Spinareovirinae | Aquareovirus | Aquatic vertebrates and invertebrates | |||
| Monopartite | Coltivirus | Invertebrates and vertebrates | ||||
| Monopartite | Cypovirus | Insects | ||||
| Monopartite | Dinovernavirus | Insects | ||||
| Monopartite | Idnoreovirus | Insects | ||||
| Monopartite | Mycoreovirus | Fungi | ||||
| Monopartite | Orthoreovirus | Vertebrates | ||||
| Multipartite | Fijivirus | Plants and insects | ||||
| Multipartite | Oryzavirus | Plants and insects |
4. Exposure of Non-Human Animals and Humans to Plant Viruses
5. Evidence of the Natural Presence and Persistence of Plant Viruses in Non-Human Mammals
| Nucleic Acid | Family | Genus | Species | Vertebrate Cells | Mammals | Humans |
|---|---|---|---|---|---|---|
| ssRNA+ | Alphaflexiviridae | N.a. | Mouse, vole and rat stools [85] | |||
| ssRNA+ | Bromoviridae | Ilarvirus | Prunus necrotic ringspot virus | Stools [86] | ||
| ssRNA+ | Closteroviridae | Closterovirus | Citrus tristeza virus | Stools [87] | ||
| ssRNA+ | Luteoviridae | N.a. | N.a. | Bat guano [88] | ||
| N.a. | N.a. | Mouse, vole and rat stools [85] | ||||
| ssRNA+ | Secoviridae | Comovirus | Cowpea mosaic virus | Human Hela cells [44] | Mice [84] | |
| Comovirus | Cowpea mosaic virus | Human huvec cells [44] | ||||
| Comovirus | Cowpea mosaic virus | Human KB cells [44] | ||||
| N.a. | N.a. | Bat guano [88] | ||||
| N.a. | N.a. | Mouse, vole and rat stools [85] | ||||
| ssRNA+ | Sobemovirus | Rice yellow mottle virus | Cows, donkeys and grass rats [89] | |||
| Sobemovirus | Subterranean clover mottle virus | Sheep [90] | ||||
| Sobemovirus | N.a | Bat guano [88] | ||||
| Sobemovirus | Cocksfoot mottle virus | Stools [86] | ||||
| ssRNA+ | Tombusviridae | Tombusvirus | Tomato bushy stunt virus | Stools [60] | ||
| Panicovirus | Panicum mosaic virus | Stools [86] | ||||
| Carmovirus | Melon necrotic spot virus | Stools [86] | ||||
| Carmovirus | Galinsoga mosaic virus | Stools [86] | ||||
| Carmovirus | Carnation mottle virus | Stools [86] | ||||
| Necrovirus | Tobacco necrosis virus | Stools [86] | ||||
| Necrovirus | Olive latent virus 1 | Stools [86] | ||||
| Aureusvirus | Pothos latent virus | Stools [86] | ||||
| Aureusvirus | Johnsongrass chlorotic stripe mosaic virus | Stools [86] | ||||
| Avenavirus | Oat chlorotic stunt virus | Stools [86] | ||||
| Machlomovirus | Maize chlorotic mottle virus | Stools [86] | ||||
| N.a. | N.a. | Mouse, vole and rat stools [85] | ||||
| ssRNA+ | Tymoviridae | Marafivirus | Grapevine asteroid mosaic-associated virus | Stools [86] | ||
| Marafivirus | Maize rayado fino virus | Stools [86] | ||||
| Marafivirus | Grapevine rupestris vein feathering virus | Stools [86] | ||||
| Marafivirus | Oat blue dwarf virus | Stools [86] | ||||
| Maculavirus | Grapevine fleck virus | Stools [86] | ||||
| Maculavirus | Grapevine red globe virus | Stools [86] | ||||
| Tymovirus | Chayote mosaic tymovirus | Stools [86] | ||||
| Tymovirus | Kennedya yellow mosaic virus | Stools [86] | ||||
| Tymovirus | Physalis mottle virus | Stools [86] | ||||
| Tymovirus | Poinsettia mosaic virus | Stools [86] | ||||
| Tymovirus | Eggplant mosaic virus | Stools [86] | ||||
| Tymovirus | Ononis yellow mosaic virus | Stools [86] | ||||
| N.a. | N.a. | Bat guano [88] | ||||
| N.a. | N.a. | Mouse, vole and rat stools [85] | ||||
| ssRNA+ | Virgariridae | Tobamovirus | Tobacco mosaic virus | Hela cells [91] | Mice [92] | Stools [86,87] |
| Mice splenocyte [93] | Skin [94] | |||||
| Mice bone marrow cells [92] | Thoracentesis fluids [95] | |||||
| Saliva [69] | ||||||
| Tobamovirus | Pepper mild mottle Virus | Stools [86,87] | ||||
| Tobamovirus | Cucumber green mottle mosaic virus | Cows stools [96] | ||||
| Tobamovirus | Tomato mosaic virus | Stools [86] | ||||
| Tobamovirus | Ribgrass mosaic virus | Stools [86] | ||||
| Tobamovirus | Turnip vein-clearing virus | Stools [86] | ||||
| Tobamovirus | Tobacco mild green mosaic virus | Stools [86] | ||||
| Tobamovirus | Odontoglossum ringspot virus | Stools [86] | ||||
| Tobamovirus | Paprika mild mottle virus | Stools [86] | ||||
| Tobamovirus | Kyuri green mottle mosaic virus | Stools [87] | ||||
| Tobamovirus | Crucifer tobamovirus | Stools [87] | ||||
| Tobamovirus | Obuda pepper virus | Stools [86] | ||||
| Tobamovirus | Nigerian tobacco latent virus | Stools [86] | ||||
| dsRNA | Partitiviridae | N.a. | N.a. | Bat guano [88] | ||
| N.a. | N.a. | Mouse, vole and rat stools [85] | ||||
| dsDNA | Phycodnaviridae | Chlorovirus | Acanthocystis turfacea chlorella virus 1 | Mouse [72] | Oropharynx samples [72] | |
| ssDNA | Nanoviridae | N.a. | N.a. | Mouse, vole and rat stools [85] | ||
| ssDNA | Geminiviridae | N.a. | N.a. | Mouse, vole and rat stools [85] |
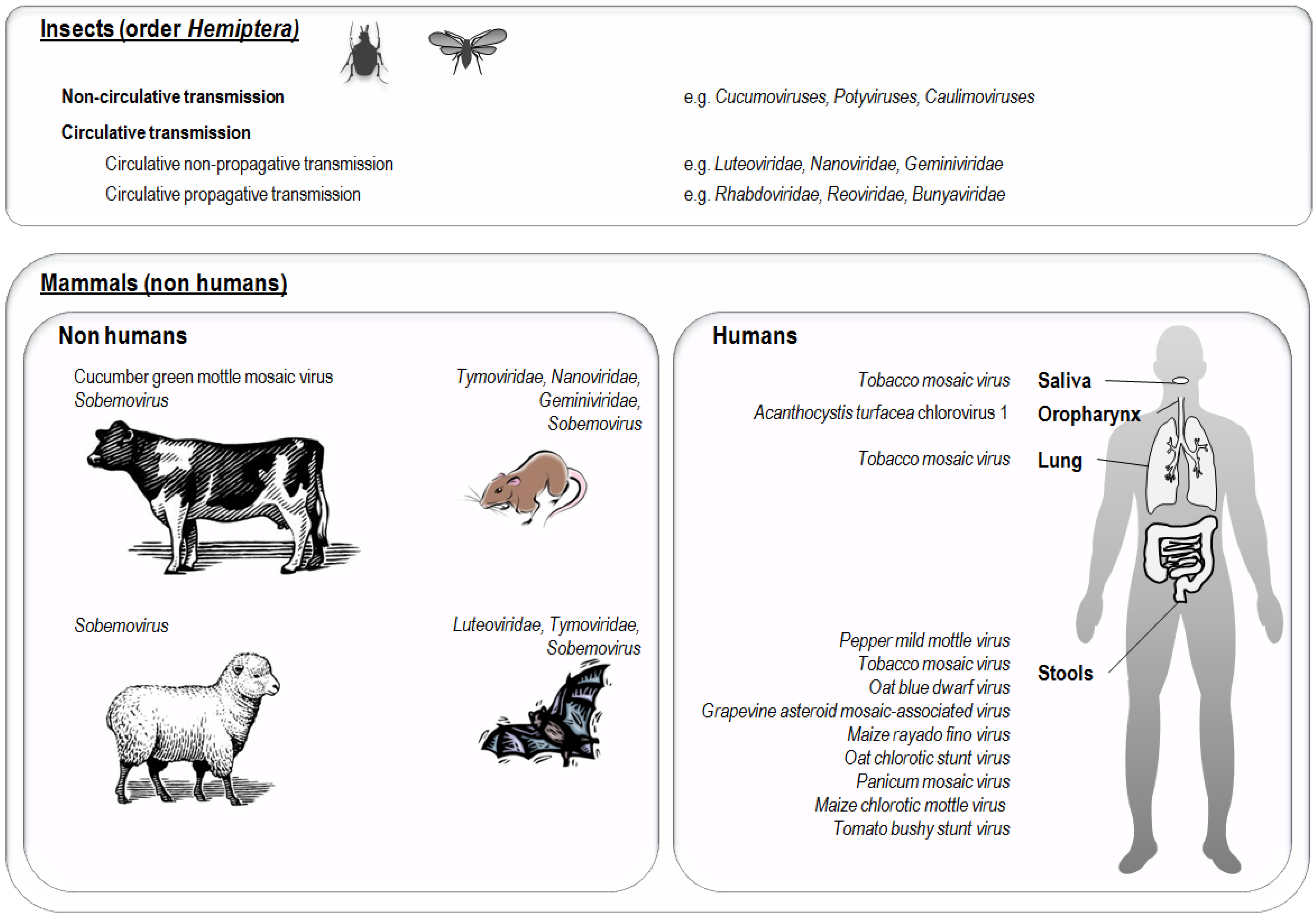
6. Presence of Plant Viruses in Human Samples
7. Immune Responses to Plant Viruses in Invertebrates, Non-Human Vertebrates and Humans
8. Evidence of the Entry of Plant Viruses or Their Genomes into Non-Human Mammal Cells and Bodies after Experimental Exposure
9. Conclusions
Author Contributions
Conflicts of Interest
References
- Soosaar, J.L.; Burch-Smith, T.M.; Dinesh-Kumar, S.P. Mechanisms of plant resistance to viruses. Nat. Rev. Microbiol. 2005, 3, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Ivanovski, D. Concerning the mosaic disease of tobacco plant. St. Petersb. Acad. Imp. Sci. Bul. 1892, 67–70. [Google Scholar]
- Beijerinck, M.W. Concerning a contagium vivum fluidum as a cause of the spot-disease of tobacco leaves. Verh. Akad. Wet. Amster. 1898, 6, 33–55. [Google Scholar]
- International Committee on Taxonomy of Viruses. Available online: http://ictvonline.org/virustaxonomy.asp (accessed on 20 March 2015).
- Wren, J.D.; Roossinck, M.J.; Nelson, R.S.; Scheets, K.; Palmer, M.W.; Melcher, U. Plant virus biodiversity and ecology. PLoS Biol. 2006, 4, e80. [Google Scholar] [CrossRef] [PubMed]
- Giampetruzzi, A.; Roumi, V.; Roberto, R.; Malossini, U.; Yoshikawa, N.; la, N.P.; Terlizzi, F.; Credi, R.; Saldarelli, P. A new grapevine virus discovered by deep sequencing of virus- and viroid-derived small RNAs in Cv Pinot gris. Virus Res. 2012, 163, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Forterre, P. Redefining viruses: Lessons from Mimivirus. Nat. Rev. Microbiol. 2008, 6, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Desnues, C.; Boyer, M.; Raoult, D. Sputnik, a virophage infecting the viral domain of life. Adv. Virus Res. 2012, 82, 63–89. [Google Scholar] [PubMed]
- Desnues, C.; Raoult, D. Virophages question the existence of satellites. Nat. Rev. Microbiol. 2012, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Ammar, E.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef] [PubMed]
- Keller, M. Les virus de plantes et les virus des animaux: Deux mondes totalement différents? Virologie 2004, 8, 251–257. [Google Scholar]
- Raccah, B. Fereres A: Plant Virus Transmission by Insects; John Wiley & Sons, Ltd.: Chichester, UK, 2009. [Google Scholar]
- Klempner, M.S.; Shapiro, D.S. Crossing the species barrier--one small step to man, one giant leap to mankind. N. Engl. J. Med. 2004, 350, 1171–1172. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Helenius, A. Virus entry at a glance. J. Cell Sci. 2013, 126, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Lazarowitz, S.D. Plant viruses. In Fields Virology, 5th ed.; Fields, B.N., Knipe, D.M., Eds.; Lippincott, Williams & Wilkins Publishers: Philadelphia, PA, USA, 2006; pp. 641–706. [Google Scholar]
- Taliansky, M.; Torrance, L.; Kalinina, N.O. Role of plant virus movement proteins. Methods Mol. Biol. 2008, 451, 33–54. [Google Scholar] [PubMed]
- Deom, C.M.; Lapidot, M.; Beachy, R.N. Plant virus movement proteins. Cell 1992, 69, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Alfonso, Y.; Faulkner, C.; Ritzenthaler, C.; Maule, A.J. Plasmodesmata: Gateways to local and systemic virus infection. Mol. Plant Microbe Interact. 2010, 23, 1403–1412. [Google Scholar] [CrossRef] [PubMed]
- Hohn, T. Plant virus transmission from the insect point of view. Proc. Natl. Acad. Sci. USA 2007, 104, 17905–17906. [Google Scholar] [CrossRef] [PubMed]
- Gaedigk, K.; Adam, G.; Mundry, K.W. The spike protein of Potato yellow dwarf virus and its functional role in the infection of insect vector cells. J. Gen. Virol. 1986, 67, 2763–2773. [Google Scholar] [CrossRef]
- Tomaru, M.; Maruyama, W.; Kikuchi, A.; Yan, J.; Zhu, Y.; Suzuki, N.; Isogai, M.; Oguma, Y.; Kimura, I.; Omura, T. The loss of outer capsid protein P2 results in nontransmissibility by the insect vector of rice dwarf phytoreovirus. J. Virol. 1997, 71, 8019–8023. [Google Scholar] [PubMed]
- Hebrard, E.; Froissart, R.; Louis, C.; Blanc, S. Les modes de transmission des virus phytopathogènes par vecteurs. Virologie 1999, 3, 25–48. [Google Scholar]
- Perry, K.L.; Zhang, L.; Palukaitis, P. Amino acid changes in the coat protein of cucumber mosaic virus differentially affect transmission by the aphids Myzus persicae and Aphis gossypii. Virology 1998, 242, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Pendon, J.A.; Canizares, M.C.; Moriones, E.; Bejarano, E.R.; Czosnek, H.; Navas-Castillo, J. Tomato yellow leaf curl viruses: Menage a trois between the virus complex, the plant and the whitefly vector. Mol. Plant Pathol. 2010, 11, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, M; Morin, S.; Zeidan, M.; Czosneck, H. Evidence for transovarial transmission of tomato yellow leaf curl virus by its vector, the whitefly Bemisia tabaci. Virology 1998, 240, 295–303. [Google Scholar] [CrossRef]
- Bosco, D.; Mason, G.; Accotto, G.P. TYLCSV DNA, but not infectivity, can be transovarially inherited by the progeny of the whitefly vector Bemisia tabaci (Gennadius). Virology 2004, 323, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Sylvester, E.S. Circulative and propagative virus transmission by aphids. Ann. Rev. Entomol. 1980, 25, 257–286. [Google Scholar] [CrossRef]
- Stafford, C.A.; Walker, G.P.; Ullman, D.E. Infection with a plant virus modifies vector feeding behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 9350–9355. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, G.; Czosnek, H. Long-term association of tomato yellow leaf curl virus with its whitefly vector Bemisia tabaci: Effect on the insect transmission capacity, longevity and fecundity. J. Gen. Virol. 1997, 78, 2683–2689. [Google Scholar] [PubMed]
- Li, J.L.; Cornman, R.S.; Evans, J.D.; Pettis, J.S.; Zhao, Y.; Murphy, C.; Peng, W.J.; Wu, J.; Hamilton, M.; Boncristiani, H.F., Jr.; et al. Systemic Spread and Propagation of a Plant-Pathogenic Virus in European Honeybees, Apis mellifera. MBio 2014, 51, e00898–13. [Google Scholar]
- Koonin, E.V. The origin and early evolution of eukaryotes in the light of phylogenomics. Genome Biol. 2010, 11, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Roger, A.J.; Simpson, A.G. Evolution: Revisiting the root of the eukaryote tree. Curr. Biol. 2009, 19, R165–R167. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2015, 43, D30–D35. [Google Scholar] [CrossRef] [PubMed]
- Maroniche, G.A.; Mongelli, V.C.; Peralta, A.V.; Distefano, A.J.; Llauger, G.; Taboga, O.A.; Hopp, E.H.; del, V.M. Functional and biochemical properties of Mal de Rio Cuarto virus (Fijivirus, Reoviridae) P9-1 viroplasm protein show further similarities to animal reovirus counterparts. Virus Res. 2010, 152, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Choi, U.Y.; Lee, S.Y.; Ma, S.H.; Jang, Y.T.; Kim, J.Y.; Kim, H.M.; Kim, J.H.; Kim, D.S.; Kim, Y.S.; Kang, J.H. Epidemiological changes in rotavirus gastroenteritis in children under 5 years of age after the introduction of rotavirus vaccines in Korea. Eur. J. Pediatr. 2013, 172, 947–952. [Google Scholar] [CrossRef] [PubMed]
- Tordo, N.; Ceccaldi, P.-E.; Gaudin, Y.; Wandeler, A.I. Rhabdoviruses: Rabies. In Topley and Wilson’s Microbiology and Microbial Infections; Mahy, B.W.J., Collier, L., Eds.; 2010; pp. 665–692. [Google Scholar]
- Kallio-Kokko, H.; Laakkonen, J.; Rizzoli, A.; Tagliapietra, V.; Cattadori, I.; Perkins, S.E.; Hudson, P.J.; Cristofolini, A.; Versini, W.; Vapalahti, O.; et al. Hantavirus and arenavirus antibody prevalence in rodents and humans in Trentino, Northern Italy. Epidemiol. Infect. 2006, 134, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Sonderegger, B.; Hachler, H.; Dobler, G.; Frei, M. Imported aseptic meningitis due to Toscana virus acquired on the island of Elba, Italy, August 2008. Euro. Surveill. 2009, 14, e19079. [Google Scholar]
- Charrel, R.N.; Gallian, P.; Navarro-Mari, J.M.; Nicoletti, L.; Papa, A.; Sanchez-Seco, M.P.; Tenorio, A.; de Lamballerie, X. Emergence of Toscana virus in Europe. Emerg. Infect. Dis. 2005, 11, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- Niehl, A.; Heinlein, M. Cellular pathways for viral transport through plasmodesmata. Protoplasma 2011, 248, 75–99. [Google Scholar] [CrossRef] [PubMed]
- Mead, D.G.; Gray, E.W.; Noblet, R.; Murphy, M.D.; Howerth, E.W.; Stallknecht, D.E. Biological transmission of vesicular stomatitis virus (New Jersey serotype) by Simulium vittatum (Diptera: Simuliidae) to domestic swine (Sus scrofa). J. Med. Entomol. 2004, 41, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Clark, A.J.; Chen, Z.; Shanks, M.; Dai, J.B.; Li, Y.; Schmidt, T.; Oxelfelt, P.; Lomonossoff, G.P.; Johnson, J.E. Structural fingerprinting: Subgrouping of comoviruses by structural studies of red clover mottle virus to 2.4-A resolution and comparisons with other comoviruses. J. Virol. 2000, 74, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Koudelka, K.J.; Destito, G.; Plummer, E.M.; Trauger, S.A.; Siuzdak, G.; Manchester, M. Endothelial targeting of cowpea mosaic virus (CPMV) via surface vimentin. PLoS Pathog. 2009, 5, e1000417. [Google Scholar] [CrossRef] [PubMed]
- Geering, A.D.; Scharaschkin, T.; Teycheney, P.Y. The classification and nomenclature of endogenous viruses of the family Caulimoviridae. Arch. Virol. 2010, 155, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V. The phylogeny of RNA-dependent RNA polymerases of positive-strand RNA viruses. J. Gen. Virol. 1991, 72, 2197–2206. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Holmes, E.C. Endogenous RNA viruses of plants in insect genomes. Virology 2012, 427, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Dolja, V.V.; Koonin, E.V. Common origins and host-dependent diversity of plant and animal viromes. Curr. Opin. Virol. 2011, 1, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.K.; Arnold, J.J.; Cameron, C.E. Structure-function relationships among RNA-dependent RNA polymerases. Curr. Top. Microbiol. Immunol. 2008, 320, 137–156. [Google Scholar] [PubMed]
- Novick, R.P. Contrasting lifestyles of rolling-circle phages and plasmids. Trends Biochem. Sci. 1998, 23, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Ilyina, T.V.; Koonin, E.V. Conserved sequence motifs in the initiator proteins for rolling circle DNA replication encoded by diverse replicons from eubacteria, eucaryotes and archaebacteria. Nucleic Acids Res. 1992, 20, 3279–3285. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, M.J.; Weiller, G.F. Evidence that a plant virus switched hosts to infect a vertebrate and then recombined with a vertebrate-infect ing virus. Proc. Natl. Acad. Sci. USA 1999, 96, 8022–8027. [Google Scholar] [CrossRef] [PubMed]
- Davidson, I.; Silva, R.F. Creation of diversity in the animal virus world by inter-species and intra-species recombinations: Lessons learned from poultry viruses. Virus Genes 2008, 36, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Hernandez, C.; Martinez de Alba, A.E.; Daros, J.A.; di, S.F. Viroids and viroid-host interactions. Annu. Rev. Phytopathol. 2005, 43, 117–139. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Ruiz-Ruiz, S.; Serra, P. Viroids and hepatitis delta virus. Semin. Liver Dis. 2012, 32, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Daros, J.A.; Elena, S.F.; Flores, R. Viroids: An Ariadne’s thread into the RNA labyrinth. EMBO Rep. 2006, 7, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Brazas, R.; Ganem, D. A cellular homolog of hepatitis delta antigen: Implications for viral replication and evolution. Science 1996, 274, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Robertson, H.D. How did replicating and coding RNAs first get together? Science 1996, 274, 66–67. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Niu, Y.; Zhang, K.; Liu, Y.; Zhou, X. Virus-derived transgenes expressing hairpin RNA give immunity to Tobacco mosaic virus and Cucumber mosaic virus. Virol. J. 2011, 8, e41. [Google Scholar] [CrossRef]
- Tomlinson, J.A.; Faithfull, E.; Flewett, T.H.; Beards, G. Isolation of infective tomato bushy stunt virus after passage through the human alimentary tract. Nature 1982, 300, 637–638. [Google Scholar] [CrossRef] [PubMed]
- Hollings, M.; Stone, O.M. Studies of pelargonium leaf curl virus. Ann. Appl. Biol. 1965, 56, 87–98. [Google Scholar] [CrossRef]
- Colson, P.; Richet, H.; Desnues, C.; Balique, F.; Moal, V.; Grob, J.J.; Berbis, P.; Lecoq, H.; Harle, J.R.; Berland, Y.; et al. Pepper mild mottle virus, a plant virus associated with specific immune responses, Fever, abdominal pains, and pruritus in humans. PLoS One 2010, 5, e10041. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization media centre, fact sheets. Available online: http://www.who.int/mediacentre/factsheets/2003/fs134/en/ (accessed on 20 March 2015).
- Bothwell, P.W. Lung cancer and tobacco mosaic virus. Lancet 1960, 1, 657–658. [Google Scholar] [CrossRef]
- Wahyuni, W.S.; Hanapi, M.; Hartana, I. The Presence of tobacco mosaic virus in the compost extract of Cigar Tobacco Debris. J. Biosci. 2008, 15, 118–22. [Google Scholar]
- Wetter, C. Tobacco mosaic virus and para-tobacco mosaic virus in cigarettes. Naturwissenschaften 1975, 62, 533. [Google Scholar] [CrossRef] [PubMed]
- Chyle, P.; Chyle, M.; Korb, J.; Papanek, M. Virus tabakove mozaiky, polyfenoly a kancerogenita tabakoveho navyku. Cesk Epidemiol. Mikrobiol. Imunol. 1971, 20, 32–42. [Google Scholar] [PubMed]
- Smith, K.M. A Textbook of Plant Virus Diseases; Elsevier; London, UK, 1957; p. 507. [Google Scholar]
- Balique, F.; Colson, P.; Raoult, D. Tobacco mosaic virus in cigarettes and saliva of smokers. J. Clin. Virol. 2012, 55, 374–376. [Google Scholar] [CrossRef] [PubMed]
- Rosario, K.; Nilsson, C.; Lim, Y.W.; Ruan, Y.; Breitbart, M. Metagenomic analysis of viruses in reclaimed water. Environ. Microbiol. 2009, 11, 2806–2820. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, O.A.; Solovyova, Y.V.; Solovyov, A.V. Results of algae viruses search in human clinical material. Ukrainica Bioorganica Acta 2011, 2, 53–56. [Google Scholar]
- Yolken, R.H.; Jones-Brando, L.; Dunigan, D.D.; Kannan, G.; Dickerson, F.; Severance, E.; Sabunciyan, S.; Talbot, C.C., Jr.; Prandovszky, E.; Gurnon, J.R.; et al. Chlorovirus ATCV-1 is part of the human oropharyngeal virome and is associated with changes in cognitive functions in humans and mice. Proc. Natl. Acad. Sci. USA 2014, 111, 16106–16111. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, V.; Castello, J. Isolation of tomato mosaic virus from waters draining forest stands in New york state. Phytopathology 1991, 81, 1112–1117. [Google Scholar] [CrossRef]
- Castello, J.D.; Rogers, S.O.; Starmer, W.T.; Catranis, C.M.; Ma, L.; Bachand, G.D.; Zhao, Y.; Smith, J.E. Detection of tomato mosaic tobamovirus RNA in ancient glacial ice. Polar biol. 1999, 22, 207–212. [Google Scholar] [CrossRef]
- Castello, J.D.; Lakhsman, D.K.; Tavantzis, S.M.; Rogers, S.O.; Bachand, G.D; Jagels, R.J.; Carlisle, J.; Liu, Y. Detection of infectious tomato mosaic tobamovirus in fog and clouds. Phytopathology 1995, 85, 1409–1412. [Google Scholar] [CrossRef]
- Rosario, K.; Symonds, E.M.; Sinigalliano, C.; Stewart, J.; Breitbart, M. Pepper mild mottle virus as an indicator of fecal pollution. Appl. Environ. Microbiol. 2009, 75, 7261–7267. [Google Scholar] [CrossRef] [PubMed]
- Hamza, I.A.; Jurzik, L.; Uberla, K.; Wilhelm, M. Methods to detect infectious human enteric viruses in environmental water samples. Int. J. Hyg. Environ. Health 2011, 214, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Haramoto, E.; Kitajima, M.; Kishida, N.; Konno, Y.; Katayama, H.; Asami, M.; Akiba, M. Occurrence of pepper mild mottle virus in drinking water sources in Japan. Appl. Environ. Microbiol. 2013, 79, 7413–7418. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, J.A.; Faithfull, E. Studies on the occurrence of tomato bushy stunt virus in English rivers. Ann. Appl. Biol. 1984, 104, 485–495. [Google Scholar] [CrossRef]
- Kramberger, P.; Peterka, M.; Boben, J.; Ravnikar, M.; Strancar, A. Short monolithic columns–A breakthrough in purification and fast quantification of tomato mosaic virus. J. Chromatogr. A 2007, 1144, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Gosalvez, B.; Navarro, J.A.; Lorca, A.; Botella, F.; Sanchez-Pina, M.A.; Pallas, V. Detection of melon necrotic spot virus in water samples and melon plants by molecular methods. J. Virol. Methods 2003, 113, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Vani, S.; Varma, A. Properties of cucumber green mottle mosaic virus isolated from water of river Yamuna. Indian Phytopathol. 1993, 46, 118–122. [Google Scholar]
- Xu, F.; Jones, T.D.; Rodgers, P.B. Potential of chimaeric plant virus particles as novel, stable vaccines. Dev. Biol. Stand. 1996, 87, 201–205. [Google Scholar] [PubMed]
- Rae, C.S.; Khor, I.W.; Wang, Q.; Destito, G.; Gonzalez, M.J.; Singh, P.; Thomas, D.M.; Estrada, M.N.; Powell, E.; Finn, M.G.; et al. Systemic trafficking of plant virus nanoparticles in mice via the oral route. Virology 2005, 343, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Kapusinszky, B.; Wang, C.; Rose, R.K.; Lipton, H.L.; Delwart, E.L. The fecal viral flora of wild rodents. PLoS Pathog. 2011, 7, e1002218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Breitbart, M.; Lee, W.H.; Run, J.Q.; Wei, C.L.; Soh, S.W.; Hibberd, M.L.; Liu, E.T.; Rohwer, F.; Ruan, Y. RNA viral community in human feces: Prevalence of plant pathogenic viruses. PLoS Biol. 2006, 4, e3. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Yang, C.S.; Sakon, N.; Ueda, M.; Tougan, T.; Yamashita, A.; Goto, N.; Takahashi, K.; Yasunaga, T.; Ikuta, K.; et al. Direct metagenomic detection of viral pathogens in nasal and fecal specimens using an unbiased high-throughput sequencing approach. PLoS One 2009, 4, e4219. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Victoria, J.G.; Wang, C.; Jones, M.; Fellers, G.M.; Kunz, T.H.; Delwart, E. Bat guano virome: Predominance of dietary viruses from insects and plants plus novel mammalian viruses. J. Virol. 2010, 84, 6955–6965. [Google Scholar] [CrossRef] [PubMed]
- Sarra, S.; Peters, D. Rice yellow mottle virus is transmitted by cows, donkeys, and grass Rats in irrigated rice crops. Plant Dis. 2003, 87, 804–808. [Google Scholar]
- MCkirdy, S.J.; Jones, R.A.C.; Sivasithamparam, K. Dertermining the effectiveness of grazing and trampling by livestock in transmitting white clover mosaic and subterranean clover mottle virus. Ann. Appl. Biol. 1998, 132, 91–105. [Google Scholar] [CrossRef]
- Li, L.; Wang, L.; Xiao, R.; Zhu, G.; Li, Y.; Liu, C.; Yang, R.; Tang, Z.; Li, J.; Huang, W.; et al. The invasion of tobacco mosaic virus RNA induces endoplasmic reticulum stress-related autophagy in HeLa cells. Biosci. Rep. 2012, 32, 171–186. [Google Scholar] [CrossRef] [PubMed]
- Balique, F.; Colson, P.; Barry, A.O.; Nappez, C.; Ferretti, A.; Moussawi, K.A.; Ngounga, T.; Lepidi, H.; Ghigo, E.; Mege, J.L.; et al. Tobacco Mosaic Virus in the Lungs of Mice following Intra-Tracheal Inoculation. PLoS One 2013, 8, e54993. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.A.; Corbo, T.A.; Wykoff-Clary, S.; Palmer, K.E.; Pogue, G.P. Chemical conjugate TMV-peptide bivalent fusion vaccines improve cellular immunity and tumor protection. Bioconjug. Chem. 2006, 17, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Katsilambros, L. Tobacco mosaic virus and lung cancer. Lancet 1960, 2, 934. [Google Scholar] [CrossRef]
- LeClair, R.A. Recovery of culturable tobacco mosaic virus from sputum and thoracentesis fluids obtained from cigarette smokers with a history of pulmonary disease. Am. Rev. Respir. Dis. 1967, 95, 510–511. [Google Scholar] [PubMed]
- Van Dorst, H.J.M. Surface water as source in the spread of cucumber green mottle mosaic virus. Neth. J. Agric. Sci. 1988, 36, 291–299. [Google Scholar]
- Hamza, I.A.; Jurzik, L.; Uberla, K.; Wilhelm, M. Evaluation of pepper mild mottle virus, human picobirnavirus and Torque teno virus as indicators of fecal contamination in river water. Water Res. 2011, 45, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Bothwell, P.W. Lung cancer and tobacco mosaic virus. Lancet 1960, 1, 923–924. [Google Scholar] [CrossRef]
- Pirié, N.W. Lung cancer and tobacco mosaic virus. Lancet 1960, 1, 707. [Google Scholar] [CrossRef]
- Pirié, N.W. Lung cancer and tobacco mosaic virus. Lancet 1960, 1, 978–979. [Google Scholar] [CrossRef]
- Carpenter, C.M.; le Clair, R. Tobacco mosaic virus and pulmonary diseases of humans. Dis. Chest 1968, 54, 42–43. [Google Scholar] [CrossRef] [PubMed]
- Falk, H.L. Chemical agents in cigarette smoke. In Handbook of Physiology; Lee, D.H.K., Ed.; 2011; pp. 199–211. [Google Scholar]
- Chyle, P.; Chyle, M.; Patocka, F. Virus tabakove mozaiky a polyfenoly jako priorozene se vyskytujici kancerogeny v Tabaku. Casopis Lekaru Ceskych 1971, 110, 189–190. [Google Scholar] [PubMed]
- Bousbia, S.; Papazian, L.; La, S.B.; Raoult, D. Detection of plant DNA in the bronchoalveolar lavage of patients with ventilator-associated pneumonia. PLoS One 2010, 5, e11298. [Google Scholar] [CrossRef] [PubMed]
- Victoria, J.G.; Kapoor, A.; Li, L.; Blinkova, O.; Slikas, B.; Wang, C.; Naeem, A.; Zaidi, S.; Delwart, E. Metagenomic analyses of viruses in stool samples from children with acute flaccid paralysis. J. Virol. 2009, 83, 4642–4651. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, R.B.; Resende, R.O.; de Avila, A.C. The plant virus Tomato Spotted Wilt Tospovirus activates the immune system of its main insect vector, Frankliniella occidentalis. J. Virol. 2004, 78, 4976–4982. [Google Scholar] [CrossRef] [PubMed]
- Florindo, M.I.; de Aragao, M.E.; da Silva, A.C.; Otoch, M.L.; Melo, D.F.; Lima, J.A.; Lima, M.G. Immune response induced in mice oral immunization with cowpea severe mosaic virus. Braz. J. Med Biol. Res. 2002, 35, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Friedland, R.P.; Tedesco, J.M.; Wilson, A.C.; Atwood, C.S.; Smith, M.A.; Perry, G.; Zagorski, M.G. Antibodies to potato virus Y bind the amyloid beta peptide: Immunohistochemical and NMR studies. J. Biol. Chem. 2008, 283, 22550–22556. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Vaishnav, R.A.; Roberts, A.M.; Friedland, R.P. Humans have antibodies against a plant virus: Evidence from tobacco mosaic virus. PLoS One 2013, 8, e60621. [Google Scholar] [CrossRef] [PubMed]
- Knowland, J. Protein synthesis directed by the RNA from a plant virus in a normal animal cell. Genetics 1974, 78, 383–394. [Google Scholar] [PubMed]
- Nedellec, P.; Vicart, P.; Laurent-Winter, C.; Martinat, C.; Prevost, M.C.; Brahic, M. Interaction of Theiler’s virus with intermediate filaments of infected cells. J. Virol. 1998, 72, 9553–9560. [Google Scholar] [PubMed]
- Kim, J.K.; Fahad, A.M.; Shanmukhappa, K.; Kapil, S. Defining the cellular target(s) of porcine reproductive and respiratory syndrome virus blocking monoclonal antibody 7G10. J. Virol. 2006, 80, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Albarino, C.G.; Price, B.D.; Eckerle, L.D.; Ball, L.A. Characterization and template properties of RNA dimers generated during flock house virus RNA replication. Virology 2001, 289, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, R.; Cheng, L.L.; Bartholomay, L.C.; Christensen, B.M. Flock house virus replicates and expresses green fluorescent protein in mosquitoes. J. Gen. Virol. 2003, 84, 1789–1797. [Google Scholar] [CrossRef] [PubMed]
- Price, B.D.; Eckerle, L.D.; Ball, L.A.; Johnson, K.L. Nodamura virus RNA replication in Saccharomyces cerevisiae: Heterologous gene expression allows replication-dependent colony formation. J. Virol. 2005, 79, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Ball, L.A. Requirements for the self-directed replication of flock house virus RNA 1. J. Virol. 1995, 69, 720–727. [Google Scholar] [PubMed]
- Gallagher, T.M.; Rueckert, R.R. Assembly-dependent maturation cleavage in provirions of a small icosahedral insect ribovirus. J. Virol. 1988, 62, 3399–3406. [Google Scholar] [PubMed]
- Miller, D.J.; Schwartz, M.D.; Ahlquist, P. Flock house virus RNA replicates on outer mitochondrial membranes in Drosophila cells. J. Virol. 2001, 75, 11664–11676. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.L.; Ball, L.A. Replication of flock house virus RNAs from primary transcripts made in cells by RNA polymerase II. J. Virol. 1997, 71, 3323–3327. [Google Scholar] [PubMed]
- De Medeiros, R.B.; Figueiredo, J.; Resende, R.O.; de Avila, A.C. Expression of a viral polymerase-bound host factor turns human cell lines permissive to a plant- and insect-infecting virus. Proc. Natl. Acad. Sci. USA 2005, 102, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Yusibov, V.; Shivprasad, S.; Turpen, T.H.; Dawson, W.; Koprowski, H. Plant viral vectors based on tobamoviruses. Curr. Top. Microbiol. Immunol. 1999, 240, 81–94. [Google Scholar] [PubMed]
- Gilleland, H.E.; Gilleland, L.B.; Staczek, J.; Harty, R.N.; Garcia-Sastre, A.; Palese, P.; Brennan, F.R.; Hamilton, W.D.; Bendahmane, M.; Beachy, R.N. Chimeric animal and plant viruses expressing epitopes of outer membrane protein F as a combined vaccine against Pseudomonas aeruginosa lung infection. FEMS Immunol. Med. Microbiol. 2000, 27, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Marusic, C.; Rizza, P.; Lattanzi, L.; Mancini, C.; Spada, M.; Belardelli, F.; Benvenuto, E.; Capone, I. Chimeric plant virus particles as immunogens for inducing murine and human immune responses against human immunodeficiency virus type 1. J. Virol. 2001, 75, 8434–8439. [Google Scholar] [CrossRef] [PubMed]
- Nuzzaci, M.; Vitti, A.; Condelli, V.; Lanorte, M.T.; Tortorella, C.; Boscia, D.; Piazzolla, P.; Piazzolla, G. In vitro stability of Cucumber mosaic virus nanoparticles carrying a Hepatitis C virus-derived epitope under simulated gastrointestinal conditions and in vivo efficacy of an edible vaccine. J. Virol. Methods 2010, 165, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Savard, C.; Guerin, A.; Drouin, K.; Bolduc, M.; Laliberte-Gagne, M.E.; Dumas, M.C.; Majeau, N.; Leclerc, D. Improvement of the trivalent inactivated flu vaccine using PapMV nanoparticles. PLoS One 2011, 6, e21522. [Google Scholar] [CrossRef] [PubMed]
- Brennan, F.R.; Gilleland, L.B.; Staczek, J.; Bendig, M.M.; Hamilton, W.D.; Gilleland, H.E., Jr. A chimaeric plant virus vaccine protects mice against a bacterial infection. Microbiology 1999, 145, 2061–2067. [Google Scholar] [CrossRef] [PubMed]
- Porta, C.; Lomonossoff, G.P. Scope for using plant viruses to present epitopes from animal pathogens. Rev. Med. Virol. 1998, 8, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Lacasse, P.; Denis, J.; Lapointe, R.; Leclerc, D.; Lamarre, A. Novel plant virus-based vaccine induces protective cytotoxic T-lymphocyte-mediated antiviral immunity through dendritic cell maturation. J. Virol. 2008, 82, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Rebolledo-Mendez, J.D.; Vaishnav, R.A.; Cooper, N.G.; Friedland, R.P. Cross-kingdom sequence similarities between human micro-RNAs and plant viruses. Commun. Integr. Biol. 2013, 6, e24951. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balique, F.; Lecoq, H.; Raoult, D.; Colson, P. Can Plant Viruses Cross the Kingdom Border and Be Pathogenic to Humans? Viruses 2015, 7, 2074-2098. https://doi.org/10.3390/v7042074
Balique F, Lecoq H, Raoult D, Colson P. Can Plant Viruses Cross the Kingdom Border and Be Pathogenic to Humans? Viruses. 2015; 7(4):2074-2098. https://doi.org/10.3390/v7042074
Chicago/Turabian StyleBalique, Fanny, Hervé Lecoq, Didier Raoult, and Philippe Colson. 2015. "Can Plant Viruses Cross the Kingdom Border and Be Pathogenic to Humans?" Viruses 7, no. 4: 2074-2098. https://doi.org/10.3390/v7042074
APA StyleBalique, F., Lecoq, H., Raoult, D., & Colson, P. (2015). Can Plant Viruses Cross the Kingdom Border and Be Pathogenic to Humans? Viruses, 7(4), 2074-2098. https://doi.org/10.3390/v7042074